
Method Article
إعادة تشكيل وتوصيف مركبات الأكتين-الأنابيب الدقيقة مع ديناميكيات وميكانيكا قابلة للضبط مدفوعة بمحرك
In This Article
Erratum Notice
Summary
يقدم هذا البحث بروتوكولات لهندسة وتوصيف الشبكات المركبة ثلاثية الأبعاد القابلة للضبط لخيوط الأكتين المتشابكة والأنابيب الدقيقة. تخضع المركبات لإعادة هيكلة نشطة وحركة باليستية ، مدفوعة بمحركات الميوسين الثاني والكينيسين ، ويتم ضبطها بواسطة التركيزات النسبية للأكتين والأنابيب الدقيقة والبروتينات الحركية والوصلات المتشابكة السلبية.
Abstract
الهيكل الخلوي المركب ، الذي يتألف من شبكات متفاعلة من خيوط الأكتين شبه المرنة والأنابيب الدقيقة الصلبة ، يعيد هيكلة ويولد قوى باستخدام البروتينات الحركية مثل الميوسين الثاني والكينيسين لدفع العمليات الرئيسية مثل الهجرة ، والتحريك الخلوي ، والالتصاق ، والاستشعار الميكانيكي. في حين أن تفاعلات الأكتين والأنابيب الدقيقة هي المفتاح لتعدد استخدامات الهيكل الخلوي وقدرته على التكيف ، فإن فهم تفاعلها مع نشاط الميوسين والكينيسين لا يزال ناشئا. يصف هذا العمل كيفية هندسة شبكات مركبة ثلاثية الأبعاد قابلة للضبط من خيوط الأكتين المتشابكة والأنابيب الدقيقة التي تخضع لإعادة هيكلة نشطة وحركة باليستية ، مدفوعة بمحركات الميوسين II والكينيسين ، ويتم ضبطها بواسطة التركيزات النسبية للأكتين والأنابيب الدقيقة والبروتينات الحركية والروابط المتشابكة السلبية. كما تم تفصيل بروتوكولات وضع العلامات الفلورية للأنابيب الدقيقة وخيوط الأكتين لتصور إعادة الهيكلة المركبة والحركة بشكل أكثر فعالية باستخدام التصوير متعدد الأطياف متحد البؤر. أخيرا ، يتم تقديم نتائج طرق تحليل البيانات التي يمكن استخدامها لتوصيف هيكل عدم التوازن وديناميكياته وميكانيكا عدم التوازن. توفر إعادة إنشاء منصة المحاكاة الحيوية القابلة للضبط والتحقيق فيها نظرة ثاقبة حول كيف يمكن أن يؤدي النشاط الحركي المقترن والميكانيكا المركبة وديناميكيات الخيوط إلى عمليات خلوية لا تعد ولا تحصى من الانقسام إلى الاستقطاب إلى الإحساس الميكانيكي.
Introduction
الهيكل الخلوي عبارة عن شبكة مركبة ديناميكية من البوليمرات الحيوية المتفاعلة التي توفر الدعم الهيكلي والميكانيكي للخلايا. تعمل المحركات الجزيئية المرتبطة والبروتينات الملزمة على إعادة هيكلة الهيكل الخلوي وتكييفه للسماح للخلايا بالنمو وتغيير الشكل والتصلب والتحرك وحتى الشفاء الذاتي ، مما يتيح عمليات خلوية لا تعد ولا تحصى تتراوح من الهجرة والانقسام إلى الاستشعار الميكانيكي 1,2. بالإضافة إلى أهميته في الفيزياء الحيوية الخلوية ، يعد الهيكل الخلوي أيضا مثالا جوهريا للمادة النشطة مع تطبيقات المواد المحتملة التي تتراوح من التئام الجروح وتوصيل الأدوية إلى الترشيح والروبوتات اللينة1،3،4،5،6،7،8،9.
السمتان الرئيسيتان اللتان تضفيان الهيكل الخلوي تنوعه الهيكلي والميكانيكي الفريد وتعدد وظائفه هما: 1) طبيعته المركبة ، التي تضم خيوط بروتينية متعددة متفاعلة ، مثل خيوط الأكتين شبه المرنة والأنابيب الدقيقة الصلبة ، بالإضافة إلى البروتينات المرتبطة بهاوالمتشابكة 3،5،10 ؛ و 2) قدرتها على إعادة الهيكلة المستمرة ، والتحرك ، والخشنة ، وأداء العمل عبر المحركات المستهلكة للطاقة ، مثل الميوسين والكينيسين ، ودفع وسحب البروتينات الخيطية1،7،11،12،13. في حين أن هذا التعقيد الأنيق يمكن الهيكل الخلوي من التوسط في عمليات متنوعة مثل حركة الخلية ، والتحريك الخلوي ، والتئام الجروح3،6،7،11 ، فإنه يعيق قدرة الباحثين على إعادة إنتاج خصائص التوقيع في الجسم الحي للهيكل الخلوي في الأنظمة المعاد تشكيلها في المختبر.
تركز جهود إعادة تكوين الحدود الحالية على مركبات خيوط الأكتين المتشابكة والمتشابكة والأنابيب الدقيقة3،10،14،15،16،17 ، وشبكات الأكتوميوسين المولدة للقوة2،8،18،19،20،21 ، والنيماتيكا النشطة التي تقودها الأنابيب الدقيقة الحركية التفاعلات22،23،24،25،26. وقد تبين أن مركبات الأكتين-الأنابيب الدقيقة ذات الحالة المستقرة تعرض الخواص الميكانيكية الناشئة15،16،27 ، مثل حركة الخيوط المحسنة وزيادة الصلابة مقارنة بالأنظمة أحادية المكون 27. أبلغت الدراسات التي أجريت على أنظمة الأكتوميوسين في المختبر عن مجموعة واسعة من الخصائص الهيكلية والديناميكية التي تعتمد على تركيزات الأكتين والميوسين والروابط المتشابكة28،29،30،31. على سبيل المثال ، مع وجود تشابك كاف ، تخضع شبكات الأكتوميوسين لانكماش واسع النطاق وخشن2،28،30،32،33،34،35،36 ، بينما بدون روابط متشابكة ، تظهر الشبكات تدفقا سريعا مزعزعا للاستقرار وتمزق 19,29 . تم الإبلاغ عن أن النيماتات النشطة القائمة على الأنابيب الدقيقة المعاد تشكيلها والتي تستخدم مجموعات من محركات kinesin للربط المتشابك وسحب حزم الأنابيب الدقيقة تظهر تدفقات مضطربة طويلة الأمد ، وتمديد ، وانبعاج ، وتكسير ، وشفاء12،22،23،24،25،37،38،39،40،41 ، 42،43،44،45،46،47.
في الآونة الأخيرة ، تبين أن مركبات الأكتين-الأنابيب الدقيقة المدفوعة بخيوط الميوسين II الصغيرة تؤدي إلى انكماش أكثر ترتيبا وسلامة الشبكة مقارنة بالتدفق المضطرب وتمزق الشبكة الذي تظهره شبكات الأكتوميوسين بدون وصلات متشابكة17،26،48. علاوة على ذلك ، يتم تحسين الجمع بين المتانة المركبة وتوليد القوة عندما يكون الأكتين والأنابيب الدقيقة موجودين بتركيزات مماثلة. تشمل السمات الناشئة الرئيسية في هذه المنطقة من مساحة الصياغة القوة الميكانيكية المحسنة 26 ، والحركة المنسقة للأكتين والأنابيب الدقيقة26 ، والانكماش المستمر المطرد ، وإعادة الهيكلة متوسطة الحجم17.
هنا ، يتم وصف البروتوكولات لهندسة وضبط المركبات المتشابكة والمتشابكة للأنابيب الدقيقة وخيوط الأكتين التي يتم دفعها خارج التوازن بواسطة خيوط الميوسين II الصغيرة ومجموعات الكينيسين التي تعمل على خيوط الأكتين والأنابيب الدقيقة ، على التوالي (الشكل 1). يمكن ضبط ديناميكيات وهيكل وميكانيكا هذه الفئة من المركبات من خلال التركيزات النسبية للخيوط والمحركات والوصلات المتشابكة لإظهار مساحة طور غنية من التدفق المضطرب والمضطرب ، والانكماش الخواص ، والتسارع ، والتباطؤ ، وإزالة الخلط ، والتصلب ، والاسترخاء ، والتمزق. ينصب تركيز هذا العمل على إعداد وضبط هذه الفئة من المركبات الهيكلية الخلوية النشطة. ومع ذلك ، لمساعدة الباحثين في قياس وتوصيف المركبات النشطة الموصوفة ، يتم أيضا تفصيل طرق التصوير الفعالة باستخدام الفحص المجهري متعدد الأطياف متحد البؤر. أخيرا ، يتم تقديم نتائج طرق التحليل الحسابي الرئيسية التي يمكن استخدامها لقياس ديناميكيات وهيكل وميكانيكا المركبات. يتم تشجيع الباحثين على اعتماد هذه الأساليب - والتي تشمل الفحص المجهري الديناميكي التفاضلي (DDM) ، والارتباط الذاتي للصور المكانية (SIA) ، وقياس سرعة صورة الجسيمات (PIV) - حيث تم تحسينها لتوصيف الديناميكيات المعقدة والتنوع الهيكلي للمركبات17،26،49.
تركز الخطوات الموضحة أدناه على تحضير المركبات وتصويرها باستخدام الفحص المجهري متحد البؤر. يمكن العثور على البروتوكولات التي تصف تحليل بيانات ما بعد الاستحواذ وقياسات الملقط البصري في الأعمال السابقة 17،26،48،50 ، ويتم توفيرها عند الطلب. يتم سرد جميع المواد في جدول المواد المقدمة.
Protocol
1. تحضير أغطية silanized وشرائح مجهرية لمنع امتزاز البروتينات على أسطح الحجرة
ملاحظة: هذه عملية لمدة 2 يوم. يمكن إعداد الشرائح Silanized تصل إلى 1 شهر قبل الاستخدام.
- ضع أغطية الأغطية رقم 1 (24 مم × 24 مم) وشرائح المجهر (1 بوصة × 3 بوصات) في رف مخصص يتناسب مع منظف البلازما. ضع الرف في منظف البلازما وقم بتشغيله لمدة 20 دقيقة.
- انقل أغطية الأغطية والشرائح إلى رف جديد مخصص فقط للاستخدام مع السيلان وضع الرف في وعاء زجاجي لتنظيف النظارات كما هو موضح أدناه.
- اغمر أغطية الأغطية والشرائح في الأسيتون بنسبة 100٪ لمدة 1 ساعة. اغمر الأغطية والشرائح في الإيثانول بنسبة 100٪ لمدة 10 دقائق.
- اغمر الأغطية والشرائح في الماء منزوع الأيونات (DI) لمدة 5 دقائق. كرر خطوات التنظيف مرتين أخريين.
- اغمر أغطية الأغطية والشرائح في 0.1 مليون KOH معدة حديثا لمدة 15 دقيقة. اغمر الأغطية والشرائح في DI جديد لمدة 5 دقائق. كرر هذه الخطوة مرتين أخريين.
- أغطية وشرائح جافة بالهواء لمدة 10 دقائق. عالج الأغطية والشرائح النظيفة باستخدام السيلان لإنتاج أسطح كارهة للماء كما هو موضح أدناه.
ملاحظة: أكمل الخطوات التالية في غطاء الدخان.- اغمر الأغطية المجففة والشرائح في 2٪ سيلان (مذاب في التولوين) لمدة 5 دقائق. استخدم قمعا لصب السيلان مرة أخرى في الزجاجة المخصصة لإعادة استخدامه حتى خمس مرات.
- اغمر الأغطية والشرائح في الإيثانول بنسبة 100٪ لمدة 5 دقائق. استبدل الإيثانول بالإيثانول الطازج. اغمر أغطية الأغطية والشرائح لمدة 5 دقائق.
- اغمر الأغطية والشرائح في DI جديد لمدة 5 دقائق. كرر خطوة غسل الإيثانول و DI مرتين أخريين باستخدام الإيثانول الطازج و DI في كل مرة. أغطية وشرائح جافة بالهواء لمدة 10 دقائق.
2. تحضير مركب أكتين - أنيبيب دقيق نشط مدفوع بخيوط الميوسين الصغيرة
- إزالة الميوسين غير النشط عن طريق ارتباط خيوط الأكتين وإجراء السحب لأسفل عن طريق الطرد المركزي الفائق كما هو موضح أدناه.
- بلمرة الأكتين إلى خيوط. باستخدام ماصة دقيقة وأطراف ماصة معقمة ، ادمج في أنبوب طرد مركزي دقيق: 1.87 ميكرولتر من DI ، 1.3 ميكرولتر من 10x G-buffer ، 1.3 ميكرولتر من 10x F-buffer ، 1.63 ميكرولتر من 4 M KCl ، 4.53 ميكرولتر من الأكتين (47.6 ميكرومتر) ، و 1.08 ميكرولتر من 100 ميكرومتر من القضيب القضيبي.
ملاحظة: لضمان البلمرة الكافية ، يجب أن يكون تركيز الأكتين ونسبة أكتين: فالويدين المولية 18.4 ميكرومتر و 2: 1 ، على التوالي. - اسكب المحلول برفق لأعلى ولأسفل حتى يختلط ثم ضعه على الثلج في الظلام لمدة ≥1 ساعة. تبريد جهاز الطرد المركزي الفائق إلى 4 درجات مئوية. إزالة حصة الميوسين من -80 درجة مئوية ووضعها على الثلج.
ملاحظة: أكمل الخطوة 2.2 في هذه المرحلة أثناء بلمرة الأكتين. - بعد ≥1 ساعة من بلمرة الأكتين ، أضف 1.3 ميكرولتر من 10 mM ATP و 2 μL من 19 μM من الميوسين إلى الأكتين المبلمر.
ملاحظة: يجب أن تكون نسبة الأكتين: الميوسين المولية >5 لضمان الإزالة الكافية لمحركات الميوسين غير النشطة (أي الرؤوس الميتة). - اسكب المحلول برفق لأعلى ولأسفل للخلط. نقل إلى أنبوب الصف الطرد المركزي الفائق.
جهاز طرد مركزي عند 4 درجات مئوية و 121,968 × جم لمدة 30 دقيقة.
- بلمرة الأكتين إلى خيوط. باستخدام ماصة دقيقة وأطراف ماصة معقمة ، ادمج في أنبوب طرد مركزي دقيق: 1.87 ميكرولتر من DI ، 1.3 ميكرولتر من 10x G-buffer ، 1.3 ميكرولتر من 10x F-buffer ، 1.63 ميكرولتر من 4 M KCl ، 4.53 ميكرولتر من الأكتين (47.6 ميكرومتر) ، و 1.08 ميكرولتر من 100 ميكرومتر من القضيب القضيبي.
- تحضير شبكة مركبة متشابكة من خيوط الأكتين والأنابيب الدقيقة كما هو موضح أدناه.
ملاحظة: ابدأ قبل 30 دقيقة من دوران الميوسين لأسفل (الخطوة 2.1.4).- اضبط كتلة الحرارة على 37 درجة مئوية. استخدم ماصة دقيقة وأطراف ماصة معقمة لإضافة ما يلي إلى أنبوب الطرد المركزي الدقيق: 13.9 ميكرولتر من PEM ، 3 ميكرولتر من 1٪ Tween20 ، 1.55 ميكرولتر من 47.6 ميكرومتر أكتين ، 0.36 ميكرولتر من 34.8 ميكرومتر R-actin ، 0.3 ميكرولتر من 250 مللي متر ATP ، 0.87 ميكرولتر من 100 ميكرومتر من القضيب ، 1.91 ميكرولتر من 5-488-توبولين ، 0.3 ميكرولتر من 100 مللي متر GTP ، و 0.75 ميكرولتر من 200 ميكرومتر تاكسول ، إلى حجم إجمالي قدره 23 ميكرولتر.
ملاحظة: تركيزات الأكتين والتوبولين المدرجة هي لمركب يحتوي على 2.9 ميكرومتر من الأكتين و 2.9 ميكرومتر من التوبولين. إجمالي تركيز البروتين هو c = c A + c T = 5.8 μM وجزء الأكتين المولي هو c A / (c A + cT) = Φ A = 0.5. راجع الخطوة 2.5 لضبط هذه القيم. - اسكب المحلول برفق لأعلى ولأسفل لخلطه وضعه على كتلة حرارية 37 درجة مئوية محمية من الضوء لمدة 1 ساعة.
- اضبط كتلة الحرارة على 37 درجة مئوية. استخدم ماصة دقيقة وأطراف ماصة معقمة لإضافة ما يلي إلى أنبوب الطرد المركزي الدقيق: 13.9 ميكرولتر من PEM ، 3 ميكرولتر من 1٪ Tween20 ، 1.55 ميكرولتر من 47.6 ميكرومتر أكتين ، 0.36 ميكرولتر من 34.8 ميكرومتر R-actin ، 0.3 ميكرولتر من 250 مللي متر ATP ، 0.87 ميكرولتر من 100 ميكرومتر من القضيب ، 1.91 ميكرولتر من 5-488-توبولين ، 0.3 ميكرولتر من 100 مللي متر GTP ، و 0.75 ميكرولتر من 200 ميكرومتر تاكسول ، إلى حجم إجمالي قدره 23 ميكرولتر.
- تحضير غرف العينات لتجارب التصوير متحد البؤر كما هو موضح أدناه.
ملاحظة: أكمل الخطوتين 2.1.4 و 2.2.2 أثناء فترات الانتظار.- ضع شريحتين مسيلانيتين جنبا إلى جنب على صفيحة ساخنة (مطفأة) ، وضع شريطين من فيلم الختم بالحرارة عبر الشرائح ~ 3 مم ، وضع غطاءين سيلانزين فوق فيلم الختم بالحرارة لتشكيل غرفة عينة.
- قم بتشغيل لوح التسخين على إعداد منخفض حتى تلتصق الأغطية بإحكام بالشرائح باستخدام فيلم مانع للتسرب بالحرارة ذائب (~ 1-2 دقيقة). اضغط لأسفل بضغط متساو لضمان الترابط مع الحفاظ على تباعد ~ 100 ميكرومتر بين السطحين.
- قم بإزالة الغرف وإيقاف تشغيل اللوحة الساخنة. غرف التسمية مع (+) و (-). ستكون الغرفة (+) للعينة النشطة (مع الميوسين) وستكون الغرفة (-) هي السيطرة (بدون الميوسين). تأكد من أن كل غرفة يمكن أن تستوعب ≤10 ميكرولتر من السوائل.
- قم بإعداد عينات للصورة كما هو موضح أدناه.
ملاحظة: من المهم إكمال هذه الخطوة مباشرة بعد إكمال الخطوتين 2.1 و 2.2.- قم بإزالة عينة الميوسين-أكتين بعناية من جهاز الطرد المركزي الفائق (الخطوة 2.1.4) وقم على الفور بسحب أعلى 7.5 ميكرولتر من المادة الطافية ونقلها إلى أنبوب طرد مركزي دقيق جديد.
- قم بإزالة عينة الأكتين والأنابيب الدقيقة من كتلة الحرارة واخلطها برفق في 1.5 ميكرولتر من 10x D-Glucose ، و 1.5 ميكرولتر من 10x GOC ، و 1.5 ميكرولتر من 1 mM blebbistatin. قسم المحلول إلى قسمين 13.7 ميكرولتر وقم بتسميته ك (+) و (-).
- امزج 1.28 ميكرولتر من المادة الطافية من الخطوة 2.4.1 إلى (+) القسمة امزج 1.28 ميكرولتر من DI إلى القسمة (-). تدفق ببطء كل محلول إلى الغرفة المقابلة (الخطوة 2.3) عن طريق العمل الشعري. احرص على عدم إدخال فقاعات الهواء في القناة.
- أغلق الطرفين المفتوحين لكل قناة بإيبوكسي سريع الجفاف أو غراء الأشعة فوق البنفسجية. تأكد من جفاف المادة اللاصقة تماما قبل وضعها على المجهر. الصورة فورا كما هو موضح في الخطوة 3.
ملاحظة: الغراء فوق البنفسجي مفيد لأنه يعالج على الفور تقريبا عند التعرض للأشعة فوق البنفسجية. ومع ذلك ، نظرا لأن البليبيستاتين حساس للأشعة فوق البنفسجية ، فمن المهم فقط إضاءة الغراء محليا (عند حواف غرفة العينة) باستخدام عصا صغيرة للأشعة فوق البنفسجية لتجنب إلغاء تنشيط البليبيستاتين.
- اختياري: قم بتغيير تركيزات البروتين لضبط ديناميكيات وهيكل المركبات.
ملاحظة: الخطوات التالية هي تعديلات مقترحة على الخطوات المذكورة أعلاه لتغيير تركيزات الأكتين والأنابيب الدقيقة والميوسين إذا رغبت في ذلك.- اتبع الخطوات الموضحة أعلاه باستثناء التعديلات التالية في الخطوتين 2.2.1 و 2.4.3.
- لتغيير تركيزات الأكتين والأنابيب الدقيقة ، وبالتالي ضبط c و ΦA ، قم بزيادة أو تقليل حجم الأكتين و R-actin و 5-488-tubulin المستخدم في الخطوة 2.2.1 ، حسب الرغبة26. عند تغيير تركيز الأكتين ، اضبط تركيزات R-actin و phalloidin المولية بشكل متناسب للحفاظ على نفس النسب المولية مع الأكتين. اضبط حجم PEM بحيث يظل الحجم النهائي للخليط 23 ميكرولتر. تظل جميع أحجام وتركيزات المكونات الأخرى كما هي.
- لتغيير تركيز الميوسين، اضبط حجم الميوسين المضاف إلى القسمة (+) في الخطوة 2.4.3 حسب الرغبة. اضبط حجم DI المضاف إلى القسمة (-) وفقا لذلك. اضبط حجم PEM في الخطوة 2.2.1 لحساب الزيادة أو النقصان في حجم الميوسين (+) و DI (-) ، مما يضمن أن الحجم النهائي لكل عينة ((+) و (-)) هو 14.98 ميكرولتر.
3. تصوير وتوصيف المركبات النشطة باستخدام المجهر متحد البؤر
- لتصوير مركبات الأكتوميوسين والأنابيب الدقيقة المحضرة في الخطوة 2 ، استخدم مجهر المسح الضوئي متحد البؤر (LSCM) ، أو مجهر مشابه ، بهدف غمر الزيت 60x 1.4 NA. لتصور خيوط الأكتين والأنابيب الدقيقة في وقت واحد في قنوات مضان منفصلة ، استخدم ليزر 561 نانومتر مع مرشحات إثارة / انبعاث 565/591 نانومتر وليزر 488 نانومتر مع مرشحات إثارة / انبعاث 488/525 نانومتر.
- ضع حجرة العينة على المجهر بحيث يتم وضع قناة التحكم مباشرة فوق الهدف. تأكد من وجود واجهة زيت بين الهدف وغطاء الغطاء.
- استخدم عناصر التحكم في المرحلة لجلب مركب التحكم إلى التركيز البؤري ، ثم ابحث عن سطحي حجرة العينة. انقل الموضع z إلى مركز حجرة العينة. تحقق من وجود شبكات خيطية واضحة كما هو موضح في الشكل 2.
- مع الاستمرار في تصور غرفة التحكم ، اضبط شدة كل ليزر للسماح بالتصور المتزامن لخيوط الأكتين والأنابيب الدقيقة. حافظ على أقل كثافة ليزر ممكنة لمنع التبييض الضوئي (أكثر انتشارا في قناة الأكتين) والنزيف (عادة من الأنابيب الدقيقة إلى قناة الأكتين).
- لتوصيف عينة التحكم غير النشطة ، اجمع ثلاث سلاسل زمنية (مقاطع فيديو) من صور 256 × 256 بكسل مربع (213 ميكرومتر × 213 ميكرومتر) بمعدل 2.65 إطارا في الثانية بإجمالي ≥1000 إطار. اجمع كل سلسلة زمنية في منطقة مختلفة من غرفة العينة مفصولة ب ≥500 ميكرومتر. تأكد من وجود الحد الأدنى من الحركة التي يمكن اكتشافها وعدم وجود تدفق أو إعادة هيكلة.
- قم بإيقاف تشغيل الليزر 488 نانومتر واستخدم عناصر التحكم في المسرح للانتقال إلى غرفة (+).
- باستخدام ليزر 568 نانومتر ، تصور الأنابيب الدقيقة في القناة (+) لضمان تكوين الشبكة بشكل صحيح (الشكل 2) وتحديد المركز المحوري لغرفة العينة (والذي قد يكون مختلفا عن موضع مركز z لغرفة التحكم).
- قم بتشغيل الليزر 488 نانومتر وكرر الخطوة 3.5 أعلاه مع التعديلات التالية. اجمع السلاسل الزمنية لمدة تصل إلى 45 دقيقة ، وتوقف عن الاكتساب عندما تتحرك العينة خارج مجال الرؤية أو التمزق أو المبيضات الضوئية. سجل 5-10 سلاسل زمنية وتتبع الوقت الذي تبدأ فيه كل سلسلة زمنية بالنسبة إلى بداية السلسلة الزمنية الأولى.
- تحليل البيانات باستخدام DDM و SIA و PIV كما هو موضح في الشكل 3 والشكل 4 والشكل 5 وسابقا17،48،50،51.
ملاحظة: ينشط ليزر 488 نانومتر نشاط الميوسين ATPase محليا عن طريق إلغاء تنشيط blebbistatin ، لذلك يجب تشغيله فقط في بداية الحصول على البيانات بحيث يكون t = 0 في بداية السلسلة الزمنية. تم تحسين معلمات الاستحواذ هذه لتحليل المجهر الديناميكي التفاضلي (DDM) كما تم سابقا26.
4. تحضير مركبات الأكتين والأنابيب الدقيقة النشطة التي تحركها محركات كينيسين
ملاحظة: تقوم الخطوات التالية بإنشاء مركبات الأكتين-الأنابيب الدقيقة التي يتم إخراجها من التوازن بواسطة محركات kinesin أو مزيج من kinesin و myosin50.
- تحضير محركات كينيسين وميوسين كما هو موضح أدناه.
- في حالة دمج الميوسين ، اتبع الخطوة 2.1.
- لتشكيل مجموعات حركية كينيسين تربط وتمارس قوى بين أزواج من الأنابيب الدقيقة ، استخدم ماصة دقيقة وأطراف ماصة معقمة لإضافة ما يلي إلى أنبوب طرد مركزي دقيق معقم سعة 1.5 مل: 1.16 ميكرولتر PEM ، 2.74 ميكرولتر 8.87 ميكرومتر كينيسين دايمرات ، 7.29 ميكرولتر 83.3 ميكرومتر نيوترافيدين ، 0.81 ميكرولتر 2 مللي متر DTT . امزج بلطف عن طريق سحب المحلول لأعلى ولأسفل واحتضانه محميا من الضوء (استخدم أنبوب طرد مركزي أسود أو لف بورق الألمنيوم) لمدة 30 دقيقة عند 4 درجات مئوية.
ملاحظة: النسبة المولية لثنائيات الكينيسين إلى NA هي 1:25.
- اتبع الخطوة 2.3 لتحضير غرف العينة وعمل ثلاث غرف بدلا من غرفتين. نفذ هذه الخطوة أثناء حضانة كينيسين (الخطوة 4.1.2) والطرد المركزي الفائق للميوسين (الخطوة 4.1.1).
- تحضير شبكة مركبة متشابكة من خيوط الأكتين والأنابيب الدقيقة.
- اضبط كتلة الحرارة على 37 درجة مئوية. استخدم ماصة دقيقة وأطراف ماصة معقمة لإضافة ما يلي إلى أنبوب طرد مركزي دقيق معقم سعة 1.5 مل: 3.21 ميكرولتر PEM ، 4.5 ميكرولتر من 1٪ Tween20 ، 2.18 ميكرولتر من 47.6 ميكرومتر أكتين ، 3.46 ميكرولتر من 5-R-tubulin ، 4.5 ميكرولتر من 100 mM ATP ، 4.5 ميكرولتر من 10 mM GTP ، 1.13 ميكرولتر من 200 ميكرومتر تاكسول ، و 1.57 ميكرولتر من 20 ميكرومتر 488-phalloidin. تأكد من أن الحجم الإجمالي هو 25 ميكرولتر.
- اسكب المحلول برفق لأعلى ولأسفل لخلطه وضعه على كتلة حرارية 37 درجة مئوية محمية من الضوء لمدة 1 ساعة. قم بإزالة الأنبوب من كتلة الحرارة واستخدم ماصة دقيقة لخلط 0.84 ميكرولتر من 100 ميكرومتر من القضيب برفق. احتضان لمدة 5-10 دقائق في درجة حرارة الغرفة ، محمية من الضوء.
ملاحظة: تؤدي إضافة phalloidin في هذه الخطوة ، بدلا من الخطوة 4.3.1 ، إلى تحسين وضع العلامات الفلورية لخيوط الأكتين ، حيث لا يتعين على 488-phalloidin التنافس مع phalloidin غير المسمى لمواقع ربط الأكتين.
- إعداد المركبات النشطة للتصوير متحد البؤر.
- أضف 1.13 ميكرولتر من 200 ميكرومتر بليبيستاتين ، و 1.35 ميكرولتر من 10x Glu ، و 1.35 ميكرولتر من 10x GOC إلى المحلول من الخطوة 4.3.2 واخلط بلطف عن طريق السحب لأعلى ولأسفل. قسم المحلول إلى ثلاثة مقسومات 10 ميكرولتر وقم بتسميتها ك (K) و (K + M) و (-).
- امزج 2.54 ميكرولتر من الميوسين من الخطوة 2.1.4 إلى القسمة (K + M). امزج في 2.54 ميكرولتر PEM إلى (K) و (-) القسمة الثنائية.
- استخدم ماصة صغيرة وأطراف ماصة معقمة لإضافة 2.5 ميكرولتر من مجموعات كينيسين من الخطوة 4.1.2 إلى (K) و (K + M) القسمة . ماصة صعودا وهبوطا لخلط. امزج 2.5 ميكرولتر PEM إلى (-) باستخدام نفس التقنية.
ملاحظة: تركيزات الأكتين والتوبولين المدرجة هي لمركب يحتوي على 2.32 ميكرومتر أكتين و 3.48 ميكرومتر توبولين. إجمالي تركيز البروتين هو c = c A + c T = 5.8 μM وجزء الأكتين المولي هو c A / (c A + cT) = Φ A = 0.4. تركيزات كينسين والميوسين هي 0.35 ميكرومتر و 0.47 ميكرومتر على التوالي. راجع الخطوة 2.5 للحصول على إرشادات عامة لضبط c A و cT و c و ΦA. - باستخدام micropipette ، قم بتدفق كل محلول ببطء في القناة المقابلة لغرف العينة المعدة (الخطوة 4.2) عبر العمل الشعري. ادفع لأسفل ببطء شديد وبرفق على الماصة حتى لا تدخل فقاعات الهواء في القناة.
- أغلق الطرفين المفتوحين لكل قناة بإيبوكسي سريع الجفاف أو غراء قابل للمعالجة بالأشعة فوق البنفسجية. تأكد من جفاف المادة اللاصقة تماما قبل وضعها على المجهر.
ملاحظة: من المهم أن تتم هذه الخطوة بسرعة لتقليل الوقت الذي يعمل فيه الكينيسين دون مراقبة. لهذا السبب ، يوصى باستخدام الإيبوكسي الذي يعالج في دقيقة واحدة (بدلا من 5 أو 10 دقائق). يعتبر الغراء القابل للمعالجة بالأشعة فوق البنفسجية مفيدا في هذا الصدد لأنه يعالج على الفور تقريبا عند التعرض للأشعة فوق البنفسجية.
- أعدت الصورة العينات على الفور ، باتباع الخطوة 3 ، باستثناء التعديلات المهمة التالية. نظرا لأن kinesin لا يتم التحكم فيه عن طريق تنشيط الضوء ، فإنه يبدأ في العمل مباشرة بعد الخطوة 4.4.3 ، لذا ضع علامة على هذه المرة ك t = 0. لتصوير المركب بالقرب من الحالة الأولية غير النشطة (t = 0) قدر الإمكان ، قم بتصوير القناتين (K) و (K + M) أولا ولاحظ الوقت المنقضي بين الخطوة 4.4.3 وبداية الحصول على البيانات (الخطوة 3.8). في الممارسة العملية ، هذا الوقت المنقضي هو ~ 5 دقائق.
5. دمج الروابط المتشابكة السلبية في المركبات النشطة
ملاحظة: تصف هذه الخطوات كيفية استخدام وحدات فرعية من الأكتين والتوبولين الحيوي و NeutrAvidin (NA) لربط الأكتين بشكل سلبي بالأكتين (A-A) أو الأنابيب الدقيقة بالأنابيب الدقيقة (M-M) في المركبات النشطة الموضحة في الخطوة 4.
- تحضير مجمعات A-A أو M-M المتشابكة مع البروتينات البيوتينية (البيوتين-أكتين أو البيوتين-توبولين) ، NA ، والبيوتين بنسبة 2: 2: 1 البيوتين-أكتين / توبولين: البيوتين: NA. ابدأ هذه العملية قبل الخطوة 4 .
- بالنسبة للروابط المتشابكة A-A ، استخدم ماصة دقيقة وأطراف ماصة معقمة لإضافة 2 ميكرولتر من 11.6 ميكرومتر من البيوتين أكتين ، و 1.39 ميكرولتر من 8.33 ميكرومتر NA ، و 2.27 ميكرولتر من 1.02 ميكرومتر من البيوتين ، و 4.34 ميكرولتر من PEM إلى أنبوب الطرد المركزي الدقيق. تخلط بلطف عن طريق سحب لأعلى ولأسفل.
- بالنسبة للروابط المتشابكة M-M ، استخدم ماصة دقيقة وأطراف ماصة معقمة لإضافة 1.86 ميكرولتر من 4.55 ميكرومتر من البيوتين توبولين ، و 1.11 ميكرولتر من 8.33 ميكرومتر NA ، و 1.82 ميكرولتر من 1.02 ميكرومتر من البيوتين ، و 5.21 ميكرولتر من PEM إلى أنبوب طرد مركزي دقيق. تخلط بلطف عن طريق سحب لأعلى ولأسفل.
- لف الأنبوب (الأنابيب) من الخطوة 5.1.1 و / أو 5.1.2 في فيلم مانع للتسرب بالحرارة لإنشاء ختم محكم الغلق. ضعه في طوف تعويم في حمام صوتي يتم التحكم في درجة حرارته مضبوط على 4 درجات مئوية.
- سونيكات لمدة 90 دقيقة عند 4 درجات مئوية. في الممارسة العملية ، من الأفضل وضع الصوتنة في غرفة باردة وإضافة عبوات ثلج إلى حمام صوتنة للحفاظ على درجة حرارة منخفضة.
- لدمج مجمعات الروابط المتشابكة في عينات للتصوير ، اتبع الخطوة 4.3 ، وتعديل الخطوة 4.3.1 كما هو موضح أدناه للربط المتشابك A-A (الخطوة 5.2.1) أو الربط المتشابك M-M (الخطوة 5.2.2).
- بالنسبة للربط المتشابك A-A ، ادمج ما يلي في أنبوب طرد مركزي دقيق: 1.94 ميكرولتر من PEM ، 4.50 ميكرولتر من 1٪ Tween20 ، 2.18 ميكرولتر من 47.6 ميكرومتر أكتين ، 3.46 ميكرولتر من 45.5 ميكرومتر 5-R-tubulin ، 1.13 ميكرولتر من الوصلات المتشابكة A-A (الخطوة 5.1.1) ، 4.50 ميكرولتر من 100 مللي متر ATP ، 4.50 ميكرولتر من 10 مللي متر GTP ، 1.13 ميكرولتر من 200 ميكرومتر تاكسول ، و 1.57 ميكرولتر من 20 ميكرومتر 488-phalloidin. تأكد من أن الحجم الإجمالي هو 25 ميكرولتر.
- بالنسبة للربط المتشابك M-M ، ادمج ما يلي في أنبوب طرد مركزي دقيق: 1.97 ميكرولتر من PEM ، 4.50 ميكرولتر من 1٪ Tween20 ، 2.18 ميكرولتر من 47.6 ميكرومتر أكتين ، 3.76 ميكرولتر من 45.5 ميكرومتر 5-R-tubulin ، 1.13 ميكرولتر من تخفيف 1: 4 من الروابط المتشابكة M-M (الخطوة 5.1.2) ، 4.50 ميكرولتر من 100 mM ATP ، 4.50 ميكرولتر من 10 mM GTP ، 1.13 ميكرولتر من 200 ميكرومتر تاكسول ، و 1.57 ميكرولتر من 20 ميكرومتر 488-phalloidin. تأكد من أن الحجم الإجمالي هو 25 ميكرولتر.
- اتبع الخطوات 4.3.2-4.5 مع التركيزات المحددة ل crosslinker: نسبة المولي الأكتين من RA = 0.02 و crosslinker: نسبة المولية tubulin من RT = 0.005. ينتج عن قيم R A و R T هذه أطوال متشابهة بين الروابط المتشابكة على طول خيوط الأكتين والأنابيب الدقيقة (d A 60 nm و d MT 67 nm) ، مقدرة باستخدام d A = I monomer / 2R A
 ، حيث Imonomer هو طول مونومر الأكتين ، و dMT = Iring / 26R T ، حيثالحلقة I هي طول حلقة مكونة من 13 أنبوبا15 ، 17.
، حيث Imonomer هو طول مونومر الأكتين ، و dMT = Iring / 26R T ، حيثالحلقة I هي طول حلقة مكونة من 13 أنبوبا15 ، 17.
النتائج
لتحديد التحضير الناجح للمركبات النشطة (الشكل 1) ، وتوصيف ديناميكياتها وهيكلها ، يتم استخدام مجهر مضان للمسح بالليزر مع قناتين مضان على الأقل لتصور خيوط الأكتين والأنابيب الدقيقة في وقت واحد (الشكل 2 والشكل 6). يتم تصنيف جميع خيوط الأكتين والأنابيب الدقيقة في المركبات بشكل ضئيل ، بدلا من تعاطي المنشطات في خيوط التتبع الساطعة ، كما يحدث غالبا في الدراسات المختبرية. تضمن هذه الطريقة أن تكون الديناميكيات والبنية المقاسة ممثلة للمركب نفسه بدلا من المتتبعات التي تتشكل في ظل ظروف مختلفة عن المركبات. لهذا السبب ، لا يمكن عادة حل خيوط الأكتين الفردية والأنابيب الدقيقة ، بل تصور الصور بنية شبكة متوسطة الحجم (الشكل 2 والشكل 6).
تم تحسين نهج وضع العلامات هذا لتحليلات الارتباط التلقائي للصور المكانية (SIA) والمجهر الديناميكي التفاضلي (DDM) التي تفحص الديناميات والبنية في فضاء فورييه المتبادل (الشكل 4 ، الشكل 5 ، والشكل 8) 52،53،54،55. يمكن أيضا استخدام قياس سرعة صورة الجسيمات (PIV) لتصوير وتوصيف الديناميات ومجالات التدفق (الشكل 3 والشكل 7) ، ولكنه يتطلب تجميع وحدات البكسل (دقة مكانية أقل) وزيادات أكبر في وقت التأخير (دقة زمنية أقل) من SIA و DDM للقضاء على المتجهات الخاطئة التي تنشأ من الضوضاء في الصور الكثيفة منخفضة الإشارة. ومع ذلك ، يوصى باستخدام PIV للفحص النوعي لمجالات التدفق وتأكيد نتائج DDM (الشكل 4 والشكل 8)26,50.
يتم توفير توصيف العينات للشبكات الموصوفة باستخدام هذه التحليلات (أي DDM و SIA و PIV) لمساعدة الباحثين في اعتماد تحليلات مماثلة لقياس وتوصيف عيناتهم. ومع ذلك ، فإن الأوصاف التفصيلية لهذه التقنيات تقع خارج نطاق هذا العمل. للحصول على أوصاف مفصلة لكيفية إجراء DDM على هذه الأنظمة وغيرها من الأنظمة المماثلة ، بما في ذلك كود Python سهل الاستخدام ، راجع الأعمال السابقة17،26،49،50 والمراجع الموجودة بداخلها. للحصول على تفاصيل حول كيفية إجراء SIA و PIV على الأنظمة الموضحة هنا ، يتم توجيه القارئ إلى الأعمال السابقة17,50.
يجب إجراء العديد من الضوابط الموضحة أدناه للتأكد من أن المركبات تعمل كما هو متوقع. يجب أن يبدو المركب الذي لا يحتوي على الميوسين أو الكينيسين ثابتا بشكل أساسي مع الحد الأدنى من التقلبات الحرارية أو الانجراف. يجب أن تظهر خيوط الأكتين والأنابيب الدقيقة متشابكة وموزعة بشكل متجانس ، مع الحد الأدنى من التجميع أو التجميع أو الفصل الطوري للأكتين والأنابيب الدقيقة في جميع أنحاء مجال رؤية ~ 200 ميكرومتر × 200 ميكرومتر (الشكل 2 ، أقصى اليسار)17. يجب على المرء أن يتوقع نتيجة مماثلة للمركبات التي تحتوي على الميوسين ولكنها لا تتعرض لضوء 488 نانومتر (لإلغاء تنشيط البليبيستاتين).
عند دمج الميوسين والتعرض لضوء 488 نانومتر ، تخضع المركبات لانكماش يكون إلى حد كبير متناحما ومشابها للأكتين والأنابيب الدقيقة ، كما هو موضح في صور المجهر الملتقطة قبل وبعد نشاط الميوسين (الشكل 2) ، وكذلك حقول تدفق PIV المقابلة لأوقات متفاوتة أثناء النشاط (الشكل 3). لتحديد ما إذا كانت الحركة باليستية ، منتشرة ، شبه منتشرة ، وما إلى ذلك ، يتم تقييم وقت الارتباط المميز τ (q) المحدد من DDM كدالة لمتجه الموجة (أي الفضاء المتبادل). انظر كما هو موضح بالتفصيل سابقا17،26،49. يوضح الشكل 4 أيضا كيفية استخدام DDM لتوصيف هذه المركبات. يشير تحجيم قانون الطاقة τ (q) ~ 1 / vq β ، مع β = 1 ، إلى الحركة الباليستية مع السرعة v. كمرجع ، يمثل β = 2 ديناميكيات الانتشار مع كون v معامل الانتشار. تظهر جميع المركبات النشطة تحجيم باليستي (الشكل 4 أ) بسرعات يتم ضبطها بواسطة تركيزات الأكتين والميوسين (الشكل 4 ب) ، والتي يمكن أن تختلف في الوقت المناسب أثناء النشاط ، إما بالتسارع أو التباطؤ (الشكل 4 ج ، د).
يمكن وصف إعادة هيكلة الشبكة وتجميعها ، المرئي في الشكل 2 والأكثر وضوحا لتركيزات الأكتين والميوسين الأعلى ، باستخدام SIA ، كما هو موضح في الشكل 5 ، والموصوف سابقا17،48،50. باختصار ، يمكن تحديد طول الارتباط ξ ، وهو مقياس للحجم المميز للمعالم في الصورة ، عن طريق ملاءمة كل منحنى ارتباط تلقائي للكثافة المكانية g (r) بدالة أسية للمسافة r بين وحدات البكسل. تشير قمم g (r) الأكبر التي تستمر لمسافات أطول إلى سمات هيكلية أكبر (أي التجميع ، وتجميع الخيوط الفردية). كما هو موضح في الشكل 5 ، بالنسبة لأجزاء الأكتين الأعلى وتركيزات الميوسين ، تنعكس إعادة الهيكلة والتجميع بشكل كبير في الزيادة في ξ بمرور الوقت.
يمكن أيضا قياس الخصائص اللزجة المرنة والاستجابة الميكانيكية غير الخطية للمركبات النشطة باستخدام علم الملقط البصري الدقيق (OTM). ومع ذلك ، فإن البروتوكولات والنتائج التمثيلية لهذه التجارب تقع خارج نطاق هذا العمل. تتم إحالة القراء المهتمين إلى الأعمال السابقة48,56 التي تصف بدقة كيفية إجراء قياسات OTM والنتائج المتوقعة.
باستخدام نفس برنامج الأدوات التجريبية والتحليلية الموضحة أعلاه ، يصف القسم التالي كيف تتغير الديناميكيات والهيكل عندما يتم دمج محركات kinesin وروابط البيوتين-NA المتشابكة في المركبات (الشكل 6 ، الشكل 7 ، والشكل 8). يوضح الشكل 6 صورا تمثيلية متحدة البؤر للمركبات التي تحركها إما الكينسين فقط (K) أو الكينيسين والميوسين (K + M) ، مع وبدون تشابك سلبي (XL) لخيوط الأكتين أو الأنابيب الدقيقة.
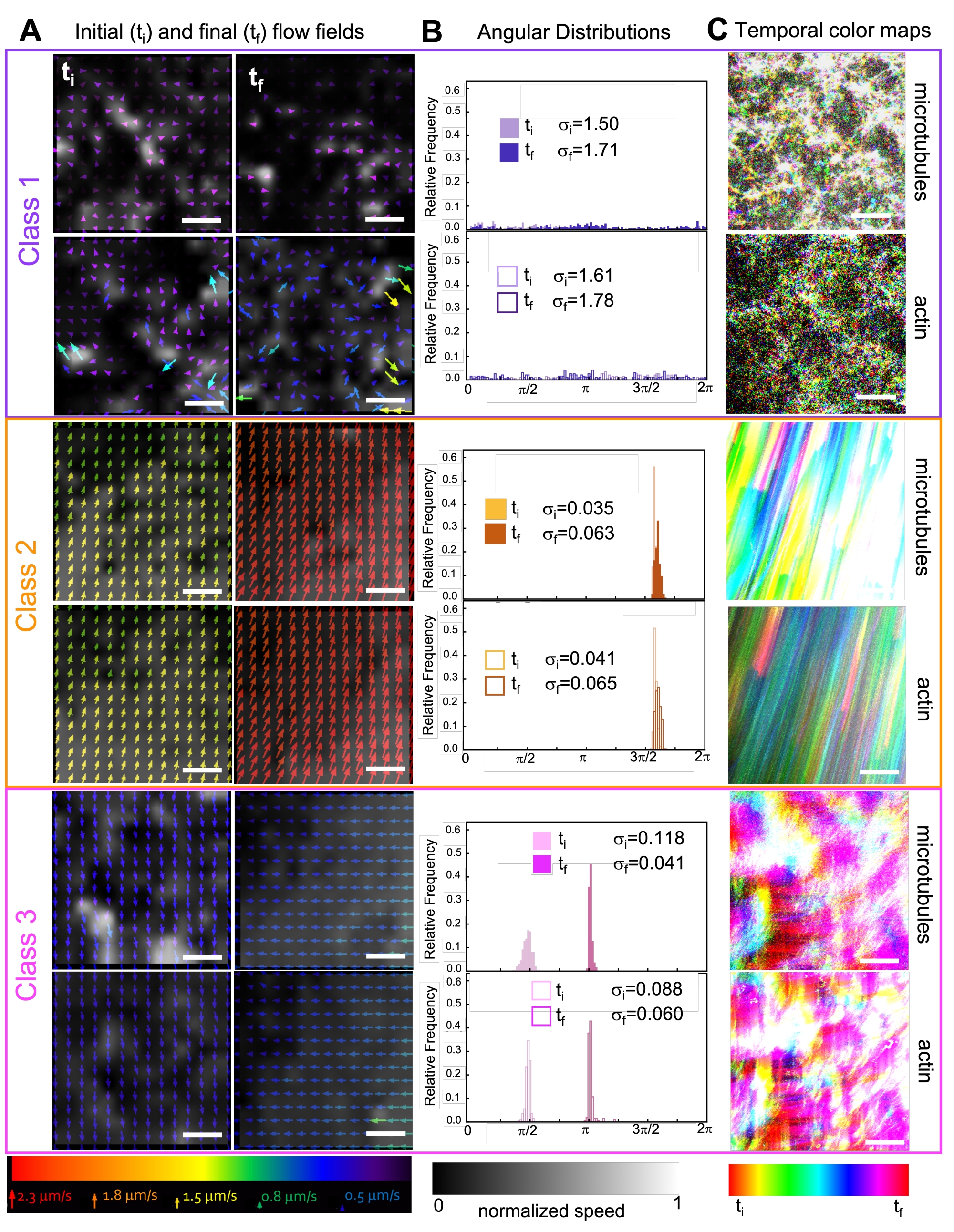
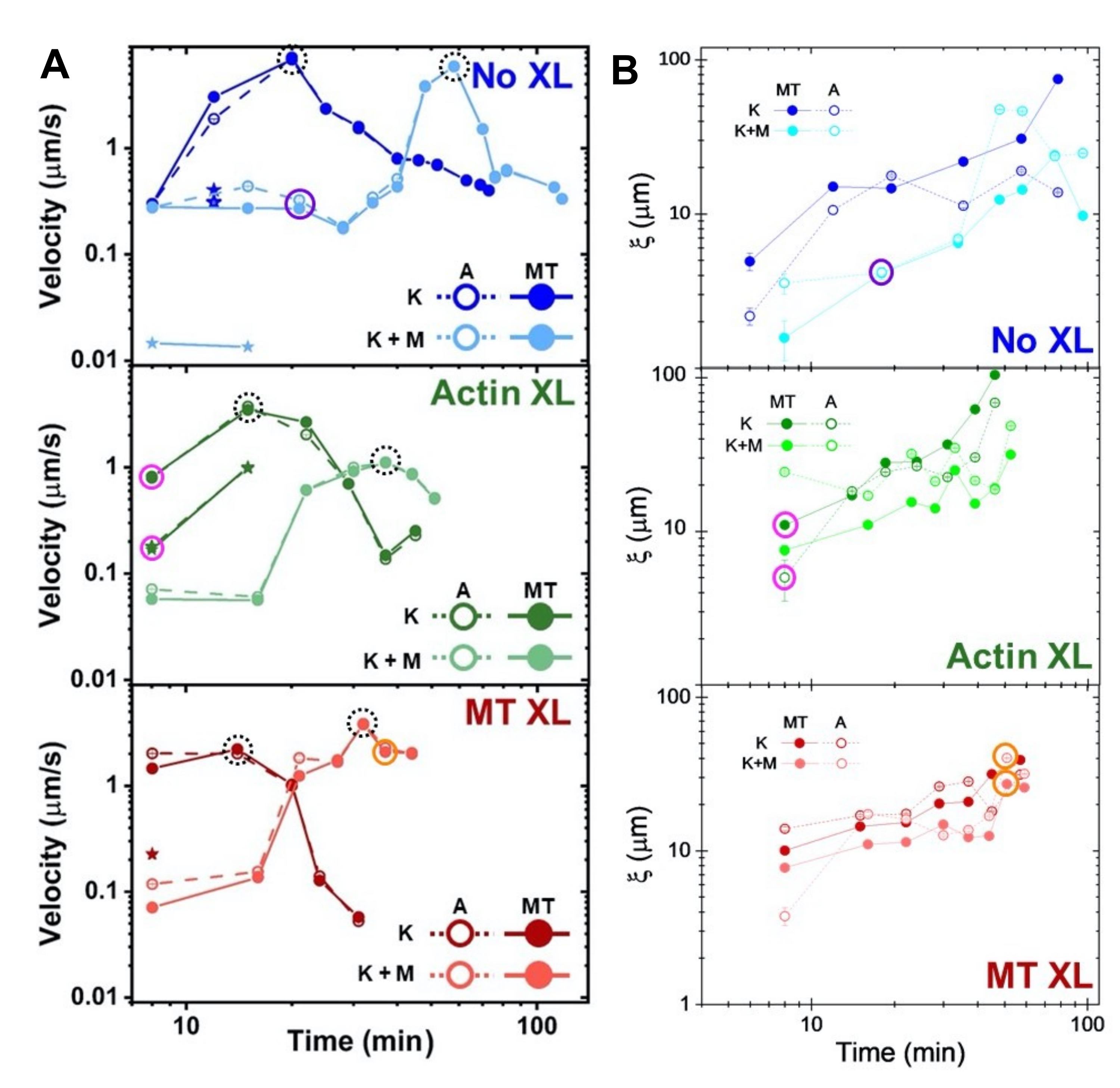
يؤدي دمج الكينيسين في المركبات في البداية إلى ديناميكيات وإعادة هيكلة مماثلة للمركبات التي يحركها الميوسين كما هو موضح في الصف العلوي من الشكل 7 (الفئة 1). ومع ذلك ، تنتقل الديناميكيات عادة إلى تدفق متباين الخواص على نطاق واسع (الشكل 7 الصف الأوسط ، الفئة 2) ، والتسارع ، والتباطؤ (الشكل 7 الصف السفلي ، الفئة 3). تقترن هذه الخصائص بالتجميع والتجميع المتوسط الحجم بعد 5-30 دقيقة (الشكل 6 والشكل 8 ب). تصور حقول التدفق التي تم إنشاؤها بواسطة PIV وخرائط الألوان الزمنية الموضحة في الشكل 7 أمثلة على إعادة الهيكلة الخواص (الفئة 1 ، اللوحة العلوية) ، والتدفق الموجه (الفئة 2 ، الألواح الوسطى) ، والتسارع ثنائي الاتجاه (الفئة 3 ، الألواح السفلية).
توضح سرعات الأكتين والأنابيب الدقيقة عند نقاط زمنية مختلفة أثناء النشاط ، والتي يتم تحديدها من خلال الملاءمة لمنحنيات τ (q) ، التسارع متبوعا بالتباطؤ (الشكل 8) ، والذي يعتمد على التشابك. كما هو موضح أيضا في الشكل 8 ، عندما يتم دمج كلا البروتينين الحركيين ، تكون الديناميكيات في الواقع أبطأ من مركبات الكينيسين فقط ، وهناك تأخر في ظهور التدفق المتوسط. يدعم الميوسين أيضا تغلغلا أكثر تجانسا لشبكات الأكتين والأنابيب الدقيقة طوال مدة النشاط ، بالإضافة إلى تجميع وإعادة هيكلة أقل. يمكن رؤية هذه التأثيرات في الصور في الشكل 6 ويتم قياسها كميا من خلال أطوال الارتباط المتغيرة زمنيا المحسوبة عبر SIA ، والتي تكون أصغر بشكل عام في وجود الميوسين (الشكل 8 ب).

الشكل 1. تصميم وتوصيف مركبات الأكتين والأنابيب الدقيقة النشطة مع محركات متعددة لتوليد القوة وروابط متشابكة سلبية. (أ) تتبلمر مونومرات الأكتين وثنائيات التوبولين بتركيزات مولية c A و c T من 0.73-11.6 μM والكسور المولية للأكتين Φ A = c A / (c A + c T) = 0 و 0.25 و 0.5 و 0.75 و 1 ، لتشكيل شبكات متشابكة من خيوط الأكتين (الخضراء) والأنابيب الدقيقة (الحمراء). يتم تحقيق التشابك السلبي باستخدام NA لربط خيوط الأكتين البيوتينيلة (Actin XL) أو الأنابيب الدقيقة (MT XL) عند الوصلات المتشابكة: نسب البروتين المولية R A = 0.01-0.08 و RMT= 0.001-0.01 للأكتين والأنابيب الدقيقة ، على التوالي. خيوط الميوسين-II الصغيرة (الأرجواني) ومجموعات الكينيسين (البرتقالية) ، بتركيزات c M = 0.12 - 0.48 μM و cK = 0.2 - 0.7 μM ، ادفع واسحب الخيوط لإخراج المركبات من الحالة المستقرة. (ب) رسم تخطيطي لحيز الصياغة. يتم دمج خيوط Myosin II الصغيرة (M) أو مجموعات الكينيسين (K) أو كلا المحركين (K + M) في المركبات بدون روابط متشابكة سلبية (No XL) ، وروابط متقاطعة للأكتين والأكتين (Actin XL) ، وروابط متقاطعة للأنابيب الدقيقة والأنابيب الدقيقة (MT XL). لا يتم رسم جميع الرسوم الكاريكاتورية على نطاق واسع. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
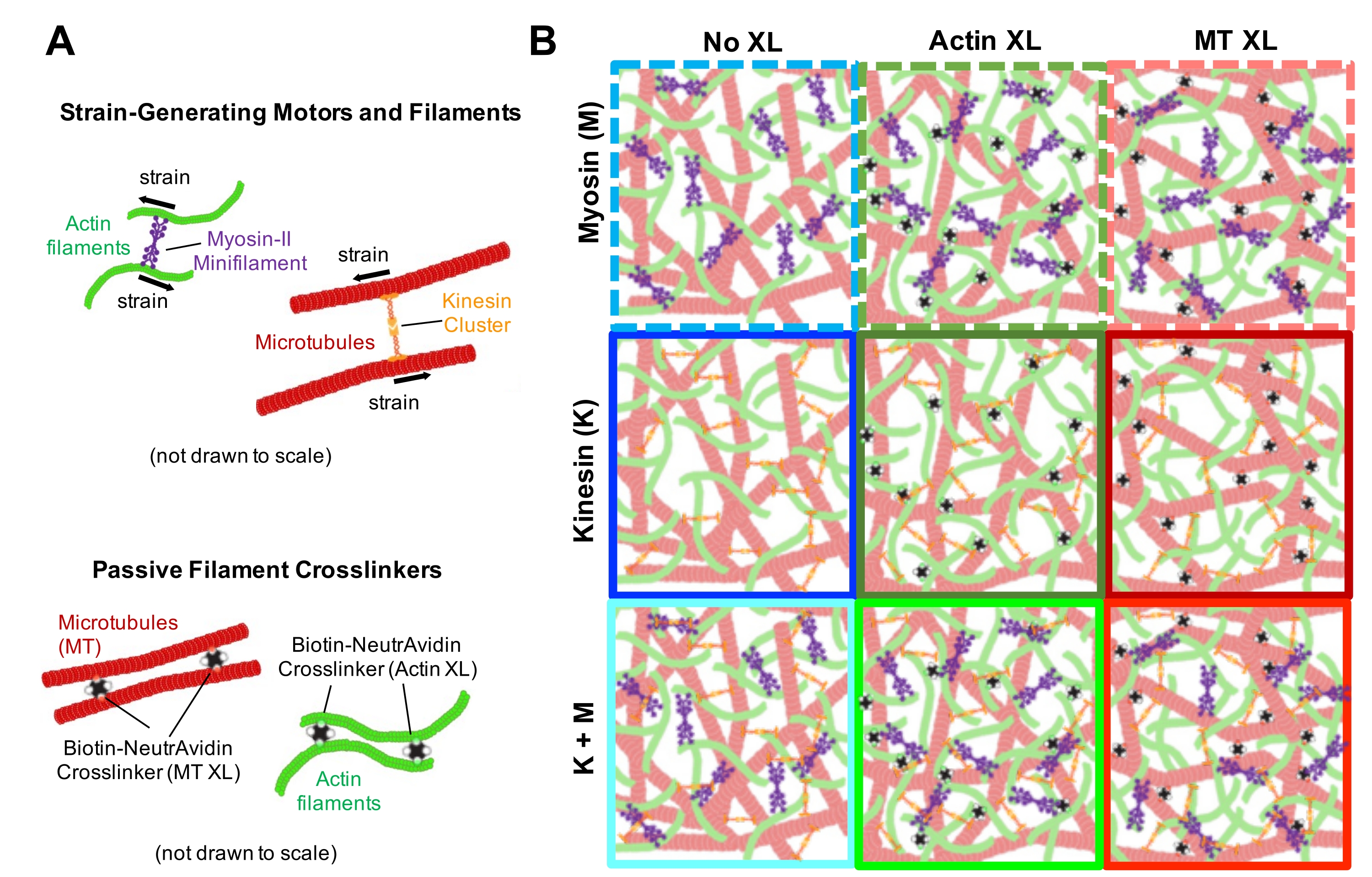{kind=link}

الشكل 2. التصوير متحد البؤر ثنائي اللون لمركبات الهيكل الخلوي التي يحركها الميوسين بتركيزات مختلفة من الميوسين cM وكسور الأكتين المولية ΦA. (أ) 256 × 128 بكسل مربع (212 × 106 ميكرومتر مربع) توضح صور الفحص المجهري متحد البؤر ثنائي اللون كيف يتم إعادة ترتيب مركبات خيوط الأكتين (الخضراء) والأنابيب الدقيقة (الحمراء) من خلال النشاط الحركي للميوسين. لا توجد محركات kinesin أو روابط متشابكة سلبية. في كل لوحة ، يتم عرض الصور التي تم التقاطها في البداية (اليسار ، قبل) والنهاية (اليمين ، بعد) لتنشيط الميوسين لمدة 45 دقيقة (عبر الإضاءة بضوء 488 نانومتر لإلغاء تنشيط blebbistatin). يتم ترتيب الألواح عن طريق زيادة التركيز المولي للميوسين (cM) ، والانتقال من اليسار إلى اليمين ، وزيادة الجزء المولي من الأكتين (ΦA) ، والانتقال من أعلى إلى أسفل. تتطابق الألوان التي تحدد كل لوحة مع الترميز اللوني المستخدم في الشكل 4 والشكل 5. قضبان المقياس 50 ميكرومتر. لالتقاط ديناميكيات وهيكل التحليل بشكل أفضل ، نستخدم معدلات إطارات من 1-5 إطارات في الثانية ، وعائد استثمار بجوانب 50-250 ميكرومتر ، وفترات سلاسل زمنية من 5-45 دقيقة ، اعتمادا على معدل الانكماش وإعادة الترتيب. تشير اللوحات التي تبدو فيها الصور قبل وبعد متشابهة إلى الحد الأدنى من إعادة الهيكلة، كما هو موضح في اللوحات الوردية والأرجوانية والسماوية. يمكن رؤية التجميع على نطاق صغير ، كما يتضح من زيادة عدم التجانس ووجود ميزات نقطية ساطعة ، في الألواح البرتقالية والخضراء والحمراء. الانكماش على نطاق واسع ، الذي ينظر إليه على أنه شبكة متقلصة بشكل موحد ، واضح في الألواح الزرقاء والأرجوانية. () عدل هذا الرقم من المرجع17. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}

الشكل 3. يظهر قياس سرعة صورة الجسيمات (PIV) أن نشاط الأكتوميوسين يؤدي إلى ديناميكيات انقباضية منسقة للأكتين والأنابيب الدقيقة في المركبات المتشابكة معا. حقول تدفق PIV للأكتين (الصف العلوي) والأنابيب الدقيقة (الصف السفلي) في مركب يحركه الميوسين مع (ΦA ، cM) = (0.5 ، 0.24) في أوقات متزايدة خلال سلسلة زمنية مدتها 6 دقائق. تم إنشاء حقول التدفق باستخدام المكون الإضافي Fiji / ImageJ PIV مع وقت تأخير يبلغ 20 ثانية وتجميع 2 بكسل × 2 بكسل. يظهر كل من الأكتين والأنابيب الدقيقة حركة متسقة موجهة نحو المنطقة المركزية لمجال الرؤية طوال مدة الفيلم. أشرطة القياس في جميع الصور هي 50 ميكرومتر. تتوافق ألوان الأسهم المختلفة مع سرعات مختلفة كما هو موضح في مقياس اللون على يمين الحقول المتجهة. تم تعديل هذا الرقم من المرجع26. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}

الشكل 4. يقيس المجهر الديناميكي التفاضلي (DDM) معدل ونوع حركة الأكتين والأنابيب الدقيقة في المركبات النشطة. (أ) يتم إجراء DDM على الأنابيب الدقيقة (أعلى ، رموز مفتوحة) والأكتين (أسفل ، رموز مملوءة) قنوات السلاسل الزمنية لتحديد أوقات الاضمحلال المميزة τ مقابل الرقم الموجي q لكل من الأكتين (الرموز المملوءة) والأنابيب الدقيقة (الرموز المفتوحة) كما هو موضح سابقا17,26. تتبع جميع المنحنيات مقياس τ ~ q-1 ، مما يشير إلى الحركة الباليستية ، مع سرعات v التي يتم تحديدها عبر الملاءمة ل τ (q) = (vq) -1. تتوافق السرعات الأسرع مع قيم τ (q) الأصغر لأي q معين. تتوافق ألوان وأشكال الرموز مع مجموعات (Φ A ، cM) الموضحة في B. (B) يتم تحديد سرعات الانكماش v من خلال الملاءمة لمنحنيات τ (q) الموضحة في A ، والتي يتم حسابها في المتوسط خلال جميع أوقات التأخير لمدة كل سلسلة زمنية مدتها 45 دقيقة. (ج) يحدد DDM الذي تم حله زمنيا (trDDM) كيف تختلف الديناميكيات بمرور الوقت من خلال تقييم τ (q) للأكتين (الرموز المملوءة ، يسار) والأنابيب الدقيقة (الرموز المفتوحة ، يمين) لفترات متتالية مدتها 6 دقائق (يشار إليها بظلال مختلفة من نفس اللون) خلال وقت التنشيط البالغ 45 دقيقة. يتم تنفيذ trDDM لكل مجموعة (ΦA ، cM) (يشار إليها برموز وألوان مختلفة) كما هو موضح في وسيلة الإيضاح في أسفل اليمين. تتبع منحنيات τ (q) الموضحة في C قياسا واتجاهات مماثلة لتلك الموجودة في A ولكنها تظهر أيضا اعتمادا على الوقت لبعض التراكيب (Φ A ، cM) ، وعلى الأخص ل Φ A = 0.75. (د) تحدد سرعات انكماش خيوط الأكتين (الرموز المغلقة) والأنابيب الدقيقة (الرموز المفتوحة) من الانحناءات إلى منحنيات τ(q) المقابلة. تمثل أشرطة الخطأ في جميع المؤامرات الخطأ القياسي للقيم عبر ثلاث إلى خمس نسخ متماثلة. () عدل هذا الرقم من المرجع17. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
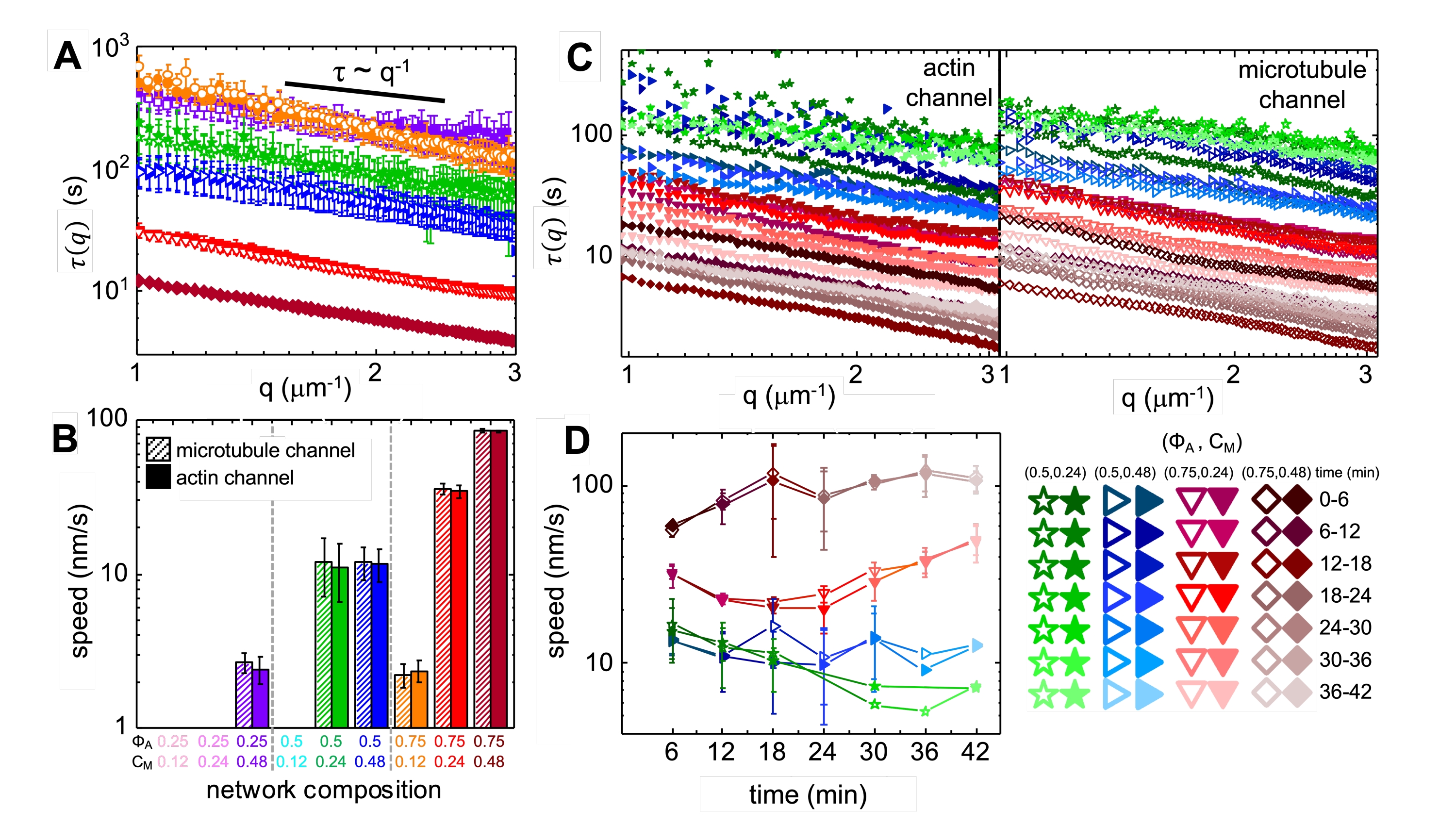{kind=link}

الشكل 5. يحدد تحليل الارتباط التلقائي للصور المكانية (SIA) إعادة الهيكلة الحركية للمركبات الهيكلية الخلوية النشطة. (أ) الارتباط الذاتي g (r) للأنابيب الدقيقة في البداية (يسار ، t = 0 دقيقة ، ظلال داكنة) والنهاية (يمين ، t = 42 دقيقة ، ظلال فاتحة) للتجربة لتركيبات (ΦA ، cM) المدرجة في وسيلة الإيضاح. أقحم: مثال يناسب البيانات في  الأوقات الأولية والنهائية ل (ΦA ، cM) = (0.75 ، 0.12). (ب) متوسط أطوال الارتباط ξ للأكتين (الرموز المغلقة) والأنابيب الدقيقة (الرموز المفتوحة) لكل (Φ A ، cM) محددة من خلال النوبات الأسية لكل منحنى g (r) ، كما هو موضح في الجزء الداخلي في A. تنقسم البيانات إلى تلك التي تظهر الحد الأدنى (يسار) مقابل إعادة هيكلة كبيرة (يمين). تمثل أشرطة الخطأ في A و B الخطأ القياسي عبر ثلاث إلى خمس نسخ متماثلة. () عدل هذا الرقم من المرجع17. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
الأوقات الأولية والنهائية ل (ΦA ، cM) = (0.75 ، 0.12). (ب) متوسط أطوال الارتباط ξ للأكتين (الرموز المغلقة) والأنابيب الدقيقة (الرموز المفتوحة) لكل (Φ A ، cM) محددة من خلال النوبات الأسية لكل منحنى g (r) ، كما هو موضح في الجزء الداخلي في A. تنقسم البيانات إلى تلك التي تظهر الحد الأدنى (يسار) مقابل إعادة هيكلة كبيرة (يمين). تمثل أشرطة الخطأ في A و B الخطأ القياسي عبر ثلاث إلى خمس نسخ متماثلة. () عدل هذا الرقم من المرجع17. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
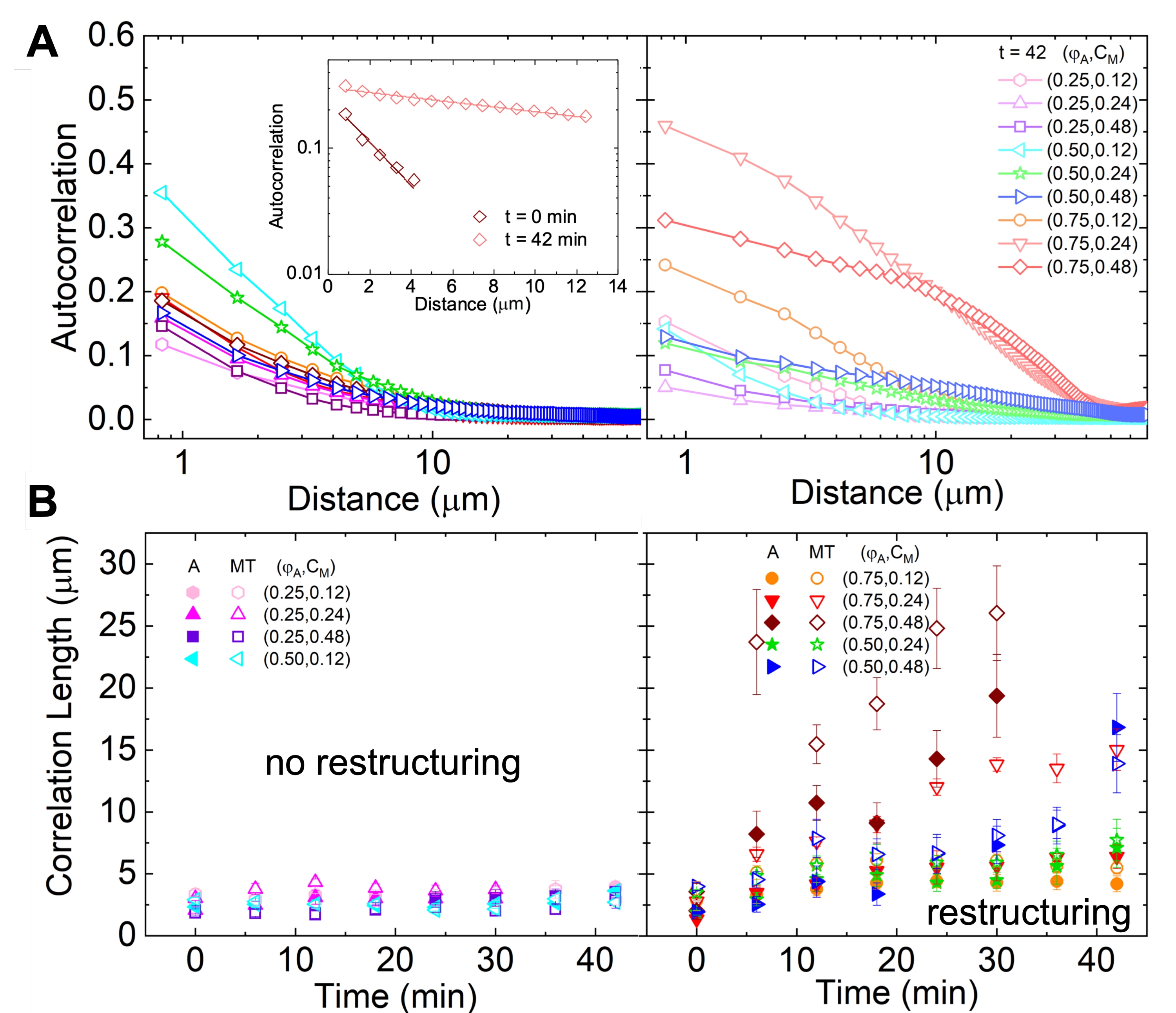{kind=link}

الشكل 6. دمج محركات kinesin والروابط المتشابكة السلبية في المركبات النشطة لزيادة قابلية البرمجة وتوسيع مساحة الطور للديناميكيات والهيكل. (أ) تظهر الصور متحدة البؤر ثنائية اللون للأكتين (الأخضر) والأنابيب الدقيقة (الحمراء) في المركبات النشطة إعادة هيكلة معقدة تعتمد على التركيب بمرور الوقت (مدرجة في دقيقة). تتوافق الصور الخمس في كل صف مع خمسة إطارات من سلسلة زمنية للإطار لعام 2000 تم الحصول عليها لمركب مدفوع بالكينيسين (K ، الصفوف 1 ، 3 ، 5) أو كينيسين وميوسين (K + M ، الصفوف 2 ، 4 ، 6) ، بما في ذلك إما عدم وجود روابط متقاطعة سلبية (No XL ، الصفوف 1 ، 2) ، روابط متقاطعة للأكتين - أكتين (Actin XL ، الصفوف 3 ، 4) ، أو روابط متقاطعة للأنابيب الدقيقة - الأنابيب الدقيقة (MT XL ، الصفوف 5 ، 6). قضبان المقياس كلها 50 ميكرومتر. تتطابق ألوان المخطط التفصيلي مع نظام الألوان في الشكل 8. (ب) تظهر قنوات تألق الأكتين والأنابيب الدقيقة المنفصلة لمركبات الكينيسين فقط تراكيب متنوعة مع كل من التوطين المشترك للأكتين-MT وفصل الطور الجزئي. الصور المعروضة هي للمركبات مع c A = 2.32 μM ، c T = 3.48 μM ، c K = 0.35 μM ، cM = 0.47 μM (الصفوف 2 ، 4 ، 6) ، R A = 0.02 (الصفوف 3 ، 4) ، و RMT = 0.005 (الصفوف 5 ، 6). تبدأ جميع المركبات بشبكات متداخلة موزعة بشكل موحد من الأكتين والأنابيب الدقيقة (العمود 1). تشكل المركبات التي يحركها Kinesin بدون روابط متشابكة (الصف 1) مجموعات غير متبلورة متصلة بشكل فضفاض وغنية ب MT. يتشارك الأكتين في توطين مراكز هذه المجاميع في البداية ولكن بعد ذلك يتم ضغطه خارج المناطق الغنية ب MT التي تستمر في الانكماش والانفصال عن بعضها البعض. يعيق تشابك الأكتين-الأكتين (الصف 3) هذا الفصل المجهري للأكتين-MT ، وبدلا من ذلك يتم توصيل الركام الغني ب MT عبر خيوط طويلة من الأكتين. يتيح تشابك الأكتين أيضا امتصاصا بطيئا للأكتين في المناطق الغنية ب MT ، بحيث يصبح المركب شبكة متصلة من مجموعات الأكتين و MT المترجمة المشتركة. يؤدي تشابك الأنابيب الدقيقة (الصف 5) إلى تجميع غير متبلور ل MTs التي تتجمع بمرور الوقت ، مما يؤدي إلى فصل طور أوسع نطاقا للأكتين و MTs. تؤدي إضافة الميوسين (الصفوف 2 و 4 و 6) إلى تقليل إزالة الخلط وإعادة الهيكلة المدفوعة بالكينيسين. بدون الروابط المتشابكة (الصف 2) ، تظهر المركبات القليل من إعادة الترتيب على مدار ساعات. يزيد التشابك من إعادة الهيكلة والتوطين المشترك للأكتين والأنابيب الدقيقة (الصفوف 4 و 6). على وجه التحديد ، عندما يتم ربط الأنابيب الدقيقة (الصف 6) ، هناك تداخل كبير وإعادة تنظيم في شبكات ألياف تشبه الويب. تم تعديل هذا الرقم من المرجع50. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
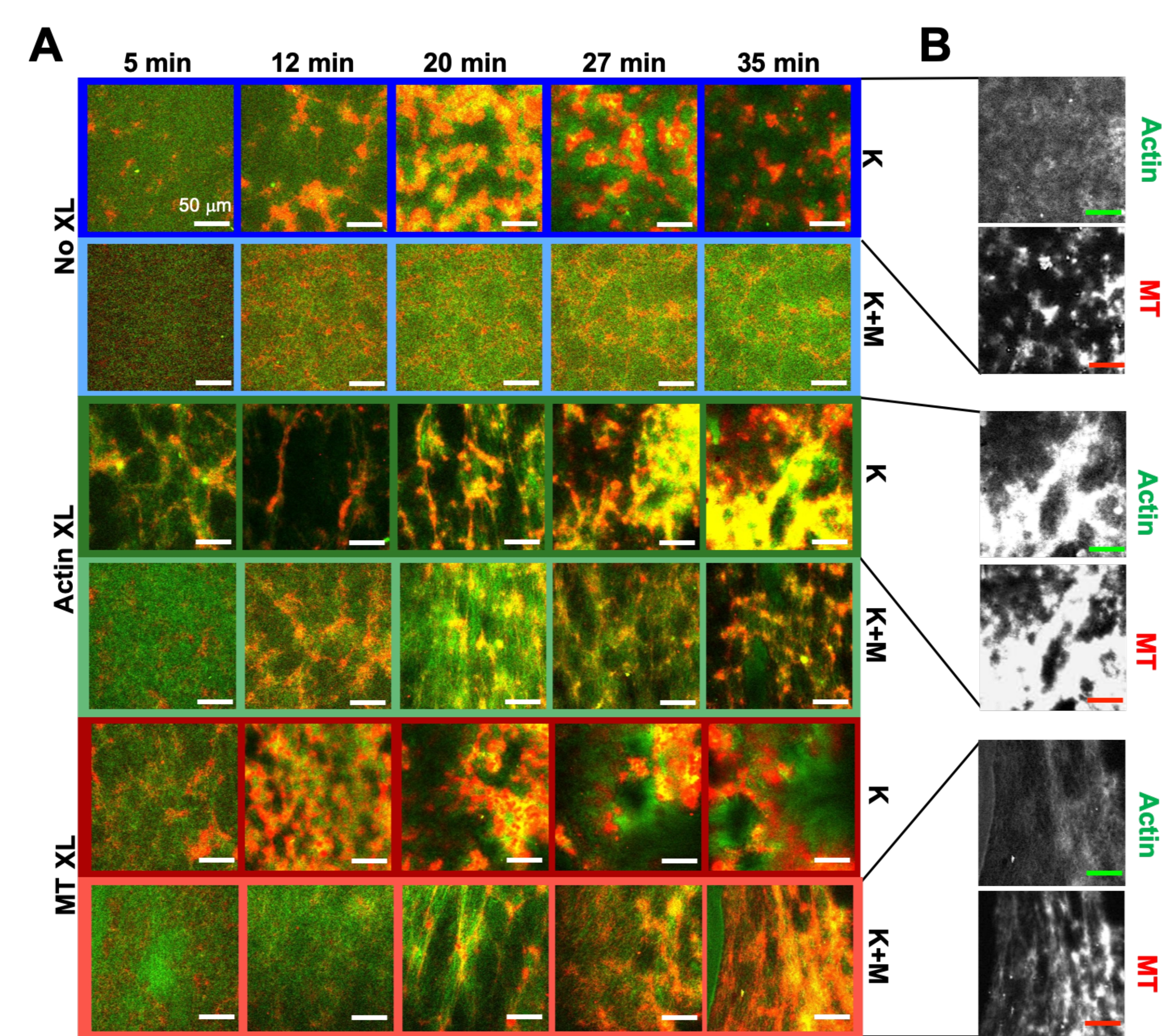{kind=link}

الشكل 7. يوضح PIV أن المركبات النشطة تظهر ثلاث فئات من حقول التدفق المتميزة زمانيا في المكان. (أ) حقول تدفق PIV للإطارات الأولى (t i) والأخيرة (tf) لثلاث سلاسل زمنية تمثيلية ، توضح الفئات الديناميكية المختلفة التي تعرضها المركبات الموضحة في الشكل 6. حقول تدفق PIV للأنابيب الدقيقة (أعلى) والأكتين (أسفل) للفئة 1 (أعلى ، أرجواني) ، الفئة 2 (وسط ، برتقالي) ، والفئة 3 (أسفل ، أرجواني) أمثلة على مقاطع الفيديو ، مع ألوان الأسهم المقابلة لمقياس السرعة العالمي في الأسفل ، وخريطة ألوان التدرج الرمادي التي توضح توزيع السرعة المكانية ، تمت تسويتها بشكل منفصل لكل حقل تدفق وفقا للمقياس الموضح في الأسفل. قضبان المقياس كلها 50 ميكرومتر. (ب) التوزيعات الزاوية لمتجهات السرعة من A (بوحدات الراديان) مع الانحرافات المعيارية الأولية والنهائية المدرجة σ i و σf. (ج) تظهر خرائط الألوان الزمنية لمقاطع الفيديو التي تم تحليلها في A و B موضع الإطار إلى الإطار لكل بكسل بالنسبة إلى نقطة البداية. تظهر خرائط الفئة 1 حركة عشوائية صغيرة الحجم. تصور خرائط الفئة 2 حركة سريعة أحادية الاتجاه مع الحد الأدنى من الاختلاف المكاني أو الزماني ؛ تعرض خرائط الفئة 3 ميزات كل من الفئة 1 و 2. تم تعديل هذا الرقم من المرجع50. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}

الشكل 8. يقيس DDM و SIA الديناميكيات المتغيرة زمنيا وهيكل مركبات الأكتين والأنابيب الدقيقة ثنائية المحركات. (أ) تظهر سرعات المركبات الموصوفة في الشكل 6 والشكل 7، المقاسة عبر DDM، تسارع وتباطؤ المركبات، المبرمجة عن طريق التشابك ونشاط الميوسين. يتم رسم سرعات الأنابيب الدقيقة (MT ، الدوائر المغلقة) والأكتين (A ، الدوائر المفتوحة) كدالة لوقت النشاط في المركبات التي لا تحتوي على تشابك (أعلى ، أزرق) ، تشابك الأكتين (وسط ، أخضر) ، تشابك الأنابيب الدقيقة (أسفل ، أحمر) ، بدون الميوسين (K ، ظلال أغمق) ، ومع الميوسين (K + M ، ظلال أفتح). بالنسبة لحالات الفئة 3 ، التي لها سرعتان ، يشار إلى السرعة الأبطأ بنجمة. تتوافق نقاط البيانات المحاطة بدوائر سوداء متقطعة مع السرعة القصوى vmaxلكل صياغة. أشرطة الخطأ (معظمها أصغر من أن نرى) هي الخطأ القياسي على نوبات قانون القوة ل τ (q) المقابلة. (ب) أطوال الارتباط الهيكلي ξ ، المحددة بواسطة SIA ، مقابل وقت النشاط ، لنفس مجموعة السلاسل الزمنية التي تم تقييمها في A. كل نقطة بيانات هي متوسط أطوال الارتباط المحددة للإطار الأول والأخير من السلسلة الزمنية المقابلة. بشكل عام ، تزداد ξ في الوقت لكل من الأكتين والأنابيب الدقيقة في جميع أنظمة المركبات ، والمركبات التي تحركها الكينيسين فقط لها أطوال ارتباط أكبر من تلك التي يوجد فيها الميوسين أيضا. نقاط البيانات في A و B التي تتوافق مع السلاسل الزمنية الثلاثة التي تم تحليلها في الشكل 7 محاطة بدائرة بلون الفئة المقابل (1 = أرجواني ، 2 = برتقالي ، 3 = أرجواني). تم تعديل هذا الرقم من المرجع50. الرجاء الضغط هنا لعرض نسخة أكبر من هذا الشكل.
{kind=link}
Discussion
يتمثل التقدم الرئيسي للنظام المعاد تشكيله الموصوف أعلاه في نمطيته وقابليته للضبط ، لذلك يتم تشجيع المستخدمين على تعديل تركيزات البروتينات والمحركات والوصلات المتشابكة وما إلى ذلك لتناسب النتائج المرجوة ، سواء كان ذلك لمحاكاة عملية خلوية معينة أو هندسة مادة ذات وظائف محددة أو خصائص ميكانيكية. يتم تعيين القيود المفروضة على نطاق تركيز الأكتين والتوبولين عند الحد الأدنى من خلال التركيز الحرج اللازم لبلمرة الأكتين (~ 0.2 ميكرومتر) 57،58،59 والتوبولين (~ 3 - 4 ميكرومتر)60 ، وعند الحد الأعلى من خلال الانتقال إلى المحاذاة النيماتيكية لخيوط الأكتين (~ 90 ميكرومتر) 61،62 أو الأنابيب الدقيقة (~ 35 ميكرومتر)63 . يجب بلمرة مونومرات الأكتين وثنائيات التوبولين في خيوط معا ، بدلا من خلطها معا بعد البلمرة ، للتأكد من أنها تشكل شبكات متجانسة متداخلة تدعم بعضها البعض بشكل تآزري. تعتمد الديناميكيات الجديدة التي تعرضها المركبات على هذا التفاعل. في حين أنه من المهم بشكل عام اتباع جميع الخطوات كما هو موضح في البروتوكول لإعادة إنتاج النتائج المعروضة بنجاح ، فإن بعض الخطوات أكثر دقة ، في حين أن البعض الآخر لديه مجال للتعديل والتعديل ليناسب الاحتياجات المحددة والموارد المتاحة.
على سبيل المثال ، تتمثل إحدى الخطوات المهمة في ضمان النتائج القابلة للتكرار في إعداد الكواشف وتخزينها بشكل صحيح باتباع الإرشادات الواردة في جدول المواد. البروتينات الهيكلية الخلوية (الأكتين ، التوبولين ، الميوسين ، الكينيسين) قابلة للذوبان ويجب اقتباسها وتجميدها بالنيتروجين السائل وتخزينها عند -80 درجة مئوية في حصص تستخدم مرة واحدة. بمجرد إزالتها من -80 درجة مئوية ، يجب الاحتفاظ بالقلص على الجليد. لا تحتفظ البروتينات الهيكلية الخلوية بوظيفتها بشكل موثوق بعد دورات التجميد والذوبان الإضافية.
الأنابيب الدقيقة أكثر حساسية لإزالة البلمرة وتغيير طبيعة الأكتين. بمجرد إزالته من -80 درجة مئوية ، يجب حفظ التوبولين على الثلج قبل البلمرة ، واستخدامه في غضون 12 ساعة. بمجرد البلمرة ، يجب حفظ الأنابيب الدقيقة في درجة حرارة الغرفة. من الأهمية بمكان أيضا تثبيت الأنابيب الدقيقة مع التاكسول لمنع إزالة البلمرة. يعد تثبيت Phalloidin لخيوط الأكتين مهما أيضا لقمع جهاز المشي الأكتين المستهلك ل ATP والذي يتنافس مع نشاط الميوسين والكينيسين.
يعد الطرد المركزي الفائق لمحركات الميوسين خطوة حاسمة أخرى ، لأنه يزيل رؤوس الميوسين الميتة غير النشطة. يؤدي عدم إزالة المونومرات غير النشطة إنزيميا إلى تشابك سلبي لشبكة الأكتين وفقدان النشاط. لإطالة نشاط ATPase للمحركات ، يمكن دمج نظام تجديد ATP مثل فوسفات الكرياتين وفسفوكينازالكرياتين 64 .
أخيرا ، يتطلب الحفاظ على النشاط المركب تثبيط امتصاص الخيوط والمحركات على جدران غرفة العينة ، والتي يمكن تحقيقها عن طريق تخميل أغطية المجهر والشرائح. البروتينات الحركية معرضة بشكل خاص للامتزاز ، مما يؤدي إلى سحب المركب إلى سطح غرفة العينة ، والخروج من مجال الرؤية ، والانهيار إلى 2D ، ولم يعد يخضع للنشاط. يعد Silanizing الأغطية والشرائح طريقة فعالة لتخميل الأسطح ومنع الامتزاز (انظر الخطوة 1). طريقة التخميل البديلة المستخدمة بشكل فعال في تجارب الهيكل الخلوي في المختبر هي طلاء السطح بطبقة ثنائية دهنية ، على غرار غشاء الخلية18. هذه الطريقة مفيدة إذا رغب المرء في ربط البروتينات بالسطح أو إدخال تفاعلات أخرى محددة بين البروتين والسطح ، لأنه يمكن تشغيل الطبقة الثنائية. بالنسبة لتجارب الملقط البصري ، يعد تخميل الكريات المجهرية أمرا بالغ الأهمية أيضا ، ويمكن تحقيقه عن طريق طلاء الكريات المجهرية الكربوكسيلية باستخدام BSA أو PEG عبر كيمياء carbodiimide crosslinker48.
هناك بعض جوانب البروتوكولات المقدمة التي قد يفكر الباحثون في تغييرها لتناسب احتياجاتهم. أولا ، قد يختار الباحثون استبدال روابط البيوتين-NA غير الأصلية بروابط متشابكة بيولوجية ، مثل alpha-actinin أو MAP65 التي تربط الأكتين والأنابيب الدقيقة ، على التوالي28،65،66. إن الدافع وراء استخدام الروابط المتشابكة غير الأصلية في المركبات الموصوفة هنا هو تعزيز قابليتها للتكاثر والاستقرار وقابلية الضبط مقارنة بالوصلات المتشابكة الأصلية. بسبب رابطة البيوتين-NA القوية ، يمكن افتراض أن الروابط المتشابكة دائمة ، بدلا من معظم الروابط المتشابكة الأصلية التي ترتبط بشكل عابر بمعدلات دوران واسعة النطاق. تعمل ديناميكيات الربط المتشابك العابر على تعقيد تحليل المساهمات من الروابط المتشابكة والمحركات إلى الديناميكيات. علاوة على ذلك ، يمكن استخدام روابط البيوتين-NA بشكل متعدد الاستخدامات لربط كل من الأكتين والأنابيب الدقيقة ، وكذلك ربط الأكتين بالأنابيب الدقيقة. بهذه الطريقة ، يمكن إجراء مقارنة لا لبس فيها بين الزخارف المتشابكة ، مع الحفاظ على جميع المتغيرات الأخرى (على سبيل المثال ، حجم الروابط المتشابكة ، وتقارب الربط ، والقياس المتكافئ ، وما إلى ذلك) ثابتة. أخيرا ، الكواشف اللازمة لدمج روابط البيوتين-NA متاحة تجاريا على نطاق واسع ، وذات خصائص جيدة ، وتستخدم بشكل شائع في العديد من مختبرات الفيزياء الحيوية. ومع ذلك ، فإن إحدى نقاط القوة الرئيسية للمنصة المختبرية الموصوفة هنا هي نمطيتها ، لذلك يجب أن يكون الباحثون قادرين على استبدال روابط البيوتين-NA بسلاسة بروابط أصلية إذا اختاروا ذلك.
ثانيا ، في البروتوكول الحالي ، يتم بلمرة مونومرات الأكتين وثنائيات التوبولين في خيوط معا في أنبوب طرد مركزي قبل إضافتها إلى غرفة العينة. قد يؤدي تدفق محلول البروتينات الخيطية المتشابكة إلى غرفة العينة إلى محاذاة التدفق ، خاصة الأنابيب الدقيقة ، مما يكسر الخواص المتجانسة المطلوبة وتجانس المركبات. في الواقع ، كان التقدم الرئيسي في العمل السابق على مركبات الأكتين والأنابيب الدقيقة في الحالة المستقرة هو القدرة على البلمرة المشتركة للأكتين والأنابيب الدقيقة في الموقع (في غرفة العينة) لضمان تكوين شبكات متداخلة الخواص من الأكتين والأنابيب الدقيقة15،16،27. ومع ذلك ، فإن توسيع هذا النهج ليشمل المركبات النشطة يتطلب إضافة المحركات إلى العينة قبل بلمرة الأكتين والتوبولين واحتضان العينة بأكملها معا عند 37 درجة مئوية قبل التجارب. وقد أدت اختبارات هذا الاختلاف في البروتوكول إلى انخفاض بلمرة الأكتين وعدم وجود نشاط حركي ملحوظ ، ويرجع ذلك على الأرجح إلى نشاط ATPase المنافس وحضانة المحركات المطولة عند 37 درجة مئوية. لحسن الحظ ، لا توجد محاذاة تدفق ملحوظة للمركبات عند اتباع البروتوكولات الحالية ، كما يتضح من الشكل 2 والشكل 3 والشكل 6. ومع ذلك ، يتم تشجيع الباحثين على تصميم بروتوكولات تسمح بتكوين مركبات نشطة في الموقع.
نقطة أخرى يجب مراعاتها هي مخطط وضع العلامات الفلورية ، والذي يستلزم وضع علامات قليلة على جميع خيوط الأكتين والأنابيب الدقيقة في الشبكة. تم تحسين نهج وضع العلامات هذا لتصور بنية الشبكة مباشرة بدلا من استنتاج البنية والديناميكيات عبر خيوط التتبع أو الكرات المجهرية. ومع ذلك ، فإن المقايضة هي أن الخيوط الفردية ليست ذات علامات زاهية وقابلة للحل. تتمثل إحدى الطرق التي يمكن للباحثين اتباعها لحل خيوط مفردة وكذلك تصور بنية الشبكة في تخدير خيوط مسبقة التشكيل تحمل علامة فلوروفور آخر ، بحيث يمكن تصوير كل من الشبكة المحيطة والخيوط الفردية في وقت واحد. ومع ذلك ، عند استخدام أكثر من اثنين من الفلوروفورات وقنوات الإثارة / الانبعاث ، غالبا ما يكون من الصعب القضاء على النزيف بين القنوات ، لذلك يجب توخي الحذر عند اختيار الفلوروفورات والمرشحات وشدة الليزر.
أحد القيود ذات الصلة هو عدم القدرة على تصور محركات الميوسين أو الكينيسين في المركبات. مونومرات الأكتين الموسومة بالفلورسنت وثنائيات التوبولين المستخدمة متاحة تجاريا ، في حين أن تصور الميوسين أو الكينيسين في المركبات يتطلب وضع العلامات الداخلية. يتم تشجيع الباحثين على اتخاذ الخطوة التالية لتسمية المحركات ، كما حدث سابقا18,67 ، ليكونوا قادرين على ربط النشاط الحركي بشكل لا لبس فيه والارتباط بالديناميكيات والهياكل التي تعرضها مركباتنا.
أخيرا ، من المهم ملاحظة أنه في البروتوكول الحالي ، لا يتم التحكم في بداية ومدة نشاط kinesin. نظرا لأن نشاط الميوسين يتم التحكم فيه باستخدام التعطيل الضوئي للبليبيستاتين ، كما هو موضح أعلاه ، لبناء تنشيط ضوئي مماثل للكينيسين ، يمكن للمرء دمج ATP المنشط بالضوء.
لبناء تعقيد التصاميم الموصوفة هنا ، لتقليد الظروف الخلوية بشكل أفضل وتوسيع مساحة معلمة البنية الديناميكية والوظيفة ، سيركز العمل المستقبلي على دمج خيوط وسيطة ، مثل vimentin68,69 ، بالإضافة إلى محركات أخرى مثل dynein13,70. سيتم أيضا دمج Gelsolin بتركيزات مختلفة للتحكم في طول الأكتين14 ، بالإضافة إلى بروتين تاو للتحكم في تصلب الأنابيب الدقيقة.
باختصار ، تصف البروتوكولات المقدمة كيفية تصميم وإنشاء وتوصيف ديناميكيات وهيكل وميكانيكا أنظمة المادة النشطة المستوحاة من الهيكل الخلوي ، والتي تحتوي على مكونين منفصلين لتوليد القوة النشطة يعملان على ركائز مختلفة في نظام واحد. تعمل هذه المنصة القابلة للضبط والمعيارية على تقريب جهود إعادة التكوين خطوة مهمة من محاكاة الهيكل الخلوي الخلوي وتوفر القدرة الفريدة على برمجة خصائصه عبر مساحة مرحلة واسعة من خلال دمج المكونات المختلفة وإزالتها وضبطها بشكل مستقل. علاوة على ذلك ، فإن جميع مكونات هذا النظام متعدد الاستخدامات متاحة تجاريا (انظر جدول المواد) ، باستثناء ثنائيات كينيسين التي يتم تنقيتها في مختبر روس ، كما هو موضح سابقا50 ، ومتاحة عند الطلب. أخيرا ، تتوفر جميع أكواد التحليل مجانا من خلال GitHub49 وتستند إلى لغات وبرامج برمجة مجانية (Python و Fiji). نأمل أن يؤدي النشر الشفاف للبروتوكولات لتصميم هذه الأنظمة إلى جعل هذه المنصة في متناول مجموعة متنوعة من المستخدمين ذوي الخبرات والخلفيات والانتماءات المؤسسية والأهداف البحثية المختلفة.
Disclosures
ليس لدى المؤلفين ما يكشفون عنه
Acknowledgements
نعرب عن تقديرنا لمايا هنديا والدكتور جوناثان ميشيل للمساعدة في تحليل البيانات ، والدكتورة جانيت شيونغ والدكتورة موميتا داس والدكتور مايكل روست على المناقشات والإرشادات المفيدة. تم دعم هذا البحث من خلال منحة أبحاث مؤسسة William M. Keck وجائزة NSF DREF (DMR 2119663) الممنوحة ل RMRA و JLR والمعاهد الوطنية للصحة R15 المنح (R15GM123420 ، 2R15GM123420-02) الممنوحة ل RMR-A و RJM.
Materials
| Name | Company | Catalog Number | Comments |
| (-)-Blebbistatin Abbreviation used in paper: blebbistatin | Sigma Aldrich | B0560 | Stock Concentration: 200 μM in DMSO Storage: dessicated, in DMSO, -20ºC Stock and Experiment Recipes: dissolve 1 mg of powder to 200 μM in DMSO Storage, Handling, Troubleshooting Notes: limited shelf-life, typically stops functioning reliably after 3-4 months. purchase and prepare new solution every 3 months. |
| 1:20 488-tubulin:tubulin mixture Abbreviation used in paper: 5-488-tubulin | NA | NA | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: mix tubulin and 488-tubulin at a 20:1 ratio, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| 1:20 R-tubulin:tubulin mixture Abbreviation used in paper: 5-R-tubulin | NA | NA | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: mix tubulin and rhodamine tubulin at a 20:1 ratio, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| actin (biotin): skeletal muscle Abbreviation used in paper: biotin-actin | Cytoskeleton | AB07 | Stock Concentration: 1 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 1 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: (1) immediately prior to use dilute to 0.5 mg/ml in PEM, (2) once removed from -80ºC, store aliquot on ice at 4ºC for up to 1 week |
| actin (rhodamine): rabbit skeletal muscle Abbreviation used in paper: R-actin | Cytoskeleton | AR05 | Stock Concentration: 1.5 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 1.5 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: once removed from -80ºC, store aliquot on ice at 4ºC, can be used for up to 1 week |
| adenosine triphosphate Abbreviation used in paper: ATP | Thermo Fisher Scientific | A1048 | Stock Concentration: 100 mM Storage: in solution (pH 7), -20ºC Stock and Experiment Recipes: reconsitute in DI H20, bring pH to 7 with NaOH Storage, Handling, Troubleshooting Notes: routinely check pH and adjust as needed, hydrolyzes over time, replace every ~6-12 months |
| AlexaFluor488 Phalloidin Abbreviation used in paper: 488-phalloidin | Thermo Fisher Scientific | A12379 | Stock Concentration: 100 μM DMSO Storage: protected from light, dessicated, -20ºC Stock and Experiment Recipes: reconstitute to 100 μM with DMSO Storage, Handling, Troubleshooting Notes: immediately prior to use dilute to 20 μM in PEM (1 μL in 4 μL PEM) |
| AlexaFluor488–labeled actin Abbreviation used in paper: 488-actin | Thermo Fisher Scientific | A12373 | Stock Concentration: 1.5 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 1.5 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: this item has been discontinued |
| Basic Plasma Cleaner Abbreviation used in paper: plasma cleaner | Harrick Plasma | PDC-32G | |
| Bemis Parafilm M Laboratory Wrapping Film Abbreviation used in paper: transparent film | Thermo Fisher Scientific | 13-374-5 | |
| D-(+)-Glucose Abbreviation used in paper: | Thermo Fisher Scientific | A1682836 | Stock Concentration: 100x Storage: store at stock concentration (100x) or 10x concentration, dessicated, at -20ºC Stock and Experiment Recipes: reconstitute powder to 4.5 mg/ml in DI H20 Storage, Handling, Troubleshooting Notes: final concentration in solution should 45 μg/mL |
| D-Biotin Abbreviation used in paper: biotin | Fisher Scientific | BP232-1 | Stock Concentration: 1.02 mM in PEM Storage: dessicated, 4ºC |
| deionized nanopure water Abbreviation used in paper: DI | |||
| Dimethyldichlorosilane Abbreviation used in paper: silane | Thermo Fisher Scientific | D/3820/PB05 | Stock Concentration: 2% dissolved in Toulene |
| Dithiothreitol Abbreviation used in paper: DTT | Thermo Fisher Scientific | R0861 | Stock Concentration: 1 M in DMSO Storage: dessicated, -20ºC Stock and Experiment Recipes: dilute to 2 mM in PEM immediately before each experiment |
| DMSO Anhydrous Abbreviation used in paper: DMSO | Thermo Fisher Scientific | D12345 | |
| F-Buffer Abbreviation used in paper: F-buffer | NA | NA | Stock Concentration: 10x Storage: dessicated, -20ºC Stock and Experiment Recipes: 10 mM Imidazole (pH 7.0), 50 mM KCl, 1 mM MgCl2, 1 mM EGTA, 0.2 mM ATP |
| G-Buffer Abbreviation used in paper: G-buffer | NA | NA | Stock Concentration: 10x Storage: dessicated, -20ºC Stock and Experiment Recipes: 2.0 mM Tris (pH 8), 0.2 mM ATP, 0.5 mM DTT, 0.1 mM CaCl2. Store at -20°C. |
| glass microscope slide Abbreviation used in paper: slide | Thermo Fisher Scientific | 22-310397 | |
| Glucose oxidase + catalase + β-mercaptoethanol Abbreviation used in paper: GOC | Sigma Aldrich | G2133-250KU, C1345, 63689 | Stock Concentration: 100x Storage: store at stock concentration (100x) or 10x concentration, dessicated, at -20ºC Stock and Experiment Recipes: For 100x: 4.3 mg/ml glucose oxidase, 0.7 mg/ml catalase, 0.5% v/v β-mercaptoethanol in DI H20 Storage, Handling, Troubleshooting Notes: final concentration in solution should be: 0.005% β-mercaptoethanol, 43 μg/mL glucose oxidase, 7 μg/mL catalase |
| glu-GOC oxygen scavenging system Abbreviation used in paper: glu-GOC | NA | NA | Stock Concentration: 100x Storage: prepare fresh each time Stock and Experiment Recipes: mix equal parts Glu and GOC and add at 1/100 final sample volume immediately before imaging Storage, Handling, Troubleshooting Notes: prepare from Glu and GOC immediately before imaging |
| Guanosine triphosphate Abbreviation used in paper: GTP | Thermo Fisher Scientific | R0461 | Stock Concentration: 100 mM Storage: 100 μL aliquots at -20ºC |
| Instant Mix 1-minute epoxy Abbreviation used in paper: epoxy | Loctite | 1366072 | |
| Kinesin-1 401 BIO 6x HIS Abbreviation used in paper: kinesin | Prepared in JL Ross Lab at Syracuse University | NA | Stock Concentration: 8.87 μM in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Storage, Handling, Troubleshooting Notes: biotinylated dimers form kinesin clusters, each aliquot can be used for up to 12 hrs stored on ice at 4ºC |
| NeutrAvidin Abbreviation used in paper: NA | Thermo Fisher Scientific | 31000 | Stock Concentration: 5 mg/ml in PEM Storage: dessicated, -20ºC Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM |
| No 1. glass coverslips (24 mm x 24 mm) Abbreviation used in paper: coverslip | Thermo Fisher Scientific | 12-548-CP | |
| Paclitaxel Abbreviation used in paper: Taxol | Thermo Fisher Scientific | P3456 | Stock Concentration: 2 mM in DMSO Storage: protected from light, dessicated, -20ºC Stock and Experiment Recipes: reconstitute to 2 mM with DMSO Storage, Handling, Troubleshooting Notes: immediately prior to use dilute to 200 μM in DMSO (0.4 μL in 3.6 μL DMSO) |
| PEM-100 Abbreviation used in paper: PEM | NA | NA | Stock Concentration: 1x Storage: room temperature (RT) Stock and Experiment Recipes: 100 mM K-PIPES (pH 6.8), 2 mM EGTA, 2 mM MgCl2 Storage, Handling, Troubleshooting Notes: use KOH to adjust pH to 6.8, recheck pH often and adjust accordingly |
| phalloidin Abbreviation used in paper: phalloidin | Thermo Fisher Scientific | P3457 | Stock Concentration: 100 μM in DMSO Storage: protected from light, dessicated, -20ºC, adhere closely to storage/handling conditions Stock and Experiment Recipes: reconstitute to 100 μM with DMSO Storage, Handling, Troubleshooting Notes: susceptible to impurities in its preparation and denaturing, identifiable as large amorphous aggregates of actin in samples |
| porcine brain tubulin Abbreviation used in paper: tubulin | Cytoskeleton | T240 | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC |
| Potassium Chloride Abbreviation used in paper: KCl | Thermo Fisher Scientific | AM9640G | Stock Concentration: 4 M Storage: RT |
| Rabbit skeletal actin Abbreviation used in paper: actin | Cytoskeleton | AKL99 | Stock Concentration: 2 mg/ml in G-buffer Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute to 2 mg/ml in G-buffer, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: once removed from -80ºC, store aliquot on ice at 4ºC, can be used for up to 1 week |
| Rabbit skeletal myosin II Abbreviation used in paper: myosin | Cytoskeleton | MY02 | Stock Concentration: 10 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 10 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: monomers form minifilaments at low KCl, each aliquot can be used for up to 12 hrs stored on ice at 4ºC |
| Tubulin (biotin): porcine brain Abbreviation used in paper: biotin-tubulin | Cytoskeleton | T333P | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: immediately prior to use dilute to 0.5 mg/ml in PEM |
| Tubulin (fluorescent HiLyte 488): porcine brain Abbreviation used in paper: 488-tubulin | Cytoskeleton | TL488M | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| tubulin (rhodamine): porcine brain Abbreviation used in paper: R-tubulin | Cytoskeleton | TL590M | Stock Concentration: 5 mg/ml in PEM Storage: single use aliquots, -80ºC, avoid freeze-thaw cycles Stock and Experiment Recipes: reconstitute powder to 5 mg/ml in PEM, flash freeze with LN2 Storage, Handling, Troubleshooting Notes: each aliquot can be used for up to 12 hrs stored on ice at 4ºC, protect from light |
| Tween 20 Abbreviation used in paper: Tween20 | Thermo Fisher Scientific | J20605.AP | Stock Concentration: 1% v/v in DI H20 Storage: RT |
| ultracentrifuge grade microtubes Abbreviation used in paper: Beckman-Coulter Optima Max XP | Beckman Coultier | 343776 | Storage, Handling, Troubleshooting Notes: 8x34 mm PC |
| UV light curing glue Abbreviation used in paper: UV glue | Pharda | SKG-2869 |
References
- Fletcher, D. A., Mullins, R. D. Cell mechanics and the cytoskeleton. Nature. 463 (7280), 485-492 (2010).
- Koenderink, G. H., Paluch, E. K. Architecture shapes contractility in actomyosin networks. Current Opinion in Cell Biology. 50, 79-85 (2018).
- Dogterom, M., Koenderink, G. H. Actin-microtubule crosstalk in cell biology. Nature Reviews Molecular Cell Biology. 20 (1), 38-54 (2019).
- Burla, F., Mulla, Y., Vos, B. E., Aufderhorst-Roberts, A., Koenderink, G. H. From mechanical resilience to active material properties in biopolymer networks. Nature Reviews Physics. 1 (4), 249-263 (2019).
- Wen, Q., Janmey, P. A. Polymer physics of the cytoskeleton. Current Opinion in Solid State and Materials Science. 15 (5), 177-182 (2011).
- Xiao, Q., Hu, X., Wei, Z., Tam, K. Y. Cytoskeleton molecular motors: structures and their functions in neuron. International Journal of Biological Sciences. 12 (9), 1083-1092 (2016).
- Ajeti, V. et al. Wound healing coordinates actin architectures to regulate mechanical work. Nature Physics. 15 (7), 696-705 (2019).
- Jung, W. et al. Dynamic motions of molecular motors in the actin cytoskeleton. Cytoskeleton. 76 (11-12), 517-531 (2019).
- Pollard, T. D., O'Shaughnessy, B. Molecular mechanism of cytokinesis. Annual Review of Biochemistry. 88 (1), 661-689 (2019).
- Huber, F., Boire, A., López, M. P., Koenderink, G. H. Cytoskeletal crosstalk: when three different personalities team up. Current Opinion in Cell Biology. 32, 39-47 (2015).
- Rivero, F. et al. The role of the cortical cytoskeleton: F-actin crosslinking proteins protect against osmotic stress, ensure cell size, cell shape and motility, and contribute to phagocytosis and development. Journal of Cell Science. 109 (11), 2679-2691 (1996).
- Duclos, G. et al. Topological structure and dynamics of three-dimensional active nematics. Science. 367 (6482), 1120-1124 (2020).
- Baclayon, M. et al. Optical tweezers-based measurements of forces and dynamics at microtubule ends. Optical Tweezers. 1486, 411-435 (2017).
- Gurmessa, B., Fitzpatrick, R., Falzone, T. T., Robertson-Anderson, R. M. Entanglement density tunes microscale nonlinear response of entangled actin. Macromolecules. 49 (10), 3948-3955 (2016).
- Francis, M. L. et al. Non-monotonic dependence of stiffness on actin crosslinking in cytoskeleton composites. Soft Matter. 15 (44), 9056-9065 (2019).
- Ricketts, S. N. et al. Varying crosslinking motifs drive the mesoscale mechanics of actin-microtubule composites. Scientific Reports. 9 (1), 12831 (2019).
- Lee, G. et al. Active cytoskeletal composites display emergent tunable contractility and restructuring. Soft Matter. 17 (47), 10765-10776 (2021).
- Murrell, M. P., Gardel, M. L. F-actin buckling coordinates contractility and severing in a biomimetic actomyosin cortex. Proceedings of the National Academy of Sciences. 109 (51), 20820-20825 (2012).
- Soares e Silva, M. et al. Active multistage coarsening of actin networks driven by myosin motors. Proceedings of the National Academy of Sciences. 108 (23), 9408-9413 (2011).
- Sonn-Segev, A., Bernheim-Groswasser, A., Roichman, Y. Dynamics in steady state in vitro acto-myosin networks. Journal of Physics: Condensed Matter. 29 (16), 163002 (2017).
- Ideses, Y., Sonn-Segev, A., Roichman, Y., Bernheim-Groswasser, A. Myosin II does it all: assembly, remodeling, and disassembly of actin networks are governed by myosin II activity. Soft Matter. 9 (29), 7127 (2013).
- Fürthauer, S. et al. Self-straining of actively crosslinked microtubule networks. Nature Physics. 15 (12), 1295-1300 (2019).
- Lemma, L. M. et al. Multiscale microtubule dynamics in active nematics. Physical Review Letters. 127 (14), 148001 (2021).
- Fan, Y., Wu, K.-T., Aghvami, S. A., Fraden, S., Breuer, K. S. Effects of confinement on the dynamics and correlation scales in kinesin-microtubule active fluids. Physical Review E. 104 (3), 034601 (2021).
- Triclin, S. et al. Self-repair protects microtubules from destruction by molecular motors. Nature Materials. 20 (6), 883-891 (2021).
- Lee, G. et al. Myosin-driven actin-microtubule networks exhibit self-organized contractile dynamics. Science Advances. 7 (6), eabe4334 (2021).
- Ricketts, S. N., Ross, J. L., Robertson-Anderson, R. M. Co-entangled actin-microtubule composites exhibit tunable stiffness and power-law stress relaxation. Biophysical Journal. 115 (6), 1055-1067 (2018).
- Bendix, P. M. et al. A quantitative analysis of contractility in active cytoskeletal protein networks. Biophysical Journal. 94 (8), 3126-3136 (2008).
- Linsmeier, I. et al. Disordered actomyosin networks are sufficient to produce cooperative and telescopic contractility. Nature Communications. 7 (1), 12615 (2016).
- Stam, S. et al. Filament rigidity and connectivity tune the deformation modes of active biopolymer networks. Proceedings of the National Academy of Sciences. 114 (47), E10037-E10045 (2017).
- Yadav, V. et al. Filament nucleation tunes mechanical memory in active polymer networks. Advanced Functional Materials. 29 (49), 1905243 (2019).
- Ennomani, H. et al. Architecture and connectivity govern actin network contractility. Current Biology. 26 (5), 616-626 (2016).
- Alvarado, J., Sheinman, M., Sharma, A., MacKintosh, F. C., Koenderink, G. H. Molecular motors robustly drive active gels to a critically connected state. Nature Physics. 9 (9), 591-597 (2013).
- Alvarado, J., Cipelletti, L., Koenderink, G. H. Uncovering the dynamic precursors to motor-driven contraction of active gels. Soft Matter. 15 (42), 8552-8565 (2019).
- Jung, W., Murrell, M. P., Kim, T. F-actin cross-linking enhances the stability of force generation in disordered actomyosin networks. Computational Particle Mechanics. 2 (4), 317-327 (2015).
- Lenz, M., Thoresen, T., Gardel, M. L., Dinner, A. R. Contractile units in disordered actomyosin bundles arise from f-actin buckling. Physical Review Letters. 108 (23), 238107 (2012).
- Memarian, F.L. et al. Active nematic order and dynamic lane formation of microtubules driven by membrane-bound diffusing motors. Proceedings of the National Academy of Sciences. 118 (52), e2117107118 (2021).
- Needleman, D., Dogic, Z. Active matter at the interface between materials science and cell biology. Nature Reviews Materials. 2 (9), 17048 (2017).
- Foster, P. J., Fürthauer, S., Shelley, M. J., Needleman, D. J. Active contraction of microtubule networks. eLife. 4, e10837 (2015).
- Thijssen, K. et al. Submersed micropatterned structures control active nematic flow, topology, and concentration. Proceedings of the National Academy of Sciences. 118 (38), e2106038118 (2021).
- Sanchez, T., Chen, D. T. N., DeCamp, S. J., Heymann, M., Dogic, Z. Spontaneous motion in hierarchically assembled active matter. Nature. 491 (7424), 431-434 (2012).
- Colen, J. et al. Machine learning active-nematic hydrodynamics. Proceedings of the National Academy of Sciences. 118 (10), e2016708118 (2021).
- Mitchell, K. A., Tan, A. J., Arteaga, J., Hirst, L. S. Fractal generation in a two-dimensional active-nematic fluid. Chaos: An Interdisciplinary Journal of Nonlinear Science. 31 (7), 073125 (2021).
- Pandolfi, R. J., Edwards, L., Johnston, D., Becich, P., Hirst, L. S. Designing highly tunable semiflexible filament networks. Physical Review E. 89 (6), 062602 (2014).
- Tan, A. J. et al. Topological chaos in active nematics. Nature Physics. 15 (10), 1033-1039 (2019).
- Roostalu, J., Rickman, J., Thomas, C., Nédélec, F., Surrey, T. Determinants of polar versus nematic organization in networks of dynamic microtubules and mitotic motors. Cell. 175 (3), 796-808.e14 (2018).
- Ndlec, F. J., Surrey, T., Maggs, A. C., Leibler, S. Self-organization of microtubules and motors. Nature. 389 (6648), 305-308 (1997).
- Sheung, J. Y. et al. Motor-driven restructuring of cytoskeleton composites leads to tunable time-varying elasticity. ACS Macro Letters. 10 (9), 1151-1158 (2021).
- McGorty, R. PyDDM v0.2.0. Zenodo. (2022).
- Achiriloaie, D. H. et al. Kinesin and myosin motors compete to drive rich multi-phase dynamics in programmable cytoskeletal composites. arXiv. (2021).
- Wulstein, D. M., Regan, K. E., Garamella, J., McGorty, R. J., Robertson-Anderson, R. M. Topology-dependent anomalous dynamics of ring and linear DNA are sensitive to cytoskeleton crosslinking. Science Advances. 5 (12), eaay5912 (2019).
- McGorty, R. Image-Correlation. at <https://github.com/rmcgorty/Image-Correlation>. (2020).
- Robertson, C. Theory and practical recommendations for autocorrelation-based image correlation spectroscopy. Journal of Biomedical Optics. 17 (8), 080801 (2012).
- McGorty, R. Differential Dynamic Microscopy - Python. at <https://github.com/rmcgorty/Differential-Dynamic-Microscopy---Python>. (2021).
- Cerbino, R., Trappe, V. Differential dynamic microscopy: probing wave vector dependent dynamics with a microscope. Physical Review Letters. 100 (18), 188102 (2008).
- Robertson-Anderson, R. M. Optical tweezers microrheology: from the basics to advanced techniques and applications. ACS Macro Letters. 7 (8), 968-975 (2018).
- Pollard, T. D. Polymerization of ADP-actin. Journal of Cell Biology. 99 (3), 769-777 (1984).
- Coué, M., Brenner, S. L., Spector, I., Korn, E. D. Inhibition of actin polymerization by latrunculin A. FEBS Letters. 213 (2), 316-318 (1987).
- Pollard, T. D. Actin and actin-binding proteins. Cold Spring Harbor Perspectives in Biology. 8 (8), a018226 (2016).
- Kumar, N. Taxol-induced polymerization of purified tubulin. Mechanism of action. Journal of Biological Chemistry. 256 (20), 10435-10441 (1981).
- Käs, J. et al. F-actin, a model polymer for semiflexible chains in dilute, semidilute, and liquid crystalline solutions. Biophysical Journal. 70 (2), 609-625 (1996).
- Viamontes, J., Narayanan, S., Sandy, A. R., Tang, J. X. Orientational order parameter of the nematic liquid crystalline phase of F -actin. Physical Review E. 73 (6), 061901 (2006).
- Hitt, A. L., Cross, A. R., Williams, R. C. Microtubule solutions display nematic liquid crystalline structure. Journal of Biological Chemistry. 265 (3), 1639-1647 (1990).
- Andexer, J. N., Richter, M. Emerging enzymes for ATP regeneration in biocatalytic processes. ChemBioChem. 16 (3), 380-386 (2015).
- Farhadi, L. et al. Actin and microtubule crosslinkers tune mobility and control co-localization in a composite cytoskeletal network. Soft Matter. 16 (31), 7191-7201 (2020).
- Falzone, T. T., Lenz, M., Kovar, D. R., Gardel, M. L. Assembly kinetics determine the architecture of α-actinin crosslinked F-actin networks. Nature Communications. 3 (1), 861 (2012).
- Thoresen, T., Lenz, M., Gardel, M. L. Reconstitution of contractile actomyosin bundles. Biophysical Journal. 100 (11), 2698-2705 (2011).
- Sanghvi-Shah, R., Weber, G. F. Intermediate filaments at the junction of mechanotransduction, migration, and development. Frontiers in Cell and Developmental Biology. 5, 81 (2017).
- Shen, Y. et al. Effects of vimentin intermediate filaments on the structure and dynamics of in vitro multicomponent interpenetrating cytoskeletal networks. Physical Review Letters. 127 (10), 108101 (2021).
- Laan, L., Roth, S., Dogterom, M. End-on microtubule-dynein interactions and pulling-based positioning of microtubule organizing centers. Cell Cycle. 11 (20), 3750-3757 (2012).
Erratum
Formal Correction: Erratum: Reconstituting and Characterizing Actin-Microtubule Composites with Tunable Motor-Driven Dynamics and Mechanics
Posted by JoVE Editors on 10/11/2022. Citeable Link.
An erratum was issued for: Reconstituting and Characterizing Actin-Microtubule Composites with Tunable Motor-Driven Dynamics and Mechanics. The Authors section was updated.
Mehrzad Sasanpour1
Daisy H. Achiriloaie1,2
Gloria Lee1
Gregor Leech1
Christopher Currie1
K. Alice Lindsay3
Jennifer L. Ross3
Ryan J. McGorty1
Rae M. Robertson-Anderson1
1Department of Physics and Biophysics, University of San Diego
2W. M. Keck Science Department, Scripps College, Pitzer College, and Claremont McKenna College
3Department of Physics, Syracuse University
to:
Mehrzad Sasanpour1
Daisy H. Achiriloaie1,2
Gloria Lee1
Gregor Leech1
Maya Hendija1
K. Alice Lindsay3
Jennifer L. Ross3
Ryan J. McGorty1
Rae M. Robertson-Anderson1
1Department of Physics and Biophysics, University of San Diego
2W. M. Keck Science Department, Scripps College, Pitzer College, and Claremont McKenna College
3Department of Physics, Syracuse University
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved