
Method Article
产妇基因产品的官能化操作使用
摘要
对于用于产妇的基因产物的操纵斑马鱼的卵母细胞的体外成熟的优化协议这里提出。
摘要
期间动物胚胎发育的早期阶段发生的细胞事件被存入发展卵母细胞来自母体的基因产物驱动。因为这些活动依赖于通常受精,鸡蛋里预先存在后,很快采取行动产妇产品,表达和功能降低涉及试剂注入受精卵标准方法通常是无效的。取而代之的是,这样的操作必须卵子发生过程之前或母体产物的积累期间执行。本文详细描述了斑马鱼未成熟卵母细胞的体外成熟和它们随后的体外受精的协议,产生一个存活至成年可行的胚胎。此方法允许卵子发生过程母体产品,如产品的表型救援和标记构建体的可视化,所述表达的功能操作以及一个S基因功能的通过反向遗传学剂的减少。
引言
在动物发育,母亲存款基因产物( 例如,RNA的蛋白质和其他生物分子)倒入蛋液;这些产品是立即施肥1次,2以下一些早期的细胞过程重要。使用试剂的注入的标准方法进受精卵3时母体产物的表达和功能的操作通常是无效的。这是因为大多数的RNA和蛋白质是由卵母细胞卵子发生过程中产生的,所以预加载产妇产品已经存在于成熟的卵子。这种预先存在的产品是不透与基因靶向剂,如吗啉代反义寡聚物(MOS)功能敲除,因为MO的靶mRNA的,而不是预先存在的蛋白质已经存在于受精卵。此外,许多早期胚胎的过程恰巧被influen受精后太早通过从注入受精卵RNA衍生的蛋白产物土木工程署,因为RNA可能无法产生足够快速地影响胚胎发生的第一事件。出于同样的原因,通过标记的mRNA表达注射入受精卵可以不在时间用于可视化在早期胚胎的积极作用期间产生蛋白质融合。注入之前,卵子活化挤出成熟的卵母细胞是可能的,但与类似的技术问题有关:这种成熟的卵子已经预装了母体蛋白质,它们不会成为翻译活性(即产生外源转录产物的蛋白质),直到鸡蛋后激活。由于这些原因,在早期胚胎发育作用产妇的基因产物的操作通常需要在成熟的卵母细胞卵子发生期间进行。
作为一个方法克服这些障碍, 在体外成熟的方法,其允许OOC的成熟ytes从阶段IV到卵形成,已经建立了在斑马鱼。早期的方法允许在体外成熟,但所得到的成熟卵均不能胜任受精4。随后,从中性至pH9.0的培养条件的操纵, 在体外成熟7,8后模仿鱼类5,6发现卵巢流体的碱性pH,允许可靠体外受精 (IVF)。实际上, 在体外卵母细胞-matured可产生存活到成年并且是可育3,8可行胚胎。这种改进的方法已经被进一步适配成包括母体基因的功能操作,标记的蛋白的斑马鱼卵子发生过程中通过外源表达的表达,和MO-介导的功能中垫击倒uring阶段IV的卵母细胞3( 图1)。
斑马鱼卵子发生显示出数特性阶段导致成熟的卵母细胞9(参照表1的快速指南的各个阶段中卵子发生)的形成的。简而言之,卵母细胞发育是通过I期的卵母细胞开始,并在减数分裂前期我的双线期被逮捕。这些卵母细胞经历通过活性转录增长,皮质肺泡的形成(也称为皮质颗粒,阶段II期间启动)(阶段IA和IB期间启动),和卵黄(阶段III期间启动)。卵母细胞生长阶段IV,当减数分裂I恢复,导致卵母细胞核的拆卸,被称为胚泡(GV)期间完成。随后,减数分裂在中期II再次被捕。卵母细胞的生长和减数分裂停滞的阶段IV期间完成导致成熟V期例如克9。卵泡膜的去除释放卵母细胞的进入子房10的内腔中发生并且是用于施肥和适当的蛋活化是必需的。一旦鸡蛋是从自然交配期间母亲挤压,它们被激活。在斑马鱼中,暴露于水是足够的全蛋激活,而不管精子11的存在。
在体外条件目前允许的卵母细胞从早期成熟IV-其可以通过它们的尺寸(690-730微米),大GV的在不对称的位置的存在,以及一个完全不透明的外观是由于积累被识别卵黄蛋白( 图2B)-to成熟V期卵,其特征在于一个完全拆卸GV和一个半透明的外观由于蛋黄蛋白处理12( 图2C)。在这种方法中,整个卵巢续在不同的发展阶段癌宁的卵母细胞是从女性删除。卵母细胞被允许在17α-20β二羟基-4- pregnen -3-酮(DHP),一种有效的成熟诱导激素8来开发。在此期间,卵母细胞成熟可以通过注射表达的( 例如,封端的, 在体外的mRNA -transcribed)或反向遗传学剂( 例如,MOS)来操纵。卵泡层不会自发地流下向熟化期结束,所以它必须被手动移除。 defolliculation后, 体外受精是通过卵的暴露于水(存在于胚胎(E3)介质)13和精子溶液触发蛋激活来实现的。得到的受精卵进行胚胎发育和允许的治疗操纵产妇基因功能的能力评价,为可视化和标记产妇产品分析( 图3)。
研究方案
所有斑马鱼严格按照良好的动物实践中处理的,由有关国家和/或当地的动物福利机构的定义,和所有的动物工作被批准通过适当委员会(威斯康星 - 麦迪逊保证数A3368-01大学)。动物在26.5℃的标准条件下维持。
女性的1.预选
注:卵母细胞在成年女性通常跨越范围发育阶段,来自阶段I至V( 图2)。通过成功自然交配吹扫预先存在的卵母细胞的女性增加卵母分段的同步,作为新开发的卵母细胞显示为第3组,14。内约8天后吹扫,最卵母细胞在早期阶段IV,其最适合在体外成熟的开始。这样的同步提高卵母细胞THA的产率t可经过体外成熟的全部过程中,便于实验操作。参见在成对交配和鱼类水组合物(在此设定为鱼细节先前的说明13,14 G海洋盐和150g的NaHCO 3每1000升反渗透水,pH值6.5〜8.5(优选的范围:6.8 -7.5)和180-360微秒的导电性)。参见Brand 等。 13,用于附加的配方。
- 8至10天前在体外培养操作,使用鱼网到期望的应变的单个雄性和单个母鱼转移到配合槽。对多套。让他们在交配缸过夜。让他们在晚上和通过第二天下午配合。
- 使用渔网到交配期间产生卵子,并将它们放到一个单独的油箱女性分开。每天两次与食物混合饲料,这些女性containi纳克盐水虾和鱼食品薄片的大致等量。食物的量应足以提供约20分钟,但不多,喂养时间。
2.成熟培养基的制备
注:在体外成熟实验当天准备成熟培养基,从阴除去卵母细胞的1个小时内(请参阅第3节)。
- 加入20毫升L-谷氨酰胺,pH 7.0的Leibovitz的L-15培养基中,至50mL锥形管中在无菌条件下。它用10N NaOH使pH为9.0。
- 添加9mL的Leibovitz的L-15培养基,pH为9.0,以2个独立的50mL锥形管中。标签1管"+ DHP",另一个"-DHP"。
- 在+ DHP管中,加入17α-20β二羟基-4- pregnen -3-酮的10μL(DHP),DH 2 O的490μL,和500μL10%牛血清白蛋白的(BSA)。
- 在-DHP管中,加入10毫克/毫升庆大霉素,400 100μL81,L卫生署2 O和10%BSA的500μL。
3.卵母细胞的解剖和体外培养开始
注: 在体外培养实验与预先选定的女性后8-10天,他们通过自然交配释放卵子进行。使用由当日成熟培养液中(见第2节)。如果夹层被开始附近的鱼昼夜循环(在实验室中通过预先设定的人工照明的鱼设施确定)13的端部,有可能通过模仿自然交配发生的过程的卵母细胞成熟的适当。这意味着,在该协议最步骤必须在晚上进行,如果鱼被容纳在用标准光循环设施( 例如,开始步骤3.2在下午6点在设施与上午08时至10时的光周期),尽管其它工作时间段是可能的与适当的时间移光周期。
- 制备用蒸馏水2 O 0.2%三卡因储备溶液,缓冲至pH 7.0用1M Tris,pH 9.0的,并保持在4℃下该溶液中。这可以提前准备。
- 发起之前在设施的日常光周期结束实验0-4小时。在250-mL烧杯中,加入20毫升的0.2%三卡因储备溶液,pH为7.0,以80毫升水鱼的并混合。
- 转移预先选定的女性,以三卡因溶液,并通过曝光过度安乐死他们。离开女性在鳃运动停止后15分钟的三卡因溶液。
- 用勺子,收集来自三卡因溶液安乐死鱼和鱼水简单冲洗一下。将鱼在纸巾吸去多余水分。
- 使用干净的刀片在胸鳍的水平斩首安乐死鱼。使用解剖剪刀,使在鱼的腹侧形成纵向切口,从其前端到肛门区延伸。
- 将鱼上入射光在解剖显微镜下的培养皿。使用一对解剖钳,转移卵巢部分以含有4 Leibovitz的L-15培养基+ DHP的mL的35×10mm的培养皿中。
注:卵巢,含有显影的卵母细胞,将显示为内部体腔内的不透明和块状结构。 - 使用解剖钳,轻轻地从滤泡群众游离于卵母细胞。排序IV早期阶段的卵母细胞( 图2B;之前GV击穿,在接近最大尺寸和特征是暗的,不透明的细胞质和一个显而易见的GV位于不对称卵母细胞内)。丢弃在早期阶段( 图2A)和半透明的,成熟V期蛋(类似于在图2C,但存在于前DHP治疗卵巢)卵母细胞。
注:卵母细胞被选择用于熟化在早期阶段IV,其导致成熟,V期oocyt的最早阶段后在体外培养条件下( 图2C和D)ES 3。因为阶段的卵母细胞IV正在积极生产产品,操纵在该阶段允许外源产物的RNA通过注射或用于经由引入的MO基因功能的降低的表达。 - 使用玻璃巴斯德吸管向初期IV的卵母细胞转移到含有4 Leibovitz的L-15培养基+ DHP毫升的第二35×10mm的塑料培养皿中。传递最小量-DHP介质的含有+ DHP溶液的培养皿中。
4.卵母细胞显微注射
注:隔离阶段的卵母细胞IV 体外成熟经历可显微注射引入用于功能操作或标记的蛋白质的表达的试剂。试剂,例如mRNA和MOS(见4)的显微注射,典型地进行时,卵母细胞正在经历体外 matu配给在+ DHP介质,以defolliculation之前。如果需要的话,为了允许更多的时间来进行前卵母细胞成熟的操作,注射也可以在-DHP介质中进行,成熟以前,至少2小时。他们随后被转移到+ DHP中。
- 制备使用标准表达试剂盒的mRNA。 (建议为200pg /μL)在RNA级水保持它们在-80℃下为100-500 pg /μL的库存用于注射在50-500 pg /μL的终浓度。根据制造商的说明在水中的4纳克/μL的浓度制备的MO。在2毫微克的终浓度它们注入/μL(或根据经验确定)。
注:注射通常在+ DHP介质之前defolliculation进行(见第5的说明)。 - 如果注入的mRNA,前立即注射,使用RNA级反渗透水和0.2M KCl的实现的0.1M的KCl的最终溶液稀释的mRNA。
- 如果注入的MO,之前立即注射,使用反渗透水和0.2M KCl的实现的0.1M的KCl的最终溶液稀释所述MOS至所需浓度。
- 手动保持用细镊子卵母细胞和注入约1纳升成使用与拉玻璃毛细吸管制成的针野生型IV期的卵母细胞。
- 通过拉动加热玻璃毛细管,加载溶液被注入,并用钳子打破针尖,如在协议标准注射到斑马鱼早期胚胎15先前所描述的制备的玻璃针。
注:这是有益的,如果针头有一个渐进的锥度,而不是突然的一个;这允许更大的灵活性在针尖断裂的位置而言,也有助于防止注射胚胎的破坏。使用相同的针和溶液作为胚胎注射,调整通过确定预的注射体积产生期望的体积微量压力设置。这些设置可由下式确定模拟注入矿物油的在显微镜阶段校准载玻片(0.01毫米),调整设置微量以获得注射溶液的丸所需直径的下降,如前所述15。
- 通过拉动加热玻璃毛细管,加载溶液被注入,并用钳子打破针尖,如在协议标准注射到斑马鱼早期胚胎15先前所描述的制备的玻璃针。
5.卵母细胞成熟和Defolliculation
注:在卵母细胞成熟,卵母细胞会逐渐变得透明,从而允许对成功培养条件的评估。
- 继续温育未成熟(和,如果合适的话,注入)在+ DHP介质的卵母细胞在26.5℃下,周期性地检查(每隔30分钟),以确保所述的卵母细胞保持完整,并通过逐渐变得半透明正在进行适当的成熟(参见图2C) 。
- 用巴斯德吸管取出所有裂解卵母细胞和丢弃在实验室废物烧杯中保持介质的质量。交换培养基用新鲜+ DHP介质的约一半体积,以保持透明溶液,这取决于卵母细胞裂解的量。
- 允许在体外成熟进行,直到大部分的卵母细胞成为半透明的,并具有一个GV即不再明显(在DHP治疗约2小时,参见图2C相比至D为)。查看它们与透射光光学解剖显微镜下。
- 从每个成熟卵母细胞中除去最外面的卵泡膜。使用特细镊子使卵泡膜撕裂的区域与卵母细胞和膜之间增加的空间。剥离膜的一部分,并滚动卵母细胞从膜的同时按住剥离部分。
注意:下面的绒毛膜将通常保持与在该过程中蛋密切关联;之后蛋行为ivation,其膨胀以形成用于胚胎的保护层。 - 转移defolliculated卵母细胞在介质的最小体积(典型地,在小于20μL转移约5-10成熟的卵母细胞)与培养(+ DHP)介质几滴到培养皿中,并进行到受精。
6. 体外培养卵母细胞的受精
注:在冰上精子溶液,制备以下,将保持其效力为约2小时。
- 制备接近卵母细胞成熟步骤的结束和前一个精子溶液使用睾丸从5名男defolliculation在Hanks'溶液的500微升,如先前16,17说明。
- 添加10-50微升精解到+ DHP培养基defolliculated卵母细胞。等待10秒。
- 使用移液管,加入胚胎(E3)中的几滴卵母细胞。等待1分钟,然后淹没PL吃了E3中。
注:E3培养基的组成如下:5mM的NaCl的,0.17毫米氯化钾,0.33毫米氯化钙2,0.33毫摩尔MgSO 4干燥,和1-5%亚甲蓝13。 - 重复步骤5.3,6.2和6.3获得受精胚胎的数量更多。
- 允许受精的胚胎发展。使用具有透过光的光学解剖显微镜通过裂解阶段,观察进展以确保成功受精,如先前对受精17和分段18描述。
结果
要确定是否上述过程是成功的,可以在切割阶段观察,确认的定型早期胚胎裂解图案18的外观的胚胎,以及在24小时后受精(HPF),以确认正确的发展基本体计划。该方法允许产妇产品的功能性研究,通过试剂,如的mRNA和MO的注射操作,卵母细胞发育过程中。
产妇产品功能研究的操作:
对野生型和突变的产物的mRNA编码可被注入到在体外成熟的卵母细胞减数分裂和早期胚胎阶段期间测试母体基因功能的操作的效果。例如,野生型胚与无线网络被注入LD-型的mRNA来测试产品的过表达的影响,或者与突变体RNA来测试潜在优势( 例如,获得性功能和antimorphic)在这些过程中的效果。在体外培养的卵母细胞都能够从外源基因产生的蛋白质在整个卵母细胞发育,如由GFP从所注射的mRNA的表达,虽然仅在卵子发生IV阶段开始培养条件的卵母细胞可以发育成成熟V期的卵母细胞( 图2B -2D,也参见Nair 等3)。野生型产物的通过注入的mRNA的表达也是工具将母体效应突变的救援定位克隆3,24期间确认基因的身份。在这种情况下,野生型基因被注入从纯合突变女性的卵母细胞测试野生型的产品是否能挽救突变表型。这种遗传拯救在示出<强>图3A-3C,其中注入AurB的mRNA被示为营救在其相应的基因, 细胞岛 (CEI)( 图3A-3C)的突变的表型效应。来自对照组的胚胎也允许开发,以显示相应的突变表型3。突变体RNA也可以被注射入卵母细胞的突变体,以测试由救援的程度进行比较来引起野生型产物突变产物是否保持部分功能。
从基因注射到卵母细胞开发蛋白质表达似乎很少或没有延迟发生。强GFP表达相应的mRNA的注射后2小时内观察到的,而不管卵母细胞3的发育阶段( 图2B-2D;也参见Nair 等3)。产品如mCherry的:SAS6和Birc5b(特利):GFP可以立即受精后和第一胚胎细胞周期3,25中观察到。 mRNA的卵子发生过程中的注射还导致产生蛋白质的,无论稠合标记部分的,是功能性的受精后立即,如图母体产品用于蜂窝岛/ aurB 3, 无效循环/ LRMP 24的情况下,和杂色/ bir5b 25。翻译阻断的MO还有一个效果受精后立即,如图用于无效循环/ Lrmp的3并在胚胎发生的后期阶段,如在不可能完成的任务/ dhx16 3的情况下。剪接阻断的MO,在卵母细胞成熟注射时,可能不会对母体功能的效果,可能是由于已经存在的成熟产妇在阶段IV的卵母细胞的转录物 3。
morphants的产生:
当使用对应于基因与已鉴定的突变的MO,所述表型morphant预计模拟突变体的表型。 Morphants进行比较,以未注射的胚胎和那些与标准注射,控制MO。翻译阻断吗啉代的喷射可以成功表型模拟已知突变表型3,如图所示由注射Lrmp的MO的卵母细胞成,它模仿相应突变的突变表型, 无效循环 (图3D-F)。 MO的特异性可以通过注入的MO成胚胎(以前的综述)19,20时所使用的相同的方法来确定。
荧光标记的融合蛋白的表达:
mRNA编码对的融合的荧光蛋白质( 例如,GFP和mCherry的)感兴趣的基因产物还可以注入到野生型或突变体的卵母细胞的早期胚胎中可视化的相应的产品( 即,反映了亚细胞定位图案; 图3G和3H)。的mRNA编码类似的融合,但涉及的突变等位基因可以类似地表示,以测试突变是否影响任何潜在的亚细胞定位。

图1: 体外卵母细胞成熟。示意图示出了涉及在体外卵母细胞成熟的各种步骤。成年女性的预选产蛋的过程之前8天,清除那些已经成熟,并有利于鸡蛋的他们微尘新的卵母细胞发育。卵巢,含有显影的卵母细胞,从女性中移出并转移到含有激素的DHP来诱导卵母细胞成熟的体外的介质。从女性和之前立即或卵母细胞成熟过程中取出后,卵母细胞可与RNA产物和其它试剂被注射的母体因素的功能的操作。成熟的卵母细胞被手动defolliculated和体外受精与精子溶液以产生受精卵,胚胎存活。 请点击此处查看该图的放大版本。
{kind=link}

图2: 在卵母细胞体外成熟和注射基因产物的表达。 (A)的卵母细胞在从卵巢中观察到的阶段I-III,女4天后清洗(DPP)。 (B)的卵母细胞在阶段IV,从女性8 DPP在卵巢观察。生发囊泡(GV,箭头)是清晰可见的,并占据的偏心位置。阶段III的卵母细胞中(A)还显示出显而易见的GV,但它是在卵母细胞发现居中。在(B)第四阶段的卵母细胞具有的尺寸是接近最大相比,在成熟的卵母细胞(C)的。 (C和D)的卵母细胞在阶段V(成熟的卵母细胞)后,在阶段IV开始,如在(B)中,用GFP-编码成熟期间注入体外成熟条件2小时, 在体外转录的mRNA(C,可见光仅; D,可见光和GFP荧光)的叠加。的GV是在第五阶段的卵母细胞,这也比阶段的卵母细胞IV较少不透明不再明显。注入卵母细胞表达GFP蛋白(D)。成熟阶段的卵母细胞IV的Defolliculation为described在协议部分。比例尺=300μm左右。所有的面板进行了转载,经许可后,从Nair 等 3。 请点击此处查看该图的放大版本。
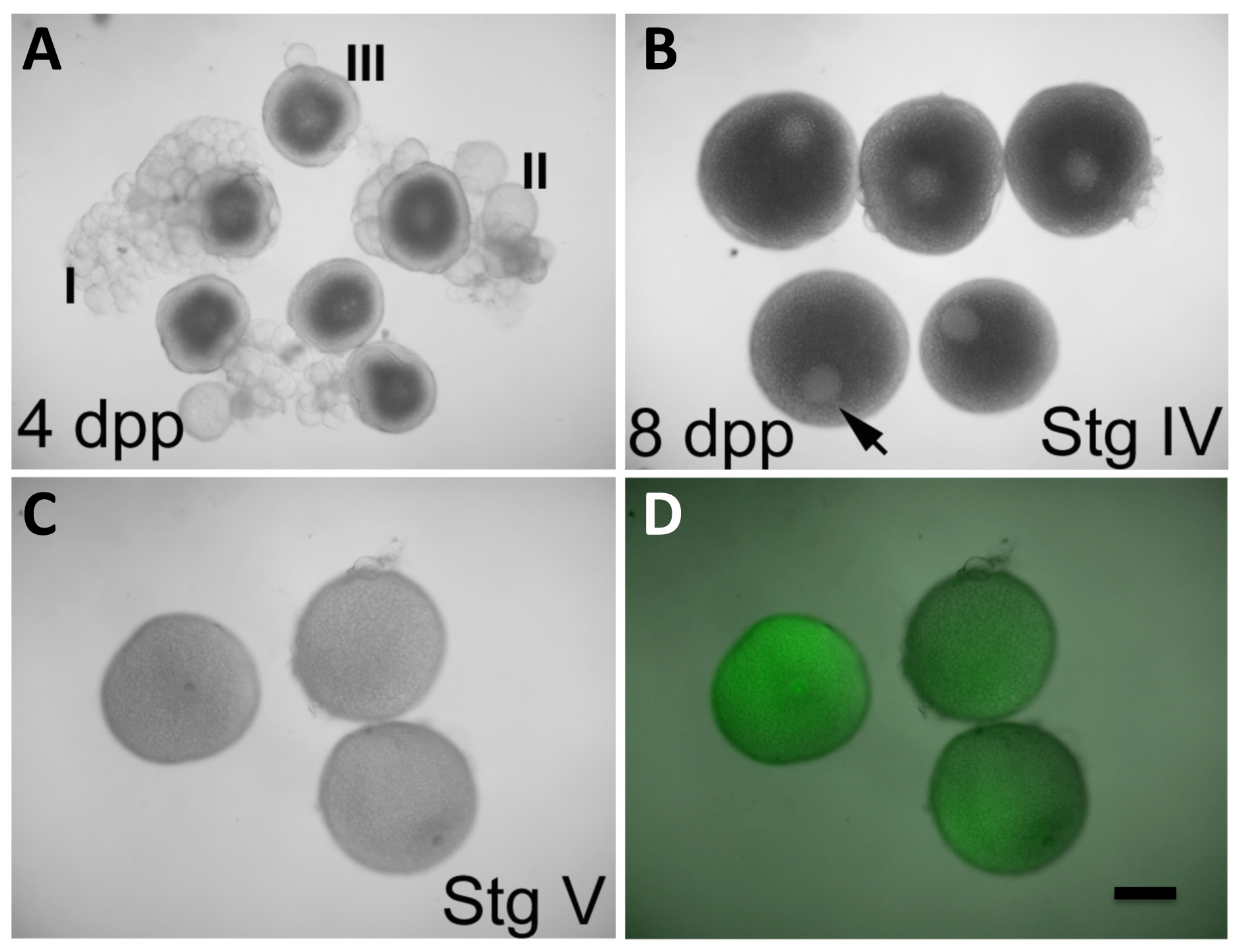{kind=link}

图3:操作和母婴产品的可视化,通过体外卵母细胞成熟。 (A - C)引起的通过野生型aurB mRNA的注射在蜂窝岛/极光B中的突变导入阶段IV CEI / aurB卵母细胞的母性影响表型的拯救。示出动物的观点65个MPF固定blastodiscs的,具有抗β-catenin的抗体可视化,以突出显示膜(绿色)合并图像,抗α微管蛋白抗体来指示微管(红色),DAPI指定DNA(蓝色)。 (A)β联蛋白积聚在野生型胚胎中,表示正常沟成熟。 (B)来自未注射的CEI / aurB阶段IV卵母甲CEI / aurB胚胎显示,不积累β连环蛋白的部分,基本的沟。从野生型的mRNA aurB示出了在沟槽健壮β连环蛋白累积喷射的CEI / aurB卵母细胞(C)一种救出CEI / aurB胚胎。 (D - F)的无效循环/ LRMP引起的突变从Lrmp的注射吗啉代入阶段的卵母细胞IV母体效应拟表型。示出的70个MPF固定blastodiscs,具有抗γ微管蛋白抗体可视化动物视图合并图像以指示中心体(红色)和DAPI来指定DNA(蓝色)。 (D)在野生型胚胎中,每个核同伙与γ微管蛋白,对于中心体材料的标记。 (E)在母体效应FUE突变体,原核聚变发生故障,从而导致对应于未融合亲代的亲核和极体为减数分裂II DNA标签的两到三个贴片。 (F)在Lrmp的morphants,其中母体Lrmp的功能被抑制,细胞核同样失败来划分,此外,不能与γ微管蛋白相关联。 (G和H)在早期胚胎母体生产的产品的可视化。从外源性的mRNA Sass6-mCherry的蛋白质注射到IV期的卵母细胞定位于中心粒。示出动物的观点的40个MPF固定blastodiscs,具有抗γ微管蛋白抗体可视化,以突出的中心体(绿色),的mCherry荧光合并图像以指示Sass6(红色),和DAPI指定DNA(蓝色)。 (G)表达Sass6-mCherry的蛋白定位癌灶由中心体标记γ微管蛋白的位点处的侧翼细胞核(箭头)进行标记。 (H )相同的图像,仅示出Sass6-mCherry的和DAPI为清楚起见。在(G和H)中的胚胎是固定的,但表示的产品,如融合的mCherry的荧光,也可以在活胚胎中观察到(未示出)。在AF比例尺=100μm,并且10μm的G和H所有板再现时,经许可,从Nair 等 3。 请点击此处查看该图的放大版本。
{kind=link}
| 卵子发生阶段 | 卵母细胞直径(μm) | 重点标志(S) |
| 1A - Prefollicle | 为7〜20 | 积极转录起始,核仁的积累 |
| 1B -卵泡 | 20至140 | 染色体的解聚 |
| II -皮质肺泡 | 140至340 | 皮质肺泡生产 |
| III -卵黄 | 340至690 | 卵黄前体蛋白和脂质的积累卵胞质变暗 |
| 早IV -卵母细胞成熟 | 690〜730(较低范围) | 胚泡的不对称本地化 |
| 早IV -卵母细胞成熟 | 690〜730(较高范围) | 胚泡消失,阻滞在中期II |
| N -成熟卵母细胞 | 〜750 | 卵质/蛋黄变成半透明 |
表1:斑马卵母细胞发育的地标鱼。
讨论
上述协议是用于施肥之前的基因产物的操作,因此允许产妇基因产物在早期斑马鱼胚胎的研究。以前的研究已经能够在体外 4成熟卵母细胞;该协议被修改,以便允许成熟卵母细胞8的体外随后施肥。这又允许用于试剂在早期胚胎3的母系遗传的产品功能性操作和可视化的注入。从这种方法产生的胚胎可以存活,能够承受成为肥沃的成年人。在初步实验中,大约一半来自该程序导出的受精胚胎的是对发展的第5天活,如通过鱼鳔通货膨胀在那个阶段,其中大约一半成为健康,肥沃的成年人(未发表的观察结果)进行评估。
有一些的关键步骤,以在协议考虑。在适当的卵母细胞阶段启动成熟后(初期IV)可以富集如果女性最近已经配合时,但8之前不DPP。使用最近没有交配的鱼可能导致变质的鸡蛋14,这不会发生的成熟。内小于8天交配的雌性将具有多数蛋在III期或更小,并且因此,卵将不能正常在体外成熟。在女性该现有8天配合卵巢将卵母细胞在早期阶段IV的最佳分数。这些可以通过它们的不透明度和GV的在偏心位置( 图2B)的存在被识别。卵母细胞阶段判定也可以通过尺寸测量的帮助下,与卵母细胞在显微镜下放置在刻度微米滑动,并与标准分级准则( 表1)。然而,早期的卵母细胞IV具有接近最大尺寸相比成熟卵母细胞(由它们的相对透明性和缺乏GV的认可; 图2C),并且很容易认识到的,如上述那样,其通常消除了直接的卵母细胞尺寸测量的需要。
在体外成熟必须几个小时到阴卵母细胞供体驯化日光周期结束的内开始。卵母细胞不能正常成熟,体现在贫困受精率,如果在光周期上午培养一段时间。其原因在体外卵母细胞成熟方法的昼夜依赖性是不理解但很可能反映了在体内卵子成熟底层昼夜偏压卵子发生21在涉及循环基因的表达。卵母细胞未能成功进行体外成熟,尽管适当的卵母细胞分期的一个常见原因是,DHP已过期。 DHP激素一年之后一般到期,并保证效果香港专业教育学院的成熟,工作批量应9个月使用内更换。
卵母细胞的注射是另一个关键步骤当该方法的目的是母体产品的功能操作。这个过程具有不同于在受精卵注射标准在0-30强积金不同的要求。在卵母细胞注射的关键因素是条件不导致过早的活化的蛋,如在Leibovitz的L-15介质22,23下进行注射的需要。因为它们嵌入在卵巢,卵母细胞成熟也对注射特殊的挑战。上述协议意味着第一从卵巢的卵母细胞解离,然后保持每个镊子分别分离卵母细胞,同时注入。这种技术已在最少的处理的适当阶段运作良好,并允许预分选的卵母细胞。替代方法也可以使用,例如为:i)将卵母细胞成标准注射琼脂糖槽15,其中,槽支撑注射过程中卵母细胞的垂直壁(在此替代方法中,卵母细胞需要转移到喷射板而保持在体外成熟培养基)和ii )使该卵母细胞保持附着到卵巢肿块,其可与钳子在注射期间被保持,以避免与注入的卵母细胞接触(在此方法中,卵母细胞应注射到两个后解离促进DHP曝光,并允许其defolliculation ,本身IVF必要的)。尽管这个特定的挑战,IV期的卵母细胞可容易地与注射针穿透,可能是因为在此阶段绒毛膜没有发育完全9。
当前成熟协议的一个限制是, 在体外成熟条件促进适当的卵母细胞发育当卵母细胞成熟是在比第四阶段早期阶段开始无法创建。当它们到达的直径为520微米,这阶段III过程中发生(卵黄,对应于340到690微米的范围)的卵母细胞成为感受态,以通过经历成熟到DHP响应9。然而,III期卵母细胞的结果在那些不能胜任受精的卵母细胞3的培养。只有卵母细胞,其体外培养在阶段IV( 图2B)开始可以发育成成熟,V期的卵母细胞( 图2C和D)。这可能与这样的事实,第三阶段是卵黄和卵母细胞增长9至关重要。因此,本文中给出的在体外卵母细胞成熟过程应该只具有阶段的初始种群IV的卵母细胞(B),而不是与在早期阶段的卵母细胞(阶段我用- III; FIGURê 2A)。出于同样的原因,此过程被限制到在阶段IV或更高表达的基因。基因,它们的产品是较早表达的官能化操作可能而不得不依赖于遗传方法(见下文)。改进体外培养成熟法可能在将来允许卵母细胞培养在早期阶段的启动,从而扩大了在体外成熟方法的能力。
在所提出的方法的另一个限制是defolliculation,这对于涉及受精后续步骤必不可少的,是目前在过程的限速步骤。卵泡膜是围绕显影的卵母细胞的透明层,和去除可能是乏味。如果该膜没有完全去除,鸡蛋不会成为完全激活和受精。这是由绒毛膜故障扩大和不规则地被安置,沟状ST的发展取得明显ructures(pseudocleavages),未受精的胚胎11的特性。酶促defolliculation方法如胶原酶,如在爪蟾使用的,已经尝试了。然而,在我们的手中,这种技术一直没有成功,因此,我们依靠手工defolliculation。因为手工defolliculation是劳动密集且耗时的,所以难以在体外卵母细胞成熟后获得的受精卵的大批量。由于人工defolliculation造成的时间上的限制,我们目前正处在一个时间施肥几defolliculated,成熟的卵子,在5-10鸡蛋小批量。酶defolliculation与此过程中的合并可能会显著提高其产量和整体易用性。
虽然在体外 -matured卵母细胞受精后产生的胚胎可以成为可行的成年人,我们注意到, 在体外受精后,胚胎的一小部分做不正常或及时划分。我们目前正在选择用于其传播包括轮体外卵母细胞成熟的,这可能导致在通过这种方法得到的胚胎发育的更稳健的图案线条。
该过程已经允许清晰抢救或蛋白质定位的可视化对于许多突变24,25。但是,对于某些基因产物表达的调控可能是至关重要的,因此通过注射基因表达的产品可能会导致不同的结果26。同样重要的是要注意到所有的控件( 例如,与具有类似的性质的那些在实验中还没有被预测用于不产生作用仅编码惰性蛋白质试剂,例如控制用MOS或RNA的注射的胚胎,例如的如GFP),作为潜在的变化可能会导致错误的结论。
NT">涉及体外操作,然后受精的一种方法是在母系沉积产品,其中注入RNA,MOS,或蛋白进入受精卵可能不能有效地降低产品的标准方法在蛋已经累积的情况下是特别有用的。其它最近开发的方法,例如CRISPR突变体的产生,也处于产生用于母本表达的基因26突变体的条件有效的。然而,这种方法需要的鱼的几代的生长。 在体外卵母细胞成熟技术允许快速功能操纵研究的母系表达的基因,包括抢救和母体效应的突变表型模拟,以及RNA和蛋白质中的早期胚胎表达的功能。披露声明
作者有没有竞争经济利益。
致谢
我们要感谢的Pelegri实验室和鱼设施的管理人员,谁在推动这个项目器乐的成员。本项目支持由美国国立卫生研究院R56 GM065303和美国国立卫生研究院2 T32 GM007133到ELW,NIH 5 F31 GM108449-02和NIH 2 T32 GM007133到CCE,和NIH RO1 GM065303到FP提供
材料
| Name | Company | Catalog Number | Comments |
| Zebrafish mating boxes | Aqua Schwarz | SpawningBox1 | |
| NaCl | Sigma | S5886 | |
| KCl | Sigma | P5405 | |
| Na2HPO4 | Sigma | S3264 | |
| KH2PO4 | Sigma | P9791 | |
| CaCl2 | Sigma | C7902 | |
| MgSO2-7H2O | Sigma | 63138 | |
| NaHCO2 | Sigma | S5761 | |
| Tricaine | Western Chemical | Tricaine-D (MS 222) | FDA approved (ANADA 200-226) |
| Tris base | Sigma | 77-86-1 | to prepare 1 M Tris pH 9.0 |
| HCl | Sigma | 920-1 | to prepare 1 M Tris pH 9.0 |
| Fish net (fine mesh) (4 - 5 in) | PennPlax | (ThatFishThatPlace # 212370) | available in ThatFishThatPlace |
| Plastic spoon | available in most standard stores | ||
| Dissecting scissors | Fine Science Tools | 14091-09 | |
| Dissecting forceps | Dumont | SS | available from Fine Science Tools |
| Dissecting stereoscope (with transmitted light source) | Nikon | SMZ645 | or equivalent |
| Reflective light source (LED arms) | Fostec | KL1600 LED | or equivalent |
| Petri plates 10 cm diameter | any maker | ||
| Eppendorf tubes 1.5 mL | any maker | ||
| Ice bucket | any maker | ||
| Narrow spatula | Fisher | 14-374 | |
| Depression glass plate | Corning Inc | 722085 (Fisher cat. No 13-748B) | available from Fisher Scientific |
| Paper towels | any maker | ||
| Kimwipes | Kimberly-Clark | 06-666-11 | available from Fisher Scientific |
| Timer stop watch | any maker | ||
| Wash bottle | Thermo Scientific | 24020500 | available from Fisher Scientific |
| beakers, 250 mL (2) | Corning Inc. | 1000250 | available from Fisher Scientific |
| Leibovitz'z L-15 medium | Thermofisher | 11415064 | |
| NaOH | Sigma | 221465 | for pH'ing |
| BSA | Sigma | A2058 | |
| 17alph-20beta-dihyroxy-4-pregnen-3-one (DHP) | Sigma | P6285 | |
| gentamicin | Sigma | G1272 | |
| Injection Apparatus | Eppendorf | FemtoJet | or equivalent |
| Capillary Tubing for injection needles | FHC | 30-30-1 | or equivalent, Borosil 1.0 mm OD x 0.5 mm ID with fiber, 100 mm |
| Needle puller | Sutter Instruments | Model P-87 | any maker |
| Micropipetor (1 - 20 µL range) with tips | any maker | ||
| Micropipetor (20 - 200 µL range) with tips | any maker | ||
| Micropipetor (100 - 1000 µL range) with tips | any maker | ||
| Conical tubes 15 mL | any maker | ||
| Conical tubes 50 mL | any maker | ||
| plastic pipette 10 mL with bulb | any maker | ||
| plastic pipette 20 mL with bulb | any maker | ||
| Microscope stage Calibration Slide 0.01 mm | AmScope | MR095 | or equivalent |
| Reagents for fish water: | |||
| Instant Ocean Salt | Drs. Foster & Smith | CD-116528 | |
| Sodium bicarbonate (cell culture tested) | Sigma | S5761-1KG | |
| Reagents for E3 medium: | |||
| NaCl | Sigma | S5886-1KG | |
| KCl | Sigma | P5405-500G | |
| CaCl2, dihydrate | Sigma | C7902-500G | |
| MgSO4, heptahydrate | Sigma | 63138-250G | |
| Methylene Blue | Sigma | M9140-25G | |
| Fish Food: | |||
| Frozen brine shrimp | Brine Shrimp Direct | FBSFKG50 | |
| Tetramin Flakes | Drs. Foster & Smith | 16623 |
参考文献
- Lindeman, R., Pelegri, F. Vertebrate maternal-effect genes: insights into fertilization, early cleavage divisions, and germ cell determinant localization from studies in the zebrafish. Mol Rep Dev. 77 (4), 299-313 (2010).
- Abrams, E. W., Mullins, M. C. Early zebrafish development: it's in the maternal genes. Curr Opin Genet Dev. 19 (4), 396-403 (2009).
- Nair, S., Lindeman, R. E., Pelegri, F. In vitro oocyte culture-based manipulation of zebrafish maternal genes. Dev Dyn. 242 (1), 44-52 (2013).
- Selman, K., Petrino, T. R., Wallace, R. A. Experimental conditions for oocyte maturation in the zebrafish, Brachydanio rerio. J Exp Zool. 269 (6), 538-550 (1994).
- Fauvel, C., Omnes, M. H., Suquet, M., Normant, Y. Reliable assessment of overripening in turbot (Scophtalmus maximus) by a simple pH measurement. Aquaculture. 117 (1-2), 107-113 (1993).
- Lahnsteiner, F., Weismann, T., Patzner, R. A. Composition of the ovarian fluid in 4 salmonid speices: Onchorhynchus mykiss, Salmo trutta flacustris, Salvelinus alpinus and Husho hucho. Reprod Nutr Dev. 35 (5), 465-474 (1995).
- Patiño, R., Bolamba, D., Thomas, P., Kumakura, N. Effects of external pH on hormonally regulated ovarian follicle maturation and ovulation in Atlantic croaker. Gen Comp Endocrinol. 141 (2), 126-134 (2005).
- Seki, S., et al. Development of a reliable in vitro maturation system for zebrafish oocytes. Reproduction. 135 (3), 285-292 (2008).
- Selman, K., Wallace, R. A., Sarka, A., Qi, X. Stages of oocyte development in the zebrafish, Brachydanio rerio. J Morphol. 218 (2), 203-224 (1993).
- Clelland, E., Peng, C. Endocrine/paracrine control of zebrafish ovarian development. Mol Cell Endocrinol. 312 (1-2), 42-52 (2009).
- Kane, D. A., Kimmel, C. B. The zebrafish midblastula transition. Development. 119 (2), 447-456 (1993).
- Kanagaraj, P., et al. Souffle/Spastizin controls secretory vesicle maturation during zebrafish oogenesis. PLoS Genet. 10 (6), e1004449 (2014).
- Brand, M., Granato, M., Nüsslein-Volhard, C., Nüsslein-Volhard, C., Dahm, R. . Keeping and raising zebrafish,Zebrafish - A Practical Approach. , 7-37 (2002).
- Connoly, M. H., Dutkosky, R. M., Heah, T. P., Sayler, G. S., Henry, T. B. Temporal dynamics of oocyte growth and vitellogenin gene expression in zebrafish (Danio rerio). Zebrafish. 11 (2), 107-114 (2014).
- Rosen, J. N., Sweeney, M. F., Mably, J. D. Microinjection of zebrafish embryos to analyze gene function. J Vis Exp. (25), e1115 (2009).
- Pelegri, F., Mullins, M. Genetic screens for mutations affecting adult traits and parental-effect genes. Meth Cell Biol. 104, 83-120 (2011).
- Baars, D. L., Takle, K. A., Heier, J., Pelegri, F. Ploidy manipulation of zebrafish embryos with Heat Shock 2 treatment. J Vis Exp. , (2016).
- Kimmel, C., Ballard, W. W., Kimmel, S. R., Ullman, B., Schilling, T. F. Stages of embryonic development in the zebrafish. Dev Dyn. 203 (3), 253-310 (1995).
- Eisen, J. S., Smith, J. C. Controlling morpholino experiments: don't stop making antisense. Development. 135 (10), 1735-1743 (2008).
- Schulte-Merker, S., Stainier, D. Y. R. Out with the old, in with the new: reassessing morpholino knockdowns in light of genome editing technology. Development. 141 (16), 3103-3104 (2014).
- Dekens, M. P. S., Santoriello, C., Vallone, D., Frassi, G., Whitmore, D., Foulkes, N. S. Light regulates the cell cycle in zebrafish. Curr Biol. 13 (23), 2051-2057 (2003).
- Sakai, N., Burgess, S., Hopkins, N. Delayed in vitro fertilization of zebrafish eggs in Hank's saline containing bovine serum albumin. Mol Mar Biotechnol. 6 (2), 84-87 (1997).
- Pelegri, F., Mullins, M. C. Genetic screens for mutations affecting adult traits and parental-effect genes. Meth Cell Biol. 135, (2016).
- Lindeman, R. E., Pelegri, F. Localized products of futile cycle/lrmp promote centrosome-nucleus attachment in the zebrafish zygote. Curr Biol. 22 (10), 843-851 (2012).
- Nair, S., Marlow, F., Abrams, E., Kapp, L., Mullins, M., Pelegri, F. The chromosomal passenger protein Birc5b organizes microfilaments and germ plasm in the zebrafish embryo. PLoS Genetics. 9 (4), e1003448 (2013).
- Eno, C., Solanki, B., Pelegri, F. aura (mid1ip1l) regulates the cytoskeleton at the zebrafish egg-to-embryo transition. Development. 143 (9), 1585-1599 (2016).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。