
Method Article
Manipolazione funzionale delle materne genica Utilizzando
In questo articolo
Riepilogo
Un protocollo ottimizzato per la maturazione in vitro di oociti zebrafish utilizzati per la manipolazione di prodotti genici materni è presentato qui.
Abstract
eventi cellulari che hanno luogo durante le prime fasi dello sviluppo embrionale degli animali sono guidati da prodotti genici di origine materna depositati nel citoplasma dell'ovocita in via di sviluppo. Poiché questi eventi si affidano ai prodotti materni che tipicamente agiscono molto presto dopo la fecondazione, che preesistono all'interno dell'uovo, approcci standard per l'espressione e la riduzione funzionale coinvolgono l'iniezione di reagenti nell'uovo fecondato sono tipicamente inefficaci. Invece, tali manipolazioni devono essere eseguite durante oogenesis, prima o durante l'accumulo di prodotti materni. Questo articolo descrive in dettaglio un protocollo per la maturazione in vitro degli ovociti immaturi zebrafish e la loro successiva fecondazione in vitro, ottenendo embrioni vitali che sopravvivono alla maturità. Questo metodo consente la manipolazione funzionale dei prodotti materni durante oogenesis, come l'espressione di prodotti per recupero fenotipico e visualizzazione costrutto contrassegnati, nonché uns la riduzione della funzione del gene attraverso agenti inversa-genetica.
Introduzione
Durante lo sviluppo degli animali, i prodotti genici depositi madre (ad esempio, RNA, proteine e altre biomolecole) nell'uovo; questi prodotti sono importanti per i primi processi cellulari subito dopo la fecondazione 1, 2. La manipolazione dell'espressione e la funzione dei prodotti materni è tipicamente inefficace quando si utilizza un approccio standard per l'iniezione dei reagenti in uova fecondate 3. Questo è perché la maggior parte RNA e proteine sono prodotte dalla ovocita durante ovogenesi, così prodotti materni precaricati sono già presenti nell'uovo maturo. Tali prodotti preesistenti sono impermeabili a knockdown funzionale con agenti gene target, come oligonucleotidi antisenso morfolino (MOS), perché MO bersaglio l'mRNA, non la proteina preesistente già presente nel uovo alla fecondazione. Inoltre, molti processi embrionale precoce accadono troppo presto dopo la fecondazione da influenced da prodotti di proteine derivate da RNA iniettato in un ovulo fecondato, come l'RNA non può essere prodotto abbastanza veloce per influenzare i primi eventi di embriogenesi. Per lo stesso motivo, etichettato fusioni di proteine espresse attraverso un'iniezione mRNA nell'uovo fecondato non possono essere prodotti in tempo per la visualizzazione durante il loro ruolo attivo nella embrione precoce. Iniezione in oociti maturi estrusi prima dell'attivazione uovo è possibile, ma è associato a problemi tecnici analoghi: tali uova mature sono già pre-caricati con proteina materna, ed essi non diventerà traslazione attiva (cioè, produrre proteine da trascrizioni esogeni) fino a dopo uovo Attivazione. Per queste ragioni, la manipolazione di prodotti genici materni che agiscono in embriogenesi precoce in genere deve essere effettuata durante ovogenesi nell'ovocita maturazione.
Come uno approccio per superare questi ostacoli, in metodi di maturazione in vitro, che consentono la maturazione di oocytes da stadio IV alla formazione di uova, sono stati stabiliti in zebrafish. Tra i primi metodi consentiti maturazione in vitro, ma le uova mature risultanti non erano competenti per la fecondazione 4. Successivamente, la manipolazione delle condizioni di coltura da neutro a pH 9,0, mimando il pH alcalino del fluido ovarico trovato in specie ittiche 5, 6, consentito affidabile fertilizzazione in vitro (IVF) dopo la maturazione in vitro 7, 8. Infatti, in vitro oociti -matured possono produrre embrioni vitali che sopravvivono alla maturità e che sono fertili 3, 8. Questo metodo migliorato è stato ulteriormente adattato per includere la manipolazione funzionale dei geni materni, l'espressione di proteine con tag durante ovogenesi zebrafish tramite espressione esogena, e MO-mediata funzionale abbattere in stuoiafase IV urante ovociti 3 (Figura 1).
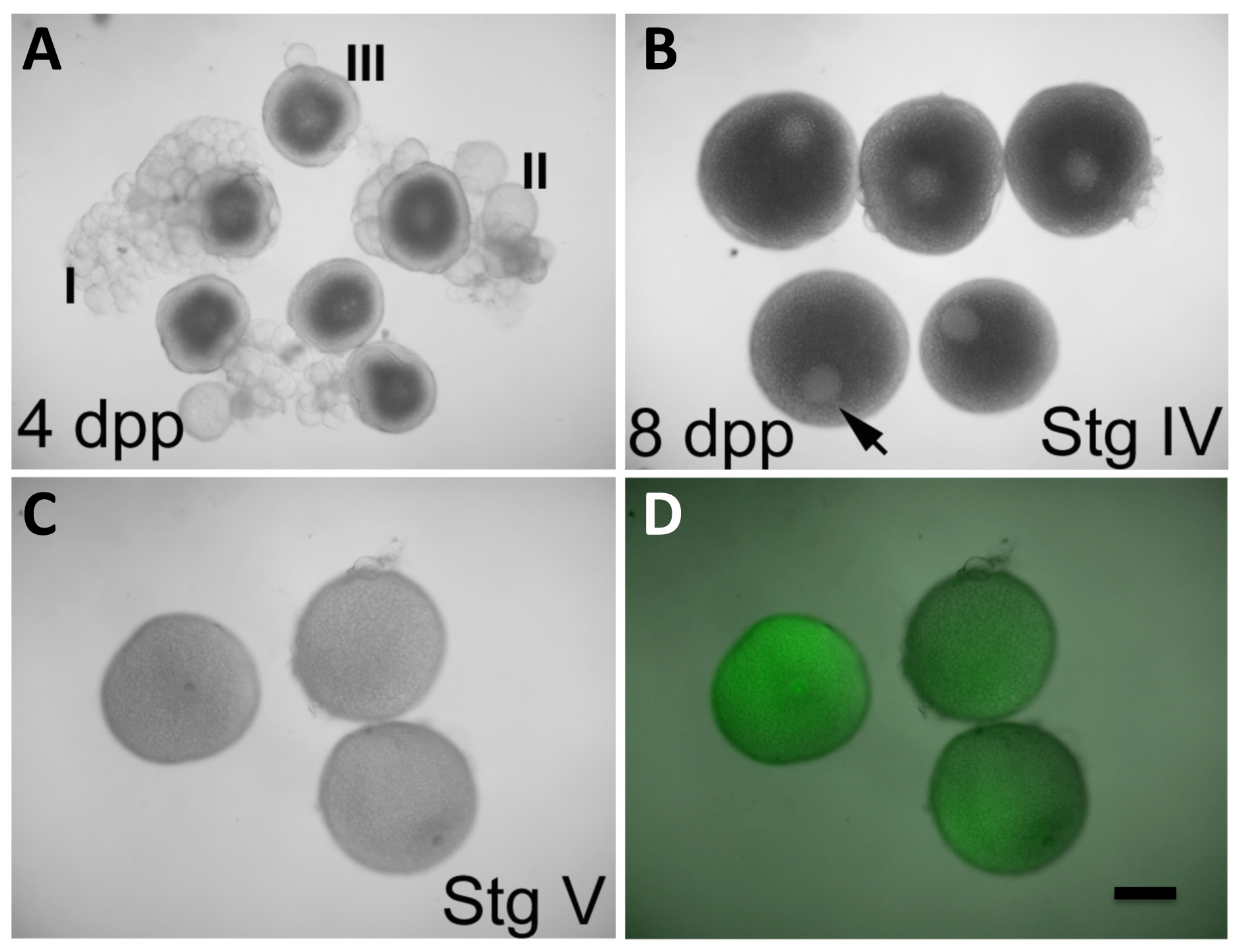
Oogenesis zebrafish presenta una serie di stadi caratteristici che portano alla formazione di ovociti maturi 9 (vedi Tabella 1 per un breve resoconto di varie fasi oogenesis). In breve, lo sviluppo degli ovociti è iniziata da stadio I ovociti e viene arrestato allo stadio di diplotene della profase I della meiosi. Questi ovociti sottoposti crescita mediante trascrizione attivo (iniziata durante le fasi IA e IB), la formazione di alveoli corticali (noto anche come granuli corticali, avviati durante la fase II), e vitellogenesi (iniziata durante la fase III). crescita oocita viene completata durante la fase IV, quando riprende la meiosi I, con conseguente smontaggio del nucleo ovocita, denominato vescicola germinale (GV). Successivamente, la meiosi viene nuovamente arrestato in metafase II. Il completamento della crescita dell'oocita e arresto meiotica durante la fase IV, porta ad un maturo fase V esg 9. La rimozione della membrana follicolare avviene durante il rilascio del ovocita nel lume dell'ovaio 10 ed è essenziale per la fertilizzazione e l'attivazione uovo corretta. Una volta che le uova sono estruse dalla madre durante l'accoppiamento naturale, diventano attivate. In zebrafish, esposizione ad acqua è sufficiente per l'attivazione completa uovo, indipendentemente dalla presenza di sperma 11.
In condizioni vitro attualmente consentire la maturazione degli oociti dalle prime fasi IV-riconoscibile dalla loro dimensione (690-730 micron), la presenza di una grande GV in posizione asimmetrica, e un aspetto completamente opaco a causa dell'accumulo di proteine tuorlo (Figura 2B) -per maturo uovo fase V, caratterizzati da un GV completamente smontato e un aspetto traslucido causa tuorlo processamento della proteina 12 (figura 2C). In questo approccio, le ovaie intere containing ovociti a diversi stadi di sviluppo vengono rimossi dalle femmine. Gli ovociti possono svilupparsi in 17α-20β-diidrossi-4 pregnen-3-one (DHP), un efficace maturazione che induce ormone 8. Durante questo periodo, ovociti maturazione possono essere manipolati attraverso l'iniezione di espressione (ad esempio, ridotta, in vitro -transcribed mRNA) o agenti reverse-genetica (ad esempio, MOS). Lo strato follicolare non spontaneamente sparso verso la fine del periodo di maturazione, quindi deve essere rimosso manualmente. Dopo defolliculation, la fecondazione in vitro viene raggiunto da innescare l'attivazione uovo attraverso l'esposizione delle uova all'acqua (presente in mezzo embrionale (E3)) 13 e una soluzione sperma. Gli zigoti risultanti subiscono sviluppo embrionale e permettono la valutazione della capacità di trattamenti di manipolare la funzione del gene materno e per la visualizzazione e l'analisi dei prodotti materni tag ( Figura 3).
Protocollo
Tutti zebrafish sono stati gestiti in stretta conformità con le buone pratiche degli animali, così come definito dagli organi nazionali e / o locali di benessere degli animali, e tutto il lavoro animale è stato approvato dalla commissione competente (University of Wisconsin-Madison numero garanzia A3368-01). Gli animali sono stati mantenuti in condizioni standard a 26,5 ° C.
1. Pre-selezione di femmine
NOTA: ovociti nelle femmine adulte tipicamente abbracciano una serie di stadi di sviluppo, dalla fase I a V (Figura 2). Spurgo le femmine di ovociti preesistenti attraverso un accoppiamento naturale successo aumenta la sincronizzazione di ovociti staging, come oociti di nuovo sviluppo appaiono come una coorte 3, 14. Da circa 8 giorni post-ventilazione, la maggior parte ovociti sono in fase iniziale IV, che è ottimale per l'inizio della maturazione in vitro. Tale sincronizzazione aumenta il rendimento di ovociti that può passare attraverso l'intero processo di maturazione in vitro, facilitando manipolazione sperimentale. Vedere le descrizioni precedenti 13 per i dettagli sulla configurazione di pesci in accoppiamenti appaiati e sulla composizione dell'acqua di pesce (qui, 14 g di sale marino e 150 g di NaHCO 3 per 1.000 litri di acqua ad osmosi inversa, pH 6,5-8,5 (intervallo preferito: 6.8 -7.5) e una conducibilità di 180-360 uS). Vedi Marca et al. 13 per le ricette supplementari.
- Otto a dieci giorni prima della manipolazione vitro in coltura, usa una rete di pesce per trasferire un singolo maschio e un singolo pesce femmina del ceppo desiderato ad un serbatoio di accoppiamento. Coppia più set. Lasciateli in una notte serbatoio di accoppiamento. Consentire loro di accoppiarsi durante la sera e per tutto il pomeriggio del giorno successivo.
- Utilizzare una rete dei pesci per separare le femmine che producono le uova durante l'accoppiamento e metterli in un serbatoio separato. Nutrire queste femmine due volte al giorno con un misto di cibo containing circa una pari quantità di artemie e scaglie di pesce cibo. La quantità di cibo dovrebbe essere sufficiente a fornire circa 20 minuti, ma non di più, di tempo di alimentazione.
2. Preparazione di Maturazione Medio
NOTA: Preparare medio di maturazione il giorno dell'esperimento maturazione in vitro, entro 1 ora di rimuovere gli ovociti dalla femmina (vedi sezione 3).
- Aggiungere 20 mL di mezzo L-15 di Leibovitz con L-glutammina, pH 7,0, ad una provetta da 50 ml in condizioni sterili. Portarlo a pH 9,0 con NaOH 10 N.
- Aggiungere 9 ml di L-15 medio di Leibovitz, pH 9,0, a 2 provette coniche separati 50 ml. Etichetta 1 tubo "+ DHP" e l'altro "-DHP".
- Nel tubo + DHP, aggiungere 10 ml di 17α-20β-diidrossi-4 pregnen-3-one (DHP), 490 ml di dH 2 O, e 500 ml di 10% di albumina sierica bovina (BSA).
- Nel tubo -DHP, aggiungere 100 ml di 10 mg / ml gentamicina, 40081; L di dH 2 O, e 500 microlitri di 10% di BSA.
3. La dissezione degli ovociti e iniziazione della coltura in vitro
NOTA: L'esperimento coltura in vitro è effettuata con le femmine preselezionati 8-10 giorni dopo che rilasciano le uova attraverso l'accoppiamento naturale. Utilizzare media stagionatura fatta lo stesso giorno (vedere paragrafo 2). Ovociti maturi appropriatamente se la dissezione è iniziata verso la fine del ciclo diurno pesce (determinata in laboratorio con luce artificiale preimpostata nella struttura pesce) 13, eventualmente mimando processi che avvengono attraverso accoppiamento naturale. Ciò implica che la maggior parte dei passaggi del protocollo devono essere eseguite in serata se i pesci sono alloggiati in una struttura con un ciclo di luce standard (ad esempio, a cominciare punto 3.2 alle 06:00 in una struttura con un periodo di luce 08:00-10:00), sebbene altri lavori periodi di tempo sono possibili con un ciclo luce opportunamente differita.
- Preparare una soluzione tricaine archivio di 0,2% in DH 2 O, tamponata a pH 7,0 con Tris 1 M, pH 9,0, e mantenere questa soluzione a 4 ° C. Questo può essere preparato prima del tempo.
- Avviare l'esperimento 0-4 h prima della fine del ciclo di luce giornaliera nella struttura. In un bicchiere da 250 ml, aggiungere 20 ml di 0,2% soluzione tricaine magazzino, pH 7,0, a 80 ml di acqua di pesce e mescolare.
- Trasferire le femmine pre-selezionati per la soluzione tricaine e l'eutanasia dalla sovraesposizione. Lasciare le femmine nella soluzione tricaine per 15 minuti dopo la cessazione del movimento Gill.
- Con un cucchiaino, raccogliere il pesce eutanasia dalla soluzione tricaine e sciacquarli brevemente in acqua i pesci. Mettere il pesce su un tovagliolo di carta per assorbire l'acqua in eccesso.
- Utilizzare una lama di rasoio pulito di decapitare i pesci eutanasia a livello della pinna pettorale. Utilizzando forbici dissezione, fare un'incisione longitudinale sul lato ventrale del pesce, che si estende dall'estremità anteriore alla zona anale.
- Mettere il pesce in una capsula di Petri sotto un microscopio da dissezione con luce incidente. Utilizzando un paio di dissettore, trasferire porzioni ovaio di una piastra da 35 mm x 10 coltura contenente 4 ml di Leibovitz L-15 medio + DHP.
NOTA: Le ovaie, che contengono ovociti di sviluppo, appariranno come strutture opache e clumpy all'interno della cavità interna del corpo. - Utilizzando dissettore, dissociarsi delicatamente gli ovociti dalle masse follicolari. In ordine le prime fasi IV ovociti (Figura 2B, prima della ripartizione GV, a dimensioni pressoché massima e caratterizzati da un buio, citoplasma opaco e prontamente apparente GV situato asimmetricamente nel ovocita). Scartare ovociti nelle fasi precedenti (Figura 2A) e, uova fase V mature traslucidi (simili a quelli nella Figura 2C ma presenti nelle ovaie prima del trattamento DHP).
NOTA: Gli ovociti vengono selezionati per la maturazione in fase iniziale IV, la prima tappa che si traduce in maturo, fase V oocytes dopo condizioni di coltura in vitro (Figura 2C e D) 3. Poiché ovociti stadio IV producono attivamente prodotti, la manipolazione in questa fase permette l'espressione di prodotti esogeni mediante iniezione o RNA per la riduzione della funzione del gene tramite MOs introdotte. - Utilizzare una pipetta Pasteur di vetro per trasferire i primi ovociti fase IV per una seconda mm piatto di plastica 35 x 10 coltura contenente 4 ml di Leibovitz L-15 medio + DHP. Trasferire quantità minime di mezzo -DHP al piatto contenente la soluzione + DHP.
4. Microiniezioni ovociti
NOTA: fase isolata IV ovociti sottoposti a maturazione in vitro possono essere microiniettati introdurre reagenti per la manipolazione o l'espressione funzionale proteina marcata. La microiniezione di reagenti, come mRNA e MOS (vedi paragrafo 4), viene tipicamente effettuata quando gli ovociti sono sottoposti in vitro maturazione in media + DHP, prima defolliculation. Se lo si desidera, in modo da consentire più tempo per effettuare manipolazioni prima della maturazione degli oociti, le iniezioni possono anche essere effettuate in mezzo -DHP, prima della maturazione, per almeno 2 ore. Possono poi essere trasferiti al mezzo + DHP.
- Preparare mRNA utilizzando un kit di espressione standard. tenerli a -80 ° C come 100-500 pg / ml magazzino per l'iniezione ad una concentrazione finale di 50-500 pg / ml (200 pg / mL è consigliato) in acqua RNA-grade. Preparare MO ad una concentrazione di 4 ng / ml in acqua secondo le istruzioni del produttore. li iniettare a concentrazioni finali di 2 ng / ml (o empiricamente determinata).
NOTA: Le iniezioni sono tipicamente condotte in terreno + DHP prima defolliculation (vedere la nota nella sezione 5). - Se l'iniezione di mRNA, immediatamente prima dell'iniezione, diluire il mRNA utilizzando acqua osmosi inversa RNA-grade e 0,2 M KCl per ottenere una soluzione finale di 0,1 M KCl.
- Se l'iniezione MO, immediatamente prima dell'iniezione, diluire i MOs alla concentrazione desiderata con acqua osmosi inversa e 0,2 M KCl per ottenere una soluzione finale di 0,1 M KCl.
- Tenere manualmente ovociti con pinza sottile e iniettare circa 1 nL in wild-type ovociti stadio IV utilizzando un ago con un capillare pipetta di vetro tirato.
- Preparare gli aghi di vetro tirando tubi capillari di vetro riscaldati, caricando la soluzione da iniettare, e rompendo la punta dell'ago con una pinza, come precedentemente descritto in protocolli di iniezione standard in zebrafish embrioni precoci 15.
NOTA: E 'utile se l'ago ha una conicità graduale piuttosto che un brusco uno; questo consente una maggiore flessibilità in termini di posizione della rottura della punta dell'ago e aiuta anche per evitare di danneggiare l'embrione iniettato. Utilizzando lo stesso ago e la soluzione, come per le iniezioni embrionali, regolare il volume iniettato per predeterminare laimpostazioni di pressione microiniettore che producono il volume desiderato. Queste impostazioni possono essere determinati finto-iniezione in una goccia di olio minerale su un vetrino di calibrazione fase di microscopio (0.01 mm) e regolare le impostazioni microinjector per ottenere il diametro desiderato nel bolo di soluzione iniettata, come descritto in precedenza 15.
- Preparare gli aghi di vetro tirando tubi capillari di vetro riscaldati, caricando la soluzione da iniettare, e rompendo la punta dell'ago con una pinza, come precedentemente descritto in protocolli di iniezione standard in zebrafish embrioni precoci 15.
5. ovociti Maturazione e Defolliculation
NOTA: durante la maturazione degli ovociti, gli ovociti diventeranno progressivamente traslucido, che consente per la valutazione delle condizioni di coltura di successo.
- Continuare incubare l'immaturo (e, se del caso, iniettato) ovociti in media + DHP a 26,5 ° C, controllando periodicamente (ogni 30 minuti) per garantire che gli ovociti rimangono intatti e sono sottoposti a corretta maturazione diventando progressivamente traslucido (vedere la Figura 2C) .
- Rimuovere eventuali ovociti lisi con una pipetta Pasteur e gettarliin un bicchiere di rifiuti di laboratorio per mantenere la qualità del mezzo. Sostituire il terreno di coltura con circa una mezza volume di fresco + DHP medio di mantenere una soluzione chiara, a seconda della quantità di ovociti lisi.
- Lasciare la maturazione in vitro procedere fino maggioranza degli ovociti diventano trasparenti e hanno una GV non più apparente (circa 2 h in trattamento DHP, vedere la Figura 2C rispetto a D). visualizzarli sotto un microscopio da dissezione con ottica luce trasmessa.
- Rimuovere la membrana follicolare più esterna di ciascun ovocita maturato. Utilizzare pinze finissimi di fare uno strappo nella membrana follicolare in una regione con maggiore spazio tra l'ovocita e la membrana. Staccare una porzione della membrana e rotolare l'ovocita fuori della membrana tenendo premuto il parte pelati.
NOTA: La membrana corionica sottostante tipicamente rimane in stretta associazione con l'uovo durante questo processo; dopo atto d'uovoivation, si espande per formare uno strato protettivo per l'embrione. - Trasferire gli ovociti defolliculated in un volume minimo di medio (tipicamente, trasferire circa 5-10 ovociti maturi in meno di 20 mL) ad una piastra di Petri con qualche goccia di coltura (+ DHP) medium e procedere alla fecondazione.
6. La fecondazione in vitro in coltura Ovociti
NOTA: La soluzione di sperma sul ghiaccio, preparata sotto, manterrà la sua potenza per circa 2 ore.
- Preparare una soluzione sperma vicino alla fine della fase di maturazione degli ovociti e prima defolliculation utilizzando testicoli da cinque maschi in 500 microlitri di soluzione di Hanks', come descritto in precedenza 16, 17.
- Aggiungere 10-50 ml di soluzione di sperma per gli ovociti defolliculated in terreno di coltura + DHP. Attendere 10 s.
- Usando una pipetta, aggiungere qualche goccia di embrionale (E3) a medio degli ovociti. Attendere 1 minuto e poi inondare il plmangiato con il mezzo E3.
NOTA: La composizione del terreno E3 è il seguente: 5 mM NaCl, 0,17 mM KCl, 0,33 mM CaCl 2, 0,33 mM MgSO 4, e 1-5% blu di metilene 13. - Ripetere i punti 5.3, 6.2, e 6.3 per ottenere un numero maggiore di embrioni fecondati.
- Lasciare gli embrioni fecondati di sviluppare. Utilizzare un microscopio da dissezione con ottica luce trasmessa per osservare la progressione attraverso le fasi scissione per assicurare la fecondazione di successo, come precedentemente descritto per la fecondazione 17 e 18 staging.
Risultati
Per determinare se la procedura sopra descritta è riuscita, gli embrioni possono essere osservati durante le fasi scissione per confermare l'aspetto del stereotipo precoce embrionale modello fenditura 18, nonché a 24 h post-fecondazione (HPF) per confermare il corretto sviluppo del piano di base del corpo. Questa procedura permette la manipolazione di prodotti materni per studi funzionali attraverso l'iniezione di reagenti, come mRNA e Mos, durante lo sviluppo dell'oocita.
Manipolazione di prodotti materni per studi funzionali:
codificante mRNA per wild-type e mutanti prodotti può essere iniettato in oociti in vitro scadenza per testare l'effetto della manipolazione della funzione genica materna durante la meiosi e le prime fasi embrionali. Per esempio, di tipo selvaggio embrioni possono essere iniettati con wild-tipo mRNA per testare l'effetto della sovraespressione prodotto, o con RNA mutante di prova per potenziali (ad esempio, il guadagno di funzione e antimorphic) effetti dominanti in tali processi. Ovociti in coltura in vitro sono in grado di produrre la proteina da mRNA esogena durante lo sviluppo degli ovociti, come dimostra l'espressione di GFP da iniettato mRNA, sebbene solo ovociti che avviano condizioni di coltura allo stadio IV dell'oogenesi possono svilupparsi in mature oociti fase V (Figura 2B -2D, puoi anche Nair et al. 3). L'espressione di wild-type prodotto attraverso iniettato mRNA è anche uno strumento per il salvataggio delle mutazioni materno-effetto per confermare l'identità del gene durante la clonazione posizionale 3, 24. In questo caso, wild-type mRNA viene iniettato in oociti da femmine omozigote mutante per verificare se i prodotti di tipo selvatico può salvare il fenotipo mutante. Questo salvataggio genetica è illustrato in Figura 3A-3C, dove iniettato AurB mRNA è mostrato per salvare gli effetti fenotipici di una mutazione nel suo gene corrispondente, isola cellulare (CEI) (Figura 3A-3C). Gli embrioni da un gruppo di controllo sono anche permesso di sviluppare per mostrare la corrispondente fenotipo mutante 3. RNA mutante può anche essere iniettato in oociti mutanti per verificare se il prodotto mutato conserva funzione parziale confrontando la misura di salvataggio a quella causata dal prodotto wild-type.
espressione della proteina da mRNA iniettato in oociti di sviluppo sembra verificarsi con poco o nessun ritardo. GFP forte si osserva entro 2 ore di iniezione del corrispondente mRNA, indipendentemente dallo stadio di sviluppo dell'oocita 3 (Figura 2B-2D, vedi anche Nair et al. 3). Prodotti come mCherry:Sas6 e Birc5b (Motley): GFP può essere osservato immediatamente dopo la fecondazione e durante il primo ciclo cellulare embrionale 3, 25. L'iniezione di mRNA durante oogenesis porta anche alla produzione di proteina che, indipendentemente dalla porzione contrassegnati fusa, è funzionale subito dopo la fecondazione, come illustrato nel caso di prodotti materni per isola cellulare / aurB 3, ciclo futile / lrmp 24, e Motley / bir5b 25. Traduzione blocco MOs hanno anche un effetto subito dopo la fecondazione, come mostrato per il ciclo futile / Lrmp 3 e nelle successive fasi di embriogenesi, come nel caso di missione impossibile / dhx16 3. Splice-blocco MOS, quando iniettato in ovociti in scadenza, non può avere un effetto sulla funzione materna, probabilmente a causa di già presenti mature trascritti materni in ovociti stadio IV 3.
Generazione di morphants:
Quando si utilizza un MO corrispondente ad un gene con una mutazione già identificati, il fenotipo morphant dovrebbe imitare il fenotipo mutante. Morphants sono confrontati con embrioni uninjected ea quelli iniettati con uno standard, controllare MO. L'iniezione di un morfolino Traduzione-bloccante può phenocopy successo il noto fenotipo mutante 3, come mostrato dalla iniezione di Lrmp MO in oociti, che imita il fenotipo mutante del corrispondente mutazione, ciclo futile (Figura 3D-F). La specificità di MOs può essere determinato attraverso gli stessi approcci usati quando si inietta MOs in embrioni (recensito in precedenza) 19, 20.
Espressione di fluorescenza etichettati proteine di fusione:
mRNA codificante per prodotti genici di interesse fusa con proteine fluorescenti (ad esempio, GFP e mCherry) può anche essere iniettato in wild-type o ovociti mutanti per visualizzare i prodotti corrispondenti ai embrione precoce (cioè, che riflette un modello localizzazione subcellulare; Figura 3G e 3H). mRNA codificanti per fusioni simili, ma che coinvolgono l'allele mutante possono essere espressi in modo simile per verificare se la mutazione influisce sui potenziali localizzazioni subcellulari.

Figura 1: in vitro maturazione degli oociti. Schema che mostra le varie fasi coinvolte nella vitro maturazione degli oociti. Le femmine adulte sono pre-selezionati per la deposizione delle uova 8 giorni prima della procedura, per loro eliminare le uova che hanno già maturato e per proMote nuovo sviluppo degli ovociti. Le ovaie, contenente ovociti di sviluppo, vengono rimossi da femmine e trasferiti in un terreno contenente l'ormone DHP per indurre la maturazione degli ovociti in vitro. Dopo la rimozione da femmine e immediatamente prima o durante la maturazione degli ovociti, ovociti possono essere iniettati con prodotti RNA e altri reagenti per la manipolazione funzionale di fattori materni. Ovociti maturi sono defolliculated manualmente e fecondati in vitro con una soluzione sperma per produrre fecondate, embrioni vitali. Si prega di cliccare qui per vedere una versione più grande di questa figura.
{kind=link}

Figura 2: In Vitro ovociti Maturazione ed espressione di mRNA Prodotti della iniettata. (A) ovociti nelle fasi I-III, osservati in ovaie difemmine 4 giorni post-spurgo (DPP). (B) ovociti in stadio IV, osservato in ovaie di femmine 8 DPP. Vescicola germinale (GV, freccia) è chiaramente visibile e occupa una posizione eccentrica. Gli ovociti stadio III in (A) mostrano anche una GV evidenti, ma si trova centrata nell'oocita. Ovociti Stage IV a (B) hanno una dimensione che è nei valori rispetto a quella di ovociti maturi in (C). (C e D) ovociti nella fase V (ovociti maturi) dopo 2 h in in vitro condizioni di maturazione avviati allo stadio IV, come in (B), e iniettate durante la maturazione con GFP-codifica, in vitro trascritto mRNA (C, luce visibile solo; D, sovrapposizione di luce visibile e fluorescenza GFP). GV è più evidente in oociti V fase, che sono anche meno opaca di ovociti stadio IV. Ovociti iniettati esprimono la proteina GFP (D). Defolliculation di maturi ovociti stadio IV è desCribed nella sezione del protocollo. bar scala = 300 um. Tutti i pannelli sono state riprodotte, con l'autorizzazione, da Nair et al 3. Si prega di cliccare qui per vedere una versione più grande di questa figura.
{kind=link}

Figura 3: manipolazione e visualizzazione dei materni prodotti attraverso in vitro maturazione degli oociti. (A - C) Salvataggio del fenotipo materno-effetto causato da una mutazione in un'isola cellulare / Aurora B attraverso l'iniezione di wild-type aurB mRNA in stadio IV CEI / aurB ovociti. fusione di immagini che mostrano viste animali di 65 blastodiscs fissi MPF visualizzati con anticorpi anti-ß-catenina per evidenziare membrane (verde), anticorpi anti-a-tubulina per indicare microtubuli (rosso) e DAPI di designare DNA (blu). (A) accumulo β-catenina in wild-type embrioni, indicativo della normale maturazione solco. (B) A CEI / aurB embrione da un non iniettato CEI / aurB stadio IV ovociti mostra parziali, solchi rudimentali che non accumulano β-catenina. (C) A cei salvato / aurB embrione da un cei / aurB ovocita iniettato con wild-type aurB mRNA mostra robusta accumulo β-catenina ai solchi. (D - F) phenocopy del materno-effetto causato da una mutazione nel ciclo futile / lrmp dall'iniezione di Lrmp morfolino nella fase IV oociti. fusione di immagini che mostrano viste animali di 70 blastodiscs fissi MPF visualizzati con anticorpi anti-gamma-tubulina per indicare centrosomi (rosso) e DAPI per designare DNA (blu). (D) in embrioni wild-type, ciascuno associa nucleo con γ-tubulina, marcatore di materiale centrosomica. (E) In materno effetto mutanti fue, fusione pronucleo fallisce, con conseguente due o tre macchie di etichette DNA corrispondenti a non fuse parentali pro-nuclei e il corpo polare per meiosi II. (F) In morphants Lrmp, dove è inibita funzione materna Lrmp, i nuclei simile riescono a dividere e, in aggiunta, sicuro associare a γ-tubulina. (G e H) Visualizzazione maternamente prodotti prodotto nell'embrione precoce. Sass6-mCherry proteina da mRNA esogena iniettato in fase IV ovociti localizza centrioli. fusione di immagini che mostrano viste animali di 40 blastodiscs fissi MPF visualizzati con anticorpi anti-gamma-tubulina per evidenziare centrosomi (verde), mCherry fluorescenza per indicare Sass6 (rosso) e DAPI di designare DNA (blu). (G) Espresso proteina Sass6-mCherry localizzati focolai etichettate dal centrosoma marcatori γ-tubulina in siti fiancheggianti nucleo (frecce). (H ) La stessa immagine, mostrando solo Sass6-mCherry e DAPI per chiarezza. Gli embrioni a (G e H) sono fisse, ma la fluorescenza dei prodotti espressa, come fusioni mCherry, possono anche essere osservati in embrioni vivi (non mostrati). Barra di scala = 100 micron di AF e 10 micron di G e H. Tutti i pannelli sono stati riprodotti, con l'autorizzazione, da Nair et al 3. Si prega di cliccare qui per vedere una versione più grande di questa figura.
{kind=link}
| Fasi di Oogenesi | Diametro ovocita (um) | Punto di riferimento chiave (s) |
| 1A - Prefollicle | 7 a 20 | Inizio della trascrizione attivo, accumulo di nucleoli |
| 1B - Follicolo | 20-140 | Decondensazione dei cromosomi |
| II - alveolo corticale | 140-340 | produzione di alveoli corticale |
| III - vitellogenesi | 340-690 | Oscuramento ooplasma dall'accumulo di tuorlo proteina precursore e lipidi |
| All'inizio IV - la maturazione degli ovociti | 690 a 730 (gamma più bassa) | localizzazione asimmetrica della vescicola germinale |
| All'inizio IV - la maturazione degli ovociti | 690 a 730 (scala) | vescicola germinale scompare, arresto in metafase II |
| V - ovocita maturo | ~ 750 | Ooplasma / tuorlo diventa traslucido |
Tabella 1: Limiti di ovociti sviluppo in Zebrapesce.
Discussione
Il protocollo di cui sopra è per la manipolazione di prodotti genici prima della fecondazione, permettendo così lo studio di prodotti genici materni nei primi anni del zebrafish. Studi precedenti hanno potuto maturare ovociti in vitro 4; Questo protocollo è stato modificato per consentire la successiva fecondazione in vitro ovociti 8 maturato. Ciò a sua volta consente per l'iniezione di reagenti per la manipolazione funzionale e la visualizzazione di prodotti maternamente ereditati nell'embrione precoce 3. Gli embrioni risultanti da questo metodo possono essere vitali e possono sopravvivere a diventare adulti fertili. In esperimenti preliminari, circa la metà degli embrioni fecondati derivati da questo procedimento fosse valida dal giorno 5 di sviluppo, come valutato dal inflazione vescica natatoria in quella fase, circa la metà dei quali è diventato sani, adulti fertili (osservazioni non pubblicate).
Ci sono una seriepassaggi di critiche da considerare nel protocollo. Gli oociti al momento opportuno per avviare la maturazione (inizio stadio IV) può essere arricchito se la femmina è stata accoppiata di recente, ma non prima di 8 DPP. Utilizzando pesce che non è accoppiate di recente potrebbe comportare degenerare uova 14, che non saranno sottoposti a maturazione. Le femmine fecondate in meno di 8 giorni avranno una maggioranza di uova in stadio III o meno, e, quindi, le uova non matureranno adeguatamente in vitro. Ovaie nelle femmine che accoppiabili 8 giorni prima avranno una frazione ottimale di ovociti in fase iniziale IV. Questi possono essere riconosciuti dalla loro opacità e la presenza del GV in posizione eccentrica (Figura 2B). Determinazione fase ovocita può anche essere aiutata da misura di formato, con gli ovociti posti su un vetrino micrometro graduato sotto un microscopio e confrontate con le linee guida stadiazione standard (Tabella 1). Tuttavia, IV ovociti fase iniziale hanno dimensioni quasi massimale rispetto a maturareovociti (riconosciuti dalla loro relativa trasparenza e la mancanza di una GV; la figura 2C) e sono facilmente riconoscibili, come sopra descritto, che ovvia generalmente la necessità di una misura diretta dimensioni ovocita.
Maturazione in vitro deve iniziare entro alcune ore dal termine del ciclo luce a cui sono acclimatati le donatrici di ovociti femminili. Gli oociti non matureranno adeguatamente, che si riflette nei tassi di fecondazione poveri, se coltivate durante la mattina del ciclo luce. La ragione per la dipendenza circadiano del metodo maturazione degli ovociti in vitro non è chiaro, ma probabilmente riflette pregiudizi circadiani sottostanti alle in vivo maturazione uovo coinvolgono bicicletta genica durante ovogenesi 21. Una causa comune di ovociti non riuscendo a subire successo maturazione in vitro nonostante corretta stadiazione ovociti è che la DHP è scaduta. ormone DHP scade in genere dopo un anno, e per assicurare l'effettola maturazione ive, il lotto di lavoro deve essere sostituita entro 9 mesi di utilizzo.
L'iniezione di ovociti è un altro aspetto critico quando lo scopo del metodo è la manipolazione funzionale dei prodotti materni. Questa procedura ha requisiti diversi da quelli per iniezioni standard nel uovo fecondato a 0-30 MPF. Un fattore chiave per iniezione degli ovociti è la necessità di eseguire le iniezioni in condizioni che non causano prematura attivazione uovo, come in L-15 medio di Leibovitz 22, 23. Perché sono incorporati in ovaie, gli ovociti in scadenza pongono anche particolari sfide per iniezione. Il protocollo sopra suggerisce primo dissociazione ovociti dalle ovaie e poi tenendosi dissociata ovocita separatamente con le pinze durante l'iniezione. Questa tecnica funziona adeguatamente e permette ovociti pre-smistamento in giusta fase con minima manipolazione. Metodi alternativi possono anche essere utilizzati, ad esempiocome: i) sistemazione oociti in iniezione standard trogoli agarosio 15, dove le pareti verticali del supporto trogolo dell'oocita durante l'iniezione (in questo metodo alternativo, gli ovociti devono essere trasferite in piastre di iniezione mentre mantenuti in vitro medio maturazione) e ii ) permettendo gli ovociti di rimanere attaccato alla massa ovarica, che può essere tenuto con una pinza durante l'iniezione per evitare il contatto con l'ovocita iniettato (in questo approccio, gli ovociti pertanto essere dissociata dopo l'iniezione sia a facilitare l'esposizione DHP e per permettere loro defolliculation , a sua volta essenziale per la fecondazione in vitro). Nonostante questo problema specifico ovociti stadio IV possono essere facilmente penetrati un ago di iniezione, probabilmente perché la membrana corionica in questa fase non è completamente sviluppato 9.
Una limitazione del protocollo maturazione attuale è che in condizioni di maturazione in vitro che promuovono corretto sviluppo dell'oocitanon può essere creato quando viene avviata la maturazione degli ovociti nelle fasi precedenti di stadio IV. Ovociti diventano competenti per rispondere a DHP subendo maturazione quando raggiungono un diametro di 520 micron, che si verifica durante la fase III (vitellogenesi, corrispondente al campo 340 a 690 micron) 9. Tuttavia, la cultura di fase III ovociti risultati in ovociti che non sono competenti per la fecondazione 3. Gli ovociti Solo cui coltura in vitro è avviato nella fase IV (Figura 2B) può svilupparsi in mature, ovociti fase V (Figura 2C e D). Questo può essere correlato al fatto che la fase III è essenziale per vitellogenesi e ovociti crescita 9. Così, la vitro procedura maturazione degli ovociti presentate in questo articolo deve essere utilizzato solo con una popolazione iniziale di stadio IV ovociti (B), e non con ovociti nelle fasi precedenti (stadio I - III; Figur e 2A). Per lo stesso motivo, questa procedura è limitato a geni che sono espressi allo stadio IV o successivo. La manipolazione funzionale di geni i cui prodotti sono espressi anteriore può invece fare affidamento su metodi genetici (vedi sotto). Miglioramenti ai metodi in vitro coltura maturazione possono in futuro consentire l'avvio di oocita coltura nelle fasi precedenti, ampliando così la potenza della vitro approccio maturazione in.
Un'altra limitazione nel metodo presentato è quello defolliculation, che è essenziale per le fasi successive che comportano la fecondazione, è attualmente un fattore limitante nella procedura. La membrana follicolare è uno strato trasparente che circonda l'oocita di sviluppo, e la rimozione può essere noioso. Se questa membrana non è completamente rimosso, l'uovo non sarà pienamente attivato e fecondato. Ciò è reso evidente dal fallimento del corion per espandere e lo sviluppo di irregolarmente livello locale, solco simile structures (pseudocleavages), caratteristici di embrioni fecondati 11. Metodi defolliculation enzimatici come collagenasi, come usati in Xenopus, sono stati provati. Tuttavia, nelle nostre mani, questa tecnica non è avuto successo, e abbiamo dunque avvalersi defolliculation manuale. Perché defolliculation manuale è e che richiede tempo ad alta intensità di lavoro, è difficile ottenere grandi lotti di uova fecondate dopo la maturazione in vitro degli ovociti. A causa della limitazione temporale imposta dal defolliculation manuale, che attualmente fertilizzare una qualche defolliculated, uova mature in un momento, in piccoli lotti di 5-10 uova. L'incorporazione di defolliculation enzimatica con questa procedura aumenterebbe probabilmente in modo significativo la sua resa e facilità d'uso.
Anche se gli embrioni prodotti dopo la fecondazione di ovociti -matured in vitro possono diventare adulti vitali, notiamo che dopo la fecondazione in vitro, una frazione di embrioni fare Non dividere normalmente o in modo tempestivo. Attualmente siamo selezione per le linee la cui propagazione comprende cicli di maturazione in vitro degli ovociti, che possono risultare in modelli più robusti di sviluppo in embrioni ottenuti tramite questo metodo.
La procedura ha consentito di soccorso chiara o visualizzazione di localizzazione delle proteine per una serie di mutazioni 24, 25. Tuttavia, per alcuni geni, la regolazione dell'espressione del prodotto può essere cruciale, così prodotti espressa con iniezione di mRNA possono portare a risultati variabili 26. E 'anche importante essere consapevoli di tutti i controlli (ad esempio, embrioni iniettati con reagenti che hanno proprietà simili a quelle utilizzate nell'esperimento ancora si prevedono di non produrre un effetto, come MOs controllo o RNA codificanti per proteine inerti solo tali come GFP), la variabilità sottostante può portare a conclusioni improprie.
nt "> Un approccio di manipolazione in vitro seguita da fecondazione è particolarmente utile nel caso di prodotti materna depositati, in cui il metodo standard di iniezione di RNA, MO, o proteine nell'uovo fecondato non può ridurre efficacemente prodotti già accumulato nell'uovo. altri metodi recentemente sviluppati, come la generazione CRISPR-mutante, sono anche in grado di generare condizioni mutanti per i geni materna espressi 26. Tuttavia, questo metodo richiede la crescita di diverse generazioni di pesce. la tecnica ovocita maturazione in vitro permette una rapida manipolazione funzionale studiare la funzione dei geni espressi materna, tra cui il salvataggio e phenocopy di mutazioni materno-effetto, così come l'espressione di RNA e proteine in embrione precoce.Divulgazioni
Gli autori non hanno interessi finanziari concorrenti.
Riconoscimenti
Vorremmo ringraziare i membri del laboratorio e pesce dirigenti impianto Pelegri, che hanno contribuito a facilitare questo progetto. Il supporto per questo progetto è stato fornito dal NIH R56 GM065303 e NIH 2 T32 GM007133 a ELW, NIH 5 F31 GM108449-02 e NIH 2 T32 GM007133 di CCE, e NIH RO1 GM065303 di FP
Materiali
| Name | Company | Catalog Number | Comments |
| Zebrafish mating boxes | Aqua Schwarz | SpawningBox1 | |
| NaCl | Sigma | S5886 | |
| KCl | Sigma | P5405 | |
| Na2HPO4 | Sigma | S3264 | |
| KH2PO4 | Sigma | P9791 | |
| CaCl2 | Sigma | C7902 | |
| MgSO2-7H2O | Sigma | 63138 | |
| NaHCO2 | Sigma | S5761 | |
| Tricaine | Western Chemical | Tricaine-D (MS 222) | FDA approved (ANADA 200-226) |
| Tris base | Sigma | 77-86-1 | to prepare 1 M Tris pH 9.0 |
| HCl | Sigma | 920-1 | to prepare 1 M Tris pH 9.0 |
| Fish net (fine mesh) (4 - 5 in) | PennPlax | (ThatFishThatPlace # 212370) | available in ThatFishThatPlace |
| Plastic spoon | available in most standard stores | ||
| Dissecting scissors | Fine Science Tools | 14091-09 | |
| Dissecting forceps | Dumont | SS | available from Fine Science Tools |
| Dissecting stereoscope (with transmitted light source) | Nikon | SMZ645 | or equivalent |
| Reflective light source (LED arms) | Fostec | KL1600 LED | or equivalent |
| Petri plates 10 cm diameter | any maker | ||
| Eppendorf tubes 1.5 mL | any maker | ||
| Ice bucket | any maker | ||
| Narrow spatula | Fisher | 14-374 | |
| Depression glass plate | Corning Inc | 722085 (Fisher cat. No 13-748B) | available from Fisher Scientific |
| Paper towels | any maker | ||
| Kimwipes | Kimberly-Clark | 06-666-11 | available from Fisher Scientific |
| Timer stop watch | any maker | ||
| Wash bottle | Thermo Scientific | 24020500 | available from Fisher Scientific |
| beakers, 250 mL (2) | Corning Inc. | 1000250 | available from Fisher Scientific |
| Leibovitz'z L-15 medium | Thermofisher | 11415064 | |
| NaOH | Sigma | 221465 | for pH'ing |
| BSA | Sigma | A2058 | |
| 17alph-20beta-dihyroxy-4-pregnen-3-one (DHP) | Sigma | P6285 | |
| gentamicin | Sigma | G1272 | |
| Injection Apparatus | Eppendorf | FemtoJet | or equivalent |
| Capillary Tubing for injection needles | FHC | 30-30-1 | or equivalent, Borosil 1.0 mm OD x 0.5 mm ID with fiber, 100 mm |
| Needle puller | Sutter Instruments | Model P-87 | any maker |
| Micropipetor (1 - 20 µL range) with tips | any maker | ||
| Micropipetor (20 - 200 µL range) with tips | any maker | ||
| Micropipetor (100 - 1000 µL range) with tips | any maker | ||
| Conical tubes 15 mL | any maker | ||
| Conical tubes 50 mL | any maker | ||
| plastic pipette 10 mL with bulb | any maker | ||
| plastic pipette 20 mL with bulb | any maker | ||
| Microscope stage Calibration Slide 0.01 mm | AmScope | MR095 | or equivalent |
| Reagents for fish water: | |||
| Instant Ocean Salt | Drs. Foster & Smith | CD-116528 | |
| Sodium bicarbonate (cell culture tested) | Sigma | S5761-1KG | |
| Reagents for E3 medium: | |||
| NaCl | Sigma | S5886-1KG | |
| KCl | Sigma | P5405-500G | |
| CaCl2, dihydrate | Sigma | C7902-500G | |
| MgSO4, heptahydrate | Sigma | 63138-250G | |
| Methylene Blue | Sigma | M9140-25G | |
| Fish Food: | |||
| Frozen brine shrimp | Brine Shrimp Direct | FBSFKG50 | |
| Tetramin Flakes | Drs. Foster & Smith | 16623 |
Riferimenti
- Lindeman, R., Pelegri, F. Vertebrate maternal-effect genes: insights into fertilization, early cleavage divisions, and germ cell determinant localization from studies in the zebrafish. Mol Rep Dev. 77 (4), 299-313 (2010).
- Abrams, E. W., Mullins, M. C. Early zebrafish development: it's in the maternal genes. Curr Opin Genet Dev. 19 (4), 396-403 (2009).
- Nair, S., Lindeman, R. E., Pelegri, F. In vitro oocyte culture-based manipulation of zebrafish maternal genes. Dev Dyn. 242 (1), 44-52 (2013).
- Selman, K., Petrino, T. R., Wallace, R. A. Experimental conditions for oocyte maturation in the zebrafish, Brachydanio rerio. J Exp Zool. 269 (6), 538-550 (1994).
- Fauvel, C., Omnes, M. H., Suquet, M., Normant, Y. Reliable assessment of overripening in turbot (Scophtalmus maximus) by a simple pH measurement. Aquaculture. 117 (1-2), 107-113 (1993).
- Lahnsteiner, F., Weismann, T., Patzner, R. A. Composition of the ovarian fluid in 4 salmonid speices: Onchorhynchus mykiss, Salmo trutta flacustris, Salvelinus alpinus and Husho hucho. Reprod Nutr Dev. 35 (5), 465-474 (1995).
- Patiño, R., Bolamba, D., Thomas, P., Kumakura, N. Effects of external pH on hormonally regulated ovarian follicle maturation and ovulation in Atlantic croaker. Gen Comp Endocrinol. 141 (2), 126-134 (2005).
- Seki, S., et al. Development of a reliable in vitro maturation system for zebrafish oocytes. Reproduction. 135 (3), 285-292 (2008).
- Selman, K., Wallace, R. A., Sarka, A., Qi, X. Stages of oocyte development in the zebrafish, Brachydanio rerio. J Morphol. 218 (2), 203-224 (1993).
- Clelland, E., Peng, C. Endocrine/paracrine control of zebrafish ovarian development. Mol Cell Endocrinol. 312 (1-2), 42-52 (2009).
- Kane, D. A., Kimmel, C. B. The zebrafish midblastula transition. Development. 119 (2), 447-456 (1993).
- Kanagaraj, P., et al. Souffle/Spastizin controls secretory vesicle maturation during zebrafish oogenesis. PLoS Genet. 10 (6), e1004449 (2014).
- Brand, M., Granato, M., Nüsslein-Volhard, C., Nüsslein-Volhard, C., Dahm, R. . Keeping and raising zebrafish,Zebrafish - A Practical Approach. , 7-37 (2002).
- Connoly, M. H., Dutkosky, R. M., Heah, T. P., Sayler, G. S., Henry, T. B. Temporal dynamics of oocyte growth and vitellogenin gene expression in zebrafish (Danio rerio). Zebrafish. 11 (2), 107-114 (2014).
- Rosen, J. N., Sweeney, M. F., Mably, J. D. Microinjection of zebrafish embryos to analyze gene function. J Vis Exp. (25), e1115 (2009).
- Pelegri, F., Mullins, M. Genetic screens for mutations affecting adult traits and parental-effect genes. Meth Cell Biol. 104, 83-120 (2011).
- Baars, D. L., Takle, K. A., Heier, J., Pelegri, F. Ploidy manipulation of zebrafish embryos with Heat Shock 2 treatment. J Vis Exp. , (2016).
- Kimmel, C., Ballard, W. W., Kimmel, S. R., Ullman, B., Schilling, T. F. Stages of embryonic development in the zebrafish. Dev Dyn. 203 (3), 253-310 (1995).
- Eisen, J. S., Smith, J. C. Controlling morpholino experiments: don't stop making antisense. Development. 135 (10), 1735-1743 (2008).
- Schulte-Merker, S., Stainier, D. Y. R. Out with the old, in with the new: reassessing morpholino knockdowns in light of genome editing technology. Development. 141 (16), 3103-3104 (2014).
- Dekens, M. P. S., Santoriello, C., Vallone, D., Frassi, G., Whitmore, D., Foulkes, N. S. Light regulates the cell cycle in zebrafish. Curr Biol. 13 (23), 2051-2057 (2003).
- Sakai, N., Burgess, S., Hopkins, N. Delayed in vitro fertilization of zebrafish eggs in Hank's saline containing bovine serum albumin. Mol Mar Biotechnol. 6 (2), 84-87 (1997).
- Pelegri, F., Mullins, M. C. Genetic screens for mutations affecting adult traits and parental-effect genes. Meth Cell Biol. 135, (2016).
- Lindeman, R. E., Pelegri, F. Localized products of futile cycle/lrmp promote centrosome-nucleus attachment in the zebrafish zygote. Curr Biol. 22 (10), 843-851 (2012).
- Nair, S., Marlow, F., Abrams, E., Kapp, L., Mullins, M., Pelegri, F. The chromosomal passenger protein Birc5b organizes microfilaments and germ plasm in the zebrafish embryo. PLoS Genetics. 9 (4), e1003448 (2013).
- Eno, C., Solanki, B., Pelegri, F. aura (mid1ip1l) regulates the cytoskeleton at the zebrafish egg-to-embryo transition. Development. 143 (9), 1585-1599 (2016).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati