
Method Article
仓鼠卵巢细胞的快速抗体糖工程
* 这些作者具有相同的贡献
摘要
抗体的糖基化模式决定了其临床表现,因此控制糖基化的工业和学术努力持续存在。由于典型的糖工程活动是时间和劳动密集型的,因此使用瞬时沉默来表征糖基化基因影响的快速方案的生成将被证明是有用的。
摘要
重组单克隆抗体结合特定的分子靶标,随后诱导免疫应答或抑制其他配体的结合。然而,单克隆抗体的功能和半衰期可能因宿主特异性糖基化的类型和分布而降低。产生优质抗体的尝试激发了合成糖优化抗体的转基因生产细胞的发展。糖工程通常需要使用诸如簇状规则间隔的短回文重复序列(CRISPR)相关蛋白9等方法生成稳定的敲除或敲除蛋白。然后使用质谱方法表征工程细胞产生的单克隆抗体,以确定是否已获得所需的糖原文件。这种策略非常耗时,技术上具有挑战性,并且需要专家。因此,利用简化的遗传糖工程和聚糖检测方案的替代策略可能有助于实现最佳抗体的努力。在这项概念验证研究中,产生IgG的中国仓鼠卵巢细胞是优化糖工程的理想宿主。将靶向 Fut8 基因的短干扰RNA递送到中国仓鼠卵巢细胞,并量化FUT8蛋白表达的变化。结果表明,这种方法的敲低是有效的,导致FUT8减少约60%。使用快速但高度灵敏的技术对抗体糖原进行补充分析:毛细管凝胶电泳和激光诱导的荧光检测。所有敲低实验都显示糖基化聚糖增加;然而,这项研究中实现的最大转变是~20%。该协议通过利用 计算机 设计工具,商业合成的基因靶向试剂和快速定量测定来简化糖工程工作,这些不需要丰富的先前经验。因此,该协议提供的时间效率可能有助于对新基因靶标的研究。
引言
N-连锁糖基化是一种酶促过程,通过该过程,寡糖部分与Asn残基共价连接。与 从头 蛋白质合成不同,聚糖合成是一种非模板化反应,导致蛋白质的非均相糖基化。聚糖的结构、组成和分布会影响蛋白质的构象和功能。事实上,免疫球蛋白G(IgG)的可结晶片段(Fc)区域中的 N-糖基化调节抗体1的治疗功效,免疫原性和半衰期。因此,用于开发重组生物治疗蛋白产品的质量源于设计(QbD)范式自然地将糖基化确定为关键质量属性(CQA)2,3。哺乳动物细胞通常是首选的表达系统,因为它们本质上比细菌,酵母,昆虫或植物细胞更能产生类似人类的糖基化模式。此外,中国仓鼠卵巢(CHO)细胞比其他哺乳动物细胞系选择,因为它们对人类病毒感染具有抗性,在高滴度下分泌产物,并且可以在悬浮培养中生长到高活细胞密度4。关于聚糖的形成,非CHO小鼠生产细胞产生免疫原性聚糖(α(1-3)连接的半乳糖[α(1-3)-Gal]和N-乙醇基神经氨酸[NeuGc]),影响单克隆抗体(mAbs)的安全使用5。这些好处使CHO细胞成为最重要的表达系统,负责在2014年至2018年期间生产超过80%的新生物治疗药物6。然而,模板无关糖基化是一种保守机制,可导致CHO衍生的生物治疗药物具有一系列糖型。
生物治疗开发策略旨在通过基因工程控制CHO细胞的异质性。一些文献例子包括唾液酶(Neu1,Neu3)7,GDP-甘露糖4,6-脱水酶(GMD)敲除8的敲除,以及糖基转移酶(GnTIII)的过表达9。由于公共资源(如CHO基因组10)的结合,以及遗传工程工具的持续发展,如转录激活子样效应核酸酶(TALENs),锌指核酸酶(ZFN)和簇状规则间隔的短回文重复序列(CRISPR)相关蛋白9(CRISPR-Cas9)11,12,13,14,14,糖工程的进步是可能的。.这些工具通常作为质粒DNA或纯化的核糖核蛋白(RNP)复合物递送到CHO细胞。相反,RNA干扰(RNAi)是一种基因工程技术,其最简单的形式只需要递送纯化的短干扰RNA(siRNA)寡核苷酸。内源性蛋白质将双链siRNA加工成单链,核酸酶RNA诱导的沉默复合物(RISC)与siRNA形成复合物以切割靶mRNA序列15,16,17。由于RNA的不稳定性,通过这种方法进行的基因沉默是短暂的,但本文的研究利用这一特征来帮助快速筛选。
为当前研究选择的模型酶α1,6-岩藻糖基转移酶(FUT8)产生具有α-1,6核连接的L-岩藻糖(Fuc)的N-聚糖。这种修饰是抗体依赖性细胞毒性(ADCC)活性的主要决定因素,正如商业抗体研究所证明的那样。在没有核心岩藻糖基化的情况下,利妥昔单抗(抗CD20 IgG1)通过改善FcgRIIIa结合18,19使曲妥珠单抗中的ADCC增加50倍并增加ADCC(抗Her2 IgG1)。因此,核心岩藻糖基化被认为是单克隆抗体的不良特征,需要努力逆转这种表型。有一些使用siRNA成功进行 Fut8 基因靶向的例子,伴随着ADCC20,21,22的增加,尽管这些例子提供了编码在质粒DNA上的 Fut8 siRNA。这样的实验产生稳定的基因沉默,因为质粒DNA是siRNA合成的模板。这允许细胞补充被细胞内RNases和磷酸酶降解的siRNA分子。相反,外源性合成siRNA的递送仅允许短暂的基因沉默,因为由于缺乏细胞内模板,siRNA无法补充。因此,用户应考虑实验设计是否与质粒衍生或合成的siRNA相容。例如,专注于峰单克隆抗体产生的研究,通常是培养物23,24的第六天,可以选择合成的siRNA,该siRNA可以在峰表达前几天递送到细胞。使用合成siRNA的瞬态方法的好处包括外包生产的能力,以及可以在质粒中生成构建体所需时间的一小部分内生成多个siRNA构建体的事实。此外,合成siRNA是有效的,正如 Fut8 基因沉默的文献实例所证明的那样,这足以降低FUT8蛋白表达25 并产生具有增加的FCgRIIIa结合和ADCC26的无糖基化IgG。
这种糖工程方案的成功取决于Fc糖基化的程度。质谱法通常是糖学分析的首选方法;然而,毛细管凝胶电泳和激光诱导荧光检测(CGE-LIF)完全适合于解析纯化IgG的糖质,并具有更高的快速性和简单性的优点。质谱实验方案必须结合适当的色谱和衍生化方法、电离源和质谱分析仪27、28、29。除了需要训练有素的专家外,质谱方案很长,方法的多样性使得具有不同设置的实验室之间难以比较数据。在生物制药的背景下,CGE-LIF是一种敏感的方法,可以提供抗体糖原文件的足够细节,并且可以轻松扩展为高通量方法。对于低丰度,高度复杂的混合物和表征不佳的糖蛋白,质谱法的优势可能仍然存在。然而,基于CGE-LIF的N-聚糖分析提供的高分辨率和高灵敏度单克隆抗体分析是试验这种方法的理由。此外,样品制备和分析在短短几个小时内完成30。最近的研究表明,CGE-LIF可用于监测来自人血浆31,小鼠32和CHO IgG33的聚糖。这些研究突出了CGE-LIF在高通量样品分析和小样品体积中的应用。
CGE-LIF方法具有应考虑的局限性。成本是使用这种和其他设备进行聚糖分析的重大障碍。然而,这些费用是外地的典型情况,CGE-LIF被认为是一种具有成本效益的备选办法34。预算较小的实验室可能会发现租赁机器或外包样品分析更实用。任何分析方法的另一个考虑因素是可重复性。使用在不同日期测定的同一样品的48个重复进行CGE-LIF的评估。测定每个毛细管的相对标准偏差,以确定配料内和配料间的可重复性。发现重复的批内比较具有6.2%的相对标准差,表明毛细管性能不均匀。此外,对产粒间数据的比较显示相对标准偏差为15.8%31,表明毛细管性能随时间而变化。所确定的操作缺陷可能不适用于当前的研究,该研究使用不同的机器和专有试剂。如果用户打算开发内部协议,那么值得考虑Ruhaak 等人的研究。31,仔细评估了CGE-LIF的试剂。因此,用于样品注射(Hi-Di甲酰胺和DMSO),聚糖标记(NaBH3CN或2-甲基吡啶硼烷)31等试剂已经过优化。
这项研究提出了一种节省时间的糖工程方案,该方案将直接RNAi的速度与下游糖组学分析相结合。由于上述原因,使用 Fut8 基因作为靶标说明了该方法。
研究方案
1. DsiRNA设计与重构
- 使用 Safari、Firefox 或 Microsoft Edge 访问 IDT 网站(https://eu.idtdna.com)。在主页上,选择"产品和服务 "选项卡,然后选择 RNA 干扰。要生成靶向 Fut8 基因的自定义干燥器底物 (DsiRNA) 构建体,请选择 "设计"工具 ,然后选择" 生成自定义 DsiRNA "选项卡。
- 输入 Fut8 加入号或手动输入基因序列即可开始。在这项研究中,从 https://chogenome.org 获得来自"CHO-K1"和"中国仓鼠"条目的 Fut8 序列,并用于产生DsiRNA。大多数基因都有几个转录本变异,重要的是要验证DsiRNA在当前组装中的所有报告变异上是否活跃。
- 选择执行"手动 BLAST"搜索的选项,因为 CHO 细胞基因组目前在 IDT 网站上无法作为"参考基因组"提供。此步骤搜索自定义DsiRNA序列和感兴趣基因组之间的匹配。
- 单击" 搜索 "以生成 DsiRNA 序列。
- 反过来,使用税号为:10029(CHO细胞系和中国仓鼠)的"手动BLAST"功能评估每种DsiRNA的特异性。查询覆盖率结果表明,DsiRNA构建体与 Fut8 (包括 Fut8 转录本变体)的匹配度为100%,而对非预期靶标的互补性为≤76%35。
- 检查DsiRNA是否专门针对 Fut8 ,以确保GC含量在30%-50%之间。低GC含量与弱和非特异性结合有关,而高GC含量抑制siRNA放卷并加载到RISC复合物36中。
- 选择三种DsiRNA用于转染研究(表1),每种都针对 Fut8 基因的不同位置。从 IDT 购买预先设计的非靶向或加扰 DsiRNA 进行对照实验。
- 到达后,将DsiRNA以13,300×g离心1分钟至沉淀,然后打开管。将无核酸酶双链缓冲液(由IDT提供)中的每种冻干DsiRNA重构到100μM储备溶液中。例如,将20μL缓冲液加入2nmol DsiRNA中以获得100μM储备液。
- 为确保充分混合,将储备溶液在室温下孵育30分钟,同时在轨道振荡器(50 rpm,16 mm轨道)上轻轻摇动。
- 通过结合20μL靶向 Fut8 的每种DsiRNA构建体(每个DsiRNA的100μM)来创建预混液。准备等分试样并储存在-20°C。
| DsiRNA靶标 | 序列 | 气相色谱(%) | ||
| 结构 A | 5' 嘎嘎 | 36% | ||
| 5' 咕 | ||||
| 结构 B | 5' 阿加奥加奥高乌克特 3' | 32% | ||
| 5' 阿加阿卡奥库库乌库加 3' | ||||
| 结构 C | 5' 阿加嘎 | 32% | ||
| 5' 瓜乌加库乌库 3' | ||||
表 1.用于 Fut8 敲低的DsiRNA序列。 IDT生成的靶向中国仓鼠和CHO K1细胞基因组中 Fut8 的序列。分别显示每个构造的检测和反义序列,并显示每个结构的 GC 内容。转载自Kotidis等人。52.
2. DsiRNA转染
- 使用适合目标细胞系的培养条件恢复表达IgG单克隆抗体37 的CHO细胞。
注意:本研究中使用的细胞是使用谷氨酰胺合成酶(GS)系统生成的,其中内源性GS被L-蛋氨酸磺酰肟(MSX)抑制,以增强稀有高产克隆的选择。因此,仅当所选细胞系需要时才使用MSX。- 在设置为37°C的水浴中解冻一小瓶细胞2-3分钟。 用70%(v / v)乙醇清洁小瓶外部,并在II类生物安全柜中继续所有工作。
- 将细胞悬浮液转移到含有9 mL预热培养基的15 mL离心管中,该培养基适合所选细胞系。通过以100×g离心5分钟沉淀细胞。
- 小心地去除并丢弃培养基,而不干扰细胞沉淀。然后,将细胞沉淀重悬于10mL预热培养基中,并取等分试样进行计数。
- 如果用血细胞计数器计数,则用台盼蓝染色细胞,或者在使用自动细胞计数器时用适当的染色剂来区分活细胞/死细胞。
- 遵循任一列举方法,将适当体积的细胞悬浮液转移到125 mL锥形瓶中,活细胞密度为3 x 105 cell·mL-1 ,在30 mL培养基中(可选补充50μM MSX)。
- 将细胞转移到设置为36.5°C,5%CO2 的培养箱中,并以150rpm(16 mm轨道)置于振荡平台上。
- 在250 mL锥形瓶中以2×105 细胞·mL-1 的接种密度和50mL的工作体积每3-4天传代一次细胞。如果使用MSX,请在第一次传代后停止补充。
- 传代细胞2x除解冻。
- 转染
- 评估细胞密度,确保细胞活力大于90%。
- 用70%(v / v)乙醇和RNase抑制剂溶液仔细清洁生物安全柜和所有设备,以避免污染。
- 以100×g沉淀细胞5分钟,重悬于预热培养基中至活细胞密度为5×106 个细胞·mL-1。
- 将8μL(相当于1μM)DsiRNA预混液或对照转移到无菌(0.4cm)电穿孔比色皿中。然后,将800μL细胞悬浮液(相当于4×106 个细胞)转移到同一比色皿中,并确保两种组分混合。
- 提供以下脉冲条件:1200 V、0.1 ms、方波。
- 将细胞悬浮液从比色皿转移到6孔板的一孔中,注意避免泡沫状材料。在培养箱(36.5°C,5%CO2)中回收细胞,而无需摇动10分钟。
- 加入800μL预热培养基,使最终体积达到每孔1.6mL,并将转染的细胞返回到培养箱中进行生长(36.5°C,5%CO2),同时以150rpm(16mm轨道)振荡。
- 转染后48小时收获上清液和细胞。
停止点:细胞培养上清液可保持在-20°C;然而,建议在细胞沉淀收集后立即进行细胞裂解,以避免蛋白质降解。上清液用于步骤3,步骤4和步骤5,而细胞沉淀用于步骤6。
3. IgG定量纯化
- 使用生物层干涉测量法或其他选择的方法测量IgG浓度。
- 在样品稀释剂中水合蛋白A生物传感器尖端10-30分钟。同时,将细胞悬浮液转移到15 mL离心管中,并以100×g沉淀细胞5分钟,或通过0.45μm硝酸纤维素过滤器过滤除去细胞。
- 在不干扰沉淀的情况下,小心地将含有IgG的分离上清液转移到干净的离心管中。
- 使用以下设置进行生物层干涉测量:振荡器速度,2200 rpm;运行时间,60 秒。这些参数将用于测量所有样品和对照。
- 一旦生物传感器尖端被水合,使用IgG标准品(每种浓度的4μL)创建标准曲线。
- 通过上样4μL细胞上清液定量样品浓度。在每个样品之间用无绒擦拭布清洁设备。
- 将标准曲线链接到未知样品,以插值未知样品的结合速率。保存数据并导出为 csv 或 PDF 文件。
- 免疫球蛋白纯化
- 制备含有0.2M甘氨酸(pH 2.5)的洗脱缓冲液,并通过0.22μm过滤器灭菌。
- 在室温下使用0.22μm微量离心机过滤管过滤1mL分离的上清液,直到整个上清液流过。
- 在1.5mL离心管中沉淀150-200μL蛋白A琼脂糖珠,以100×g离心3分钟并弃去上清液。然后,用150-200μL细胞培养基洗涤蛋白质A珠并重复离心。弃去上清液。
- 将平衡蛋白A珠与步骤3.2.3中制备的1mL上清液重悬。并装入1 mL聚丙烯管中。将聚丙烯管固定在旋转混合器(15 mm轨道)上,并在室温下以30rpm旋转60-90分钟或在4°C下过夜。
- 孵育完成后,收集流通物并用1 mL 1x PBS洗涤磁珠以除去任何未结合的蛋白质。同时收集洗涤部分。
- 通过在聚丙烯柱中加入3mL洗脱缓冲液,从蛋白质A珠中洗脱IgG。收集从色谱柱(每个500 μL)排出到标记管中的顺序级分。
- 可选:如果要储存纯化的抗体,请通过添加25μL 1 M Tris pH 9.5来中和洗脱缓冲液。如果抗体将通过缓冲液交换等立即处理,请跳过此步骤。
注意:纯化的所有部分(流通,洗涤和所有洗脱部分)应保持保持,以确保目标蛋白不会丢失。如果纯化不成功,这些样品也可以在故障排除过程中提供帮助。 - 使用第一次洗脱(洗脱液A)进行下游处理,但如果将来需要,请保留所有其他馏分。
4. 缓冲液更换和样品浓缩
- 将IgG洗脱缓冲液与1x PBS交换。
- 将洗脱液A加载到3 kDa分子量截止离心浓缩器上。选择合适的 MWCO 色谱柱时,请参阅制造商指南。在该实验中,较小的孔径确保了分泌蛋白质的最大保留,但也增加了离心时间。
- 在4°C下以13,300×g离心40-50分钟。 一旦残余体积等于或小于50μL,离心就完成了。
- 弃去流通液并加入500μL预稠化(4°C)1x PBS以稀释残留的上清液。使用相同的条件(13,300×g在4°C下40-50分钟)再次离心样品,直到残留50μL残留上清液。
- 加入500μL预沸(4°C)1x PBS以稀释残留的上清液,并再次重复离心过程以获得100x稀释的上清液。
- 将上清液浓缩至终浓度~2.5 g·L-1 在40μL或更低,以确保与聚糖分析方法的相容性。
注意:需要加载100μg用于聚糖分析。
停止点:浓缩的IgG(1x PBS)可以在-20°C下储存,并在聚糖分析之前解冻。
5. 聚糖分析
- 根据制造商的说明使用毛细管凝胶电泳进行聚糖分析。
- 将200μL磁珠溶液转移到0.2mL PCR管中,并置于磁性支架上以将磁珠与上清液分离。
- 小心地除去上清液,并从磁性支架上取出管子。加入纯化的蛋白质样品和涡旋,以确保完全混合。
- 将变性缓冲液(提供)加入样品管中,并在60°C下孵育8分钟。 保持样品管打开,以获得最佳反应性能。
- 加入PNGase F(每个样品500单位)并在60°C下孵育20分钟以从纯化的抗体中切割聚糖。
- 在释放N-聚糖后,关闭样品管并涡旋。加入乙腈,涡旋,并在室温下孵育1分钟。
- 将样品管放在磁性支架中,以将磁珠与溶液分离。使用移液器小心地除去上清液,不要接触珠子。
- 在通风橱中,将含有荧光团的聚糖标记溶液加入样品中。涡旋以确保充分混合并在60°C下孵育20分钟(打开盖子)。
- 将样品在乙腈中洗涤3次以除去多余的染料。然后,在DDI水中洗脱标记的聚糖(供应)。
- 将样品管置于磁性支架中,以将珠子与含有纯化和标记聚糖的上清液分离。
- 准备葡萄糖梯标准品,包围标准品和样品并将其装载到指定的托盘位置。运行聚糖分析方案。
- 使用适当的软件来分析和鉴定样品中存在的聚糖。
注意:确保加热块中的温度准确,以实现高效孵育。
6. 免疫印迹
- 使用抗α-1,6-岩藻糖基转移酶抗体进行蛋白质印迹分析,量化敲低效率。聚丙烯酰胺凝胶和缓冲液配方可从38,39商业供应商处获得。
- 在转染后48小时,使用血细胞计数器或自动细胞计数器计数细胞,并确定相当于5×106 个细胞的细胞悬浮液体积。
- 将适当体积的细胞悬浮液从每个实验条件转移到无菌的1.5mL离心管中,并在4°C下以13,200×g沉淀10分钟。 弃去上清液。
- 在室温下用含有1%v / v蛋白酶抑制剂混合物的200μL裂解缓冲液(适用于从哺乳动物细胞中提取蛋白质)裂解细胞10分钟。在孵育期间轻轻摇动混合物(50rpm,16 mm轨道)。
- 为了除去细胞碎片,将裂解物以13,200×g离心10分钟,然后将清除的裂解物转移到无菌的1.5mL管中。
- 使用280nm处的分光光度计测量每种裂解物的蛋白质浓度。随后,制备调整为具有相同蛋白质浓度的每种样品的等分试样。
- 通过在DTT-SDS样品上样染料中在100°C下孵育10-15分钟使蛋白质样品变性;最终染料浓度为1x。
- 将15μL变性样品和5μL预染蛋白分子量标准品上样到具有12.5%分辨凝胶的SDS-PAGE凝胶中,以实现高效分离。以每块凝胶25 mA的电流运行样品90分钟或直到染料前端到达凝胶的末端。
- 小心地从凝胶盒中取出凝胶,并在含有甲醇的1x转膜缓冲液中孵育。
- 准备湿转印系统并用甲醇激活PVDF膜。组装凝胶和PVDF膜进行湿转印,在转印罐中放置一个冰块,并将整个转印槽浸没在冰中以确保转印条件保持低温。在 350 mA/100 V 下运行 60 分钟。
- 通过在室温下在封闭溶液中孵育30分钟来阻断膜,同时在轨道振荡器上以50rpm(16mm轨道)轻轻摇动。
- 用无菌水冲洗膜5分钟并重复。然后,使用干净的手术刀以〜50 kDa的速度小心地水平切割膜,使用可见的蛋白质分子量标准作为指导。
- 将含有大于50 kDa的蛋白质的膜与抗α-1,6-岩藻糖基转移酶抗体在抗体稀释剂缓冲液中以1:1000孵育。在稀释剂缓冲液中用小于50 kDa的固定化蛋白质孵育膜,并在1:10,000处具有抗GAPDH。膜可以在室温下孵育1小时或在4°C下过夜。
- 洗涤膜5分钟(x3),然后在室温下与适当的二抗一起孵育至少30分钟。
- 用洗涤缓冲液进行3次洗涤5分钟,然后用水洗涤3次。然后,用显色底物(或适当的检测试剂)显影膜,直到条带出现(1-60分钟)。
- 用水冲洗膜2次,然后让其干燥。捕获膜的图像并执行密度分析40。
- 要计算每个样品中的相对蛋白表达,首先计算样品中GAPDH信号的比率。这是每个样本的归一化因子,用于校正样本加载量的差异。

- 将 FUT8 信号强度(对于每个泳道)除以相应泳道的归一化因子,得到相对 FUT8 蛋白表达。

结果
蛋白质印迹分析显示,用三种Fut8 DsiRNA构建体的混合物转染的细胞中 FUT8 蛋白表达降低。在用非靶向DsiRNA转染的对照样品中,FUT8在~65和70 kDa处表现为双条带。由于FUT8的预测分子量为66 kDa,因此低分子量条带信号强度的降低表明基因沉默。为了确认和量化基因沉默,FUT8蛋白的水平被归一化到相对GAPDH蛋白水平。蛋白质印迹分析在~37和35 kDa处检测到GAPDH的两条条带。较高的分子量条带对应于预测的蛋白质大小,因此用于归一化计算。当针对GAPDH蛋白水平进行归一化时,FUT8蛋白表达降低高达60%(图1)。
根据转染后48 h基因敲低的观察结果,通过CGE-LIF处理相应的单克隆抗体样品进行分析。来自敲低细胞的聚糖结构显示岩藻糖基化减少。这种趋势在无半乳糖基化结构(G0F)中最为明显,在半乳糖基化结构(G1F,G1F'和G2F)中观察到的程度较小。从该数据集中,总IgG核心岩藻糖基化从阴性对照条件下观察到的〜95%核心岩藻糖基化降至约75%(图2)。鉴于FUT8蛋白水平降低约60%,预计核心岩藻糖基化将有更大的减少。经过反思,值得注意的是,糖质前列代表自转染以来在48小时内积累的糖基化mAb,而基因沉默仅代表收获时存在的蛋白质水平。
对这种敲低方法的进一步检查涉及改变DsiRNA浓度,收获时间和电穿孔条件。对每个因素进行单独探测以确定其相关性。电穿孔脉冲条件对核心岩藻糖基化和细胞活力的影响在实验B,C,D和E中捕获。这些结果表明,与单个方波脉冲(实验B)相比,使用两个方波脉冲(实验C)进行电穿孔的岩藻糖基化减少了两倍,细胞活力没有显着差异(表2)。电穿孔条件e3(实验D)导致该时间点的细胞活力(〜90%)和IgG产量最低。然而,在电穿孔事件中幸存下来的细胞被适度转染,正如核心岩藻糖基化减少约10%所证明的那样(表2)。有趣的是,实验D使用的电穿孔条件使核心岩藻糖基化的最大减少(14.7%),但显然对细胞活力有害(91%-93%的活力)。这组有限的实验表明需要确定电穿孔设置,以使细胞膜具有足够的透化性而不会造成不可挽回的损伤。同样有趣的是,注意到siRNA浓度和收获时间对核心岩藻糖基化的作用。总体而言,增加siRNA浓度对核心岩藻糖基化的影响大于增加收获时间(实验B,F,G与实验A,B,H)。在未来的实验中,滴定电穿孔方法e2传递的siRNA浓度将是有趣的。

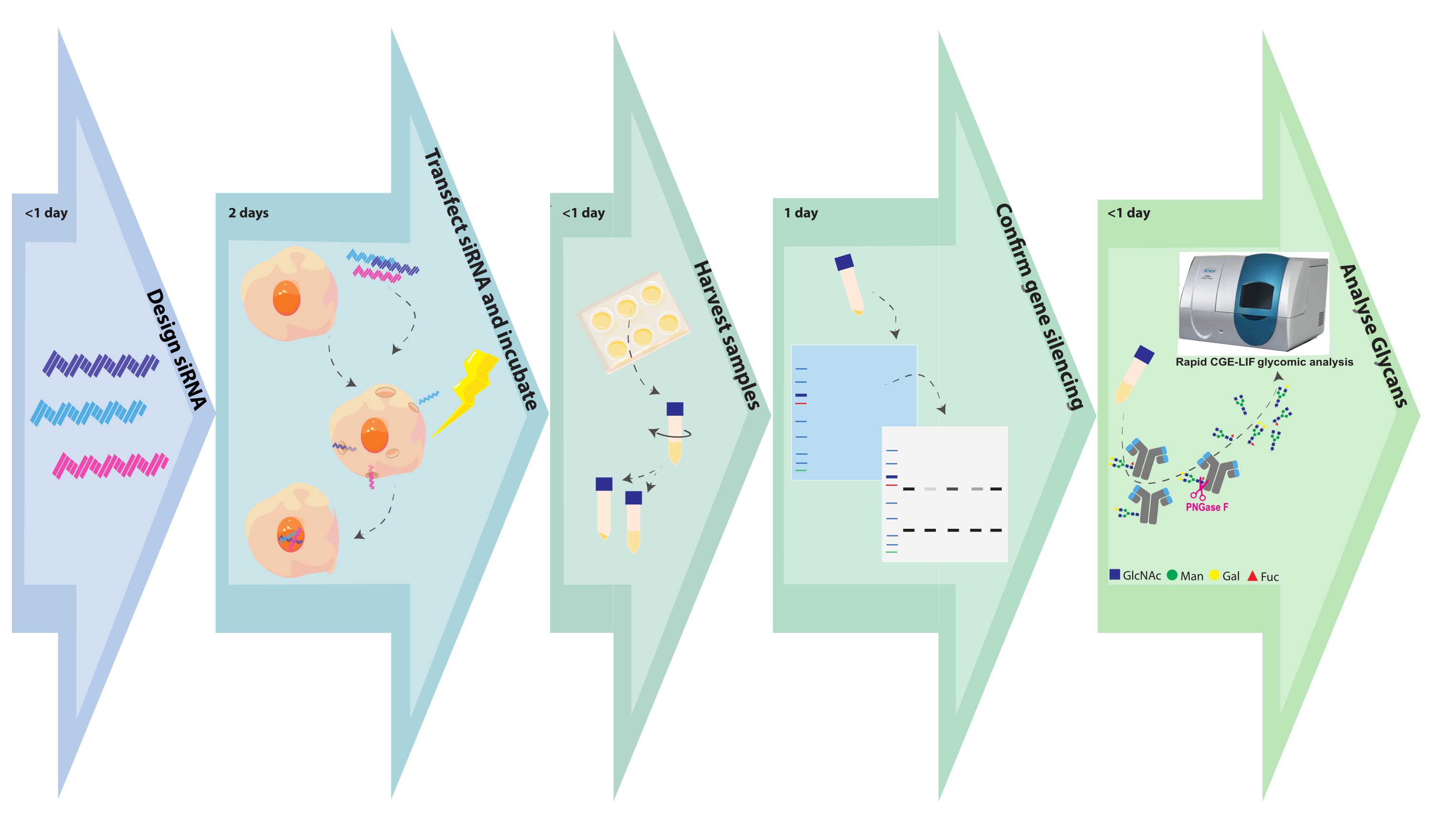
图 1.实验流程图。 糖工程和样品分析步骤与每个步骤所需的相关时间一起描述。SiRNA设计需要几个小时,这取决于每个基因靶标的基因靶标或构建体的数量。CHO细胞用siRNA转染在几个小时内完成,转化后的细胞生长48小时。细胞沉淀和上清液在几个小时内收获。裂解细胞沉淀,并在SDS PAGE上分离细胞内蛋白质,然后用针对靶基因的抗体进行印迹和探测。聚糖从纯化的抗体中切割出来,并用CGE-LIF进行分析。这些测定可能需要每次1天。 请点击此处查看此图的大图。
{kind=link}

图 2.确认RNA干扰。 蛋白质印迹检测用 Fut8 或非靶向对照DsiRNA处理的样品中的α-1,6-岩藻糖基转移酶(FUT8)蛋白水平。与 FUT8 对应的波段在控制上比 Fut8 敲低样本更强烈。还评估了GAPDH蛋白水平,以使靶基因表达正常化。所有样品均取自实验G(见 表2)。转载自Kotidis等人。52. 请按此浏览此图的大图。
{kind=link}

图 3. Fut8 敲低对48 h时累积IgG糖基化的影响. 在敲低样品中检测到聚糖分布的变化。特别是,在敲低实验中,主要核心岩藻糖基化结构(G0F)的相对丰度降低,而无糖基化物质增加。从实验G样品中进行测量(见 表2)。收获后对每个实验进行的生物三份混合,以减轻下游分析的负担。转载自Kotidis 等人。52. 请按此浏览此图的大图。
{kind=link}
| 实验名称 | 电穿孔法 | 双锡核糖核酸浓度(nΜ) | 收获时间(h) | 生存能力(%) | 十五 (106节・mL-1) | IgG滴度(毫克·L-1) | 岩藻糖基化率差异(%) | ||||||
| ExpA_Negative | e1 | 500 | 24 | 98.3 | 4.71 | 122.5 | - | ||||||
| ExpA_Knockdown | e1 | 500 | 24 | 98.3 | 4.9 | 110.3 | 4.08 | ||||||
| ExpB_Negative | e1 | 500 | 48 | 95.6 | 9.55 | 453.3 | - | ||||||
| ExpB_Knockdown | e1 | 500 | 48 | 96.7 | 9.61 | 469 | 5.38 | ||||||
| ExpC_Negative | e2 | 500 | 48 | 96.3 | 9.91 | 449.3 | - | ||||||
| ExpC_Knockdown | e2 | 500 | 48 | 96.7 | 11 | 454.6 | 11.42 | ||||||
| ExpD_Negative | e3 | 500 | 48 | 90.6 | 6.25 | 318.5 | - | ||||||
| ExpD_Knockdown | e3 | 500 | 48 | 89.1 | 6.09 | 311.85 | 9.71 | ||||||
| ExpE_Negative | e4 | 500 | 48 | 91.1 | 7.2 | 380.3 | - | ||||||
| ExpE_Knockdown | e4 | 500 | 48 | 93.3 | 7.79 | 422.8 | 14.7 | ||||||
| ExpF_Negative | e1 | 750 | 48 | 96.2 | 9.7 | 501 | - | ||||||
| ExpF_Knockdown | e1 | 750 | 48 | 95.7 | 9.76 | 504.6 | 9.9 | ||||||
| ExpG_Negative | e1 | 1000 | 48 | 96.1 | 11.1 | 422.6 | - | ||||||
| ExpG_Knockdown | e1 | 1000 | 48 | 95.9 | 9.73 | 499.3 | 17.26 | ||||||
| ExpH_Negative | e1 | 500 | 72 | 94.4 | 14.3 | 925.8 | - | ||||||
| ExpH_Knockdown | e1 | 500 | 72 | 95 | 13.5 | 1018.4 | 7.37 | ||||||
表 2.转染优化。 电穿孔方法、DsiRNA浓度和收获时间的迭代修饰导致细胞活力、活细胞密度、收获时IgG滴度和核心岩藻糖基化的差异发生变化。每个实验都比较了敲低和各自的阴性对照,以确定修饰是否产生所需的效果(即岩藻糖基化的减少)。电穿孔设置如下:e1:1200 V,0.1 ms,方波;e2: 1200 V, 2x 0.1 ms, 脉冲间5 s, 方波;e3: 150 V, 20 ms, 方波;e4:250 V,500 μF,指数衰减。转载自Kotidis等人。52.
讨论
糖基化途径涉及酶和辅助蛋白的复杂代谢网络。如果仅依靠传统的敲除或敲除基因工程策略,剖析途径成分的功能是令人生畏的。另一种方法是使用瞬时功能丧失测定法初步筛选通路成员。为此,将RNAi和CGE-LIF检测两种快速方案相结合,以创建一种更有效的方法来表征糖基化基因。所描述的方法需要5-7天才能完成,而传统方法可能需要数周才能完成。此外,具有自动化能力的研究环境可以利用这种方法筛选更多的候选基因,而不是手动处理。
瞬时糖工程活动的成功很大程度上取决于siRNA的设计。定制的DsiRNA设计必须遵循前面概述的规则,或者为了方便起见,用户可以选择市售的预先设计的序列。与其他基因修饰策略一样,RNAi具有脱靶效应的潜力。因此,鼓励用户通过计算方法41评估无意的基因靶向。实验设计选择也有助于限制脱靶效应。Kittler 等人 表明,siRNA的多重递送导致脱靶效应的减少42。虽然这似乎有悖常理,但有人建议预混液降低每个siRNA构建体的浓度,从而限制了脱靶基因沉默的机会。另一个好处是,同时转染靶向相同基因的siRNA结构增加了RNAi成功的可能性。使用预混液还可以确保样品和复制品之间的一致性,并加快转染过程。在对混合siRNA构建体进行初始筛选之后,可以使用单个构建体进行另一个实验以确定每个序列的RNAi效率。在这项和其他敲低研究中,多达三个siRNA被合并并递送到细胞43,44,45。然而,可能需要同时筛选三个以上的siRNA以有效地靶向单个基因或靶向多个基因。事实上,一项研究证明,多重siRNA介导的沉默多达六个基因的水平与单个基因的沉默水平相当46。然而,需要进一步的研究来确定可以在池中使用的siRNA构建体的最大数量,而不会影响沉默效率。Martin 等人 提出了多重策略,以加快RNAi文库筛选实验46的速度,类似的概念可能被证明对糖基化基因的筛选是有用的。
这里描述的协议作为概念验证,期望将进行后续实验以验证其他糖基化基因。感兴趣的新基因可能未表征或不如 Fut8受欢迎,并且检测基因沉默的一抗可能较差或不可用。在这种情况下,RT-PCR等替代方法可用于量化基因沉默47,但应该注意的是,RT-PCR检测的是mRNA而不是蛋白质。当有用于蛋白质印迹的抗体可用时,一个常见的问题是检测不良或存在非特异性条带。有帮助用户解决常见问题的故障排除指南,这些指南往往包括一系列解决方案,例如一抗滴定、替代阻断和检测条件48,49。在这项研究中,FUT8出乎意料地出现在~65和~70 kDa的双频带中。~70 kDa条带可能代表糖基化的FUT8。来自人类细胞系的文献证据描述了Thr 56450,51的O连锁糖基化,这是一个在中国仓鼠中保守的位点,以及CHO K1 FUT8序列。
如前所述,糖基化途径通常涉及复杂的酶阵列。目前的方案已经开发,优化,并证明使用由 Fut8控制的单基因糖基化反应。因此,当靶基因编码具有替代动力学和表达水平的酶或由具有冗余功能的同工酶调节的途径时,需要进一步的研究来确认该方法的稳健性。
总而言之,快速沉默基因和检测修饰的IgG糖原文件的能力是实现定制糖工程抗体的有用工具。来自类似短期研究的见解可以应用于生成稳定的糖工程细胞,用于长期测定,如补料分批培养。在药物环境之外,该方法有助于聚糖生物学的研究,并突出了聚糖在发育,健康和疾病中的重要功能。
披露声明
作者没有什么可透露的。
致谢
PK感谢伦敦帝国理工学院化学工程系的奖学金。RD感谢英国生物技术和生物科学研究委员会对他的学生身份。MM由英国生物技术和生物科学研究委员会资助(资助编号:BB/S006206/1)。IAG感谢爱尔兰研究委员会(奖学金编号:GOIPG/2017/1049)和CONACyT(奖学金编号438330)。
材料
| Name | Company | Catalog Number | Comments |
| 32 Karat software | SCIEX | contact manufacturer | Software for glycan data acquisition and analysis using the Fast Glycan analysis protocol and separation method. |
| Acetonitrile, HPLC grade | Sigma Aldrich | 34851 | Solvent. |
| Anti-FUT8 antibody | AbCam | ab198741 | Rabbit polycloncal to Fut8. Use this antibody to quantify Fut8 protein expression; replace this antibody if using siRNA targeting a different gene. |
| Anti-GAPDH antibody | AbCam | ab181602 | Rabbit monoclonal to GAPDH. Alternative housekeeping genes exist and might be preferred by the user. |
| BioDrop Spectrophotometer | Biochrom | 80 3006 55 | Instrument used to quantify protein concentration. |
| BLItz | ForteBio | 45 5000 | Instrument. Label-Free Protein Analysis System. |
| BRAND Haemocytometer | Sigma Alrich | BR717810 | Counting chamber device |
| Capillary cartridge | SCIEX | A55625 | Pre-assembled capillary cartridge with window (30 cm total length, 375 µm outer diameter (o.d), x 50 µm inner diameter (i.d). |
| C100HT Glycan analysis—capillary electrophoresis | SCIEX | contact manufacturer | Capillary gel electrophoresis instrument, the CESI 8000 Plus instrument is now used. |
| CD CHO Medium | Thermo Fisher Scientific | 10743029 | Replace this with a culture medium appropriate for the cell line of choice. |
| Centrifuge tubes, 15 mL | Greiner Bio | 188261 | Sterile polypropylene tube. |
| Centrifuge tubes, 50 mL | Greiner Bio | 227270 | Sterile polypropylene tube. |
| CHO IgG | MedImmune | Gift | Chinese Hamster Ovary cells expressing an IgG monoclonal antibody (CHO T127). Created using the GS system. |
| Dulbecco's phosphate-buffered saline (DPBS) | Gibco | 14190144 | 1x PBS, without calcium or magnesium. |
| Erlenmeyer Flasks with Vent Cap, 125 mL | Corning | 431143 | Replace this with a culture vessel suitable for growing the cell line of choice. |
| Erlenmeyer Flasks with Vent Cap, 250 mL | Corning | 431144 | Replace this with a culture vessel suitable for growing the cell line of choice. |
| Fast Glycan Labelling and Analysis kit | SCIEX | B94499PTO | Labels N-glycans with APTS and then uses a magnetic-bead based clean up system to remove excess APTS. |
| Fut8 DsiRNA | IDT | Custom | Custom designed DsiRNA targetting Fut8. |
| Gene Pulser cuvettes, 0.4 cm | Bio-Rad | 1652088 | Electroporation cuvette. |
| Gene Pulser Xcell Eukaryotic System | Bio-Rad | 165 2661 | Insturment. Xcell main unit with Capacitance Extender (CE) Mocdule and ShockPod. |
| Immobilon-FL PVDF membrane | Merck-Millipore | IPFL00010 | Immunoblot transfer membrane, low background. |
| L-Methionine sulfoximine (MSX) | Sigma Aldrich | M5379 | Only necessary for CHO cell lines using the glutamine synthetase (GS) selection system. |
| Kimwipes | Thermo Fisher Scientific | 10623111 | Low-lint, high absorbency and chemically inert wipes. |
| M-PER Mammalian Protein Extraction Reagent | Thermo Fisher Scientific | 78505 | Alternative lysis buffers such as RIPA are also appropriate. |
| Methanol, HPLC grade | Fisher Scientific | 10365710 | Solvent. |
| Microcentrifuge tubes, 1.5 mL | Eppendorf | 616201 | Autoclavable tubes. |
| Mini-PROTEAN Tetra Vertical Electrophoresis Cell system | Bio-Rad | 1658035FC | Instrument. 4-gel capacity, for 1.0 mm thick handcast gels, with Mini Trans-Blot Module and PowerPac HC Power Supply. |
| NC-Slide A8 | ChemoMetec | 942 0003 | 8-chamber slide for use with NucleoCounter NC 250. |
| Negative Control DsiRNA, 5 nmol | IDT | 51 01 14 04 | Non-targeting DsiRNA. |
| Nuclease-free duplex buffer | IDT | 11-01-03-01 | Reconstitution buffer for DsiRNA. |
| NucleoCounter NC-250 | Chemometec | contact manufacturer | Instrument. Automated Cell Analyzer |
| Page-Ruler ladder, 10 to 180 kDa | Thermo Fisher Scientific | 26616 | Mixed blue, orange and green protein standards for SDS PAGE and western blotting. |
| PCR tubes | Greiner Bio | 608281 | Autoclavable tubes for DsiRNA aliqouts and glycan preparation. |
| Pipette filter tips sterilised (10, 200, 1000 µL) | Gibson | F171203, F171503, F171703 | Sterile filter tips to avoid RNA contamination. |
| PNGase F enzyme | New England Biolabs | P0704S | Enzymatic cleavage of glycans from glycoproteins. |
| Polypropylene columns, 1 mL | Qiagen | 34924 | Columns for gravity-flow chromatography. |
| Protease Inhibitor Cocktail | Sigma Aldrich | P8340 | Inhibition of serine, cysteine, aspartic proteases and aminopeptidases |
| Protein-A Agarose Beads | Merck-Millipore | 16 125 | For purification of human, mouse and rabbit immunoglobulins. |
| Protein-A biosensor | ForteBio | 18 5010 | Tips functionalised with Protein A for rapid antibody quantification. |
| RNaseZap | Invitrogen | AM9780 | Removes RNAse contamination. |
| Sample dilutent | ForteBio | 18 1104 | Activate Protein A tips. |
| Serological pipets (5, 10, 25 mL) | Corning | 4487, 4488, 4489 | Used for sterile cell culture tecniques. |
| Sodium cyanoborohydride solution 1 M in THF | Sigma Aldrich | 296813 | Reducing agent. |
| Solution 18 | ChemoMetec | 910-3018 | Staining reagent containing acridine orange (AO) and 4',6-diamidino-2-phenylindole (DAPI) |
| Spin-X Centrifuge Tube Filters | Corning | 8161 | 0.22 µm pore, Cellulose Acetate membrane. |
| Suspension plate with lid, 6-well | Greiner Bio | 657 185 | Hydrophobic culture plate for growth of suspension cultures. |
| Syringe filters, 0.22 μm | Sartorius | 514 7011 | Surfactant-free cellulose acetate (SFCA) |
| Syringes with Luer lock tip, 20 mL | Fisher Scientific | 10569215 | For secure connection with syringe filter. |
| Trypan Blue solution | Gibco | 15250061 | Stains dead and dying cells. |
| Vivaspin 500, 3,000 MWCO | Sartorius | VS0191 | Polyethersulfone |
| WesternBreeze Chromogenic Kit, anti-rabbit | Thermo Fisher Scientific | WB7105 | Western blot detection kit, alternative blocking buffers and antibody diluents can be made by the user using recipes available online. |
参考文献
- Jefferis, R. Glycosylation as a strategy to improve antibody-based therapeutics. Nature Reviews Drug Discovery. 8 (3), 226-234 (2009).
- Rathore, A. S., Winkle, H. Quality by design for biopharmaceuticals. Nature Biotechnology. 27 (1), 26-34 (2009).
- Eon-Duval, A., Broly, H., Gleixner, R. Quality attributes of recombinant therapeutic proteins: An assessment of impact on safety and efficacy as part of a quality by design development approach. Biotechnology Progress. 28 (3), 608-622 (2012).
- Lalonde, M. -. E., Durocher, Y. Therapeutic glycoprotein production in mammalian cells. Journal of Biotechnology. 251, 128-140 (2017).
- Mimura, Y., et al. Glycosylation engineering of therapeutic IgG antibodies: challenges for the safety, functionality and efficacy. Protein & Cell. 9 (1), 47-62 (2018).
- Walsh, G. Biopharmaceutical benchmarks 2018. Nature Biotechnology. 36 (12), 1136-1145 (2018).
- Zhang, M., Koskie, K., Ross, J. S., Kayser, K. J., Caple, M. V. Enhancing glycoprotein sialylation by targeted gene silencing in mammalian cells. Biotechnology and Bioengineering. , (2010).
- Kanda, Y., et al. et al. of a GDP-mannose 4,6-dehydratase (GMD) knockout host cell line: a new strategy for generating completely non-fucosylated recombinant therapeutics. Journal of Biotechnology. 130 (3), 300-310 (2007).
- Umaña, P., Jean-Mairet, J., Moudry, R., Amstutz, H., Bailey, J. E. Engineered glycoforms of an antineuroblastoma IgG1 with optimized antibody-dependent cellular cytotoxic activity. Nature Biotechnology. 17 (2), 176-180 (1999).
- Xu, X., et al. et al. genomic sequence of the Chinese hamster ovary (CHO)-K1 cell line. Nature Biotechnology. 29 (8), 735-741 (2011).
- Chan, K. F., et al. et al. of GDP-fucose transporter gene (Slc35c1) in CHO cells by ZFNs, TALENs and CRISPR-Cas9 for production of fucose-free antibodies. Biotechnology Journal. 11 (3), 399-414 (2016).
- Zhang, P., et al. Identification of functional elements of the GDP-fucose transporter SLC35C1 using a novel Chinese hamster ovary mutant. Glycobiology. 22 (7), 897-911 (2012).
- Amann, T., et al. Genetic engineering approaches to improve posttranslational modification of biopharmaceuticals in different production platforms. Biotechnology and Bioengineering. 116 (10), 2778-2796 (2019).
- Karottki, K. J., et al. Awakening dormant glycosyltransferases in CHO cells with CRISPRa. Biotechnology and Bioengineering. 117 (2), 593-598 (2020).
- Fire, A., et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 391 (6669), 806-811 (1998).
- Hammond, S. M., Bernstein, E., Beach, D., Hannon, G. J. An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells. Nature. 404 (6775), 293-296 (2000).
- Bernstein, E., Caudy, A. A., Hammond, S. M., Hannon, G. J. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature. 409 (6818), 363-366 (2001).
- Shields, R. L., et al. et al. of fucose on human IgG1 N-linked oligosaccharide improves binding to human FcγRIII and antibody-dependent cellular toxicity. Journal of Biological Chemistry. 277 (30), 26733-26740 (2002).
- Shinkawa, T., et al. et al. Absence of fucose but not the presence of galactose or bisecting N-acetylglucosamine of human IgG1 complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity. Journal of Biological Chemistry. 278 (5), 3466-3473 (2003).
- Mori, K., et al. Engineering Chinese hamster ovary cells to maximize effector function of produced antibodies using FUT8 siRNA. Biotechnology and Bioengineering. 88 (7), 901-908 (2004).
- Imai-Nishiya, H., et al. Double knockdown of alpha1,6-fucosyltransferase (FUT8) and GDP-mannose 4,6-dehydratase (GMD) in antibody-producing cells: a new strategy for generating fully non-fucosylated therapeutic antibodies with enhanced ADCC. BMC Biotechnology. 7, 84 (2007).
- Beuger, V., et al. et al. silencing of fucosyltransferase 8 in Chinese-hamster ovary cells for the production of antibodies with enhanced antibody immune effector function. Biotechnology and Applied Biochemistry. 53 (1), 31 (2009).
- Templeton, N., Dean, J., Reddy, P., Young, J. D. Peak antibody production is associated with increased oxidative metabolism in an industrially relevant fed-batch CHO cell culture. Biotechnology and Bioengineering. 110 (7), 2013-2024 (2013).
- Hussain, H., et al. A comparative analysis of recombinant Fab and full-length antibody production in Chinese hamster ovary cells. Biotechnology and Bioengineering. 118 (12), 4815-4828 (2021).
- Shimoyama, H., et al. Partial silencing of fucosyltransferase 8 gene expression inhibits proliferation of Ishikawa cells, a cell line of endometrial cancer. Biochemistry and Biophysics Reports. 22, 100740 (2020).
- Tummala, S., et al. Evaluation of exogenous siRNA addition as a metabolic engineering tool for modifying biopharmaceuticals. Biotechnology Progress. 29 (2), 415-424 (2013).
- Carillo, S., et al. Comparing different domains of analysis for the characterisation of N-glycans on monoclonal antibodies. Journal of Pharmaceutical Analysis. 10 (1), 23-34 (2020).
- Reusch, D., et al. Comparison of methods for the analysis of therapeutic immunoglobulin G Fc-glycosylation profiles-Part 2: Mass spectrometric methods. mAbs. 7 (4), 732-742 (2015).
- Donini, R., Haslam, S. M., Kontoravdi, C. Glycoengineering Chinese hamster ovary cells: a short history. Biochemical Society Transactions. 49 (2), 915-931 (2021).
- Szigeti, M., Chapman, J., Borza, B., Guttman, A. Quantitative assessment of mAb Fc glycosylation of CQA importance by capillary electrophoresis. ELECTROPHORESIS. 39 (18), 2340-2343 (2018).
- Ruhaak, L. R., et al. Optimized workflow for preparation of APTS-labeled N-glycans allowing high-throughput analysis of human plasma glycomes using 48-channel multiplexed CGE-LIF. Journal of Proteome Research. 9 (12), 6655-6664 (2010).
- Patenaude, A. -. M., et al. N-glycosylation analysis of mouse immunoglobulin G isolated from dried blood spots. Electrophoresis. 42 (24), 2615-2618 (2021).
- Karst, D. J., et al. Modulation and modeling of monoclonal antibody N-linked glycosylation in mammalian cell perfusion reactors. Biotechnology and Bioengineering. 114 (9), 1978-1990 (2017).
- Dadouch, M., Ladner, Y., Perrin, C. Analysis of monoclonal antibodies by capillary electrophoresis: sample preparation, separation, and detection. Separations. 8 (1), 4 (2021).
- Fakhr, E., Zare, F., Teimoori-Toolabi, L. Precise and efficient siRNA design: a key point in competent gene silencing. Cancer Gene Therapy. 23 (4), 73-82 (2016).
- Birmingham, A., et al. A protocol for designing siRNAs with high functionality and specificity. Nature Protocols. 2 (9), 2068-2078 (2007).
- Makrydaki, E., et al. . Immobilised enzyme cascade for targeted glycosylation. , (2022).
- . Bulletin_6201.pdf Available from: https://www.bio-rad.com/webroot/web/pdf/lsr/literature/Bulletin_6201.pdf (2022)
- . Bulletin_6376.pdf Available from: https://www.bio-rad.com/webroot/web/pdf/lsr/literature/Bulletin_6201.pdf (2022)
- . Dot Blot Analysis Available from: https://imagej.nih.gov/ij/docs/examples/dot-blot/index.html (2022)
- Yilmazel, B., et al. Online GESS: prediction of miRNA-like off-target effects in large-scale RNAi screen data by seed region analysis. BMC Bioinformatics. 15 (1), 192 (2014).
- Kittler, R., et al. Genome-wide resources of endoribonuclease-prepared short interfering RNAs for specific loss-of-function studies. Nature Methods. 4 (4), 337-344 (2007).
- Stec, E., et al. A multiplexed siRNA screening strategy to identify genes in the PARP pathway. Journal of Biomolecular Screening. 17 (10), 1316-1328 (2012).
- Brown, J. M., et al. Ligand conjugated multimeric siRNAs enable enhanced uptake and multiplexed gene silencing. Nucleic Acid Therapeutics. 29 (5), 231-244 (2019).
- Parsons, B. D., Schindler, A., Evans, D. H., Foley, E. A direct phenotypic comparison of siRNA pools and multiple individual duplexes in a functional assay. PLoS One. 4 (12), 8471 (2009).
- Martin, S. E., et al. Multiplexing siRNAs to compress RNAi-based screen size in human cells. Nucleic Acids Research. 35 (8), 57 (2007).
- Popp, O., Moser, S., Zielonka, J., Rüger, P., Hansen, S., Plöttner, O. Development of a pre-glycoengineered CHO-K1 host cell line for the expression of antibodies with enhanced Fc mediated effector function. mAbs. 10 (2), 290-303 (2018).
- Yang, P. -. C., Mahmood, T. Western blot: Technique, theory, and trouble shooting. North American Journal of Medical Sciences. 4 (9), 429 (2012).
- Troubleshooting Western Blots with the Western Blot Doctor. Bio-Rad Laboratories Available from: https://www.bio-rad.com/en-uk/applications-technologies/troubleshooting-western-blots-with-western-blot-doctor?ID=MIW4HR15 (2022)
- Steentoft, C., et al. Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology. EMBO. 32 (10), 1478-1488 (2013).
- . FUT8 - Alpha-(1,6)-fucosyltransferase - Proteomics Available from: https://www.nextprot.org/entry/NX_Q9BYC5/proteomics (2022)
- Kotidis, P., et al. Rapid antibody glycoengineering in CHO cells via RNA interference and CGE-LIF N-glycomics. Methods in Molecular Biology. 2370, 147-167 (2022).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。