
Method Article
Schnelles Antikörper-Glykoengineering in chinesischen Hamster-Eierstockzellen
* Diese Autoren haben gleichermaßen beigetragen
In diesem Artikel
Zusammenfassung
Das Glykosylierungsmuster eines Antikörpers bestimmt seine klinische Leistung, so dass die industriellen und akademischen Bemühungen zur Kontrolle der Glykosylierung bestehen bleiben. Da typische Glykoengineering-Kampagnen zeit- und arbeitsintensiv sind, würde sich die Erstellung eines schnellen Protokolls zur Charakterisierung der Auswirkungen von Glykosylierungsgenen mittels transienter Silencing als nützlich erweisen.
Zusammenfassung
Rekombinante monoklonale Antikörper binden spezifische molekulare Ziele und induzieren anschließend eine Immunantwort oder hemmen die Bindung anderer Liganden. Die monoklonale Antikörperfunktionalität und Halbwertszeit kann jedoch durch die Art und Verteilung der wirtsspezifischen Glykosylierung verringert werden. Versuche, überlegene Antikörper zu produzieren, haben die Entwicklung gentechnisch veränderter Produzentenzellen inspiriert, die glykooptimierte Antikörper synthetisieren. Glykoengineering erfordert typischerweise die Erzeugung einer stabilen Knockout- oder Knockin-Zelllinie unter Verwendung von Methoden wie dem Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)-assoziierten Protein 9. Monoklonale Antikörper, die von künstlichen Zellen hergestellt werden, werden dann mit massenspektrometrischen Methoden charakterisiert, um festzustellen, ob das gewünschte Glykoprofil erhalten wurde. Diese Strategie ist zeitaufwendig, technisch anspruchsvoll und erfordert Spezialisten. Daher kann eine alternative Strategie, die optimierte Protokolle für das genetische Glykoengineering und den Glykannachweis verwendet, die Bemühungen um optimale Antikörper unterstützen. In dieser Proof-of-Concept-Studie diente eine IgG-produzierende chinesische Hamster-Eierstockzelle als idealer Wirt, um das Glykoengineering zu optimieren. Kurze störende RNA, die auf das Fut8-Gen abzielte, wurden an chinesische Hamster-Eierstockzellen abgegeben, und die daraus resultierenden Veränderungen in der FUT8-Proteinexpression wurden quantifiziert. Die Ergebnisse deuten darauf hin, dass der Knockdown mit dieser Methode effizient war, was zu einer ~ 60% igen Reduktion von FUT8 führte. Die komplementäre Analyse des Antikörperglykoprofils wurde mit einer schnellen, aber hochempfindlichen Technik durchgeführt: Kapillargelelektrophorese und laserinduzierter Fluoreszenzdetektion. Alle Knockdown-Experimente zeigten einen Anstieg der afucosylierten Glykane; Die größte Verschiebung, die in dieser Studie erreicht wurde, betrug jedoch ~ 20%. Dieses Protokoll vereinfacht die Glykoengineering-Bemühungen durch die Nutzung von In-silico-Design-Tools , kommerziell synthetisierten Gen-Targeting-Reagenzien und schnellen Quantifizierungsassays, die keine umfangreichen Vorkenntnisse erfordern. Daher können die von diesem Protokoll gebotenen Zeiteffizienzen die Untersuchung neuer Genziele unterstützen.
Einleitung
Die N-verknüpfte Glykosylierung ist ein enzymatischer Prozess, bei dem Oligosaccharid-Einheiten kovalent an Asn-Reste gebunden werden. Im Gegensatz zur De-novo-Proteinsynthese ist die Glykansynthese eine nicht vorlagenbasierte Reaktion, die zu einer heterogenen Glykosylierung von Proteinen führt. Die Struktur, Zusammensetzung und Verteilung von Glykanen kann die Proteinkonformation und -funktion beeinflussen. Tatsächlich reguliert die N-Glykosylierung in der Region des Immunglobulins G (IgG) im Bereich des kristallisierbaren Fragments (Fc) die therapeutische Wirksamkeit, Immunogenität und Halbwertszeit des Antikörpers1. Daher identifiziert das QbD-Paradigma (Quality by Design) für die Entwicklung rekombinanter biotherapeutischer Proteinprodukte die Glykosylierung auf natürliche Weise als kritisches Qualitätsmerkmal (CQA)2,3. Säugetierzellen sind oft die bevorzugten Expressionssysteme, da sie von Natur aus menschenähnliche Glykosylierungsmuster stärker produzieren als Bakterien, Hefen, Insekten oder Pflanzenzellen. Darüber hinaus werden Chinesische Hamster-Eierstockzellen (CHO) über anderen Säugetierzelllinien ausgewählt, da sie resistent gegen menschliche Virusinfektionen sind, Produkte bei hohen Titern absondern und in Suspensionskultur zu hohen lebensfähigen Zelldichten gezüchtetwerden können 4. In Bezug auf die Glykanbildung erzeugen nicht-CHO-murine Produktionszellen immunogene Glykane (α(1-3)-verknüpfte Galactose [α(1-3)-Gal] und N-Glykolylneuraminsäure [NeuGc]), die die sichere Verwendung von monoklonalen Antikörpern (mAbs)5 beeinträchtigen. Diese Vorteile machen CHO-Zellen zum führenden Expressionssystem, das für die Produktion von über 80% der neuen Biotherapeutika zwischen 2014 und 2018 verantwortlichist 6. Die vorlagenunabhängige Glykosylierung ist jedoch ein konservierter Mechanismus, der zu CHO-abgeleiteten Biotherapeutika mit einer Reihe von Glykoformen führt.
Biotherapeutische Entwicklungsstrategien zielen darauf ab, die Heterogenität in CHO-Zellen durch Gentechnik zu kontrollieren. Einige Literaturbeispiele sind der Knockdown von Sialidasen (Neu1, Neu3)7, GDP-Mannose 4,6-Dehydratase (GMD) Knockout8 und Überexpression von Glykosyltransferasen (GnTIII)9. Fortschritte im Glykoengineering sind möglich durch eine Kombination von öffentlich zugänglichen Ressourcen, wie dem CHO-Genom10, und der laufenden Entwicklung gentechnischer Werkzeuge, wie Transkriptionsaktivator-ähnliche Effektornukleasen (TALENs), Zinkfingernukleasen (ZFNs) und geclusterte regelmäßig interspaced short palindromic repeats (CRISPR)-assoziiertes Protein 9 (CRISPR-Cas9)11,12,13,14 . Diese Werkzeuge werden typischerweise als Plasmid-DNA oder als gereinigte Ribonukleoprotein (RNP) -Komplexe an CHO-Zellen abgegeben. Umgekehrt ist die RNA-Interferenz (RNAi) eine gentechnische Technologie, die in ihrer einfachsten Form nur die Abgabe von gereinigten kurzen störenden RNA (siRNA) Oligonukleotiden erfordert. Endogene Proteine verarbeiten doppelsträngige siRNA zu Einzelsträngen, und die Nuklease, RNA-induzierter Silencing-Komplex (RISC), bildet einen Komplex mit siRNA, um die Ziel-mRNA-Sequenzen15,16,17 zu spalten. Das Gen-Silencing über diese Methode ist aufgrund der RNA-Instabilität vorübergehend, aber die Untersuchung hierin nutzt diese Funktion, um ein schnelles Screening zu unterstützen.
Das für die aktuelle Studie ausgewählte Modellenzym, die α1,6-Fucosyltransferase (FUT8), produziert N-Glykane mit α-1,6-kerngebundener L-Fucose (Fuc). Diese Modifikation ist eine primäre Determinante der antikörperabhängigen Zellzytotoxizität (ADCC), wie Studien an kommerziellen Antikörpern belegen. In Ermangelung einer Kernfukosylierung erhöht Rituximab (Anti-CD20-IgG1) den ADCC um das 50-fache und erhöht den ADCC in Trastuzumab (Anti-Her2-IgG1) durch Verbesserung der FcgRIIIa-Bindung18,19. Die Kernfucosylierung wird daher als unerwünschtes Merkmal von mAbs angesehen, das Bemühungen zur Umkehrung dieses Phänotyps rechtfertigt. Es gibt Beispiele für erfolgreiches Fut8-Gen-Targeting unter Verwendung von siRNA mit gleichzeitigem Anstieg des ADCC20,21,22, obwohl diese Beispiele Fut8-siRNA liefern, die auf Plasmid-DNA kodiert ist. Solche Experimente erzeugen ein stabiles Gen-Silencing, da Plasmid-DNA als Vorlage für die siRNA-Synthese dient. Dies ermöglicht es den Zellen, siRNA-Moleküle aufzufüllen, die durch intrazelluläre RNasen und Phosphatasen abgebaut werden. Umgekehrt erlaubt die Abgabe von exogener synthetischer siRNA nur ein vorübergehendes Gen-Silencing, da siRNA aufgrund des Fehlens einer intrazellulären Vorlage nicht wieder aufgefüllt werden kann. Daher sollten Anwender überlegen, ob experimentelle Designs mit plasmidabgeleiteter oder synthetischer siRNA kompatibel sind. Zum Beispiel können Studien, die sich auf die maximale mAb-Produktion konzentrieren, typischerweise Tag sechs der Kultur23,24, sich für synthetische siRNA entscheiden, die einige Tage vor der Spitzenexpression an Zellen abgegeben werden kann. Zu den Vorteilen eines transienten Ansatzes unter Verwendung synthetischer siRNA gehören die Möglichkeit, die Produktion auszulagern, und die Tatsache, dass mehrere siRNA-Konstrukte in einem Bruchteil der Zeit erzeugt werden können, die zur Erzeugung von Konstrukten in Plasmiden benötigt wird. Darüber hinaus ist synthetische siRNA wirksam, wie Literaturbeispiele für Fut8-Gen-Silencing belegen, die ausreichen, um die FUT8-Proteinexpression25 zu reduzieren und afucosylierte IgGs mit erhöhter FCgRIIIa-Bindung und ADCC26 zu erhalten.
Der Erfolg dieses Glykoengineering-Protokolls wurde durch den Grad der Fc-Glykosylierung bestimmt. Die Massenspektrometrie ist normalerweise die Methode der Wahl für glykomische Analysen; Die Kapillargelelektrophorese und die laserinduzierte Fluoreszenzdetektion (CGE-LIF) eignen sich jedoch hervorragend zum Auflösen des Glykoprofils gereinigter IgGs und haben den Vorteil einer größeren Schnelligkeit und Einfachheit. Massenspektrometrieprotokolle müssen die entsprechenden chromatographischen und Derivatisierungsmethoden, Ionisationsquellen und Massenanalysatoren 27,28,29 kombinieren. Massenspektrometrieprotokolle erfordern nicht nur einen ausgebildeten Spezialisten, sondern sind auch langwierig, und die Vielfalt der Methoden macht es schwierig, Daten zwischen Labors mit unterschiedlichen Setups zu vergleichen. Im Kontext von Biopharmazeutika ist CGE-LIF eine empfindliche Methode, die ausreichende Details eines Antikörperglykoprofils liefern kann und für Hochdurchsatzmethoden leicht skalierbar ist. Bei niederfrequenten, hochkomplexen Gemischen mit schlecht charakterisierten Glykoproteinen könnten die Vorteile der Massenspektrometrie bestehen bleiben. Die hochauflösende und hochempfindliche mAb-Analytik, die durch die CGE-LIF-basierte N-Glykan-Analyse ermöglicht wird, dient jedoch als Begründung für die Erprobung dieser Methode. Darüber hinaus sind die Probenvorbereitung und -analyse in nur wenigen Stundenabgeschlossen 30. Jüngste Studien haben gezeigt, dass CGE-LIF zur Überwachung von Glykanen aus menschlichem Plasma31, Maus32 und CHO IgGs33 verwendet werden kann. Diese Studien unterstreichen die Verwendung von CGE-LIF für die Hochdurchsatz-Probenanalyse und kleine Probenvolumina.
Die CGE-LIF-Methode weist Einschränkungen auf, die berücksichtigt werden sollten. Die Kosten sind ein erhebliches Hindernis für die Verwendung dieses und anderer Geräte für die Glykananalyse. Diese Kosten sind jedoch typisch für das Feld, und CGE-LIF gilt als kostengünstige Option34. Labore mit kleineren Budgets können es praktischer finden, Maschinen zu leasen oder die Probenanalyse auszulagern. Eine weitere Überlegung jeder analytischen Methode ist die Wiederholbarkeit. Die Evaluierung von CGE-LIF wurde unter Verwendung von 48 Replikaten derselben Probe durchgeführt, die an verschiedenen Tagen untersucht wurden. Die relative Standardabweichung pro Kapillare wurde für die Intrabatch- und Interbatch-Wiederholbarkeit bestimmt. Der Intrabatch-Vergleich von Replikaten wies eine relative Standardabweichung von 6,2% auf, was darauf hindeutet, dass die Kapillarleistung nicht einheitlich ist. Darüber hinaus zeigte ein Vergleich der Interbatch-Daten eine relative Standardabweichung von 15,8 %31, was darauf hindeutet, dass sich die Kapillarleistung im Laufe der Zeit ändert. Die festgestellten betrieblichen Mängel gelten möglicherweise nicht für die aktuelle Studie, bei der andere Maschinen und proprietäre Reagenzien verwendet werden. Wenn Benutzer beabsichtigen, ein internes Protokoll zu entwickeln, lohnt es sich, die Studie von Ruhaak et al. in Betracht zu ziehen. 31, die die Reagenzien für CGE-LIF sorgfältig bewertete. Daher wurden die Reagenzien für die Probeninjektion (Hi-Di-Formamid und DMSO), die Glykanmarkierung (NaBH3CN oder 2-Picolin-Boran)31 und andere optimiert.
Diese Studie präsentiert ein zeiteffizientes Glykoengineering-Protokoll, das die Schnelligkeit der direkten RNAi mit der nachgeschalteten glykomischen Analyse kombiniert. Die Methodik wird anhand des Fut8-Gens als Ziel aus den oben genannten Gründen veranschaulicht.
Protokoll
1. DsiRNA-Design und Rekonstitution
- Verwenden Sie Safari, Firefox oder Microsoft Edge, um auf die IDT-Website (https://eu.idtdna.com) zuzugreifen. Wählen Sie auf der Startseite die Registerkarte Produkte und Dienstleistungen gefolgt von RNA-Interferenz. Um benutzerdefinierte DsiRNA-Konstrukte (DsiRNA) zu generieren, die auf das Fut8-Gen abzielen, wählen Sie das Design-Tool gefolgt von der Registerkarte Benutzerdefinierte DsiRNA generieren.
- Geben Sie die Fut8-Zugangsnummer ein oder geben Sie die Gensequenz manuell ein, um zu beginnen. In dieser Studie wurden die vorgeschlagenen Fut8-Sequenzen sowohl aus "CHO-K1"- als auch aus "Chinese Hamster"-Einträgen aus https://chogenome.org gewonnen und zur Erzeugung von DsiRNA verwendet. Die meisten Gene haben mehrere Transkriptvarianten, und es ist wichtig zu überprüfen, ob DsiRNA auf allen berichteten Varianten in der aktuellen Assemblierung aktiv ist.
- Wählen Sie die Option, um eine "Manuelle BLAST" -Suche durchzuführen, da das CHO-Zellgenom derzeit nicht als "Referenzgenom" auf der IDT-Website verfügbar ist. In diesem Schritt wird nach Übereinstimmungen zwischen benutzerdefinierten DsiRNA-Sequenzen und dem interessierenden Genom gesucht.
- Klicken Sie auf Suchen , um DsiRNA-Sequenzen zu generieren.
- Bewerten Sie wiederum die Spezifität jeder DsiRNA mit der Funktion "Manual BLAST" mit der Steuer-ID:10029 (CHO-Zelllinien und chinesischer Hamster). Die Ergebnisse der Abfrageabdeckung zeigen, dass DsiRNA-Konstrukte mit Fut8 (einschließlich Fut8-Transkriptvarianten) zu 100% übereinstimmen, während die Komplementarität gegenüber unbeabsichtigten Zielen ≤76% beträgt35.
- Überprüfen Sie, ob die DsiRNA speziell auf Fut8 abzielt, um sicherzustellen, dass der GC-Gehalt zwischen 30% und 50% liegt. Ein niedriger GC-Gehalt ist mit einer schwachen und unspezifischen Bindung verbunden, während ein hoher GC-Gehalt das Abwickeln und Laden von siRNA in den RISC-Komplex36 hemmt.
- Wählen Sie drei DsiRNA für die Verwendung in Transfektionsstudien aus (Tabelle 1), die jeweils auf einen anderen Ort des Fut8-Gens abzielen. Kaufen Sie eine vorgefertigte Nicht-Targeting- oder verschlüsselte DsiRNA von IDT für Kontrollexperimente.
- Bei der Ankunft zentrifugieren Sie die DsiRNA bei 13.300 x g für 1 min zum Pellet, bevor Sie das Röhrchen öffnen. Rekonstituieren Sie jede lyophilisierte DsiRNA in einem nukleasefreien Duplexpuffer (bereitgestellt von IDT) zu einer 100 μM-Stammlösung. Fügen Sie beispielsweise 20 μL Puffer zu 2 nmol DsiRNA hinzu, um einen 100 μM-Vorrat zu erhalten.
- Um eine ausreichende Durchmischung zu gewährleisten, inkubieren Sie die Stammlösungen bei Raumtemperatur für 30 Minuten und schütteln Sie sie vorsichtig auf einem Orbitalschüttler (50 U / min, 16 mm Umlaufbahn).
- Erstellen Sie einen Master-Mix, indem Sie 20 μL jedes DsiRNA-Konstrukts kombinieren, das auf Fut8 abzielt (100 μM jeder DsiRNA). Aliquots vorbereiten und bei −20 °C lagern.
| DsiRNA-Target | Reihenfolge | GC (%) | ||
| Struktur A | 5' GAGAAGAUAGAAACAGUCAAAUACC 3' | 36% | ||
| 5' GGUAUUUGACUGUUUCUAUCUUCUCUCUCUCUC 3' | ||||
| Struktur B | 5' AGAAUGAGAAUGGAUGUUUUUCCTT 3' | 32% | ||
| 5' AAGGAAAAACAUCCAUUCUCAUUCUGA 3' | ||||
| Struktur C | 5' AGAGAAGAUAGAAACAGUCAAAUAC 3' | 32% | ||
| 5' GUAUUUGACUGUUUCUAUCUUCUCUCG 3' | ||||
Tabelle 1. DsiRNA-Sequenzen, die für Fut8-Knockdown verwendet werden. Von IDT generierte Sequenzen, die auf Fut8 in chinesischen Hamster- und CHO-K1-Zellgenomen abzielen. Die Sense- und Antisense-Sequenzen für jedes Konstrukt werden (jeweils) angezeigt, und der GC-Inhalt jeder Struktur wird angezeigt. Nachdruck von Kotidis et al. 52.
2. DsiRNA-Transfektion
- Wiederbelebung von CHO-Zellen, die einen monoklonalen IgG-Antikörper37 unter Verwendung von Kulturbedingungen exprimieren, die für die interessierende Zelllinie geeignet sind.
HINWEIS: Die in dieser Studie verwendeten Zellen wurden mit dem Glutaminsynthetase (GS) -System erzeugt, bei dem endogenes GS durch L-Methioninsulfoximin (MSX) gehemmt wird, um die Auswahl seltener hochproduzierender Klone zu verbessern. Verwenden Sie MSX daher nur, wenn dies für die Zelllinie Ihrer Wahl erforderlich ist.- Tauen Sie eine Durchstechflasche mit Zellen für 2-3 Minuten in einem auf 37 °C eingestellten Wasserbad auf. Reinigen Sie die Außenseite der Durchstechflasche mit 70% (v/v) Ethanol und setzen Sie alle Arbeiten in einer Biosicherheitskabine der Klasse II fort.
- Überführen Sie die Zellsuspension in ein 15-ml-Zentrifugenröhrchen, das 9 ml vorgewärmtes Medium enthält, das für die Zelllinie der Wahl geeignet ist. Pelletieren Sie die Zellen durch Zentrifugation bei 100 x g für 5 min.
- Entfernen Sie das Medium vorsichtig und entsorgen Sie es, ohne das Zellpellet zu stören. Dann suspendieren Sie das Zellpellet in 10 ml vorgewärmtem Medium und nehmen Sie ein Aliquot zum Zählen.
- Färben Sie die Zellen mit Trypanblau, wenn Sie mit einem Hämozytometer zählen, oder mit einer geeigneten Färbung, um lebende / tote Zellen zu unterscheiden, wenn Sie einen automatisierten Zellzähler verwenden.
- Nach einer der beiden Aufzählungsverfahren wird ein geeignetes Volumen der Zellsuspension bei einer lebensfähigen Zelldichte von 3 x 105 Zellen·mL-1 in 30 ml Medium (mit optionaler Ergänzung von 50 μM MSX) in einen 125 ml Erlenmeyer-Schüttelkolben übertragen.
- Die Zellen werden auf 36,5 °C, 5 % CO2 eingestellt und mit 150 U/min (16 mm Umlaufbahn) auf eine Schüttelplattform gestellt.
- Passagezellen alle 3-4 Tage bei einer Aussaatdichte von 2 x 105 Zellen·mL-1 und einem Arbeitsvolumen von 50 mL in einem 250 mL Erlenmeyerkolben. Wenn Sie MSX verwenden, brechen Sie die Supplementierung nach der ersten Passage ab.
- Durchgangszellen 2x zusätzlich zum Auftauen.
- Transfektion
- Bewerten Sie die Zelldichte und stellen Sie sicher, dass die Zelllebensfähigkeit mehr als 90% beträgt.
- Reinigen Sie die Biosicherheitswerkbank und alle Geräte sorgfältig mit 70% (v/v) Ethanol und einer RNase-Inhibitorlösung, um eine Kontamination zu vermeiden.
- Pelletzellen bei 100 x g für 5 min und resuspendieren in vorgewärmtem Medium bis zu einer lebensfähigen Zelldichte von 5 x 106 Zellen·mL-1.
- Übertragen Sie 8 μL (entspricht 1 μM) der DsiRNA-Mastermischung oder -kontrolle auf eine sterile (0,4 cm) Elektroporationsküvette. Übertragen Sie dann 800 μL der Zellsuspension (entspricht 4 x 106 Zellen) auf dieselbe Küvette und stellen Sie sicher, dass beide Komponenten gemischt sind.
- Liefern Sie die folgenden Impulsbedingungen: 1200 V, 0,1 ms, Rechteckwellenform.
- Übertragen Sie die Zellsuspension von der Küvette auf eine Vertiefung einer 6-Well-Platte und achten Sie darauf, schaumartiges Material zu vermeiden. Stellen Sie die Zellen im Inkubator (36,5 °C, 5% CO2) ohne Schütteln für 10 min wieder her.
- Fügen Sie 800 μL vorgewärmtes Medium hinzu, um ein Endvolumen von 1,6 ml pro Vertiefung zu erhalten, und bringen Sie die transfizierten Zellen zum Wachstum (36,5 ° C, 5% CO2) in den Inkubator zurück, während sie bei 150 U / min (16 mm Umlaufbahn) schütteln.
- Ernten Sie die Überstände und Zellen bei 48 h nach der Transfektion.
Haltepunkt: Der Zellkulturüberstand kann bei -20 °C gehalten werden; Es ist jedoch ratsam, dass die Zelllyse unmittelbar nach der Zellpelletsammlung durchgeführt werden sollte, um einen Proteinabbau zu vermeiden. Überstände werden in Schritt 3, Schritt 4 und Schritt 5 verwendet, während Zellpellets in Schritt 6 verwendet werden.
3. IgG-Quantifizierung und -Reinigung
- Messen Sie die IgG-Konzentration mit Biolayer-Interferometrie oder einer anderen Methode Ihrer Wahl.
- Hydratprotein Ein Biosensor kippt in Probenverdünnungsmittel für 10-30 min. In der Zwischenzeit können Zellsuspensionen in 15-ml-Zentrifugenröhrchen und Pelletzellen bei 100 x g für 5 min übertragen oder Zellen durch Filtration durch einen 0,45-μm-Nitrocellulosefilter entfernt werden.
- Ohne das Pellet zu stören, geben Sie den isolierten Überstand, der IgG enthält, vorsichtig in ein sauberes Zentrifugenröhrchen.
- Verwenden Sie die folgenden Einstellungen für die Biolayer-Interferometrie: Shaker-Geschwindigkeit, 2200 U/min; Laufzeit, 60 s. Diese Parameter werden verwendet, um alle Proben und Kontrollen zu messen.
- Sobald die Biosensorspitzen hydratisiert sind, erstellen Sie eine Standardkurve mit einem IgG-Standard (4 μL jeder Konzentration).
- Quantifizierung der Probenkonzentrationen durch Beladung von 4 μL des Zellüberstandes. Reinigen Sie das Gerät mit fusselfreien Tüchern zwischen jeder Probe.
- Verknüpfen Sie die Standardkurve mit den unbekannten Stichproben, um die Bindungsrate der unbekannten Stichproben zu interpolieren. Speichern Sie die Daten und exportieren Sie sie als CSV- oder PDF-Datei.
- IgG-Reinigung
- Es wird ein Elutionspuffer mit 0,2 Mio. Glycin (pH 2,5) hergestellt und durch einen 0,22-μm-Filter sterilisiert.
- Filtern Sie 1 ml des isolierten Überstands bei Raumtemperatur mit 0,22 μm Mikrozentrifugenfilterröhrchen, bis der gesamte Überstand durchfließt.
- Pellet 150-200 μL Protein A Agaroseperlen in 1,5 mL Zentrifugenröhrchen bei 100 x g für 3 min und entsorgen den Überstand. Dann waschen Sie die Protein-A-Perlen mit 150-200 μL Zellkulturmedium und wiederholen Sie die Zentrifugation. Verwerfen Sie den Überstand.
- Resuspendiert die gleichgestellten Protein-A-Perlen mit 1 ml der hergestellten Überstände aus Schritt 3.2.3. und in ein 1 ml Polypropylenrohr laden. Befestigen Sie das Polypropylenrohr an einem Drehmischer (15 mm Umlaufbahn) und drehen Sie es bei 30 U / min für 60-90 min bei Raumtemperatur oder über Nacht bei 4 ° C.
- Nachdem die Inkubation abgeschlossen ist, sammeln Sie den Durchfluss und waschen Sie die Perlen mit 1 ml 1x PBS, um ungebundene Proteine zu entfernen. Sammeln Sie auch die Waschfraktion.
- Eluieren Sie IgG aus den Protein-A-Perlen durch Zugabe von 3 ml Elutionspuffer zur Polypropylensäule. Sammeln Sie sequentielle Fraktionen, die aus der Säule abfließen (jeweils 500 μl), in markierte Röhrchen.
- Optional: Wenn der gereinigte Antikörper gelagert werden soll, neutralisieren Sie den Elutionspuffer durch Zugabe von 25 μL von 1 M Tris pH 9,5. Überspringen Sie diesen Schritt, wenn der Antikörper sofort über Pufferaustausch usw. verarbeitet wird.
HINWEIS: Alle Teile der Reinigung (Durchfluss, Waschungen und alle Elutionfraktionen) sollten beibehalten werden, um sicherzustellen, dass das Zielprotein nicht verloren geht. Diese Beispiele können auch bei der Fehlerbehebung helfen, wenn die Reinigung nicht erfolgreich ist. - Verwenden Sie die erste Elution (Elution A) für die nachgelagerte Verarbeitung, behalten Sie jedoch alle anderen Fraktionen bei, falls sie in Zukunft benötigt werden.
4. Pufferwechsel und Probenkonzentration
- Tauschen Sie den IgG-Elutionspuffer mit 1x PBS aus.
- Laden Sie die Elution A auf einen 3 kDa Molekulargewichts-Cutoff-Zentrifugalkonzentrator. Beachten Sie bei der Auswahl einer geeigneten MWCO-Spalte den Leitfaden des Herstellers. In diesem Experiment sorgt eine kleinere Porengröße für eine maximale Retention des sezernierten Proteins, erhöht aber auch die Zentrifugationszeit.
- Zentrifuge bei 13.300 x g für 40-50 min bei 4 °C. Die Zentrifugation ist abgeschlossen, sobald das Restvolumen gleich oder kleiner als 50 μL ist.
- Verwerfen Sie den Durchfluss und fügen Sie 500 μL vorgekühlt (4 °C) 1x PBS hinzu, um den verbleibenden Überstand zu verdünnen. Die Probe wird unter den gleichen Bedingungen (13.300 x g für 40-50 min bei 4 °C) erneut zentrifugiert, bis 50 μL Restüberstand übrig sind.
- 500 μL vorgekühltes (4 °C) 1x PBS hinzufügen, um den Restüberstand zu verdünnen, und wiederholen Sie den Zentrifugationsprozess erneut, um eine 100-fache Verdünnung des Überstands zu erhalten.
- Konzentrieren Sie den Überstand auf eine Endkonzentration von ~2,5 g· L-1 in 40 μL oder weniger, um die Kompatibilität mit der Glykananalysemethode zu gewährleisten.
HINWEIS: Für die Glykananalyse müssen 100 μg geladen werden.
Haltepunkt: Das konzentrierte IgG (in 1x PBS) kann bei −20 °C gelagert und vor der Glykananalyse aufgetaut werden.
5. Glykananalyse
- Führen Sie eine Glykananalyse mit Kapillargelelektrophorese gemäß den Anweisungen des Herstellers durch.
- Übertragen Sie 200 μL der magnetischen Kügelchenlösung auf eine 0,2 mL PCR-Röhre und legen Sie sie auf den Magnetständer, um die Perlen vom Überstand zu trennen.
- Entfernen Sie vorsichtig den Überstand und entfernen Sie die Röhren vom Magnetständer. Fügen Sie die gereinigte Proteinprobe und den Wirbel hinzu, um eine vollständige Vermischung zu gewährleisten.
- Denaturierungspuffer (mitgeliefert) in das Probenröhrchen geben und 8 min bei 60 °C inkubieren. Halten Sie die Probenröhrchen offen, um eine optimale Reaktionsleistung zu erzielen.
- Fügen Sie PNGase F (500 Einheiten pro Probe) hinzu und inkubieren Sie für 20 Minuten bei 60 ° C, um Glykane von den gereinigten Antikörpern zu trennen.
- Nach der Freisetzung von N-Glykanen das Probenröhrchen und den Wirbel schließen. Acetonitril, Wirbel und 1 min bei Raumtemperatur inkubieren.
- Legen Sie die Probenröhrchen in den Magnetständer, um die Perlen von der Lösung zu trennen. Verwenden Sie eine Pipette, um den Überstand vorsichtig zu entfernen, ohne die Perlen zu berühren.
- In einem Abzug wird die Glykanmarkierungslösung, die ein Fluorophor enthält, in die Probe gegeben. Vortex zur Sicherstellung einer ausreichenden Durchmischung und Inkubation bei 60 °C für 20 min (offene Deckel).
- Waschen Sie die Probe 3x in Acetonitril, um überschüssigen Farbstoff zu entfernen. Dann eluieren Sie die markierten Glykane in DDI-Wasser (mitgeliefert).
- Legen Sie das Probenröhrchen in den Magnetständer, um die Perlen von dem Überstand zu trennen, der gereinigte und markierte Glykane enthält.
- Bereiten Sie den Glukoseleiterstandard, die Halterungsstandards und die Proben vor und laden Sie sie in die dafür vorgesehenen Tray-Positionen. Führen Sie das Glykananalyseprotokoll aus.
- Verwenden Sie geeignete Software, um die in der Probe vorhandenen Glykane zu analysieren und zu identifizieren.
HINWEIS: Stellen Sie sicher, dass die Temperatur im Wärmeblock für eine effiziente Inkubation genau ist.
6. Westlicher Schandfleck
- Quantifizieren Sie die Knockdown-Effizienz mithilfe der Western-Blot-Analyse mit einem Anti-α-1,6-Fucosyltransferase-Antikörper. Polyacrylamidgele und Pufferrezepturen sind bei kommerziellen Anbieternerhältlich 38,39.
- Zählen Sie die Zellen nach 48 h nach der Transfektion mit einem Hämozytometer oder einem automatisierten Zellzähler und bestimmen Sie das Volumen der Zellsuspension, das 5 x 106 Zellen entspricht.
- Übertragen Sie das entsprechende Volumen der Zellsuspension aus jedem Versuchszustand auf ein steriles 1,5 ml Zentrifugenröhrchen und Pellet bei 13.200 x g für 10 min bei 4 °C. Verwerfen Sie den Überstand.
- Lyse der Zellen mit 200 μL Lysepuffer (geeignet für die Extraktion von Proteinen aus Säugetierzellen), der 1% v/v Proteaseinhibitor-Cocktail enthält, bei Raumtemperatur für 10 min. Schütteln Sie die Mischung vorsichtig während der Inkubation (50 U / min, 16 mm Umlaufbahn).
- Um die Zelltrümmer zu entfernen, zentrifugieren Sie das Lysat bei 13.200 x g für 10 min und geben Sie dann das gereinigte Lysat in ein steriles 1,5-ml-Röhrchen.
- Messen Sie die Proteinkonzentration jedes Lysats mit einem Spektralphotometer bei 280 nm. Bereiten Sie anschließend Aliquots jeder Probe vor, die an die gleiche Proteinkonzentration angepasst sind.
- Denaturierung der Proteinproben durch Inkubation bei 100 °C für 10-15 min in DTT-SDS-Probenladefarbstoff; Die endgültige Farbstoffkonzentration beträgt 1x.
- Laden Sie 15 μL der denaturierten Proben und 5 μL vorgefärbte Proteinleiter in ein SDS-PAGE-Gel mit 12,5% auflösendem Gel für eine effiziente Trennung. Lassen Sie die Proben mit 25 mA pro Gel für 90 min oder bis die Farbstofffront das Ende des Gels erreicht.
- Entfernen Sie das Gel vorsichtig aus der Kassette und inkubieren Sie es in 1x Transferpuffer, der Methanol enthält.
- Bereiten Sie das Nasstransfersystem vor und aktivieren Sie eine PVDF-Membran mit Methanol. Montieren Sie das Gel und die PVDF-Membran für den Nasstransfer, legen Sie einen Eisblock in den Transfertank und tauchen Sie den gesamten Tank in Eis, um sicherzustellen, dass die Transferbedingungen kalt bleiben. 60 min lang mit 350 mA/100 V laufen.
- Blockieren Sie die Membran, indem Sie in einer blockierenden Lösung für 30 min bei Raumtemperatur inkubieren, während Sie sanft auf einem Orbitalschüttler bei 50 U / min (16 mm Umlaufbahn) schütteln.
- Spülen Sie die Membran 5 Minuten lang mit sterilem Wasser ab und wiederholen Sie dies. Verwenden Sie dann ein sauberes Skalpell, um die Membran vorsichtig horizontal bei ~ 50 kDa zu schneiden, wobei Sie die sichtbare Proteinleiter als Leitfaden verwenden.
- Inkubieren Sie die Membran, die Proteine von mehr als 50 kDa beherbergt, mit einem Anti-α-1,6-Fucosyltransferase-Antikörper bei 1:1000 in einem Antikörper-Verdünnungspuffer. Inkubieren Sie die Membran mit immobilisierten Proteinen unter 50 kDa mit Anti-GAPDH bei 1:10.000 im Verdünnungspuffer. Membranen können für 1 h bei Raumtemperatur oder über Nacht bei 4 °C inkubiert werden.
- Waschen Sie die Membranen für 5 min (x3) und inkubieren Sie dann mit einem geeigneten sekundären Antikörper für mindestens 30 min bei Raumtemperatur.
- Führen Sie 3x Wäschen mit einem Waschpuffer für 5 Minuten durch, gefolgt von 3x Wäschen mit Wasser. Entwickeln Sie dann die Membran mit einem chromogenen Substrat (oder einem geeigneten Nachweisreagenz), bis Banden auftreten (1-60 min).
- Spülen Sie die Membran 2x mit Wasser ab und lassen Sie sie trocknen. Nehmen Sie ein Bild der Membran auf und führen Sie eine densitometrische Analysedurch 40.
- Um die relative Proteinexpression in jeder Probe zu berechnen, berechnen Sie zunächst das Verhältnis des GAPDH-Signals in Proben. Dies ist der Normalisierungsfaktor für jede Probe, der Diskrepanzen beim Laden von Proben korrigiert.

- Teilen Sie die FUT8-Signalintensität (für jede Spur) durch den Normalisierungsfaktor der entsprechenden Spur, um die relative FUT8-Proteinexpression zu erhalten.

Ergebnisse
Die Western-Blot-Analyse zeigte eine reduzierte FUT8-Proteinexpression in Zellen, die mit einer Mischung aus drei Fut8-DsiRNA-Konstrukten transfiziert wurden. In Kontrollproben, die mit nicht zielgerichteter DsiRNA transfiziert wurden, erschien FUT8 als Doppelband bei ~65 und 70 kDa. Da das vorhergesagte Molekulargewicht von FUT8 66 kDa beträgt, ist eine Verringerung der Signalintensität des unteren Molekulargewichtsbandes ein Hinweis auf Gen-Silencing. Um das Gen-Silencing zu bestätigen und zu quantifizieren, wurde der FUT8-Proteinspiegel auf den relativen GAPDH-Proteinspiegel normalisiert. Die Western-Blot-Analyse ergab zwei Banden für GAPDH bei ~37 und 35 kDa. Das höhere Molekulargewichtsband entspricht der vorhergesagten Proteingröße und wird daher in Normalisierungsberechnungen verwendet. Bei Normalisierung gegen GAPDH-Proteinspiegel wurde die FUT8-Proteinexpression um bis zu 60% reduziert (Abbildung 1).
Im Einklang mit der Beobachtung des Gen-Knockdowns bei 48 h nach der Transfektion wurden entsprechende mAb-Proben für die Analyse mittels CGE-LIF aufbereitet. Glykanstrukturen aus Knockdown-Zellen zeigten eine Abnahme der Fukosylierung. Dieser Trend war in agalaktosylierten Strukturen (G0F) am ausgeprägtesten und in geringerem Maße in galaktosylierten Strukturen (G1F, G1F' und G2F) zu beobachten. Aus diesem Datensatz ging hervor, dass die gesamte IgG-Kernfucosylierung auf ~ 75% sank, verglichen mit ~ 95% der Kernfucosylierung, die für die negative Kontrollbedingung beobachtet wurde (Abbildung 2). Eine stärkere Verringerung der Kernfucosylierung wurde angesichts der ~ 60% igen Abnahme des FUT8-Proteinspiegels erwartet. Bei näherer Betrachtung ist es bemerkenswert, dass das Glykoprofil glykosylierte mAbs darstellt, die sich über einen Zeitraum von 48 h seit der Transfektion angesammelt haben, während Gen-Silencing nur Proteinspiegel darstellt, die zum Zeitpunkt der Ernte vorhanden sind.
Eine weitere Untersuchung dieser Knockdown-Methode beinhaltete die Variation der DsiRNA-Konzentration, der Erntezeit und der Elektroporationsbedingungen. Jeder Faktor wurde einzeln untersucht, um seine Relevanz zu bestimmen. Der Einfluss von Elektroporationspulsbedingungen auf die Kernfucosylierung und die Zelllebensfähigkeit wird in den Experimenten B, C, D und E erfasst. Diese Ergebnisse zeigen eine zweifache Reduktion der Kernfucosylierung durch Elektroporation unter Verwendung von zwei Rechteckwellenpulsen (Experiment C) im Vergleich zu einem einzelnen Rechteckwellenpuls (Experiment B), ohne signifikante Unterschiede in der Zelllebensfähigkeit (Tabelle 2). Die Elektroporationsbedingung e3 (Experiment D) führte zu diesem Zeitpunkt zu der niedrigsten Zelllebensfähigkeit (~ 90%) und IgG-Ausbeute. Zellen, die das Elektroporationsereignis überlebten, waren jedoch mäßig transfiziert, wie die ~ 10% ige Abnahme der Kernfucosylierung zeigt (Tabelle 2). Interessanterweise verwendete Experiment D Elektroporationsbedingungen, die die größte Verringerung der Kernfucosylierung (14,7%) lieferten, aber offensichtlich schädlich für die Zelllebensfähigkeit waren (91% -93% Lebensfähigkeit). Diese begrenzte Anzahl von Experimenten veranschaulicht die Notwendigkeit, Elektroporationseinstellungen zu bestimmen, die eine ausreichende Permeabilisierung der Zellmembran ermöglichen, ohne unwiderrufliche Schäden zu verursachen. Es ist auch interessant, die Rolle der siRNA-Konzentration und der Erntezeit bei der Kernfukosylierung zu beachten. Insgesamt hat die Erhöhung der siRNA-Konzentration einen größeren Einfluss auf die Kernfucosylierung als die Erhöhung der Erntezeit (Experimente B, F, G versus Experimente A, B, H). In zukünftigen Experimenten wäre es interessant, die siRNA-Konzentrationen der Elektroporationsmethode e2 zu titrieren.

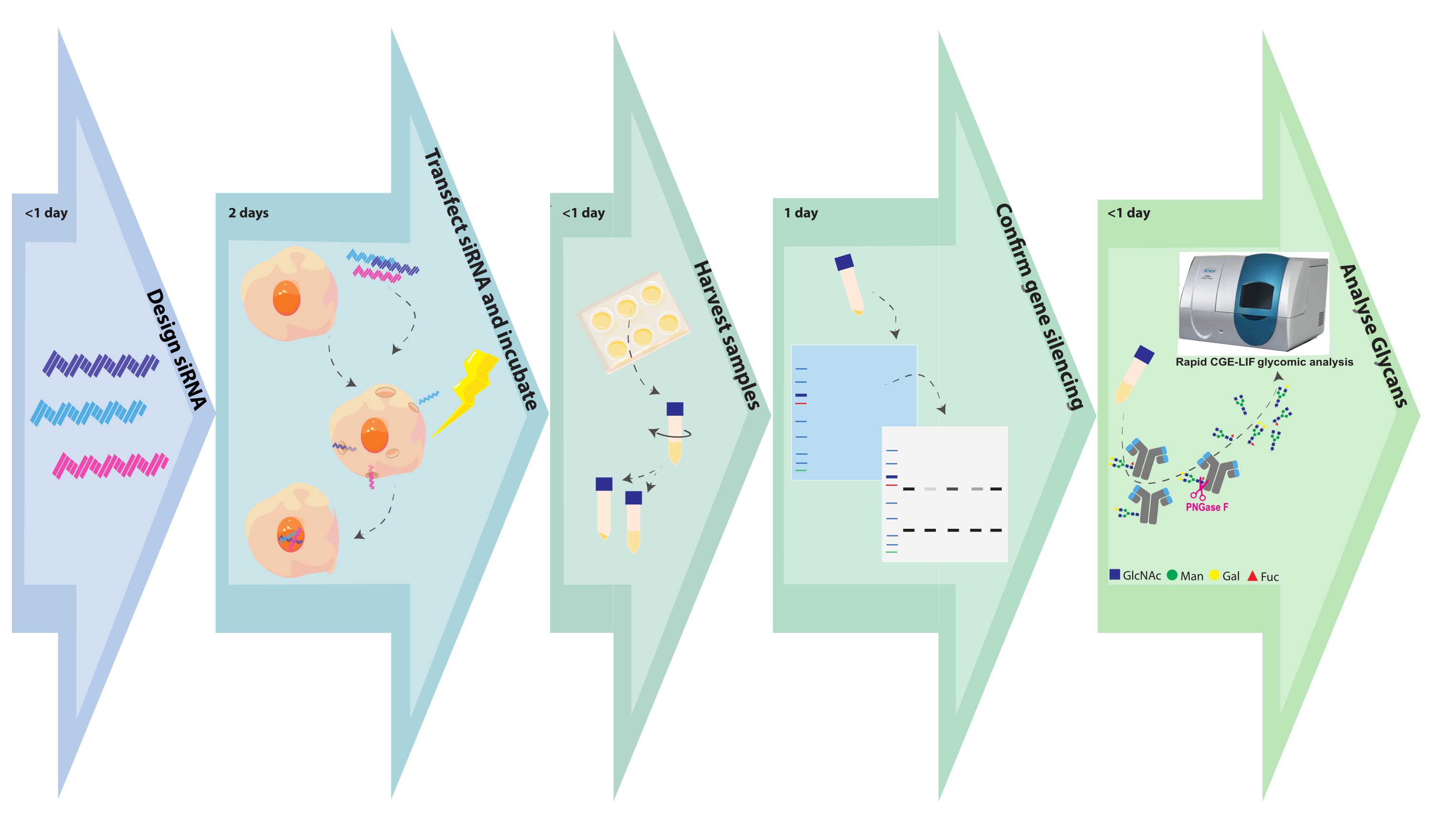
Abbildung 1. Experimentelles Flussdiagramm. Glykoengineering- und Probenanalyseschritte werden mit der zugehörigen Zeit dargestellt, die für jeden Schritt benötigt wird. Das SiRNA-Design dauert einige Stunden, abhängig von der Anzahl der Genziele oder -konstrukte pro Genziel. Die CHO-Zelltransfektion mit siRNA ist in wenigen Stunden abgeschlossen, und transformierte Zellen können 48 Stunden lang wachsen. Zellpellets und Überstände werden innerhalb weniger Stunden geerntet. Zellpellets werden lysiert, und die intrazellulären Proteine werden auf einer SDS-SEITE getrennt und anschließend mit Antikörpern gegen das Zielgen abgetupft und untersucht. Glykane werden aus gereinigten Antikörpern gespalten und mit CGE-LIF analysiert. Diese Assays können jeweils 1 Tag dauern. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 2. Bestätigung der RNA-Interferenz. Western Blot-Nachweis von α-1,6-Fucosyltransferase (FUT8)-Proteinspiegeln in Proben, die mit Fut8 oder Non-Targeting-Kontroll-DsiRNA behandelt wurden. Banden, die FUT8 entsprechen, sind intensiver in der Kontrolle als Fut8-Knockdown-Samples. Der GAPDH-Proteinspiegel wurde ebenfalls bewertet, um die Expression des Zielgens zu normalisieren. Alle Proben wurden aus Experiment G entnommen (siehe Tabelle 2). Nachdruck von Kotidis et al. 52. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}

Abbildung 3. Wirkung von Fut8-Knockdown auf die kumulative IgG-Glykosylierung bei 48 h. Eine Verschiebung der Glykanverteilung wird in Knockdown-Proben nachgewiesen. Insbesondere wird die relative Häufigkeit der wichtigsten kernfucosylierten Strukturen (G0F) reduziert, während die afucosylierten Spezies im Knockdown-Experiment erhöht werden. Die Messungen wurden anhand von Proben aus Experiment G durchgeführt (siehe Tabelle 2). Biologische Verdreifachungen, die für jedes Experiment durchgeführt wurden, wurden nach der Ernte gemischt, um die Belastung durch nachgelagerte Analysen zu reduzieren. Nachdruck von Kotidis et al. 52. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
{kind=link}
| Name des Experiments | Elektroporationsverfahren | DsiRNA-Konzentration (nΜ) | Erntezeit (h) | Lebensfähigkeit (%) | Xv (106 Zellen·mL-1) | IgG-Titer (mg· L-1) | Unterschied in der Kern-Fucosylierung (%) | ||||||
| ExpA_Negative | e1 | 500 | 24 | 98.3 | 4.71 | 122.5 | - | ||||||
| ExpA_Knockdown | e1 | 500 | 24 | 98.3 | 4.9 | 110.3 | 4.08 | ||||||
| ExpB_Negative | e1 | 500 | 48 | 95.6 | 9.55 | 453.3 | - | ||||||
| ExpB_Knockdown | e1 | 500 | 48 | 96.7 | 9.61 | 469 | 5.38 | ||||||
| ExpC_Negative | e2 | 500 | 48 | 96.3 | 9.91 | 449.3 | - | ||||||
| ExpC_Knockdown | e2 | 500 | 48 | 96.7 | 11 | 454.6 | 11.42 | ||||||
| ExpD_Negative | E3 | 500 | 48 | 90.6 | 6.25 | 318.5 | - | ||||||
| ExpD_Knockdown | E3 | 500 | 48 | 89.1 | 6.09 | 311.85 | 9.71 | ||||||
| ExpE_Negative | E4 | 500 | 48 | 91.1 | 7.2 | 380.3 | - | ||||||
| ExpE_Knockdown | E4 | 500 | 48 | 93.3 | 7.79 | 422.8 | 14.7 | ||||||
| ExpF_Negative | e1 | 750 | 48 | 96.2 | 9.7 | 501 | - | ||||||
| ExpF_Knockdown | e1 | 750 | 48 | 95.7 | 9.76 | 504.6 | 9.9 | ||||||
| ExpG_Negative | e1 | 1000 | 48 | 96.1 | 11.1 | 422.6 | - | ||||||
| ExpG_Knockdown | e1 | 1000 | 48 | 95.9 | 9.73 | 499.3 | 17.26 | ||||||
| ExpH_Negative | e1 | 500 | 72 | 94.4 | 14.3 | 925.8 | - | ||||||
| ExpH_Knockdown | e1 | 500 | 72 | 95 | 13.5 | 1018.4 | 7.37 | ||||||
Tabelle 2. Transfektionsoptimierung. Iterative Modifikationen der Elektroporationsmethode, der DsiRNA-Konzentration und der Erntezeit führten zu Veränderungen der Zelllebensfähigkeit, der lebensfähigen Zelldichte, des IgG-Titers zum Erntezeitpunkt und Unterschieden in der Kernfukosylierung. Jedes Experiment verglich den Knockdown und die jeweilige Negativkontrolle, um festzustellen, ob die Modifikation den gewünschten Effekt erzeugt (d.h. eine Abnahme der Fukosylierung). Die Elektroporationseinstellungen waren wie folgt: e1: 1200 V, 0,1 ms, Rechteckwellenform; e2: 1200 V, 2x 0,1 ms, 5 s zwischen den Impulsen, Rechteckwellenform; e3: 150 V, 20 ms, Rechteckwellenform; e4: 250 V, 500 μF, exponentieller Zerfall. Nachdruck von Kotidis et al. 52.
Diskussion
Glykosylierungswege beinhalten ein komplexes metabolisches Netzwerk von Enzymen und akzessorischen Proteinen. Die Sezierung der Funktion von Signalwegsbestandteilen ist entmutigend, wenn sie allein auf konventionelle Knockout- oder Knockin-Gentechnik-Strategien angewiesen ist. Ein alternativer Ansatz besteht darin, Mitglieder eines Pfades vorläufig mit einem transienten Funktionsverlust-Assay zu screenen. Zu diesem Zweck wurden zwei schnelle Protokolle, RNAi- und CGE-LIF-Detektion, kombiniert, um eine effizientere Möglichkeit zur Charakterisierung von Glykosylierungsgenen zu schaffen. Die beschriebene Methode erfordert 5-7 Tage für den Abschluss im Vergleich zu herkömmlichen Methoden, die möglicherweise mehrere Wochen dauern. Darüber hinaus könnten Forschungsumgebungen mit Automatisierungsfunktionen diese Methode nutzen, um mehr Genkandidaten zu screenen, als mit manueller Handhabung möglich ist.
Der Erfolg einer transienten Glykoengineering-Kampagne hängt maßgeblich vom siRNA-Design ab. Benutzerdefinierte DsiRNA-Designs müssen den zuvor beschriebenen Regeln entsprechen, oder der Einfachheit halber können sich Benutzer für kommerziell erhältliche vorgefertigte Sequenzen entscheiden. Wie andere Genmodifikationsstrategien hat RNAi das Potenzial für Off-Target-Effekte. Daher werden die Nutzer ermutigt, unbeabsichtigtes Gen-Targeting mit Berechnungsmethoden zu bewerten41. Experimentelle Designentscheidungen können auch dazu beitragen, Off-Target-Effekte zu begrenzen. Kittler et al. zeigten, dass die gemultiplexte Abgabe von siRNA zu einer Verringerung der Off-Target-Effekte führte42. Obwohl dies kontraintuitiv erscheint, wird vorgeschlagen, dass ein Master-Mix die Konzentration jedes siRNA-Konstrukts reduziert und somit die Möglichkeit eines Off-Target-Gen-Silencings einschränkt. Ein weiterer Vorteil ist, dass die gleichzeitige Transfektion von siRNA-Strukturen, die auf dasselbe Gen abzielen, die Wahrscheinlichkeit einer erfolgreichen RNAi erhöht. Die Verwendung eines Master-Mixes gewährleistet auch die Konsistenz zwischen Proben und Replikaten und beschleunigt den Transfektionsprozess. Nach einem ersten Screening von gemischten siRNA-Konstrukten kann ein weiteres Experiment mit einzelnen Konstrukten durchgeführt werden, um die RNAi-Effizienz jeder Sequenz zu bestimmen. In dieser und anderen Knockdown-Studien wurden bis zu drei siRNA gepoolt und an die Zellenabgegeben 43,44,45. Es kann jedoch wünschenswert sein, mehr als drei siRNA gleichzeitig zu screenen, um effizient auf ein einzelnes Gen oder mehrere Gene abzuzielen. Tatsächlich zeigte eine Studie ein gemultiplextes siRNA-vermitteltes Silencing von bis zu sechs Genen in Konzentrationen, die mit dem Silencing einzelner Gene vergleichbar sind46. Es sind jedoch weitere Studien erforderlich, um die maximale Anzahl von siRNA-Konstrukten zu bestimmen, die in einem Pool verwendet werden können, ohne die Stummschaltungseffizienz zu beeinträchtigen. Die Multiplex-Strategie wurde von Martin et al. vorgeschlagen, um das Tempo der RNAi-Bibliotheks-Screening-Experimente zu verbessern46, und ein ähnliches Konzept könnte sich als nützlich erweisen, um Glykosylierungsgene zu screenen.
Das hierin beschriebene Protokoll dient als Proof-of-Concept mit der Erwartung, dass nachfolgende Experimente durchgeführt werden, um andere Glykosylierungsgene zu validieren. Neue Gene von Interesse können uncharakterisiert oder weniger beliebt sein als Fut8, und primäre Antikörper zum Nachweis von Gen-Silencing können schlecht oder nicht verfügbar sein. In diesem Szenario können alternative Methoden wie die RT-PCR verwendet werden, um das Gen-Silencing47 zu quantifizieren, aber es sollte beachtet werden, dass RT-PCR eher mRNA als Protein nachweist. Wenn Antikörper für Western Blotting verfügbar sind, ist ein häufiges Problem ein schlechter Nachweis oder das Vorhandensein unspezifischer Banden. Anleitungen zur Fehlerbehebung, die Benutzern bei der Lösung häufiger Probleme helfen, sind verfügbar, und diese umfassen in der Regel eine Reihe von Lösungen wie primäre Antikörpertitrierung, alternative Blockierung und Nachweisbedingungen48,49. In dieser Studie erschien FUT8 unerwartet als Doppelband bei ~65 und ~70 kDa. Es ist möglich, dass das ~70 kDa-Band glykosyliertes FUT8 darstellt. Literaturnachweise aus menschlichen Zelllinien beschreiben die O-verknüpfte Glykosylierung bei Thr 56450,51, einer Stelle, die in chinesischen Hamster-Sequenzen konserviert ist, und CHO K1 FUT8-Sequenzen.
Wie bereits erwähnt, beinhalten Glykosylierungswege oft eine komplexe Reihe von Enzymen. Das aktuelle Protokoll wurde unter Verwendung einer monogenen Glykosylierungsreaktion, die von Fut8 kontrolliert wird, entwickelt, optimiert und demonstriert. Daher sind weitere Studien erforderlich, um die Robustheit dieser Methode zu bestätigen, wenn das Zielgen für ein Enzym mit abwechselnder Kinetik und Expressionsniveaus oder einen durch Isoenzyme mit redundanten Funktionen regulierten Weg kodiert.
Zusammengenommen ist die Fähigkeit, Gene schnell zum Schweigen zu bringen und modifizierte IgG-Glykoprofile zu erkennen, ein nützliches Werkzeug bei den Bemühungen um maßgeschneiderte glykotechnische Antikörper. Erkenntnisse aus ähnlichen Kurzzeitstudien können angewendet werden, um stabile glykotechnisch hergestellte Zellen für den Einsatz in Langzeitassays wie der Fed-Batch-Kultur zu erzeugen. Außerhalb des pharmazeutischen Kontextes trägt diese Methode zum Studium der Glykanbiologie bei und hebt die wichtige Funktion von Glykanen bei Entwicklung, Gesundheit und Krankheit hervor.
Offenlegungen
Die Autoren haben nichts offenzulegen.
Danksagungen
PK dankt dem Department of Chemical Engineering, Imperial College London, für sein Stipendium. RD dankt dem U.K. Biotechnology and Biological Sciences Research Council für sein Studium. MM wird vom U.K. Biotechnology and Biological Sciences Research Council finanziert (Grant reference: BB/S006206/1). IAG dankt dem Irish Research Council (Stipendium Nr. GOIPG/2017/1049) und CONACyT (Stipendium Nr. 438330).
Materialien
| Name | Company | Catalog Number | Comments |
| 32 Karat software | SCIEX | contact manufacturer | Software for glycan data acquisition and analysis using the Fast Glycan analysis protocol and separation method. |
| Acetonitrile, HPLC grade | Sigma Aldrich | 34851 | Solvent. |
| Anti-FUT8 antibody | AbCam | ab198741 | Rabbit polycloncal to Fut8. Use this antibody to quantify Fut8 protein expression; replace this antibody if using siRNA targeting a different gene. |
| Anti-GAPDH antibody | AbCam | ab181602 | Rabbit monoclonal to GAPDH. Alternative housekeeping genes exist and might be preferred by the user. |
| BioDrop Spectrophotometer | Biochrom | 80 3006 55 | Instrument used to quantify protein concentration. |
| BLItz | ForteBio | 45 5000 | Instrument. Label-Free Protein Analysis System. |
| BRAND Haemocytometer | Sigma Alrich | BR717810 | Counting chamber device |
| Capillary cartridge | SCIEX | A55625 | Pre-assembled capillary cartridge with window (30 cm total length, 375 µm outer diameter (o.d), x 50 µm inner diameter (i.d). |
| C100HT Glycan analysis—capillary electrophoresis | SCIEX | contact manufacturer | Capillary gel electrophoresis instrument, the CESI 8000 Plus instrument is now used. |
| CD CHO Medium | Thermo Fisher Scientific | 10743029 | Replace this with a culture medium appropriate for the cell line of choice. |
| Centrifuge tubes, 15 mL | Greiner Bio | 188261 | Sterile polypropylene tube. |
| Centrifuge tubes, 50 mL | Greiner Bio | 227270 | Sterile polypropylene tube. |
| CHO IgG | MedImmune | Gift | Chinese Hamster Ovary cells expressing an IgG monoclonal antibody (CHO T127). Created using the GS system. |
| Dulbecco's phosphate-buffered saline (DPBS) | Gibco | 14190144 | 1x PBS, without calcium or magnesium. |
| Erlenmeyer Flasks with Vent Cap, 125 mL | Corning | 431143 | Replace this with a culture vessel suitable for growing the cell line of choice. |
| Erlenmeyer Flasks with Vent Cap, 250 mL | Corning | 431144 | Replace this with a culture vessel suitable for growing the cell line of choice. |
| Fast Glycan Labelling and Analysis kit | SCIEX | B94499PTO | Labels N-glycans with APTS and then uses a magnetic-bead based clean up system to remove excess APTS. |
| Fut8 DsiRNA | IDT | Custom | Custom designed DsiRNA targetting Fut8. |
| Gene Pulser cuvettes, 0.4 cm | Bio-Rad | 1652088 | Electroporation cuvette. |
| Gene Pulser Xcell Eukaryotic System | Bio-Rad | 165 2661 | Insturment. Xcell main unit with Capacitance Extender (CE) Mocdule and ShockPod. |
| Immobilon-FL PVDF membrane | Merck-Millipore | IPFL00010 | Immunoblot transfer membrane, low background. |
| L-Methionine sulfoximine (MSX) | Sigma Aldrich | M5379 | Only necessary for CHO cell lines using the glutamine synthetase (GS) selection system. |
| Kimwipes | Thermo Fisher Scientific | 10623111 | Low-lint, high absorbency and chemically inert wipes. |
| M-PER Mammalian Protein Extraction Reagent | Thermo Fisher Scientific | 78505 | Alternative lysis buffers such as RIPA are also appropriate. |
| Methanol, HPLC grade | Fisher Scientific | 10365710 | Solvent. |
| Microcentrifuge tubes, 1.5 mL | Eppendorf | 616201 | Autoclavable tubes. |
| Mini-PROTEAN Tetra Vertical Electrophoresis Cell system | Bio-Rad | 1658035FC | Instrument. 4-gel capacity, for 1.0 mm thick handcast gels, with Mini Trans-Blot Module and PowerPac HC Power Supply. |
| NC-Slide A8 | ChemoMetec | 942 0003 | 8-chamber slide for use with NucleoCounter NC 250. |
| Negative Control DsiRNA, 5 nmol | IDT | 51 01 14 04 | Non-targeting DsiRNA. |
| Nuclease-free duplex buffer | IDT | 11-01-03-01 | Reconstitution buffer for DsiRNA. |
| NucleoCounter NC-250 | Chemometec | contact manufacturer | Instrument. Automated Cell Analyzer |
| Page-Ruler ladder, 10 to 180 kDa | Thermo Fisher Scientific | 26616 | Mixed blue, orange and green protein standards for SDS PAGE and western blotting. |
| PCR tubes | Greiner Bio | 608281 | Autoclavable tubes for DsiRNA aliqouts and glycan preparation. |
| Pipette filter tips sterilised (10, 200, 1000 µL) | Gibson | F171203, F171503, F171703 | Sterile filter tips to avoid RNA contamination. |
| PNGase F enzyme | New England Biolabs | P0704S | Enzymatic cleavage of glycans from glycoproteins. |
| Polypropylene columns, 1 mL | Qiagen | 34924 | Columns for gravity-flow chromatography. |
| Protease Inhibitor Cocktail | Sigma Aldrich | P8340 | Inhibition of serine, cysteine, aspartic proteases and aminopeptidases |
| Protein-A Agarose Beads | Merck-Millipore | 16 125 | For purification of human, mouse and rabbit immunoglobulins. |
| Protein-A biosensor | ForteBio | 18 5010 | Tips functionalised with Protein A for rapid antibody quantification. |
| RNaseZap | Invitrogen | AM9780 | Removes RNAse contamination. |
| Sample dilutent | ForteBio | 18 1104 | Activate Protein A tips. |
| Serological pipets (5, 10, 25 mL) | Corning | 4487, 4488, 4489 | Used for sterile cell culture tecniques. |
| Sodium cyanoborohydride solution 1 M in THF | Sigma Aldrich | 296813 | Reducing agent. |
| Solution 18 | ChemoMetec | 910-3018 | Staining reagent containing acridine orange (AO) and 4',6-diamidino-2-phenylindole (DAPI) |
| Spin-X Centrifuge Tube Filters | Corning | 8161 | 0.22 µm pore, Cellulose Acetate membrane. |
| Suspension plate with lid, 6-well | Greiner Bio | 657 185 | Hydrophobic culture plate for growth of suspension cultures. |
| Syringe filters, 0.22 μm | Sartorius | 514 7011 | Surfactant-free cellulose acetate (SFCA) |
| Syringes with Luer lock tip, 20 mL | Fisher Scientific | 10569215 | For secure connection with syringe filter. |
| Trypan Blue solution | Gibco | 15250061 | Stains dead and dying cells. |
| Vivaspin 500, 3,000 MWCO | Sartorius | VS0191 | Polyethersulfone |
| WesternBreeze Chromogenic Kit, anti-rabbit | Thermo Fisher Scientific | WB7105 | Western blot detection kit, alternative blocking buffers and antibody diluents can be made by the user using recipes available online. |
Referenzen
- Jefferis, R. Glycosylation as a strategy to improve antibody-based therapeutics. Nature Reviews Drug Discovery. 8 (3), 226-234 (2009).
- Rathore, A. S., Winkle, H. Quality by design for biopharmaceuticals. Nature Biotechnology. 27 (1), 26-34 (2009).
- Eon-Duval, A., Broly, H., Gleixner, R. Quality attributes of recombinant therapeutic proteins: An assessment of impact on safety and efficacy as part of a quality by design development approach. Biotechnology Progress. 28 (3), 608-622 (2012).
- Lalonde, M. -. E., Durocher, Y. Therapeutic glycoprotein production in mammalian cells. Journal of Biotechnology. 251, 128-140 (2017).
- Mimura, Y., et al. Glycosylation engineering of therapeutic IgG antibodies: challenges for the safety, functionality and efficacy. Protein & Cell. 9 (1), 47-62 (2018).
- Walsh, G. Biopharmaceutical benchmarks 2018. Nature Biotechnology. 36 (12), 1136-1145 (2018).
- Zhang, M., Koskie, K., Ross, J. S., Kayser, K. J., Caple, M. V. Enhancing glycoprotein sialylation by targeted gene silencing in mammalian cells. Biotechnology and Bioengineering. , (2010).
- Kanda, Y., et al. et al. of a GDP-mannose 4,6-dehydratase (GMD) knockout host cell line: a new strategy for generating completely non-fucosylated recombinant therapeutics. Journal of Biotechnology. 130 (3), 300-310 (2007).
- Umaña, P., Jean-Mairet, J., Moudry, R., Amstutz, H., Bailey, J. E. Engineered glycoforms of an antineuroblastoma IgG1 with optimized antibody-dependent cellular cytotoxic activity. Nature Biotechnology. 17 (2), 176-180 (1999).
- Xu, X., et al. et al. genomic sequence of the Chinese hamster ovary (CHO)-K1 cell line. Nature Biotechnology. 29 (8), 735-741 (2011).
- Chan, K. F., et al. et al. of GDP-fucose transporter gene (Slc35c1) in CHO cells by ZFNs, TALENs and CRISPR-Cas9 for production of fucose-free antibodies. Biotechnology Journal. 11 (3), 399-414 (2016).
- Zhang, P., et al. Identification of functional elements of the GDP-fucose transporter SLC35C1 using a novel Chinese hamster ovary mutant. Glycobiology. 22 (7), 897-911 (2012).
- Amann, T., et al. Genetic engineering approaches to improve posttranslational modification of biopharmaceuticals in different production platforms. Biotechnology and Bioengineering. 116 (10), 2778-2796 (2019).
- Karottki, K. J., et al. Awakening dormant glycosyltransferases in CHO cells with CRISPRa. Biotechnology and Bioengineering. 117 (2), 593-598 (2020).
- Fire, A., et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 391 (6669), 806-811 (1998).
- Hammond, S. M., Bernstein, E., Beach, D., Hannon, G. J. An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells. Nature. 404 (6775), 293-296 (2000).
- Bernstein, E., Caudy, A. A., Hammond, S. M., Hannon, G. J. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature. 409 (6818), 363-366 (2001).
- Shields, R. L., et al. et al. of fucose on human IgG1 N-linked oligosaccharide improves binding to human FcγRIII and antibody-dependent cellular toxicity. Journal of Biological Chemistry. 277 (30), 26733-26740 (2002).
- Shinkawa, T., et al. et al. Absence of fucose but not the presence of galactose or bisecting N-acetylglucosamine of human IgG1 complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity. Journal of Biological Chemistry. 278 (5), 3466-3473 (2003).
- Mori, K., et al. Engineering Chinese hamster ovary cells to maximize effector function of produced antibodies using FUT8 siRNA. Biotechnology and Bioengineering. 88 (7), 901-908 (2004).
- Imai-Nishiya, H., et al. Double knockdown of alpha1,6-fucosyltransferase (FUT8) and GDP-mannose 4,6-dehydratase (GMD) in antibody-producing cells: a new strategy for generating fully non-fucosylated therapeutic antibodies with enhanced ADCC. BMC Biotechnology. 7, 84 (2007).
- Beuger, V., et al. et al. silencing of fucosyltransferase 8 in Chinese-hamster ovary cells for the production of antibodies with enhanced antibody immune effector function. Biotechnology and Applied Biochemistry. 53 (1), 31 (2009).
- Templeton, N., Dean, J., Reddy, P., Young, J. D. Peak antibody production is associated with increased oxidative metabolism in an industrially relevant fed-batch CHO cell culture. Biotechnology and Bioengineering. 110 (7), 2013-2024 (2013).
- Hussain, H., et al. A comparative analysis of recombinant Fab and full-length antibody production in Chinese hamster ovary cells. Biotechnology and Bioengineering. 118 (12), 4815-4828 (2021).
- Shimoyama, H., et al. Partial silencing of fucosyltransferase 8 gene expression inhibits proliferation of Ishikawa cells, a cell line of endometrial cancer. Biochemistry and Biophysics Reports. 22, 100740 (2020).
- Tummala, S., et al. Evaluation of exogenous siRNA addition as a metabolic engineering tool for modifying biopharmaceuticals. Biotechnology Progress. 29 (2), 415-424 (2013).
- Carillo, S., et al. Comparing different domains of analysis for the characterisation of N-glycans on monoclonal antibodies. Journal of Pharmaceutical Analysis. 10 (1), 23-34 (2020).
- Reusch, D., et al. Comparison of methods for the analysis of therapeutic immunoglobulin G Fc-glycosylation profiles-Part 2: Mass spectrometric methods. mAbs. 7 (4), 732-742 (2015).
- Donini, R., Haslam, S. M., Kontoravdi, C. Glycoengineering Chinese hamster ovary cells: a short history. Biochemical Society Transactions. 49 (2), 915-931 (2021).
- Szigeti, M., Chapman, J., Borza, B., Guttman, A. Quantitative assessment of mAb Fc glycosylation of CQA importance by capillary electrophoresis. ELECTROPHORESIS. 39 (18), 2340-2343 (2018).
- Ruhaak, L. R., et al. Optimized workflow for preparation of APTS-labeled N-glycans allowing high-throughput analysis of human plasma glycomes using 48-channel multiplexed CGE-LIF. Journal of Proteome Research. 9 (12), 6655-6664 (2010).
- Patenaude, A. -. M., et al. N-glycosylation analysis of mouse immunoglobulin G isolated from dried blood spots. Electrophoresis. 42 (24), 2615-2618 (2021).
- Karst, D. J., et al. Modulation and modeling of monoclonal antibody N-linked glycosylation in mammalian cell perfusion reactors. Biotechnology and Bioengineering. 114 (9), 1978-1990 (2017).
- Dadouch, M., Ladner, Y., Perrin, C. Analysis of monoclonal antibodies by capillary electrophoresis: sample preparation, separation, and detection. Separations. 8 (1), 4 (2021).
- Fakhr, E., Zare, F., Teimoori-Toolabi, L. Precise and efficient siRNA design: a key point in competent gene silencing. Cancer Gene Therapy. 23 (4), 73-82 (2016).
- Birmingham, A., et al. A protocol for designing siRNAs with high functionality and specificity. Nature Protocols. 2 (9), 2068-2078 (2007).
- Makrydaki, E., et al. . Immobilised enzyme cascade for targeted glycosylation. , (2022).
- . Bulletin_6201.pdf Available from: https://www.bio-rad.com/webroot/web/pdf/lsr/literature/Bulletin_6201.pdf (2022)
- . Bulletin_6376.pdf Available from: https://www.bio-rad.com/webroot/web/pdf/lsr/literature/Bulletin_6201.pdf (2022)
- . Dot Blot Analysis Available from: https://imagej.nih.gov/ij/docs/examples/dot-blot/index.html (2022)
- Yilmazel, B., et al. Online GESS: prediction of miRNA-like off-target effects in large-scale RNAi screen data by seed region analysis. BMC Bioinformatics. 15 (1), 192 (2014).
- Kittler, R., et al. Genome-wide resources of endoribonuclease-prepared short interfering RNAs for specific loss-of-function studies. Nature Methods. 4 (4), 337-344 (2007).
- Stec, E., et al. A multiplexed siRNA screening strategy to identify genes in the PARP pathway. Journal of Biomolecular Screening. 17 (10), 1316-1328 (2012).
- Brown, J. M., et al. Ligand conjugated multimeric siRNAs enable enhanced uptake and multiplexed gene silencing. Nucleic Acid Therapeutics. 29 (5), 231-244 (2019).
- Parsons, B. D., Schindler, A., Evans, D. H., Foley, E. A direct phenotypic comparison of siRNA pools and multiple individual duplexes in a functional assay. PLoS One. 4 (12), 8471 (2009).
- Martin, S. E., et al. Multiplexing siRNAs to compress RNAi-based screen size in human cells. Nucleic Acids Research. 35 (8), 57 (2007).
- Popp, O., Moser, S., Zielonka, J., Rüger, P., Hansen, S., Plöttner, O. Development of a pre-glycoengineered CHO-K1 host cell line for the expression of antibodies with enhanced Fc mediated effector function. mAbs. 10 (2), 290-303 (2018).
- Yang, P. -. C., Mahmood, T. Western blot: Technique, theory, and trouble shooting. North American Journal of Medical Sciences. 4 (9), 429 (2012).
- Troubleshooting Western Blots with the Western Blot Doctor. Bio-Rad Laboratories Available from: https://www.bio-rad.com/en-uk/applications-technologies/troubleshooting-western-blots-with-western-blot-doctor?ID=MIW4HR15 (2022)
- Steentoft, C., et al. Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology. EMBO. 32 (10), 1478-1488 (2013).
- . FUT8 - Alpha-(1,6)-fucosyltransferase - Proteomics Available from: https://www.nextprot.org/entry/NX_Q9BYC5/proteomics (2022)
- Kotidis, P., et al. Rapid antibody glycoengineering in CHO cells via RNA interference and CGE-LIF N-glycomics. Methods in Molecular Biology. 2370, 147-167 (2022).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenWeitere Artikel entdecken
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten