
Method Article
チャイニーズハムスター卵巣細胞における迅速な抗体糖工学
* これらの著者は同等に貢献しました
要約
抗体のグリコシル化パターンは、その臨床的性能を決定し、したがって、グリコシル化を制御するための産業的および学術的努力は持続する。典型的な糖工学キャンペーンは時間と労力を要するため、一過性サイレンシングを使用してグリコシル化遺伝子の影響を特徴付けるための迅速なプロトコルの生成が有用であることが証明されるであろう。
要約
組換えモノクローナル抗体は、特定の分子標的に結合し、続いて、免疫応答を誘導するか、または他のリガンドの結合を阻害する。しかしながら、モノクローナル抗体の機能性および半減期は、宿主特異的グリコシル化の種類および分布によって低下し得る。優れた抗体を産生する試みは、糖最適化抗体を合成する遺伝子組み換え生産細胞の開発を鼓舞してきた。糖工学は、典型的には、クラスター化された規則的に間隔をあけた短い回文反復(CRISPR)関連タンパク質9などの方法を用いて、安定なノックアウトまたはノックイン細胞株の生成を必要とする。次いで、操作された細胞によって産生されたモノクローナル抗体は、質量分析法を用いて特徴付けられ、所望のグリコプロファイルが得られたかどうかを決定する。この戦略は時間がかかり、技術的に困難であり、専門家が必要です。したがって、遺伝子グリコエンジニアリングおよびグリカン検出のための合理化されたプロトコルを利用する代替戦略は、最適な抗体に向けた努力を支援する可能性がある。この概念実証研究では、IgG産生チャイニーズハムスター卵巣細胞が糖工学を最適化するための理想的な宿主として役立った。 Fut8 遺伝子を標的とする短い干渉RNAをチャイニーズハムスター卵巣細胞に送達し、その結果生じるFUT8タンパク質発現の変化を定量した。結果は、この方法によるノックダウンが効率的であり、FUT8の〜60%の減少をもたらしたことを示している。抗体グリコプロファイルの相補的解析は、迅速でありながら高感度な技術(キャピラリーゲル電気泳動およびレーザー誘導蛍光検出)を用いて実施した。すべてのノックダウン実験は、アフコシル化グリカンの増加を示した。しかし、この研究で達成された最大のシフトは約20%でした。このプロトコルは、 インシリコ 設計ツール、商業的に合成された遺伝子標的化試薬、および広範な事前経験を必要としない迅速な定量アッセイを利用することにより、糖工学の取り組みを簡素化します。したがって、このプロトコルによって提供される時間効率は、新しい遺伝子標的の調査に役立つ可能性がある。
概要
N結合型グリコシル化は、オリゴ糖部分がAsn残基に共有結合で結合される酵素プロセスである。de novoタンパク質合成とは異なり、グリカン合成は、タンパク質の不均一なグリコシル化をもたらす非テンプレート化反応である。グリカンの構造、組成、および分布は、タンパク質の立体構造および機能に影響を及ぼし得る。実際、免疫グロブリンG(IgG)の結晶化可能なフラグメント(Fc)領域におけるN-グリコシル化は、抗体1の治療有効性、免疫原性、および半減期を調節する。このように、組換えバイオ治療用タンパク質製品の開発のための設計による品質(QbD)パラダイムは、グリコシル化を重要な品質属性(CQA)として自然に識別します2,3。哺乳動物細胞は、細菌、酵母、昆虫、または植物細胞よりも本質的にヒト様のグリコシル化パターンをより密接に産生するため、しばしば好ましい発現系である。さらに、チャイニーズハムスター卵巣(CHO)細胞は、ヒトウイルス感染に耐性があり、高力価で産物を分泌し、浮遊培養で高い生細胞密度に増殖させることができるため、他の哺乳動物細胞株よりも選択される4。グリカン形成に関して、非CHOマウス産生細胞は、モノクローナル抗体(mAbs)5の安全な使用に影響を与える免疫原性グリカン(α(1-3)結合ガラクトース[α(1-3)-Gal]およびN-グリコリルノイラミン酸[NeuGc])を生成する。これらの利点により、CHO細胞は最も重要な発現系となり、2014年から2018年の間に新しいバイオ医薬品の80%以上の生産を担っています6。しかしながら、鋳型非依存性グリコシル化は、一連のグリコフォームを有するCHO由来の生物治療薬をもたらす保存された機構である。
バイオ医薬品開発戦略は、遺伝子工学によってCHO細胞の不均一性を制御することを目指しています。いくつかの文献例には、シアリダーゼ(Neu1、Neu3)7のノックダウン、GDP-マンノース4,6-デヒドラターゼ(GMD)ノックアウト8、および糖転移酵素(GnTIII)9の過剰発現が含まれる。CHOゲノム10のような公的に入手可能な資源と、転写アクチベーター様エフェクターヌクレアーゼ(TALEN)、ジンクフィンガーヌクレアーゼ(ZFN)、およびクラスター化された規則的に間隔をあけた短い回文反復(CRISPR)関連タンパク質9(CRISPR-Cas9)などの遺伝子工学ツールの継続的な開発の組み合わせにより、糖工学の進歩が可能である11、12、13、14.これらのツールは、典型的には、プラスミドDNAまたは精製リボヌクレオタンパク質(RNP)複合体としてCHO細胞に送達される。逆に、RNA干渉(RNAi)は、その最も単純な形態では、精製された短い干渉RNA(siRNA)オリゴヌクレオチドの送達のみを必要とする遺伝子工学技術である。内因性タンパク質は二本鎖siRNAを一本鎖に加工し、ヌクレアーゼであるRNA誘導サイレンシング複合体(RISC)はsiRNAと複合体を形成して標的mRNA配列を切断する15、16、17。この方法による遺伝子サイレンシングは、RNA不安定性のために一過性であるが、本明細書における調査は、迅速なスクリーニングを支援するためにこの特徴を利用する。
現在の研究のために選択されたモデル酵素であるα1,6-フコース転移酵素(FUT8)は、α-1,6コア結合L-フコース(Fuc)を有するN-グリカンを生成する。この修飾は、市販の抗体の研究によって証明されるように、抗体依存性細胞傷害(ADCC)活性の主要な決定因子である。コアフコシル化がない場合、リツキシマブ(抗CD20 IgG1)は、FcgRIIIa結合を改善することによってADCCを50倍に増加させ、トラスツズマブ(抗Her2 IgG1)中のADCCを増加させる18,19。したがって、コアフコシル化は、この表現型を逆転させる努力を正当化するmAbsの望ましくない特徴と考えられている。ADCC 20、21、22の増加を伴うsiRNAを用いたFut8遺伝子ターゲティングの成功例があるが、これらの例はプラスミドDNA上にコードされるFut8 siRNAを送達する。このような実験は、プラスミドDNAがsiRNA合成の鋳型として機能するように、安定した遺伝子サイレンシングを生成する。これにより、細胞は、細胞内RNaseおよびホスファターゼによって分解されるsiRNA分子を補充することができる。逆に、外因性の合成siRNAの送達は、siRNAが細胞内テンプレートの欠如のために補充できないため、一過性の遺伝子サイレンシングのみを可能にする。したがって、ユーザーは、実験計画がプラスミド由来または合成siRNAと互換性があるかどうかを検討する必要があります。例えば、ピークmAb産生に焦点を当てた研究は、典型的には培養23、24の6日目に、ピーク発現の数日前に細胞に送達され得る合成siRNAを選択し得る。合成siRNAを用いた一過性アプローチの利点には、生産を外部委託する能力と、プラスミド中の構築物を生成するのにかかる時間のほんの一部で複数のsiRNA構築物を生成できるという事実が含まれる。さらに、合成siRNAは、FUT8タンパク質発現25を減少させ、FCgRIIIa結合およびADCC26の増加を伴うアフコシル化IgGsを生じるのに十分であるFut8遺伝子サイレンシングの文献例によって証明されるように、有効である。
この糖工学プロトコールの成功は、Fcグリコシル化の程度によって決定された。質量分析は、通常、グリコミック分析に最適な方法です。しかし、キャピラリーゲル電気泳動およびレーザー誘起蛍光検出(CGE-LIF)は、精製されたIgGのグリコプロファイルの解決に完全に適しており、より迅速でシンプルであるという利点を有する。質量分析プロトコルは、適切なクロマトグラフィーおよび誘導体化方法、イオン化源、および質量分析器27、28、29を組み合わせる必要があります。訓練を受けた専門家を必要とすることに加えて、質量分析プロトコルは時間がかかり、方法の多様性は異なるセットアップのラボ間でデータを比較することを困難にします。バイオ医薬品の文脈では、CGE-LIFは抗体グリコプロファイルの十分な詳細を提供することができ、ハイスループット法のために容易に拡張可能な高感度な方法である。存在量が少なく、特性が不十分な糖タンパク質との高度に複雑な混合物の場合、質量分析の利点が残る可能性があります。しかし、CGE-LIFベースのN-グリカン分析によってもたらされる高解像度で高感度のmAb分析は、この方法を試行する理論的根拠として役立ちます。さらに、サンプル調製と分析はわずか数時間30で完了します。最近の研究は、CGE-LIFがヒト血漿31、マウス32、およびCHO IgGs33に由来するグリカンをモニターするために使用できることを示している。これらの研究は、高スループットのサンプル分析と少量のサンプルにCGE-LIFを使用することを強調しています。
CGE-LIF法には、考慮すべき制限があります。コストは、グリカン分析のためのこのデバイスおよび他のデバイスの使用に対する大きな障壁である。しかし、これらのコストは現場では典型的であり、CGE-LIFは費用対効果の高い選択肢であると考えられている34。予算の少ないラボでは、機械をリースしたり、サンプル分析を外部委託したりする方が現実的かもしれません。分析方法のもう1つの考慮事項は、再現性です。CGE−LIFの評価は、異なる日にアッセイされた同じサンプルの48反復を用いて実施した。キャピラリーあたりの相対標準偏差を、バッチ内およびバッチ間の再現性について決定した。反復のバッチ内比較では、6.2%の相対標準偏差を有することが判明し、毛細管性能が均一ではないことを示している。さらに、バッチ間データの比較は、15.8%31の相対標準偏差を示し、毛細管性能が経時的に変化することを示している。特定された運用上の欠点は、異なる機械と独自の試薬を使用する現在の研究では当てはまらない可能性があります。ユーザーが社内プロトコルを開発するつもりなら、Ruhaakらによる研究を検討する価値があるだろう 。図31において、CGE-LIF用の試薬を注意深く評価した。そのようにして、試料注入用の試薬(ハイジホルムアミドおよびDMSO)、グリカン標識(NaBH3CNまたは2−ピコリンボラン)31、およびその他が最適化されている。
この研究は、直接RNAiの迅速さと下流のグリコミック分析を組み合わせた、時間効率の良いグリコエンジニアリングプロトコルを提示します。この方法論は、上記で概説した理由により、 Fut8 遺伝子を標的として用いて例示される。
プロトコル
1. DsiRNAの設計と再構成
- Safari、Firefox、または Microsoft Edge を使用して、IDT の Web サイト (https://eu.idtdna.com) にアクセスします。ホームページから、[製品とサービス ] タブを選択し、その後に RNA 干渉を選択します。 Fut8遺伝子 を標的とするカスタムダイサー基質(DsiRNA)コンストラクトを生成するには、 デザインツール を選択してから カスタムDsiRNAを生成 タブを選択します。
- Fut8アクセッション番号を入力するか、開始する遺伝子配列を手動で入力します。この研究では、「CHO-K1」および「チャイニーズハムスター」の両方のエントリから提案されたFut8配列を https://chogenome.org から取得し、DsiRNAの生成に使用した。ほとんどの遺伝子にはいくつかの転写バリアントがあり、DsiRNAが現在のアセンブリで報告されたすべてのバリアントで活性であることを確認することが重要です。
- CHO細胞ゲノムは現在、IDTのウェブサイトで「参照ゲノム」として利用できないため、「手動BLAST」検索を実行するオプションを選択します。このステップでは、カスタム DsiRNA 配列と目的のゲノムとの間の一致を検索します。
- 「 検索 」をクリックして、DsiRNA 配列を生成します。
- 次に、税番号:10029(CHO細胞株およびチャイニーズハムスター)の「手動BLAST」機能を使用して、各DsiRNAの特異性を評価します。クエリカバレッジの結果は、DsiRNA構築物がFut8(Fut8転写変異体を含む)に100%一致することを示していますが、意図しない標的に対する相補性は≤76%35です。
- DsiRNAが Fut8 を特異的に標的としていることを確認し、GC含量が30%〜50%であることを確認します。低いGC含量は弱く非特異的な結合と関連しており、一方、GC含量が高いとsiRNAの巻き戻しおよびRISC複合体への装填が阻害される36。
- トランスフェクション研究で使用するために3つのDsiRNAを選択し(表1)、それぞれが Fut8 遺伝子の異なる位置を標的とする。コントロール実験のために、事前に設計された非ターゲティングまたはスクランブルされたDsiRNAをIDT社から購入してください。
- 到着したら、DsiRNAを13,300 x gで1分間遠心分離してペレットにし、チューブを開きます。凍結乾燥した各DsiRNAをヌクレアーゼ非含有二本鎖緩衝液(IDT社提供)中で100μMストック溶液に再構成する。例えば、2 nmolのDsiRNAに20 μLの緩衝液を加えて、100 μMストックを得る。
- 十分な混合を確実にするために、オービタルシェーカー(50rpm、16mm軌道)上で穏やかに振とうしながら、ストック溶液を室温で30分間インキュベートする。
- Fut8 を標的とする各 DsiRNA 構築物 20 μL (各 DsiRNA の 100 μM) を組み合わせてマスターミックスを作成します。アリコートを用意し、-20°Cで保存する。
| DsiRNAターゲット | 順序 | GC (%) | ||
| 構造 A | 5' ガガアガアガアカグカウアアアック 3' | 36% | ||
| 5' グァウウガクグウクウクウク 3' | ||||
| 構造 B | 5' アガアウガガウガウガウガウグウウウウウククト 3' | 32% | ||
| 5' AAGGAAAAACAUCCAUUCAUUCAUUCUGA 3' | ||||
| 構造 C | 5' アガガガアウアガアカガアカアウアック 3' | 32% | ||
| 5' グアウウガクグウクウクアウクウクウククウ | ||||
表 1. Fut8 ノックダウンに使用したDsiRNA配列。 チャイニーズハムスターおよびCHO K1細胞ゲノム中の Fut8 を標的とするIDTによって生成された配列。各構築物のセンス配列およびアンチセンス配列が(それぞれ)示され、各構造のGC含量が表示される。Kotidisらより転載。52。
2. DsiRNAトランスフェクション
- IgGモノクローナル抗体37 を発現するCHO細胞を目的の細胞株に適した培養条件を用いて復活させる。
注:この研究で使用された細胞は、内因性GSがL-メチオニンスルホキシミン(MSX)によって阻害され、希少な高生産クローンの選択を強化するグルタミン合成酵素(GS)システムを使用して生成された。したがって、MSXは、選択した細胞株で必要な場合にのみ使用してください。- 細胞のバイアルを37°Cに設定した水浴中で2〜3分間解凍する。 バイアルの外側を70%(v/v)エタノールで清掃し、クラスIIバイオセーフティキャビネットですべての作業を続行します。
- 細胞懸濁液を、選択した細胞株に適した9mLの予備加温培地を含む15mL遠沈管に移す。100 x gで5分間遠心分離することにより細胞をペレット化する。
- 細胞ペレットを乱すことなく、培地を慎重に取り出して廃棄する。次いで、細胞ペレットを予め加温した培地10mLに再懸濁し、計数のためにアリコートを取る。
- 血球計数器で計数する場合はトリパンブルーで細胞を染色し、自動細胞カウンターを使用する場合は生細胞と死細胞を区別するために適切な染色を行います。
- いずれかの列挙方法に従って、適量の細胞懸濁液を、30 mL の培地中 3 x 10 5 cells·mL-1 の生細胞密度で125 mL の三角振とうフラスコに移します (オプションで 50 μM MSX を補充します)。
- 細胞を36.5°Cに設定したインキュベーターに移し、5%CO2 、150rpm(16mm軌道)で振とうプラットフォーム上に置いた。
- 250 mL 三角振とうフラスコに 2 x 105 cells·mL-1 の播種密度で 50 mL の作業容量で 3 ~ 4 日ごとに細胞を継代します。MSXを使用する場合は、最初の継代後に補充を中止してください。
- 継代細胞2xは融解に加えて行う。
- トランスフェクション
- 細胞密度を評価し、細胞生存率が90%より大きいことを確認する。
- 汚染を避けるために、バイオセーフティキャビネットとすべての機器を70%(v/v)エタノールとRNase阻害剤溶液で慎重に清掃してください。
- ペレット細胞を100 x gで5分間、予温した培地に再懸濁して、5 x106 細胞・mL-1の生細胞密度にする。
- DsiRNAマスターミックスまたはコントロールの8 μL(1 μMに相当)を滅菌(0.4 cm)エレクトロポレーションキュベットに移す。次いで、800μLの細胞懸濁液(4 x106 細胞に相当)を同じキュベットに移し、両方の成分が混合されていることを確認する。
- 次のパルス条件を提供します:1200 V、0.1 ms、方形波形。
- 細胞懸濁液をキュベットから6ウェルプレートの1ウェルに移し、泡状物質を避けるように注意する。細胞をインキュベーター(36.5°C、5%CO2)で10分間振盪することなく回収する。
- 800 μLの予備加温培地を加えて1ウェルあたり1.6 mLの最終容量を作り、トランスフェクトした細胞をインキュベーターに戻し、150 rpm(16 mm軌道)で振とうしながら増殖させます(36.5°C、5%CO2)。
- トランスフェクション後48時間で上清および細胞を回収する。
停止点:細胞培養上清を-20°Cに保持することができる;ただし、タンパク質分解を避けるために、細胞ペレット収集直後に細胞溶解を行うことをお勧めします。上清はステップ3、ステップ4、およびステップ5で使用され、細胞ペレットはステップ6で使用される。
3. IgG の定量と精製
- 生体層干渉法または選択した別の方法を使用してIgG濃度を測定します。
- タンパク質Aバイオセンサーチップをサンプル希釈液に10〜30分間水和させる。その間、細胞懸濁液を15 mL遠沈管に移し、細胞を100 x gで5分間ペレット化するか、0.45 μmニトロセルロースフィルターでろ過して細胞を除去します。
- ペレットを乱すことなく、IgGを含む単離された上清を清浄な遠沈管に注意深く移す。
- バイオレイヤー干渉計には、シェーカー速度、2200rpmの設定を使用します。実行時間、60秒。これらのパラメータは、すべてのサンプルとコントロールを測定するために使用されます。
- バイオセンサーチップが水和したら、IgG 標準 (各濃度の 4 μL) を使用して標準曲線を作成します。
- 4 μLの細胞上清を装填してサンプル濃度を定量する。各サンプル間の糸くずの出ないワイプで装置を清掃します。
- 標準曲線を未知サンプルにリンクして、未知サンプルの結合率を補間します。データを保存し、csv または PDF ファイルとしてエクスポートします。
- IgG 精製
- 0.2 Mグリシン(pH 2.5)を含む溶出バッファーを調製し、0.22 μmフィルターを通して滅菌します。
- 単離された上清のフィルター1 mLを、上清全体が流れるまで0.22 μm微量遠心フィルターチューブを用いて室温で分析した。
- プロテインAアガロースビーズ150〜200 μLを100 x gの1.5 mL遠沈管に3分間ペレット化し、上清を廃棄する。次いで、プロテインAビーズを150〜200μLの細胞培養培地で洗浄し、遠心分離を繰り返す。上清を捨てる。
- 平衡化したプロテインAビーズを、ステップ3.2.3から調製した上清の1mLで再懸濁する。1mLポリプロピレンチューブにロードします。ポリプロピレンチューブをロータリーミキサー(15mm軌道)に貼り付け、室温で30〜90分間、または4°Cで一晩回転させる。
- インキュベーションが完了したら、フロースルーを収集し、ビーズを1mLの1x PBSで洗浄して、結合していないタンパク質を除去します。洗浄画分も集めてください。
- ポリプロピレンカラムに溶出バッファー 3 mL を加えることにより、プロテイン A ビーズから IgG を溶出します。カラムから排出される連続画分 (それぞれ 500 μL) を標識チューブに集めます。
- オプション: 精製抗体を保存する場合は、1 M Tris pH 9.5 を 25 μL 加えて溶出バッファーを中和します。抗体がバッファー交換などによって直ちに処理される場合は、この手順をスキップしてください。
注:精製のすべての部分(フロースルー、洗浄、およびすべての溶出画分)は、標的タンパク質が失われないように保管する必要があります。これらのサンプルは、精製が成功しなかった場合のトラブルシューティングにも役立ちます。 - 下流処理には最初の溶出(溶出A)を使用しますが、将来必要になる場合は他のすべての画分を保持します。
4. バッファー交換とサンプル濃度
- IgG 溶出バッファーを 1x PBS と交換します。
- 溶出 A を 3 kDa 分子量の分画遠心濃縮器にロードします。適切なMWCOカラムを選択する際には、製造元のガイドを参照してください。この実験では、孔径が小さいほど、分泌タンパク質の最大の保持が保証されますが、遠心分離時間も長くなります。
- 13,300 x gで4°Cで40〜50分間遠心分離する。 遠心分離は、残留容量が50μL以下になると完了します。
- フロースルーを破棄し、500 μLのプレチルド(4°C)1x PBSを加えて残留上清を希釈する。同じ条件(13,300 x g、4°Cで40-50分間)を用いてサンプルを再び遠心分離し、50 μLの残留上清が残るまで遠心分離した。
- 500 μLのプレチルド(4°C)1x PBSを加えて残留上清を希釈し、再度遠心分離処理を繰り返して上清の100倍希釈液を得た。
- 上清を終濃度〜2.5g·L-1を 40 μL以下で、糖鎖分析法との相溶性を確保した。
注:グリカン分析には100μgをロードする必要があります。
停止点:濃縮されたIgG(1x PBS中)は−20°Cで保存し、グリカン分析の前に解凍することができます。
5. 糖鎖分析
- 製造元の指示に従ってキャピラリーゲル電気泳動を用いてグリカン分析を行う。
- 磁気ビーズ溶液200 μLを0.2 mL PCRチューブに移し、磁気スタンドの上に置き、ビーズを上清から分離した。
- 上澄み液を慎重に取り除き、磁気スタンドからチューブを取り外します。精製されたタンパク質サンプルと渦を加えて、完全な混合を確実にします。
- 変性緩衝液(付属)をサンプルチューブに加え、60°Cで8分間インキュベートした。 最適な反応性能を得るために、サンプルチューブを開いたままにしておきます。
- PNGase F(サンプルあたり500単位)を加え、60°Cで20分間インキュベートして、精製抗体からグリカンを切断する。
- N-グリカンの放出に続いて、サンプルチューブを閉じ、渦巻きする。アセトニトリル、ボルテックスを加え、室温で1分間インキュベートした。
- サンプルチューブを磁気スタンドに置き、ビーズを溶液から分離します。ピペットを使用して、ビーズに触れることなく上清を慎重に取り除きます。
- ヒュームフード内で、フルオロフォアを含むグリカン標識溶液をサンプルに加える。ボルテックスは十分な混合を確実にし、60°Cで20分間インキュベートする(蓋を開ける)。
- サンプル3xをアセトニトリルで洗浄し、過剰な色素を除去した。次いで、標識グリカンをDDI水(供給)に溶出させる。
- 磁気スタンドにサンプル管を置き、精製および標識されたグリカンを含む上清からビーズを分離する。
- グルコースはしご標準、ブラケット標準、およびサンプルを準備し、指定されたトレイ位置にロードします。糖鎖分析プロトコルを実行します。
- 適切なソフトウェアを使用して、サンプル中に存在するグリカンを分析および同定します。
メモ: ヒートブロック内の温度が正確であることを確認して、効率的なインキュベーションを行います。
6. ウェスタンブロット
- 抗α-1,6-フコシルトランスフェラーゼ抗体を用いたウェスタンブロット分析を用いてノックダウン効率を定量する。ポリアクリルアミドゲルおよび緩衝液レシピは、商業的供給業者から入手可能である38,39。
- トランスフェクション後 48 時間で、血球計数器または自動セルカウンターを使用して細胞をカウントし、5 x106 細胞に相当する細胞懸濁液の体積を決定します。
- 各実験条件からの適量の細胞懸濁液を滅菌1.5mL遠沈管に移し、13,200 x gで4°Cで10分間ペレット化する。 上清を捨てる。
- 1%v/vプロテアーゼ阻害剤カクテルを含む200 μLの溶解バッファー(哺乳動物細胞からのタンパク質の抽出に適している)で室温で10分間細胞を溶解する。インキュベーション中に混合物を穏やかに振る(50rpm、16mm軌道)。
- 細胞破片を除去するには、溶解液を13,200 x gで10分間遠心分離し、次いで、除去した溶解物を滅菌1.5mLチューブに移す。
- 各溶解液のタンパク質濃度を280nmで分光光度計を用いて測定する。続いて、同じタンパク質濃度となるように調整した各試料のアリコートを調製する。
- DTT-SDSサンプルローディング色素中で100°Cで10~15分間インキュベートすることによりタンパク質サンプルを変性させる;最終染料濃度は1xです。
- 15 μL の変性サンプルと 5 μL のプレステインタンパク質ラダーを、効率的な分離のために 12.5% の分割ゲルを含む SDS-PAGE ゲルにロードします。サンプルをゲルあたり 25 mA で 90 分間、または色素の前面がゲルの端に達するまで実行します。
- ゲルをカセットから慎重に取り出し、メタノールを含む1xトランスファーバッファー中でインキュベートします。
- 湿式移送システムを準備し、メタノールでPVDF膜を活性化する。湿式転写用のゲルとPVDF膜を組み立て、移送タンクにアイスブロックを配置し、タンク全体を氷に沈めて、移送条件が冷たいままであることを確認します。350 mA/100 V で 60 分間実行します。
- 50rpm(16mm軌道)のオービタルシェーカー上で穏やかに振とうしながら、室温で30分間ブロッキング溶液中でインキュベートすることによって膜を遮断する。
- 滅菌水で膜を5分間すすぎ、繰り返します。次に、きれいなメスを使用して、目に見えるタンパク質はしごをガイドとして使用して、膜を〜50kDaで水平に慎重に切断します。
- 50kDaを超えるタンパク質を保有する膜を抗α-1,6-フコシルトランスフェラーゼ抗体と共に抗体希釈バッファー中で1:1000でインキュベートする。希釈バッファー中の1:10,000で抗GAPDHと共に50kDa未満の固定化タンパク質と共に膜をインキュベートする。膜は、室温で1時間、または4°Cで一晩インキュベートすることができる。
- メンブレンを5分間洗浄し(x3)、適切な二次抗体とともに室温で少なくとも30分間インキュベートします。
- 洗浄バッファーで3回洗浄を5分間行い、続いて水で3回洗浄します。次いで、バンドが現れるまで(1〜60分間)、発色基質(または適切な検出試薬)で膜を発達させる。
- メンブレン2xを水ですすぎ、乾燥させます。膜の画像をキャプチャし、濃度測定分析40を実行する。
- 各サンプルにおける相対タンパク質発現を計算するには、まずサンプル中のGAPDHシグナルの比率を計算する。これは、サンプル負荷量の不一致を補正する各サンプルの正規化係数です。

- (各レーンの)FUT8シグナル強度を対応するレーンの正規化係数で除算し、相対的なFUT8タンパク質発現を得る。

結果
ウェスタンブロット分析は、3つの Fut8 DsiRNA構築物の混合物でトランスフェクトされた細胞におけるFUT8タンパク質発現の低下を示した。非標的性DsiRNAを導入した対照試料において、FUT8は〜65および70kDaでダブルバンドとして現れた。FUT8の予測分子量は66kDaであるため、低分子量バンドのシグナル強度の低下は遺伝子サイレンシングの指標である。遺伝子サイレンシングを確認および定量するために、FUT8タンパク質のレベルを相対GAPDHタンパク質レベルに正規化した。ウェスタンブロット分析では、GAPDHの2つのバンドが〜37および35kDaで検出されました。より高い分子量のバンドは、予測されたタンパク質サイズに対応し、したがって、正規化計算に使用される。GAPDHタンパク質レベルに対して標準化すると、FUT8タンパク質発現は最大60%減少した(図1)。
トランスフェクション後48時間での遺伝子ノックダウンの観察に沿って、対応するmAbサンプルをCGE−LIFによる分析のために処理した。ノックダウン細胞由来のグリカン構造は、フコシル化の減少を示した。この傾向は、アガラクトシル化構造(G0F)において最も顕著であり、ガラクトシル化構造(G1F、G1F'およびG2F)においてより少ない程度で観察された。このデータセットから、IgGコアフコシル化の総量は、陰性対照条件で観察されたコアフコシル化の約95%から〜75%に減少した(図2)。コアフコシル化のより大きな減少は、FUT8タンパク質レベルの〜60%の減少を考えると予想された。熟考すると、グリコプロファイルはトランスフェクション以来48時間にわたって蓄積したグリコシル化mAbsを表し、遺伝子サイレンシングは収穫時に存在するタンパク質レベルのみを表すことは注目に値する。
このノックダウン法のさらなる精査には、DsiRNA濃度、収穫時間、およびエレクトロポレーション条件の変化が含まれていました。各因子を個別に調査し、その関連性を判断しました。コアフコシル化および細胞生存率に対するエレクトロポレーションパルス条件の影響は、実験B、C、D、およびEで捕捉される。これらの結果は、2つの方形波パルスを用いたエレクトロポレーション(実験C)からのコアフコシル化が、単一の方形波パルス(実験B)と比較して2倍に減少したことを実証し、細胞生存率に有意差はない(表2)。エレクトロポレーション条件e3(実験D)は、この時点での細胞生存率(〜90%)およびIgG収率が最も低いことをもたらした。しかしながら、エレクトロポレーション事象を生存した細胞は、コアフコシル化の〜10%の減少によって証明されるように、適度にトランスフェクトされた(表2)。興味深いことに、実験Dは、コアフコシル化(14.7%)を最大に減少させたが、明らかに細胞生存率(91%〜93%生存率)に有害であったエレクトロポレーション条件を使用した。この限られた実験セットは、取り返しのつかない損傷を引き起こすことなく細胞膜の十分な透過を可能にするエレクトロポレーション設定を決定する必要性を示しています。また、コアフコシル化におけるsiRNA濃度と収穫時間の役割に注目することも興味深い。全体として、siRNA濃度の増加は、収穫時間を増加させるよりもコアフコシル化に大きな影響を与える(実験B、F、G対実験A、B、H)。将来の実験では、エレクトロポレーション法e2によって送達されるsiRNA濃度を滴定することは興味深いであろう。

図1.実験フローチャート。 糖工学およびサンプル分析ステップは、各ステップに必要な関連する時間とともに描かれている。siRNAの設計は、遺伝子標的の数または遺伝子標的ごとの構築物の数に応じて、数時間かかる。siRNAによるCHO細胞トランスフェクションは数時間で完了し、形質転換細胞は48時間増殖するように放置される。細胞ペレットおよび上清は、数時間以内に回収される。細胞ペレットを溶解し、細胞内タンパク質をSDS PAGE上で分離し、続いて標的遺伝子に対する抗体でブロットおよびプローブする。グリカンは精製された抗体から切断され、CGE−LIFによって分析される。これらのアッセイは、それぞれ1日を必要とし得る。 この図の拡大版を表示するには、ここをクリックしてください。
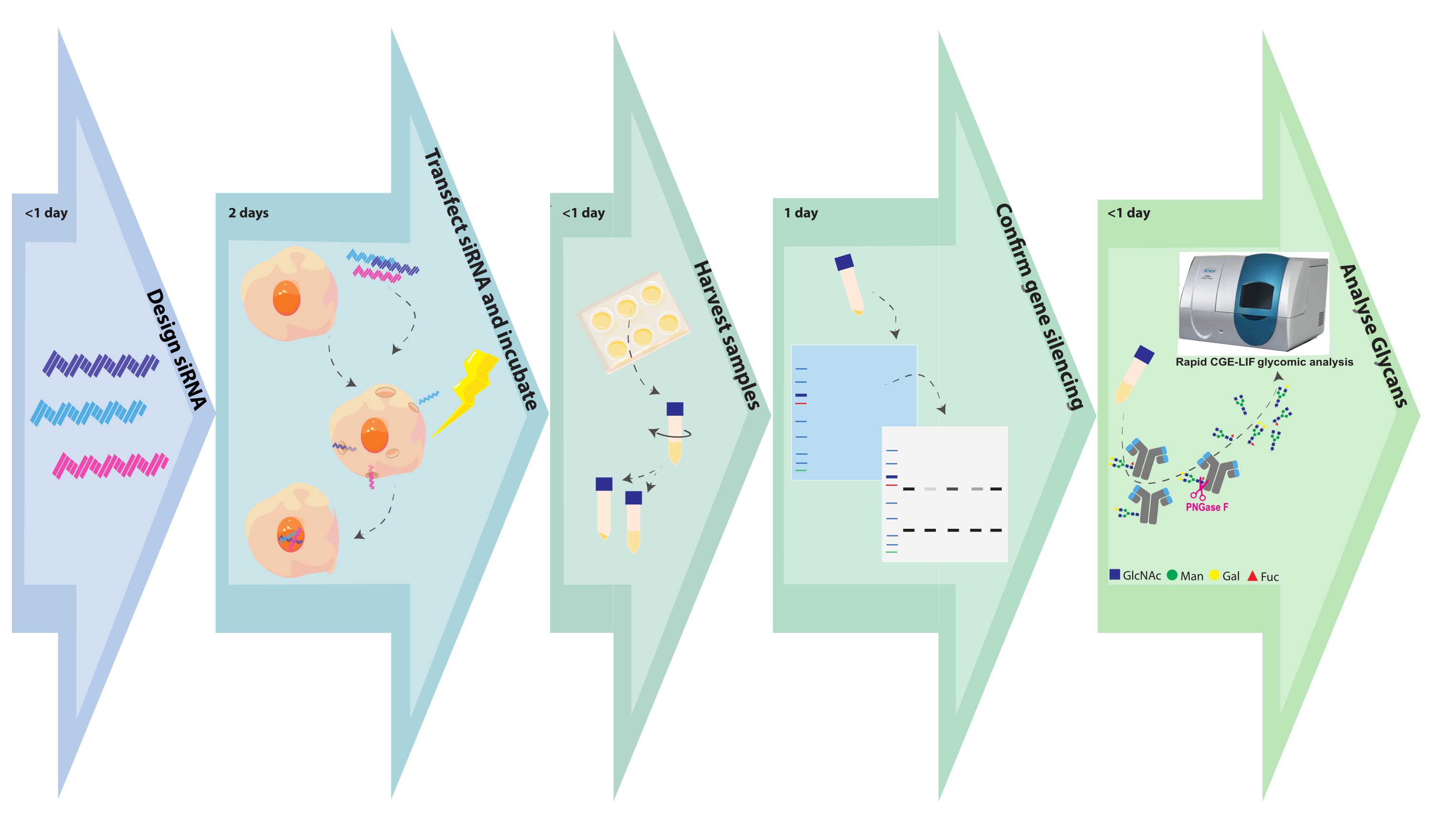{kind=link}

図2.RNA干渉の確認。Fut8または非標的対照DsiRNAで処理したサンプル中のα-1,6-フコシルトランスフェラーゼ(FUT8)タンパク質レベルのウェスタンブロット検出。FUT8に対応するバンドは、Fut8ノックダウンサンプルよりも対照においてより強い。標的遺伝子発現を正常化するために、GAPDHタンパク質レベルも評価した。全ての試料を実験Gから採取した(表2参照)。Kotidisらより転載。52. この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

図3.48時間における累積IgGグリコシル化に対する Fut8 ノックダウンの効果。 グリカン分布の変化は、ノックダウンサンプルで検出されます。特に、主コアフコシル化構造(G0F)の相対的存在量は減少し、アフコシル化種はノックダウン実験において増加する。測定は実験Gサンプルから行った( 表2参照)。各実験に対して実施した生物学的3連を採取後に混合し、下流分析の負担を軽減した。Kotidisらより転 載。52. この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
| 実験名 | エレクトロポレーション法 | DsiRNA濃度(nΜ) | 収穫時期(h) | 生存率(%) | Xv (106セル ・mL-1) | IgG 力価 (mg·L-1) | コアフコシル化の違い(%) | ||||||
| ExpA_Negative | E1 · | 500 | 24 | 98.3 | 4.71 | 122.5 | - | ||||||
| ExpA_Knockdown | E1 · | 500 | 24 | 98.3 | 4.9 | 110.3 | 4.08 | ||||||
| ExpB_Negative | E1 · | 500 | 48 | 95.6 | 9.55 | 453.3 | - | ||||||
| ExpB_Knockdown | E1 · | 500 | 48 | 96.7 | 9.61 | 469 | 5.38 | ||||||
| ExpC_Negative | E2 · | 500 | 48 | 96.3 | 9.91 | 449.3 | - | ||||||
| ExpC_Knockdown | E2 · | 500 | 48 | 96.7 | 11 | 454.6 | 11.42 | ||||||
| ExpD_Negative | e3 · | 500 | 48 | 90.6 | 6.25 | 318.5 | - | ||||||
| ExpD_Knockdown | e3 · | 500 | 48 | 89.1 | 6.09 | 311.85 | 9.71 | ||||||
| ExpE_Negative | E4 · | 500 | 48 | 91.1 | 7.2 | 380.3 | - | ||||||
| ExpE_Knockdown | E4 · | 500 | 48 | 93.3 | 7.79 | 422.8 | 14.7 | ||||||
| ExpF_Negative | E1 · | 750 | 48 | 96.2 | 9.7 | 501 | - | ||||||
| ExpF_Knockdown | E1 · | 750 | 48 | 95.7 | 9.76 | 504.6 | 9.9 | ||||||
| ExpG_Negative | E1 · | 1000 | 48 | 96.1 | 11.1 | 422.6 | - | ||||||
| ExpG_Knockdown | E1 · | 1000 | 48 | 95.9 | 9.73 | 499.3 | 17.26 | ||||||
| ExpH_Negative | E1 · | 500 | 72 | 94.4 | 14.3 | 925.8 | - | ||||||
| ExpH_Knockdown | E1 · | 500 | 72 | 95 | 13.5 | 1018.4 | 7.37 | ||||||
表 2.トランスフェクションの最適化。 エレクトロポレーション法の反復的改変、DsiRNA濃度、および回収時間は、細胞生存率、生細胞密度、回収時のIgG力価、およびコアフコシル化の違いをもたらした。各実験は、ノックダウンとそれぞれの陰性対照とを比較して、修飾が所望の効果(すなわち、フコシル化の減少)を生じるかどうかを決定する。エレクトロポレーションの設定は以下の通りであった: e1: 1200 V, 0.1 ms, 方形波形;e2: 1200 V, 2x 0.1 ms, パルス間5秒, 方形波形;e3: 150 V, 20 ms, 方形波形;e4: 250 V、500 μF、指数関数的減衰。Kotidisらより転載。52。
ディスカッション
グリコシル化経路は、酵素および補助タンパク質の複雑な代謝ネットワークを含む。経路構成成分の機能を解剖することは、従来のノックアウトまたはノックイン遺伝子工学戦略のみに依存する場合、困難である。別のアプローチは、一過性機能喪失アッセイを用いて経路のメンバーを予めスクリーニングすることである。この目的のために、RNAiおよびCGE-LIF検出の2つの迅速なプロトコルを組み合わせて、グリコシル化遺伝子を特徴付けるより効率的な方法を作り出した。記載されている方法は、完了に数週間かかる可能性のある従来の方法と比較して、完了に5〜7日かかります。さらに、自動化機能を備えた研究環境は、この方法を利用して、手動処理で実現可能なよりも多くの遺伝子候補をスクリーニングする可能性があります。
一過性糖工学キャンペーンの成功は、siRNAデザインに大きく依存します。カスタムDsiRNAデザインは、以前に概説した規則に従わなければならないか、または容易にするために、ユーザーは市販の事前設計された配列を選択することができる。他の遺伝子改変戦略と同様に、RNAiはオフターゲット効果の可能性を秘めています。したがって、使用者は、計算方法41によって意図しない遺伝子標的化を評価することが奨励される。実験計画の選択は、ターゲット外効果を制限するのにも役立ちます。Kittlerらは、siRNAの多重送達がオフターゲット効果の低下をもたらしたことを示した42。これは直感に反しているように思えるが、マスターミックスは各siRNA構築物の濃度を低下させ、オフターゲット遺伝子サイレンシングの機会を制限することが示唆されている。さらなる利点は、同じ遺伝子を標的とするsiRNA構造の同時トランスフェクションにより、RNAiが成功する可能性が高まることです。また、マスターミックスを使用することで、サンプルと複製の間の一貫性が確保され、トランスフェクションプロセスが高速化されます。混合siRNA構築物の初期スクリーニングに続いて、各配列のRNAi効率を確認するために、個々の構築物を用いて別の実験を実施してもよい。このおよび他のノックダウン研究において、最大3つのsiRNAがプールされ、細胞43、44、45に送達された。しかし、単一の遺伝子を効率的に標的にしたり、複数の遺伝子を標的にしたりするために、3つ以上のsiRNAを同時にスクリーニングすることが望ましい場合があります。実際、ある研究では、個々の遺伝子のサイレンシングに匹敵するレベルで、最大6つの遺伝子の多重化siRNA媒介サイレンシングが実証されました46。しかし、サイレンシング効率を損なうことなくプール内で使用できるsiRNA構築物の最大数を決定するためには、さらなる研究が必要である。この多重化戦略は、RNAiライブラリースクリーニング実験46のペースを速めるためにMartinらによって提案され、同様の概念がグリコシル化遺伝子のスクリーニングに有用であることが証明される可能性がある。
本明細書に記載されるプロトコルは、他のグリコシル化遺伝子を検証するためにその後の実験が行われることを期待して、概念実証として役立つ。関心のある新しい遺伝子は、Fut8よりも特徴付けられていないか、または一般的ではない可能性があり、遺伝子サイレンシングを検出するための一次抗体は不十分であるか、または利用できない可能性がある。このシナリオでは、遺伝子サイレンシング47を定量するためにRT-PCRなどの代替方法を使用してもよいが、RT-PCRはタンパク質ではなくmRNAを検出することに留意すべきである。ウェスタンブロッティング用の抗体が利用可能な場合、一般的な問題は検出不良または非特異的バンドの存在です。ユーザーが一般的な問題を解決するのに役立つトラブルシューティングガイドが利用可能であり、これらには、一次抗体滴定、代替ブロッキング、および検出条件48,49などの幅広いソリューションが含まれる傾向がある。この研究では、FUT8は予想外に〜65および〜70kDaでダブルバンドとして現れた。〜70kDaバンドはグリコシル化FUT8を表す可能性がある。ヒト細胞株からの文献証拠は、チャイニーズハムスターにおいて保存されている部位であるThr 56450、51、およびCHO K1 FUT8配列におけるO結合型グリコシル化を記載している。
前述のように、グリコシル化経路は、しばしば複雑な酵素配列を伴う。現在のプロトコールは、 Fut8によって制御される単遺伝子グリコシル化反応を用いて開発、最適化、および実証されている。したがって、標的遺伝子が代替動態および発現レベルを有する酵素、または冗長な機能を有するアイソザイムによって調節される経路をコードする場合、この方法の堅牢性を確認するためにさらなる研究が必要である。
まとめると、遺伝子を迅速にサイレンシングし、修飾IgGグリコプロファイルを検出する能力は、カスタムグリコエンジニアリング抗体に向けた取り組みにおいて有用なツールです。同様の短期研究からの洞察は、流加培養などの長期アッセイで使用するための安定した糖操作細胞を生成するために適用することができる。薬学的文脈の外では、この方法はグリカン生物学の研究に貢献し、発達、健康、および疾患におけるグリカンの重要な機能を強調する。
開示事項
著者らは開示するものは何もありません。
謝辞
PKは、彼の奨学金のために、インペリアルカレッジロンドンの化学工学科に感謝します。RDは、彼の学生時代のために英国のバイオテクノロジーと生物科学研究評議会に感謝します。MMは、英国バイオテクノロジー生物科学研究評議会(Grant reference: BB/S006206/1)から資金提供を受けています。IAGはアイルランド研究評議会(奨学金番号)に感謝します。GOIPG/2017/1049)およびCONACyT(奨学金番号438330)。
資料
| Name | Company | Catalog Number | Comments |
| 32 Karat software | SCIEX | contact manufacturer | Software for glycan data acquisition and analysis using the Fast Glycan analysis protocol and separation method. |
| Acetonitrile, HPLC grade | Sigma Aldrich | 34851 | Solvent. |
| Anti-FUT8 antibody | AbCam | ab198741 | Rabbit polycloncal to Fut8. Use this antibody to quantify Fut8 protein expression; replace this antibody if using siRNA targeting a different gene. |
| Anti-GAPDH antibody | AbCam | ab181602 | Rabbit monoclonal to GAPDH. Alternative housekeeping genes exist and might be preferred by the user. |
| BioDrop Spectrophotometer | Biochrom | 80 3006 55 | Instrument used to quantify protein concentration. |
| BLItz | ForteBio | 45 5000 | Instrument. Label-Free Protein Analysis System. |
| BRAND Haemocytometer | Sigma Alrich | BR717810 | Counting chamber device |
| Capillary cartridge | SCIEX | A55625 | Pre-assembled capillary cartridge with window (30 cm total length, 375 µm outer diameter (o.d), x 50 µm inner diameter (i.d). |
| C100HT Glycan analysis—capillary electrophoresis | SCIEX | contact manufacturer | Capillary gel electrophoresis instrument, the CESI 8000 Plus instrument is now used. |
| CD CHO Medium | Thermo Fisher Scientific | 10743029 | Replace this with a culture medium appropriate for the cell line of choice. |
| Centrifuge tubes, 15 mL | Greiner Bio | 188261 | Sterile polypropylene tube. |
| Centrifuge tubes, 50 mL | Greiner Bio | 227270 | Sterile polypropylene tube. |
| CHO IgG | MedImmune | Gift | Chinese Hamster Ovary cells expressing an IgG monoclonal antibody (CHO T127). Created using the GS system. |
| Dulbecco's phosphate-buffered saline (DPBS) | Gibco | 14190144 | 1x PBS, without calcium or magnesium. |
| Erlenmeyer Flasks with Vent Cap, 125 mL | Corning | 431143 | Replace this with a culture vessel suitable for growing the cell line of choice. |
| Erlenmeyer Flasks with Vent Cap, 250 mL | Corning | 431144 | Replace this with a culture vessel suitable for growing the cell line of choice. |
| Fast Glycan Labelling and Analysis kit | SCIEX | B94499PTO | Labels N-glycans with APTS and then uses a magnetic-bead based clean up system to remove excess APTS. |
| Fut8 DsiRNA | IDT | Custom | Custom designed DsiRNA targetting Fut8. |
| Gene Pulser cuvettes, 0.4 cm | Bio-Rad | 1652088 | Electroporation cuvette. |
| Gene Pulser Xcell Eukaryotic System | Bio-Rad | 165 2661 | Insturment. Xcell main unit with Capacitance Extender (CE) Mocdule and ShockPod. |
| Immobilon-FL PVDF membrane | Merck-Millipore | IPFL00010 | Immunoblot transfer membrane, low background. |
| L-Methionine sulfoximine (MSX) | Sigma Aldrich | M5379 | Only necessary for CHO cell lines using the glutamine synthetase (GS) selection system. |
| Kimwipes | Thermo Fisher Scientific | 10623111 | Low-lint, high absorbency and chemically inert wipes. |
| M-PER Mammalian Protein Extraction Reagent | Thermo Fisher Scientific | 78505 | Alternative lysis buffers such as RIPA are also appropriate. |
| Methanol, HPLC grade | Fisher Scientific | 10365710 | Solvent. |
| Microcentrifuge tubes, 1.5 mL | Eppendorf | 616201 | Autoclavable tubes. |
| Mini-PROTEAN Tetra Vertical Electrophoresis Cell system | Bio-Rad | 1658035FC | Instrument. 4-gel capacity, for 1.0 mm thick handcast gels, with Mini Trans-Blot Module and PowerPac HC Power Supply. |
| NC-Slide A8 | ChemoMetec | 942 0003 | 8-chamber slide for use with NucleoCounter NC 250. |
| Negative Control DsiRNA, 5 nmol | IDT | 51 01 14 04 | Non-targeting DsiRNA. |
| Nuclease-free duplex buffer | IDT | 11-01-03-01 | Reconstitution buffer for DsiRNA. |
| NucleoCounter NC-250 | Chemometec | contact manufacturer | Instrument. Automated Cell Analyzer |
| Page-Ruler ladder, 10 to 180 kDa | Thermo Fisher Scientific | 26616 | Mixed blue, orange and green protein standards for SDS PAGE and western blotting. |
| PCR tubes | Greiner Bio | 608281 | Autoclavable tubes for DsiRNA aliqouts and glycan preparation. |
| Pipette filter tips sterilised (10, 200, 1000 µL) | Gibson | F171203, F171503, F171703 | Sterile filter tips to avoid RNA contamination. |
| PNGase F enzyme | New England Biolabs | P0704S | Enzymatic cleavage of glycans from glycoproteins. |
| Polypropylene columns, 1 mL | Qiagen | 34924 | Columns for gravity-flow chromatography. |
| Protease Inhibitor Cocktail | Sigma Aldrich | P8340 | Inhibition of serine, cysteine, aspartic proteases and aminopeptidases |
| Protein-A Agarose Beads | Merck-Millipore | 16 125 | For purification of human, mouse and rabbit immunoglobulins. |
| Protein-A biosensor | ForteBio | 18 5010 | Tips functionalised with Protein A for rapid antibody quantification. |
| RNaseZap | Invitrogen | AM9780 | Removes RNAse contamination. |
| Sample dilutent | ForteBio | 18 1104 | Activate Protein A tips. |
| Serological pipets (5, 10, 25 mL) | Corning | 4487, 4488, 4489 | Used for sterile cell culture tecniques. |
| Sodium cyanoborohydride solution 1 M in THF | Sigma Aldrich | 296813 | Reducing agent. |
| Solution 18 | ChemoMetec | 910-3018 | Staining reagent containing acridine orange (AO) and 4',6-diamidino-2-phenylindole (DAPI) |
| Spin-X Centrifuge Tube Filters | Corning | 8161 | 0.22 µm pore, Cellulose Acetate membrane. |
| Suspension plate with lid, 6-well | Greiner Bio | 657 185 | Hydrophobic culture plate for growth of suspension cultures. |
| Syringe filters, 0.22 μm | Sartorius | 514 7011 | Surfactant-free cellulose acetate (SFCA) |
| Syringes with Luer lock tip, 20 mL | Fisher Scientific | 10569215 | For secure connection with syringe filter. |
| Trypan Blue solution | Gibco | 15250061 | Stains dead and dying cells. |
| Vivaspin 500, 3,000 MWCO | Sartorius | VS0191 | Polyethersulfone |
| WesternBreeze Chromogenic Kit, anti-rabbit | Thermo Fisher Scientific | WB7105 | Western blot detection kit, alternative blocking buffers and antibody diluents can be made by the user using recipes available online. |
参考文献
- Jefferis, R. Glycosylation as a strategy to improve antibody-based therapeutics. Nature Reviews Drug Discovery. 8 (3), 226-234 (2009).
- Rathore, A. S., Winkle, H. Quality by design for biopharmaceuticals. Nature Biotechnology. 27 (1), 26-34 (2009).
- Eon-Duval, A., Broly, H., Gleixner, R. Quality attributes of recombinant therapeutic proteins: An assessment of impact on safety and efficacy as part of a quality by design development approach. Biotechnology Progress. 28 (3), 608-622 (2012).
- Lalonde, M. -. E., Durocher, Y. Therapeutic glycoprotein production in mammalian cells. Journal of Biotechnology. 251, 128-140 (2017).
- Mimura, Y., et al. Glycosylation engineering of therapeutic IgG antibodies: challenges for the safety, functionality and efficacy. Protein & Cell. 9 (1), 47-62 (2018).
- Walsh, G. Biopharmaceutical benchmarks 2018. Nature Biotechnology. 36 (12), 1136-1145 (2018).
- Zhang, M., Koskie, K., Ross, J. S., Kayser, K. J., Caple, M. V. Enhancing glycoprotein sialylation by targeted gene silencing in mammalian cells. Biotechnology and Bioengineering. , (2010).
- Kanda, Y., et al. et al. of a GDP-mannose 4,6-dehydratase (GMD) knockout host cell line: a new strategy for generating completely non-fucosylated recombinant therapeutics. Journal of Biotechnology. 130 (3), 300-310 (2007).
- Umaña, P., Jean-Mairet, J., Moudry, R., Amstutz, H., Bailey, J. E. Engineered glycoforms of an antineuroblastoma IgG1 with optimized antibody-dependent cellular cytotoxic activity. Nature Biotechnology. 17 (2), 176-180 (1999).
- Xu, X., et al. et al. genomic sequence of the Chinese hamster ovary (CHO)-K1 cell line. Nature Biotechnology. 29 (8), 735-741 (2011).
- Chan, K. F., et al. et al. of GDP-fucose transporter gene (Slc35c1) in CHO cells by ZFNs, TALENs and CRISPR-Cas9 for production of fucose-free antibodies. Biotechnology Journal. 11 (3), 399-414 (2016).
- Zhang, P., et al. Identification of functional elements of the GDP-fucose transporter SLC35C1 using a novel Chinese hamster ovary mutant. Glycobiology. 22 (7), 897-911 (2012).
- Amann, T., et al. Genetic engineering approaches to improve posttranslational modification of biopharmaceuticals in different production platforms. Biotechnology and Bioengineering. 116 (10), 2778-2796 (2019).
- Karottki, K. J., et al. Awakening dormant glycosyltransferases in CHO cells with CRISPRa. Biotechnology and Bioengineering. 117 (2), 593-598 (2020).
- Fire, A., et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 391 (6669), 806-811 (1998).
- Hammond, S. M., Bernstein, E., Beach, D., Hannon, G. J. An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells. Nature. 404 (6775), 293-296 (2000).
- Bernstein, E., Caudy, A. A., Hammond, S. M., Hannon, G. J. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature. 409 (6818), 363-366 (2001).
- Shields, R. L., et al. et al. of fucose on human IgG1 N-linked oligosaccharide improves binding to human FcγRIII and antibody-dependent cellular toxicity. Journal of Biological Chemistry. 277 (30), 26733-26740 (2002).
- Shinkawa, T., et al. et al. Absence of fucose but not the presence of galactose or bisecting N-acetylglucosamine of human IgG1 complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity. Journal of Biological Chemistry. 278 (5), 3466-3473 (2003).
- Mori, K., et al. Engineering Chinese hamster ovary cells to maximize effector function of produced antibodies using FUT8 siRNA. Biotechnology and Bioengineering. 88 (7), 901-908 (2004).
- Imai-Nishiya, H., et al. Double knockdown of alpha1,6-fucosyltransferase (FUT8) and GDP-mannose 4,6-dehydratase (GMD) in antibody-producing cells: a new strategy for generating fully non-fucosylated therapeutic antibodies with enhanced ADCC. BMC Biotechnology. 7, 84 (2007).
- Beuger, V., et al. et al. silencing of fucosyltransferase 8 in Chinese-hamster ovary cells for the production of antibodies with enhanced antibody immune effector function. Biotechnology and Applied Biochemistry. 53 (1), 31 (2009).
- Templeton, N., Dean, J., Reddy, P., Young, J. D. Peak antibody production is associated with increased oxidative metabolism in an industrially relevant fed-batch CHO cell culture. Biotechnology and Bioengineering. 110 (7), 2013-2024 (2013).
- Hussain, H., et al. A comparative analysis of recombinant Fab and full-length antibody production in Chinese hamster ovary cells. Biotechnology and Bioengineering. 118 (12), 4815-4828 (2021).
- Shimoyama, H., et al. Partial silencing of fucosyltransferase 8 gene expression inhibits proliferation of Ishikawa cells, a cell line of endometrial cancer. Biochemistry and Biophysics Reports. 22, 100740 (2020).
- Tummala, S., et al. Evaluation of exogenous siRNA addition as a metabolic engineering tool for modifying biopharmaceuticals. Biotechnology Progress. 29 (2), 415-424 (2013).
- Carillo, S., et al. Comparing different domains of analysis for the characterisation of N-glycans on monoclonal antibodies. Journal of Pharmaceutical Analysis. 10 (1), 23-34 (2020).
- Reusch, D., et al. Comparison of methods for the analysis of therapeutic immunoglobulin G Fc-glycosylation profiles-Part 2: Mass spectrometric methods. mAbs. 7 (4), 732-742 (2015).
- Donini, R., Haslam, S. M., Kontoravdi, C. Glycoengineering Chinese hamster ovary cells: a short history. Biochemical Society Transactions. 49 (2), 915-931 (2021).
- Szigeti, M., Chapman, J., Borza, B., Guttman, A. Quantitative assessment of mAb Fc glycosylation of CQA importance by capillary electrophoresis. ELECTROPHORESIS. 39 (18), 2340-2343 (2018).
- Ruhaak, L. R., et al. Optimized workflow for preparation of APTS-labeled N-glycans allowing high-throughput analysis of human plasma glycomes using 48-channel multiplexed CGE-LIF. Journal of Proteome Research. 9 (12), 6655-6664 (2010).
- Patenaude, A. -. M., et al. N-glycosylation analysis of mouse immunoglobulin G isolated from dried blood spots. Electrophoresis. 42 (24), 2615-2618 (2021).
- Karst, D. J., et al. Modulation and modeling of monoclonal antibody N-linked glycosylation in mammalian cell perfusion reactors. Biotechnology and Bioengineering. 114 (9), 1978-1990 (2017).
- Dadouch, M., Ladner, Y., Perrin, C. Analysis of monoclonal antibodies by capillary electrophoresis: sample preparation, separation, and detection. Separations. 8 (1), 4 (2021).
- Fakhr, E., Zare, F., Teimoori-Toolabi, L. Precise and efficient siRNA design: a key point in competent gene silencing. Cancer Gene Therapy. 23 (4), 73-82 (2016).
- Birmingham, A., et al. A protocol for designing siRNAs with high functionality and specificity. Nature Protocols. 2 (9), 2068-2078 (2007).
- Makrydaki, E., et al. . Immobilised enzyme cascade for targeted glycosylation. , (2022).
- . Bulletin_6201.pdf Available from: https://www.bio-rad.com/webroot/web/pdf/lsr/literature/Bulletin_6201.pdf (2022)
- . Bulletin_6376.pdf Available from: https://www.bio-rad.com/webroot/web/pdf/lsr/literature/Bulletin_6201.pdf (2022)
- . Dot Blot Analysis Available from: https://imagej.nih.gov/ij/docs/examples/dot-blot/index.html (2022)
- Yilmazel, B., et al. Online GESS: prediction of miRNA-like off-target effects in large-scale RNAi screen data by seed region analysis. BMC Bioinformatics. 15 (1), 192 (2014).
- Kittler, R., et al. Genome-wide resources of endoribonuclease-prepared short interfering RNAs for specific loss-of-function studies. Nature Methods. 4 (4), 337-344 (2007).
- Stec, E., et al. A multiplexed siRNA screening strategy to identify genes in the PARP pathway. Journal of Biomolecular Screening. 17 (10), 1316-1328 (2012).
- Brown, J. M., et al. Ligand conjugated multimeric siRNAs enable enhanced uptake and multiplexed gene silencing. Nucleic Acid Therapeutics. 29 (5), 231-244 (2019).
- Parsons, B. D., Schindler, A., Evans, D. H., Foley, E. A direct phenotypic comparison of siRNA pools and multiple individual duplexes in a functional assay. PLoS One. 4 (12), 8471 (2009).
- Martin, S. E., et al. Multiplexing siRNAs to compress RNAi-based screen size in human cells. Nucleic Acids Research. 35 (8), 57 (2007).
- Popp, O., Moser, S., Zielonka, J., Rüger, P., Hansen, S., Plöttner, O. Development of a pre-glycoengineered CHO-K1 host cell line for the expression of antibodies with enhanced Fc mediated effector function. mAbs. 10 (2), 290-303 (2018).
- Yang, P. -. C., Mahmood, T. Western blot: Technique, theory, and trouble shooting. North American Journal of Medical Sciences. 4 (9), 429 (2012).
- Troubleshooting Western Blots with the Western Blot Doctor. Bio-Rad Laboratories Available from: https://www.bio-rad.com/en-uk/applications-technologies/troubleshooting-western-blots-with-western-blot-doctor?ID=MIW4HR15 (2022)
- Steentoft, C., et al. Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology. EMBO. 32 (10), 1478-1488 (2013).
- . FUT8 - Alpha-(1,6)-fucosyltransferase - Proteomics Available from: https://www.nextprot.org/entry/NX_Q9BYC5/proteomics (2022)
- Kotidis, P., et al. Rapid antibody glycoengineering in CHO cells via RNA interference and CGE-LIF N-glycomics. Methods in Molecular Biology. 2370, 147-167 (2022).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved