Method Article
Glycoingénierie rapide des anticorps dans les cellules ovaires du hamster chinois
* Ces auteurs ont contribué à parts égales
Dans cet article
Résumé
Le schéma de glycosylation d’un anticorps détermine ses performances cliniques, de sorte que les efforts industriels et académiques pour contrôler la glycosylation persistent. Étant donné que les campagnes typiques de glyco-ingénierie nécessitent beaucoup de temps et de main-d’œuvre, la génération d’un protocole rapide pour caractériser l’impact des gènes de glycosylation à l’aide d’un silence transitoire s’avérerait utile.
Résumé
Les anticorps monoclonaux recombinants lient des cibles moléculaires spécifiques et, par la suite, induisent une réponse immunitaire ou inhibent la liaison d’autres ligands. Cependant, la fonctionnalité et la demi-vie des anticorps monoclonaux peuvent être réduites par le type et la distribution de la glycosylation spécifique à l’hôte. Les tentatives de produire des anticorps supérieurs ont inspiré le développement de cellules productrices génétiquement modifiées qui synthétisent des anticorps glyco-optimisés. La glyco-ingénierie nécessite généralement la génération d’une lignée cellulaire knockin ou knockin stable à l’aide de méthodes telles que la protéine 9 associée à de courtes répétitions palindromiques régulièrement espacées en grappes (CRISPR). Les anticorps monoclonaux produits par les cellules modifiées sont ensuite caractérisés à l’aide de méthodes de spectrométrie de masse pour déterminer si le glycoprofil souhaité a été obtenu. Cette stratégie prend beaucoup de temps, est techniquement difficile et nécessite des spécialistes. Par conséquent, une stratégie alternative qui utilise des protocoles rationalisés pour la glyco-ingénierie génétique et la détection des glycanes peut aider les efforts vers des anticorps optimaux. Dans cette étude de preuve de concept, une cellule d’ovaire de hamster chinois produisant des IgG a servi d’hôte idéal pour optimiser la glyco-ingénierie. Un ARN interférent court ciblant le gène Fut8 a été livré aux cellules de l’ovaire de hamster chinois, et les changements qui en ont résulté dans l’expression de la protéine FUT8 ont été quantifiés. Les résultats indiquent que le knockdown par cette méthode était efficace, conduisant à une réduction d’environ 60% de FUT8. Une analyse complémentaire du glycoprofil d’anticorps a été réalisée à l’aide d’une technique rapide mais très sensible : l’électrophorèse sur gel capillaire et la détection de fluorescence induite par laser. Toutes les expériences de knockdown ont montré une augmentation des glycanes afucosylés; cependant, le plus grand changement réalisé dans cette étude était d’environ 20 %. Ce protocole simplifie les efforts de glyco-ingénierie en exploitant des outils de conception in silico , des réactifs de ciblage de gènes synthétisés commercialement et des tests de quantification rapide qui ne nécessitent pas d’expérience préalable approfondie. En tant que tel, les gains de temps offerts par ce protocole peuvent aider les recherches sur de nouvelles cibles génétiques.
Introduction
La glycosylation liée à l’azote est un processus enzymatique par lequel les fractions oligosaccharidiques sont liées de manière covalente aux résidus d’Asn. Contrairement à la synthèse des protéines de novo, la synthèse des glycanes est une réaction non modélisée qui entraîne une glycosylation hétérogène des protéines. La structure, la composition et la distribution des glycanes peuvent affecter la conformation et la fonction des protéines. En effet, la N-glycosylation dans la région du fragment cristallisable (Fc) de l’immunoglobuline G (IgG) régule l’efficacité thérapeutique, l’immunogénicité et la demi-vie de l’anticorps1. En tant que tel, le paradigme de la qualité dès la conception (QbD) pour le développement de produits protéiques biothérapeutiques recombinants identifie naturellement la glycosylation comme un attribut de qualité critique (CQA)2,3. Les cellules de mammifères sont souvent les systèmes d’expression préférés car elles produisent intrinsèquement des modèles de glycosylation semblables à ceux de l’homme plus étroitement que les bactéries, les levures, les insectes ou les cellules végétales. De plus, les cellules de l’ovaire de hamster chinois (CHO) sont sélectionnées par rapport à d’autres lignées cellulaires de mammifères parce qu’elles sont résistantes à l’infection virale humaine, sécrètent des produits à titre élevé et peuvent être cultivées en culture en suspension à des densités cellulaires viables élevées4. En ce qui concerne la formation de glycanes, les cellules de production murine non CHO génèrent des glycanes immunogènes (galactose lié à la α(1-3)-Gal [α(1-3)-Gal] et acide N-glycolylneuraminique [NeuGc]) qui empiètent sur l’utilisation sûre des anticorps monoclonaux (mAbs)5. Ces avantages font des cellules CHO le premier système d’expression, responsable de la production de plus de 80% de nouvelles biothérapies entre 2014 et 20186. Cependant, la glycosylation indépendante du modèle est un mécanisme conservé qui conduit à des biothérapies dérivées de CHO avec un éventail de glycoformes.
Les stratégies de développement biothérapeutique visent à contrôler l’hétérogénéité des cellules CHO par génie génétique. Parmi les exemples de littérature, citons l’élimination des sialidases (Neu1, Neu3) 7, le knockout8 de la GDP-mannose 4,6-déshydratase (GMD) et la surexpression des glycosyltransférases (GnTIII)9. Les progrès de la glyco-ingénierie sont possibles grâce à une combinaison de ressources accessibles au public, comme le génomeCHO 10, et le développement continu d’outils de génie génétique, tels que les nucléases effectrices de type activateur de transcription (TALEN), les nucléases à doigts de zinc (ZFN) et les courtes répétitions palindromiques intercalaires groupées (CRISPR) associées à la protéine 9 (CRISPR-Cas9)11,12,13,14 . Ces outils sont généralement livrés aux cellules CHO sous forme d’ADN plasmidique ou de complexes ribonucléoprotéiques purifiés (RNP). Inversement, l’interférence ARN (ARNi) est une technologie de génie génétique qui, dans sa forme la plus simple, ne nécessite que la livraison d’oligonucléotides d’ARN interférent court purifié (siRNA). Les protéines endogènes transforment le siRNA double brin en brins simples, et la nucléase, le complexe de silençage induit par l’ARN (RISC), forme un complexe avec siRNA pour cliver les séquences d’ARNm cibles 15,16,17. Le silençage génique via cette méthode est transitoire en raison de l’instabilité de l’ARN, mais l’étude ici tire parti de cette fonctionnalité pour faciliter un dépistage rapide.
L’enzyme modèle sélectionnée pour la présente étude, l’α1,6-fucosyltransférase (FUT8), produit des N-glycanes avec α-1,6 L-fucose liée au noyau (Fuc). Cette modification est un déterminant primaire de l’activité de cytotoxicité cellulaire dépendante des anticorps (ADCC), comme en témoignent les études sur les anticorps commerciaux. En l’absence de fucosylation du noyau, le rituximab (IgG1 anti-CD20) multiplie par 50 l’ADCC et augmente l’ADCC dans le trastuzumab (IgG1 anti-Her2) en améliorantla liaison fcgRIIIa 18,19. La fucosylation de base est donc considérée comme une caractéristique indésirable des mAbs qui justifie des efforts pour inverser ce phénotype. Il existe des exemples de ciblage réussi du gène Fut8 à l’aide de siRNA avec des augmentations concomitantes de l’ADCC 20,21,22, bien que ces exemples fournissent du SiRNA Fut8 codé sur l’ADN plasmidique. De telles expériences génèrent un silençage génique stable car l’ADN plasmidique sert de modèle pour la synthèse du siRNA. Cela permet aux cellules de reconstituer les molécules de siRNA dégradées par les RNases intracellulaires et les phosphatases. Inversement, l’administration de siRNA synthétique exogène ne permet que le silençage transitoire des gènes, car le siRNA ne peut pas être reconstitué en raison de l’absence d’un modèle intracellulaire. Ainsi, les utilisateurs doivent déterminer si les plans expérimentaux sont compatibles avec le siRNA dérivé de plasmides ou synthétique. Par exemple, les études axées sur le pic de production de mAb, généralement le sixième jour de la culture23,24, peuvent opter pour un siRNA synthétique qui peut être livré aux cellules quelques jours avant le pic d’expression. Les avantages d’une approche transitoire utilisant du siRNA synthétique incluent la capacité d’externaliser la production et le fait que plusieurs constructions de siRNA peuvent être générées en une fraction du temps nécessaire pour générer des constructions dans les plasmides. En outre, le siRNA synthétique est efficace, comme en témoignent les exemples de silençage du gène Fut8 qui sont suffisants pour réduire l’expression de la protéine FUT825 et produire des IgG afucosylées avec une liaison accrue à FCgRIIIa et ADCC26.
Le succès de ce protocole de glyco-ingénierie a été déterminé par le degré de glycosylation Fc. La spectrométrie de masse est normalement la méthode de choix pour les analyses glycomiques; cependant, l’électrophorèse sur gel capillaire et la détection de fluorescence induite par laser (CGE-LIF) se prêtent parfaitement à la résolution du glycoprofil des IgG purifiées et présentent l’avantage d’une plus grande rapidité et simplicité. Les protocoles de spectrométrie de masse doivent combiner les méthodes de chromatographie et de dérivatisation appropriées, les sources d’ionisation et les analyseurs de masse 27,28,29. En plus de nécessiter un spécialiste qualifié, les protocoles de spectrométrie de masse sont longs et la diversité des méthodes rend les données difficiles à comparer entre les laboratoires avec des configurations différentes. Dans le contexte des produits biopharmaceutiques, le CGE-LIF est une méthode sensible qui peut fournir suffisamment de détails sur un glycoprofil d’anticorps et est facilement évolutive pour les méthodes à haut débit. Pour les mélanges à faible abondance et très complexes avec des glycoprotéines mal caractérisées, les avantages de la spectrométrie de masse pourraient demeurer. Cependant, l’analyse mAb haute résolution et haute sensibilité offerte par l’analyse N-glycane basée sur CGE-LIF sert de justification pour tester cette méthode. De plus, la préparation et l’analyse des échantillons sont terminées en quelques heures30 seulement. Des études récentes ont montré que le CGE-LIF peut être utilisé pour surveiller les glycanes dérivés du plasma humain31, de la souris32 et des IGG CHO33. Ces études mettent en évidence l’utilisation du CGE-LIF pour l’analyse d’échantillons à haut débit et de petits volumes d’échantillons.
La méthode CGE-LIF présente des limites à prendre en considération. Le coût est un obstacle important à l’utilisation de cet appareil et d’autres pour l’analyse des glycanes. Cependant, ces coûts sont typiques dans le domaine, et CGE-LIF est considéré comme une option rentable34. Les laboratoires disposant de petits budgets peuvent trouver plus pratique de louer des machines ou d’externaliser l’analyse d’échantillons. Une autre considération de toute méthode analytique est la répétabilité. L’évaluation du CGE-LIF a été réalisée à l’aide de 48 répétitions du même échantillon qui ont été analysées à des jours différents. L’écart-type relatif par capillaire a été déterminé pour la répétabilité intrabatch et interbatch. La comparaison intrabatch des réplicats s’est avérée avoir un écart-type relatif de 6,2%, ce qui indique que la performance capillaire n’est pas uniforme. De plus, une comparaison des données interbatch a montré un écart-type relatif de 15,8 %31, ce qui indique que le rendement capillaire change au fil du temps. Les lacunes opérationnelles identifiées peuvent ne pas s’appliquer dans la présente étude, qui utilise différentes machines et réactifs propriétaires. Si les utilisateurs ont l’intention de développer un protocole interne, il serait utile d’envisager l’étude de Ruhaak et al. 31, qui a soigneusement évalué les réactifs pour CGE-LIF. Ainsi, les réactifs pour l’injection d’échantillons (Hi-Di Formamide et DMSO), le marquage des glycanes (NaBH3CN ou 2-picoline borane)31, et d’autres ont été optimisés.
Cette étude présente un protocole de glycoingénierie rapide qui combine la rapidité de l’ARNi direct avec l’analyse glycomique en aval. La méthodologie est illustrée en utilisant le gène Fut8 comme cible pour les raisons décrites ci-dessus.
Protocole
1. Conception et reconstitution de l’ARNdsi
- Utilisez Safari, Firefox ou Microsoft Edge pour accéder au site Web IDT (https://eu.idtdna.com). Sur la page d’accueil, sélectionnez l’onglet Produits et services suivi de l’interférence ARN. Pour générer des constructions DsiRNA (dicer-substrates) personnalisées ciblant le gène Fut8 , sélectionnez Outil de conception suivi de l’onglet Générer un DsiRNA personnalisé .
- Entrez le numéro d’acquisition Fut8 ou entrez manuellement la séquence de gènes pour commencer. Dans cette étude, les séquences Fut8 proposées à partir des entrées « CHO-K1 » et « Hamster chinois » ont été obtenues à partir de https://chogenome.org et utilisées pour générer de l’ARNdsi. La plupart des gènes ont plusieurs variantes de transcription, et il est important de vérifier que l’ARNdsi serait actif sur toutes les variantes signalées dans l’assemblage actuel.
- Sélectionnez l’option pour effectuer une recherche « Manual BLAST » car le génome de la cellule CHO n’est actuellement pas disponible en tant que « génome de référence » sur le site Web de l’IDT. Cette étape recherche des correspondances entre les séquences d’ARNdS personnalisées et le génome d’intérêt.
- Cliquez sur Rechercher pour générer des séquences d’ARNds.
- À son tour, évaluez la spécificité de chaque DsiRNA à l’aide de la fonction « Manual BLAST » avec l’id fiscal: 10029 (lignées cellulaires CHO et hamster chinois). Les résultats de la couverture des requêtes indiquent que les constructions DsiRNA correspondent à Fut8 (y compris les variantes de transcription Fut8 ) à 100%, tandis que la complémentarité contre les cibles non intentionnelles est de ≤76%35.
- Vérifiez que le DsiRNA cible spécifiquement Fut8 pour vous assurer que la teneur en GC est comprise entre 30% et 50%. Une faible teneur en GC est associée à une liaison faible et non spécifique, tandis qu’une teneur élevée en GC inhibe le déroulement et la charge de siRNA dans le complexe RISC36.
- Sélectionner trois ARNdsi à utiliser dans les études de transfection (tableau 1), chacun ciblant un emplacement différent du gène Fut8 . Achetez un DsiRNA préconçu non ciblé ou brouillé auprès d’IDT pour des expériences de contrôle.
- À votre arrivée, centrifugez l’ARNdsi à 13 300 x g pendant 1 min pour granuler avant d’ouvrir le tube. Reconstituer chaque DsiRNA lyophilisé dans un tampon duplex sans nucléase (fourni par IDT) en une solution mère de 100 μM. Par exemple, ajoutez 20 μL de tampon à 2 nmol d’ARNdsi pour obtenir une souche de 100 μM.
- Pour assurer un mélange suffisant, incuber les solutions mères à température ambiante pendant 30 min tout en agitant doucement sur un agitateur orbital (50 tr/min, orbite de 16 mm).
- Créez un mélange maître en combinant 20 μL de chaque construction d’ARNdsi qui cible Fut8 (100 μM de chaque DsiRNA). Préparer les aliquotes et conserver à −20 °C.
| Cible DsiRNA | Séquence | GC (%) | ||
| Structure A | 5' GAGAAGAUAGAAACAGUCAAAUACC 3' | 36% | ||
| 5' GGUAUUUGACUGUUUCUAUCUUCUCUC 3' | ||||
| Structure B | 5' AGAAUGAGAAUGGAUGUUUUUCCTT 3' | 32% | ||
| 5' AAGGAAAAACAUCCAUUCUCAUUCUGA 3' | ||||
| Structure C | 5' AGAGAAGAUAGAAACAGUCAAAUAC 3' | 32% | ||
| 5' GUAUUUGACUGUUUCUAUCUUCUCUCG 3' | ||||
Tableau 1. Séquences d’ARNdi utilisées pour l’élimination de Fut8. Séquences générées par IDT qui ciblent Fut8 dans les génomes des hamsters chinois et des cellules CHO K1. Les séquences de sens et d’antisens pour chaque construction sont affichées (respectivement), et le contenu GC de chaque structure est affiché. Reproduit à partir de Kotidis et al. 52.
2. Transfection d’ARNdsi
- Raviver les cellules CHO exprimant un anticorps monoclonal IgG37 en utilisant des conditions de culture adaptées à la lignée cellulaire d’intérêt.
REMARQUE: Les cellules utilisées dans cette étude ont été générées à l’aide du système de glutamine synthétase (GS), où le GS endogène est inhibé par la L-méthionine sulfoximine (MSX) pour améliorer la sélection de clones rares à haute production. Par conséquent, n’utilisez MSX que si la lignée cellulaire de votre choix l’exige.- Décongeler un flacon de cellules pendant 2-3 min dans un bain-marie réglé à 37 °C. Nettoyez l’extérieur du flacon avec de l’éthanol à 70 % (v/v) et continuez tous les travaux dans une armoire de biosécurité de classe II.
- Transférer la suspension cellulaire dans un tube centrifuge de 15 mL contenant 9 mL de milieu préavertis approprié à la lignée cellulaire de choix. Granulés les cellules par centrifugation à 100 x g pendant 5 min.
- Retirez et jetez soigneusement le milieu sans perturber la pastille de cellule. Ensuite, remettez en suspension la pastille cellulaire dans 10 mL de milieu préavertissé et prenez une aliquote pour le comptage.
- Colorez les cellules avec du bleu Trypan si vous comptez avec un hémocytomètre, ou une coloration appropriée pour distinguer les cellules vivantes / mortes lors de l’utilisation d’un compteur de cellules automatisé.
- En suivant l’une ou l’autre méthode de dénombrement, transférer un volume approprié de suspension cellulaire dans une fiole agitée d’Erlenmeyer de 125 mL à une densité cellulaire viable de 3 x 105 cellules·mL-1 dans 30 mL de milieu (avec supplémentation facultative de 50 μM MSX).
- Transférer les cellules dans un incubateur réglé à 36,5 °C, 5 % de CO2 et les placer sur une plate-forme de secousse à 150 tr/min (orbite de 16 mm).
- Cellules de passage tous les 3-4 jours à une densité d’ensemencement de 2 x 105 cellules·mL-1 et un volume de travail de 50 mL dans une fiole agitée Erlenmeyer de 250 mL. Si vous utilisez MSX, arrêtez la supplémentation après le premier passage.
- Cellules de passage 2x en plus de la décongélation.
- Transfection
- Évaluez la densité cellulaire et assurez-vous que la viabilité cellulaire est supérieure à 90 %.
- Nettoyez soigneusement l’armoire de biosécurité et tout l’équipement avec de l’éthanol à 70 % (v/v) et une solution d’inhibiteur de la RNase pour éviter toute contamination.
- Cellules à granulés à 100 x g pendant 5 min et remises en suspension dans un milieu préavertissé à une densité cellulaire viable de 5 x 106 cellules·mL-1.
- Transférer 8 μL (équivalent à 1 μM) du mélange maître ou du témoin DsiRNA sur une cuvette d’électroporation stérile (0,4 cm). Ensuite, transférez 800 μL de la suspension cellulaire (équivalent à 4 x 106 cellules) dans la même cuvette et assurez-vous que les deux composants sont mélangés.
- Fournir les conditions d’impulsion suivantes: 1200 V, 0,1 ms, forme d’onde carrée.
- Transférer la suspension cellulaire de la cuvette dans un puits d’une plaque à 6 puits, en prenant soin d’éviter tout matériau semblable à de la mousse. Récupérer les cellules dans l’incubateur (36,5 °C, 5% CO2) sans secouer pendant 10 min.
- Ajouter 800 μL de milieu préavertis pour obtenir un volume final de 1,6 mL par puits et renvoyer les cellules transfectées à l’incubateur pour la croissance (36,5 °C, 5 % de CO2) tout en secouant à 150 tr/min (orbite de 16 mm).
- Récoltez les surnageants et les cellules 48 h après la transfection.
Point d’arrêt : Le surnageant de culture cellulaire peut être maintenu à -20 °C ; cependant, il est conseillé d’effectuer la lyse cellulaire immédiatement après la collecte des granulés cellulaires afin d’éviter la dégradation des protéines. Les surnageants sont utilisés aux étapes 3, 4 et 5, tandis que les granulés cellulaires sont utilisés à l’étape 6.
3. Quantification et purification des IgG
- Mesurer la concentration d’IgG à l’aide de l’interférométrie de biocouche ou d’une autre méthode de choix.
- Protéine hydratée Un biocapteur pointe dans l’échantillon de diluant pendant 10-30 min. En attendant, transférez les suspensions cellulaires dans des tubes centrifuges de 15 mL et des cellules à granulés à 100 x g pendant 5 min ou retirez les cellules par filtration à travers un filtre à nitrocellulose de 0,45 μm.
- Sans perturber la pastille, transférez soigneusement le surnageant isolé contenant des IgG dans un tube de centrifugeuse propre.
- Utilisez les paramètres suivants pour l’interférométrie de biocouche : vitesse de l’agitateur, 2200 tr/min ; durée d’exécution, 60 s. Ces paramètres seront utilisés pour mesurer tous les échantillons et les contrôles.
- Une fois les extrémités du biocapteur hydratées, créez une courbe standard à l’aide d’un étalon IgG (4 μL de chaque concentration).
- Quantifier les concentrations de l’échantillon en chargeant 4 μL du surnageant cellulaire. Nettoyez l’appareil avec des lingettes non pelucheuses entre chaque échantillon.
- Liez la courbe standard aux échantillons inconnus pour interpoler le taux de liaison des échantillons inconnus. Enregistrez les données et exportez-les sous forme de fichier csv ou PDF.
- Purification des IgG
- Préparer un tampon d’élution contenant 0,2 M de glycine (pH 2,5) et stériliser en passant à travers un filtre de 0,22 μm.
- Filtrer 1 mL du surnageant isolé à température ambiante à l’aide de tubes filtrants de microcentrifugeuse de 0,22 μm jusqu’à ce que le surnageant entier s’écoule.
- Granulé 150-200 μL de perles d’agarose de protéine A dans des tubes centrifuges de 1,5 mL à 100 x g pendant 3 min et jeter le surnageant. Ensuite, lavez les billes de protéine A avec 150-200 μL de milieu de culture cellulaire et répétez la centrifugation. Jetez le surnageant.
- Remettre en suspension les billes de protéine A équilibrées avec 1 mL des surnageants préparés de l’étape 3.2.3. et charger dans un tube en polypropylène de 1 mL. Fixez le tube en polypropylène à un mélangeur rotatif (orbite de 15 mm) et tournez à 30 tr/min pendant 60 à 90 minutes à température ambiante ou pendant la nuit à 4 °C.
- Une fois l’incubation terminée, recueillir le flux et laver les billes avec 1 mL de 1x PBS pour éliminer les protéines non liées. Collectez également la fraction de lavage.
- Éluez les IgG des billes de protéine A en ajoutant 3 mL de tampon d’élution à la colonne de polypropylène. Recueillir les fractions séquentielles qui s’écoulent de la colonne (500 μL chacune) dans des tubes étiquetés.
- Facultatif : Si l’anticorps purifié doit être stocké, neutraliser le tampon d’élution en ajoutant 25 μL de 1 M Tris pH 9,5. Sautez cette étape si l’anticorps sera traité immédiatement via un échange de tampon, etc.
REMARQUE: Toutes les parties de la purification (écoulement, lavages et toutes les fractions d’élution) doivent être conservées en veillant à ce que la protéine cible ne soit pas perdue. Ces exemples peuvent également aider lors du dépannage si la purification échoue. - Utilisez la première élution (élution A) pour le traitement en aval, mais conservez toutes les autres fractions si elles sont nécessaires à l’avenir.
4. Échange de tampons et concentration de l’échantillon
- Tampon d’élution IgG Exchange avec 1x PBS.
- Charger l’élution A sur un concentrateur centrifuge de coupure de poids moléculaire de 3 kDa. Reportez-vous au guide du fabricant lorsque vous sélectionnez une colonne MWCO appropriée. Dans cette expérience, une taille de pore plus petite assure une rétention maximale des protéines sécrétées, mais augmente également le temps de centrifugation.
- Centrifuger à 13 300 x g pendant 40-50 min à 4 °C. La centrifugation est terminée lorsque le volume résiduel est égal ou inférieur à 50 μL.
- Jeter l’écoulement et ajouter 500 μL de PBS précraissé (4 °C) 1x pour diluer le surnageant résiduel. Centrifuger à nouveau l’échantillon dans les mêmes conditions (13 300 x g pendant 40-50 min à 4 °C) jusqu’à ce qu’il reste 50 μL de surnageant résiduel.
- Ajouter 500 μL de PBS précroché (4 °C) pour diluer le surnageant résiduel et répéter le processus de centrifugation pour obtenir une dilution de 100x du surnageant.
- Concentrer le surnageant à une concentration finale de ~2,5 g· L-1 dans 40 μL ou moins pour assurer la compatibilité avec la méthode d’analyse des glycanes.
REMARQUE: 100 μg doivent être chargés pour l’analyse du glycane.
Point d’arrêt : Les IgG concentrées (dans 1x PBS) peuvent être stockées à −20 °C et décongelées avant l’analyse du glycane.
5. Analyse des glycanes
- Effectuer une analyse du glycane à l’aide de l’électrophorèse sur gel capillaire conformément aux instructions du fabricant.
- Transférer 200 μL de la solution de billes magnétiques dans un tube PCR de 0,2 mL et placer sur le support magnétique pour séparer les billes du surnageant.
- Retirez soigneusement le surnageant et retirez les tubes du support magnétique. Ajouter l’échantillon de protéine purifiée et le vortex pour assurer un mélange complet.
- Ajouter le tampon de dénaturation (fourni) au tube d’échantillonnage et incuber pendant 8 min à 60 °C. Gardez les tubes d’échantillonnage ouverts pour des performances de réaction optimales.
- Ajouter PNGase F (500 unités par échantillon) et incuber pendant 20 min à 60 °C pour cliver les glycanes des anticorps purifiés.
- Après la libération de N-glycanes, fermez le tube d’échantillonnage et le vortex. Ajouter l’acétonitrile, le vortex et incuber à température ambiante pendant 1 min.
- Placez les tubes d’échantillonnage dans le support magnétique pour séparer les billes de la solution. Utilisez une pipette pour retirer soigneusement le surnageant sans toucher les perles.
- Dans une hotte aspirante, ajouter à l’échantillon la solution d’étiquetage du glycane contenant un fluorophore. Vortex pour assurer un mélange suffisant et incuber à 60 °C pendant 20 min (couvercles ouverts).
- Lavez l’échantillon 3x dans l’acétonitrile pour éliminer l’excès de colorant. Ensuite, éluez les glycanes étiquetés dans de l’eau DDI (fournie).
- Placez le tube d’échantillonnage dans le support magnétique pour séparer les perles du surnageant contenant des glycanes purifiés et étiquetés.
- Préparez et chargez l’étalon de l’échelle de glucose, les étalons de bracketing et les échantillons dans les positions de plateau désignées. Exécutez le protocole d’analyse des glycanes.
- Utilisez un logiciel approprié pour analyser et identifier les glycanes présents dans l’échantillon.
REMARQUE: Assurez-vous que la température dans le bloc thermique est précise pour une incubation efficace.
6. Transfert western
- Quantifier l’efficacité de l’élimination à l’aide de l’analyse par transfert western avec un anticorps anti-α-1,6-fucosyltransférase. Les gels de polyacrylamide et les recettes tampons sont disponibles auprès des fournisseurs commerciaux38,39.
- À 48 h après la transfection, comptez les cellules à l’aide d’un hémocytomètre ou d’un compteur cellulaire automatisé et déterminez le volume de suspension cellulaire équivalent à 5 x 106 cellules.
- Transférer le volume approprié de suspension cellulaire de chaque condition expérimentale dans un tube de centrifugeuse stérile de 1,5 mL et une pastille à 13 200 x g pendant 10 min à 4 °C. Jetez le surnageant.
- Lyser les cellules avec 200 μL de tampon de lyse (adapté à l’extraction de protéines de cellules de mammifères) contenant un cocktail d’inhibiteurs de protéase de 1% v/v à température ambiante pendant 10 min. Agiter doucement le mélange pendant l’incubation (50 tr/min, orbite de 16 mm).
- Pour éliminer les débris cellulaires, centrifugez le lysat à 13 200 x g pendant 10 min, puis transférez le lysat éliminé dans un tube stérile de 1,5 mL.
- Mesurer la concentration en protéines de chaque lysat à l’aide d’un spectrophotomètre à 280 nm. Par la suite, préparez des aliquotes de chaque échantillon ajustées pour avoir la même concentration en protéines.
- Dénaturer les échantillons de protéines par incubation à 100 °C pendant 10 à 15 min dans un colorant de chargement d’échantillons DTT-SDS; la concentration finale de colorant est de 1x.
- Chargez 15 μL des échantillons dénaturés et 5 μL d’échelle protéique précolorée dans un gel SDS-PAGE avec 12,5% de gel résolvant pour une séparation efficace. Faites couler les échantillons à 25 mA par gel pendant 90 min ou jusqu’à ce que le front de teinture atteigne la fin du gel.
- Retirez soigneusement le gel de la cassette et incubez dans 1x tampon de transfert contenant du méthanol.
- Préparez le système de transfert humide et activez une membrane PVDF avec du méthanol. Assemblez le gel et la membrane PVDF pour le transfert humide, placez un bloc de glace dans le réservoir de transfert et immergez l’ensemble du réservoir dans la glace pour vous assurer que les conditions de transfert restent froides. Fonctionner à 350 mA/100 V pendant 60 min.
- Bloquez la membrane en incubant dans une solution bloquante pendant 30 min à température ambiante tout en secouant doucement sur un agitateur orbital à 50 tr/min (orbite de 16 mm).
- Rincez la membrane à l’eau stérile pendant 5 min et répétez. Ensuite, utilisez un scalpel propre pour couper soigneusement la membrane horizontalement à ~ 50 kDa, en utilisant l’échelle de protéines visible comme guide.
- Incuber la membrane hébergeant des protéines supérieures à 50 kDa avec un anticorps anti-α-1,6-fucosyltransférase à 1:1000 dans un tampon diluant anticorps. Incuber la membrane avec des protéines immobilisées inférieures à 50 kDa avec de l’anti-GAPDH à 1:10 000 dans un tampon diluant. Les membranes peuvent être incubées pendant 1 h à température ambiante ou pendant la nuit à 4 °C.
- Lavez les membranes pendant 5 min (x3), puis incubez avec un anticorps secondaire approprié pendant au moins 30 min à température ambiante.
- Effectuez 3x lavages avec un tampon de lavage pendant 5 min, suivis de 3x lavages à l’eau. Ensuite, développez la membrane avec un substrat chromogène (ou un réactif de détection approprié) jusqu’à l’apparition de bandes (1-60 min).
- Rincez la membrane 2x avec de l’eau, puis laissez-la sécher. Capturez une image de la membrane et effectuez une analyse densitométrique40.
- Pour calculer l’expression relative des protéines dans chaque échantillon, calculez d’abord le rapport du signal GAPDH dans les échantillons. Il s’agit du facteur de normalisation pour chaque échantillon qui corrige les écarts dans le chargement de l’échantillon.

- Divisez l’intensité du signal FUT8 (pour chaque voie) par le facteur de normalisation de la voie correspondante pour obtenir l’expression relative de la protéine FUT8.

Résultats
L’analyse par transfert western a montré une réduction de l’expression de la protéine FUT8 dans les cellules transfectées avec un mélange de trois constructions d’ARNds Fut8 . Dans les échantillons témoins transfectés avec de l’ARNdsi non ciblé, FUT8 est apparu comme une double bande à ~65 et 70 kDa. Étant donné que le poids moléculaire prédit de FUT8 est de 66 kDa, une réduction de l’intensité du signal de la bande de poids moléculaire inférieure est révélatrice d’un silençage génique. Pour confirmer et quantifier le silençage génique, le niveau de protéine FUT8 a été normalisé au niveau relatif de la protéine GAPDH. L’analyse par transfert Western a détecté deux bandes pour LE GAPDH à ~37 et 35 kDa. La bande de poids moléculaire plus élevée correspond à la taille prédite des protéines et est donc utilisée dans les calculs de normalisation. Lorsqu’elle est normalisée par rapport aux niveaux de protéines GAPDH, l’expression de la protéine FUT8 a été réduite jusqu’à 60 % (Figure 1).
Conformément à l’observation de l’élimination des gènes à 48 h après la transfection, les échantillons mAb correspondants ont été traités pour analyse par CGE-LIF. Les structures glycanes des cellules knockdown ont montré une diminution de la fucosylation. Cette tendance était plus prononcée dans les structures agalactosylées (G0F) et observée dans une moindre mesure dans les structures galactosylées (G1F, G1F' et G2F). À partir de cet ensemble de données, la fucosylation totale du noyau IgG a diminué à ~75 %, en baisse par rapport à ~95 % de fucosylation du noyau observée pour la condition de contrôle négative (Figure 2). Une plus grande réduction de la fucosylation du noyau était prévue compte tenu de la diminution d’environ 60% des niveaux de protéines FUT8. Après réflexion, il convient de noter que le glycoprofil représente des mAbs glycosylés qui se sont accumulés sur une période de 48 h depuis la transfection, tandis que le silençage génique représente les niveaux de protéines présents au moment de la récolte seulement.
Un examen plus approfondi de cette méthode d’élimination impliquait de faire varier la concentration d’ARNdsi, le temps de récolte et les conditions d’électroporation. Chaque facteur a été examiné individuellement pour déterminer sa pertinence. L’impact des conditions d’impulsion d’électroporation sur la fucosylation du noyau et la viabilité cellulaire est capturé dans les expériences B, C, D et E. Ces résultats démontrent une réduction de deux fois de la fucosylation du noyau due à l’électroporation à l’aide de deux impulsions d’onde carrées (expérience C) par rapport à une seule impulsion d’onde carrée (expérience B), sans différences significatives dans la viabilité cellulaire (tableau 2). La condition d’électroporation e3 (expérience D) a conduit à la viabilité cellulaire la plus faible (~ 90%) et au rendement en IgG à ce moment-là. Cependant, les cellules qui ont survécu à l’événement d’électroporation ont été modérément transfectées, comme en témoigne la diminution d’environ 10% de la fucosylation du noyau (tableau 2). Fait intéressant, l’expérience D a utilisé des conditions d’électroporation qui ont fourni la plus grande réduction de la fucosylation du noyau (14,7%), mais qui étaient manifestement préjudiciables à la viabilité cellulaire (viabilité de 91% à 93%). Cet ensemble limité d’expériences illustre la nécessité de déterminer les paramètres d’électroporation qui permettent une perméabilisation suffisante de la membrane cellulaire sans causer de dommages irrévocables. Il est également intéressant de noter le rôle de la concentration en siRNA et du temps de récolte sur la fucosylation du noyau. Dans l’ensemble, l’augmentation de la concentration de siRNA a une plus grande influence sur la fucosylation du noyau que l’augmentation du temps de récolte (expériences B, F, G par rapport aux expériences A, B, H). Dans de futures expériences, il serait intéressant de titrer les concentrations de siRNA délivrées par la méthode d’électroporation e2.

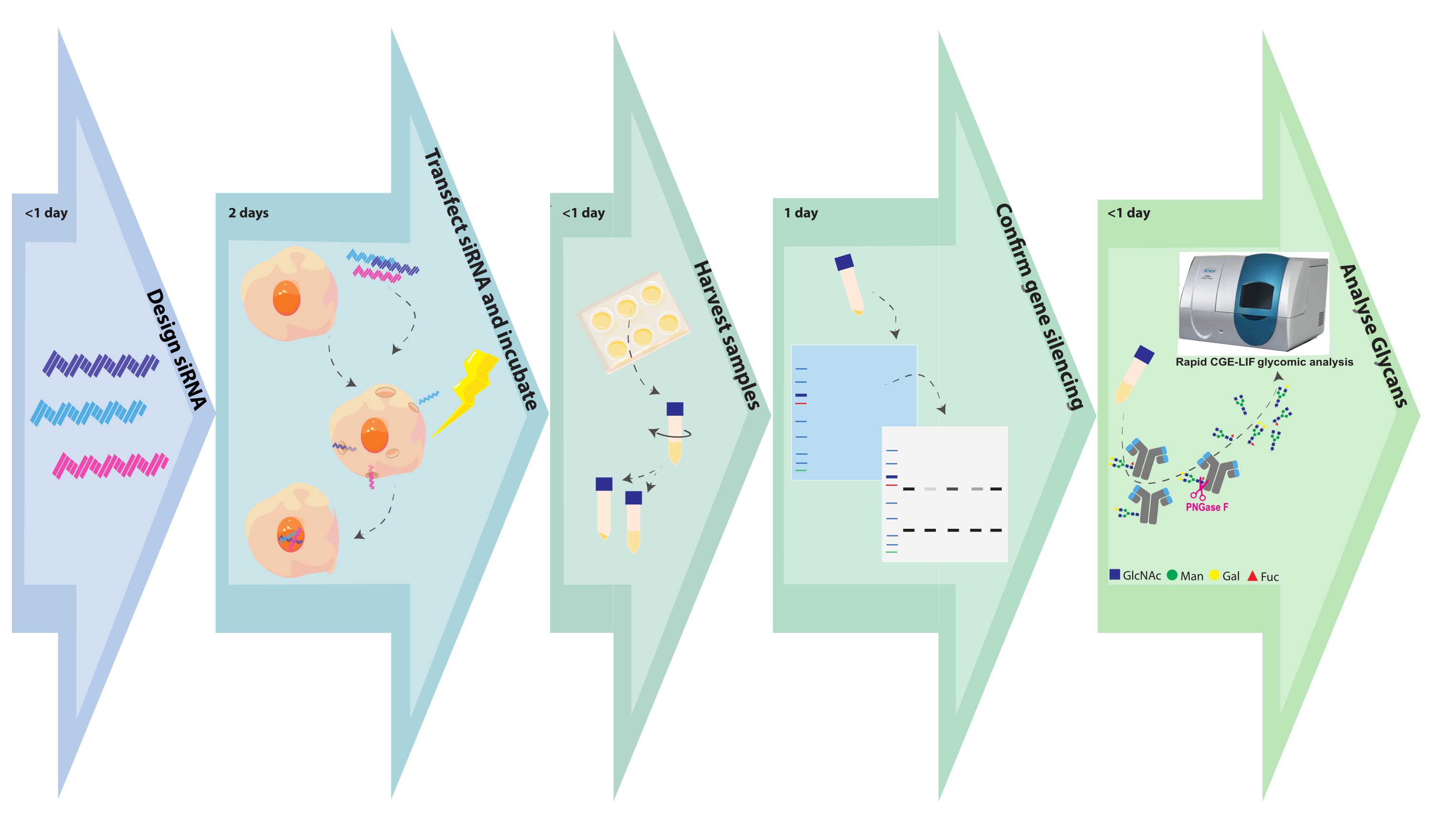
Graphique 1. Organigramme expérimental. Les étapes de glyco-ingénierie et d’analyse des échantillons sont représentées avec le temps associé requis pour chaque étape. La conception de SiRNA prend quelques heures, en fonction du nombre de cibles ou de constructions de gènes par cible de gène. La transfection des cellules CHO avec siRNA est terminée en quelques heures et les cellules transformées sont laissées à croître pendant 48 h. Les granulés cellulaires et les surnageants sont récoltés en quelques heures. Les pastilles cellulaires sont lysées et les protéines intracellulaires sont séparées sur un SDS PAGE, puis effacées et sondées avec des anticorps contre le gène cible. Les glycanes sont clivés des anticorps purifiés et analysés par CGE-LIF. Ces tests peuvent nécessiter 1 jour chacun. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Graphique 2. Confirmation de l’interférence ARN. Détection par transfert western des niveaux de protéines α-1,6-fucosyltransférase (FUT8) dans des échantillons traités avec Fut8 ou avec de l’ARNdsi témoin non ciblé. Les bandes correspondant à FUT8 sont plus intenses en contrôle que les échantillons de Knockdown Fut8. Le niveau de protéine GAPDH a également été évalué afin de normaliser l’expression du gène cible. Tous les échantillons ont été prélevés à partir de l’expérience G (voir le tableau 2). Reproduit à partir de Kotidis et al. 52. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Graphique 3. Effet de Fut8 knockdown sur la glycosylation cumulative des IgG à 48 h. Un changement dans la distribution des glycanes est détecté dans les échantillons de knockdown. En particulier, l’abondance relative des principales structures fucosylées du noyau (G0F) est réduite tandis que les espèces afucosylées sont augmentées dans l’expérience de knockdown. Les mesures ont été effectuées à partir d’échantillons de l’expérience G (voir le tableau 2). Les triples biologiques effectués pour chaque expérience ont été mélangés après la récolte afin de réduire le fardeau de l’analyse en aval. Reproduit à partir de Kotidis et al. 52. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
| Nom de l’expérience | Méthode d’électroporation | Concentration d’ARNdsi (nΜ) | Temps de récolte (h) | Viabilité (%) | Xv (106 cellules·mL-1) | Titre IgG (mg· L-1) | Différence dans la fucosylation du noyau (%) | ||||||
| ExpA_Negative | e1 | 500 | 24 | 98.3 | 4.71 | 122.5 | - | ||||||
| ExpA_Knockdown | e1 | 500 | 24 | 98.3 | 4.9 | 110.3 | 4.08 | ||||||
| ExpB_Negative | e1 | 500 | 48 | 95.6 | 9.55 | 453.3 | - | ||||||
| ExpB_Knockdown | e1 | 500 | 48 | 96.7 | 9.61 | 469 | 5.38 | ||||||
| ExpC_Negative | e2 | 500 | 48 | 96.3 | 9.91 | 449.3 | - | ||||||
| ExpC_Knockdown | e2 | 500 | 48 | 96.7 | 11 | 454.6 | 11.42 | ||||||
| ExpD_Negative | e3 | 500 | 48 | 90.6 | 6.25 | 318.5 | - | ||||||
| ExpD_Knockdown | e3 | 500 | 48 | 89.1 | 6.09 | 311.85 | 9.71 | ||||||
| ExpE_Negative | e4 | 500 | 48 | 91.1 | 7.2 | 380.3 | - | ||||||
| ExpE_Knockdown | e4 | 500 | 48 | 93.3 | 7.79 | 422.8 | 14.7 | ||||||
| ExpF_Negative | e1 | 750 | 48 | 96.2 | 9.7 | 501 | - | ||||||
| ExpF_Knockdown | e1 | 750 | 48 | 95.7 | 9.76 | 504.6 | 9.9 | ||||||
| ExpG_Negative | e1 | 1000 | 48 | 96.1 | 11.1 | 422.6 | - | ||||||
| ExpG_Knockdown | e1 | 1000 | 48 | 95.9 | 9.73 | 499.3 | 17.26 | ||||||
| ExpH_Negative | e1 | 500 | 72 | 94.4 | 14.3 | 925.8 | - | ||||||
| ExpH_Knockdown | e1 | 500 | 72 | 95 | 13.5 | 1018.4 | 7.37 | ||||||
Tableau 2. Optimisation de la transfection. Les modifications itératives de la méthode d’électroporation, de la concentration d’ARNdsi et du temps de récolte ont entraîné des changements dans la viabilité cellulaire, la densité cellulaire viable, le titre d’IgG au moment de la récolte et des différences dans la fucosylation du noyau. Chaque expérience a comparé le knockdown et le contrôle négatif respectif pour déterminer si la modification produit l’effet souhaité (c’est-à-dire une diminution de la fucosylation). Les réglages d’électroporation étaient les suivants: e1: 1200 V, 0,1 ms, forme d’onde carrée; e2: 1200 V, 2x 0,1 ms, 5 s entre les impulsions, forme d’onde carrée; e3: 150 V, 20 ms, forme d’onde carrée; e4 : 250 V, 500 μF, désintégration exponentielle. Reproduit à partir de Kotidis et al. 52.
Discussion
Les voies de glycosylation impliquent un réseau métabolique complexe d’enzymes et de protéines accessoires. Disséquer la fonction des constituants de la voie est intimidant s’il dépend uniquement des stratégies conventionnelles de génie génétique knockin ou knockin. Une autre approche consiste à dépister au préalable les membres d’une voie à l’aide d’un test transitoire de perte de fonction. À cette fin, deux protocoles rapides ont été combinés, la détection de l’ARNi et du CGE-LIF, afin de créer un moyen plus efficace de caractériser les gènes de glycosylation. La méthode décrite nécessite 5 à 7 jours pour l’achèvement par rapport aux méthodes conventionnelles qui prennent potentiellement plusieurs semaines pour l’achèvement. En outre, les environnements de recherche dotés de capacités d’automatisation pourraient exploiter cette méthode pour dépister plus de gènes candidats que possible avec une manipulation manuelle.
Le succès d’une campagne de glyco-ingénierie transitoire dépend en grande partie de la conception du siRNA. Les conceptions DsiRNA personnalisées doivent suivre les règles décrites précédemment, ou pour plus de facilité, les utilisateurs peuvent opter pour des séquences prédéfinies disponibles dans le commerce. Comme d’autres stratégies de modification génétique, l’ARNi a le potentiel d’avoir des effets hors cible. Par conséquent, les utilisateurs sont encouragés à évaluer le ciblage involontaire des gènes par des méthodes de calcul41. Les choix de conception expérimentale peuvent également aider à limiter les effets hors cible. Kittler et al. ont montré que l’administration multiplexée de siRNA entraînait une réduction des effets hors cible42. Bien que cela semble contre-intuitif, il est suggéré qu’un mélange maître réduit la concentration de chaque construction de siRNA, limitant ainsi la possibilité de silençage génique hors cible. Un autre avantage est que la transfection simultanée des structures de siRNA qui ciblent le même gène augmente la probabilité de succès de l’ARNi. L’utilisation d’un mélange maître assure également la cohérence entre les échantillons et les répliques et accélère le processus de transfection. Après un criblage initial de constructions mixtes de siRNA, une autre expérience peut être menée en utilisant des constructions individuelles pour déterminer l’efficacité de l’ARNi de chaque séquence. Dans cette étude et d’autres, jusqu’à trois siRNA ont été regroupés et livrés aux cellules 43,44,45. Cependant, il peut être souhaitable de dépister plus de trois siRNA simultanément pour cibler efficacement un seul gène ou plusieurs gènes. En effet, une étude a démontré un silence multiplexé médié par le siRNA de jusqu’à six gènes à des niveaux comparables au silence de gènes individuels46. Cependant, d’autres études sont nécessaires pour déterminer le nombre maximal de constructions de siRNA pouvant être utilisées dans un pool sans compromettre l’efficacité du silencieux. La stratégie multiplex a été proposée par Martin et al. pour améliorer le rythme des expériences de criblage de bibliothèque d’ARNi46, et un concept similaire pourrait s’avérer utile pour dépister les gènes de glycosylation.
Le protocole décrit ici sert de preuve de concept avec l’espoir que des expériences ultérieures seront effectuées pour valider d’autres gènes de glycosylation. Les nouveaux gènes d’intérêt peuvent être non caractérisés ou moins populaires que Fut8, et les anticorps primaires pour détecter le silençage génique peuvent être médiocres ou indisponibles. Dans ce scénario, d’autres méthodes telles que la RT-PCR peuvent être utilisées pour quantifier le silençage génique47, mais il convient de noter que la RT-PCR détecte l’ARNm plutôt que les protéines. Lorsque des anticorps pour le transfert western sont disponibles, un problème courant est une mauvaise détection ou la présence de bandes non spécifiques. Des guides de dépannage pour aider les utilisateurs à résoudre les problèmes courants sont disponibles, et ceux-ci ont tendance à inclure une gamme de solutions telles que le titrage primaire des anticorps, le blocage alternatif et les conditions de détection48,49. Dans cette étude, FUT8 est apparu de manière inattendue sous la forme d’une double bande à ~65 et ~70 kDa. Il est possible que la bande ~70 kDa représente fut8 glycosylé. Les preuves de la littérature provenant de lignées cellulaires humaines décrivent la glycosylation liée à l’O à Thr 564 50,51, un site conservé dans le hamster chinois, et les séquences CHO K1 FUT8.
Comme mentionné précédemment, les voies de glycosylation impliquent souvent un ensemble complexe d’enzymes. Le protocole actuel a été développé, optimisé et démontré à l’aide d’une réaction de glycosylation monogénique contrôlée par Fut8. Par conséquent, d’autres études sont nécessaires pour confirmer la robustesse de cette méthode lorsque le gène cible code pour une enzyme avec des niveaux alternatifs de cinétique et d’expression ou une voie régulée par des isoenzymes aux fonctions redondantes.
Dans l’ensemble, la capacité de faire taire rapidement les gènes et de détecter les glycoprofils IgG modifiés est un outil utile dans l’effort visant à obtenir des anticorps glyco-modifiés personnalisés. Les résultats d’études similaires à court terme peuvent être appliqués pour générer des cellules glyco-modifiées stables à utiliser dans des essais à long terme tels que la culture par lots nourris. En dehors du contexte pharmaceutique, cette méthode contribue à l’étude de la biologie des glycanes et met en évidence la fonction importante des glycanes dans le développement, la santé et la maladie.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
PK remercie le Département de génie chimique de l’Imperial College de Londres pour sa bourse. RD remercie le Conseil de recherches en biotechnologie et en sciences biologiques du Royaume-Uni pour sa bourse d’études. LE MM est financé par le Conseil de recherches en biotechnologie et en sciences biologiques du Royaume-Uni (référence de la subvention : BB/S006206/1). L’IAG remercie l’Irish Research Council (Scholarship No. GOIPG/2017/1049) et CONACyT (Bourse n° 438330).
matériels
| Name | Company | Catalog Number | Comments |
| 32 Karat software | SCIEX | contact manufacturer | Software for glycan data acquisition and analysis using the Fast Glycan analysis protocol and separation method. |
| Acetonitrile, HPLC grade | Sigma Aldrich | 34851 | Solvent. |
| Anti-FUT8 antibody | AbCam | ab198741 | Rabbit polycloncal to Fut8. Use this antibody to quantify Fut8 protein expression; replace this antibody if using siRNA targeting a different gene. |
| Anti-GAPDH antibody | AbCam | ab181602 | Rabbit monoclonal to GAPDH. Alternative housekeeping genes exist and might be preferred by the user. |
| BioDrop Spectrophotometer | Biochrom | 80 3006 55 | Instrument used to quantify protein concentration. |
| BLItz | ForteBio | 45 5000 | Instrument. Label-Free Protein Analysis System. |
| BRAND Haemocytometer | Sigma Alrich | BR717810 | Counting chamber device |
| Capillary cartridge | SCIEX | A55625 | Pre-assembled capillary cartridge with window (30 cm total length, 375 µm outer diameter (o.d), x 50 µm inner diameter (i.d). |
| C100HT Glycan analysis—capillary electrophoresis | SCIEX | contact manufacturer | Capillary gel electrophoresis instrument, the CESI 8000 Plus instrument is now used. |
| CD CHO Medium | Thermo Fisher Scientific | 10743029 | Replace this with a culture medium appropriate for the cell line of choice. |
| Centrifuge tubes, 15 mL | Greiner Bio | 188261 | Sterile polypropylene tube. |
| Centrifuge tubes, 50 mL | Greiner Bio | 227270 | Sterile polypropylene tube. |
| CHO IgG | MedImmune | Gift | Chinese Hamster Ovary cells expressing an IgG monoclonal antibody (CHO T127). Created using the GS system. |
| Dulbecco's phosphate-buffered saline (DPBS) | Gibco | 14190144 | 1x PBS, without calcium or magnesium. |
| Erlenmeyer Flasks with Vent Cap, 125 mL | Corning | 431143 | Replace this with a culture vessel suitable for growing the cell line of choice. |
| Erlenmeyer Flasks with Vent Cap, 250 mL | Corning | 431144 | Replace this with a culture vessel suitable for growing the cell line of choice. |
| Fast Glycan Labelling and Analysis kit | SCIEX | B94499PTO | Labels N-glycans with APTS and then uses a magnetic-bead based clean up system to remove excess APTS. |
| Fut8 DsiRNA | IDT | Custom | Custom designed DsiRNA targetting Fut8. |
| Gene Pulser cuvettes, 0.4 cm | Bio-Rad | 1652088 | Electroporation cuvette. |
| Gene Pulser Xcell Eukaryotic System | Bio-Rad | 165 2661 | Insturment. Xcell main unit with Capacitance Extender (CE) Mocdule and ShockPod. |
| Immobilon-FL PVDF membrane | Merck-Millipore | IPFL00010 | Immunoblot transfer membrane, low background. |
| L-Methionine sulfoximine (MSX) | Sigma Aldrich | M5379 | Only necessary for CHO cell lines using the glutamine synthetase (GS) selection system. |
| Kimwipes | Thermo Fisher Scientific | 10623111 | Low-lint, high absorbency and chemically inert wipes. |
| M-PER Mammalian Protein Extraction Reagent | Thermo Fisher Scientific | 78505 | Alternative lysis buffers such as RIPA are also appropriate. |
| Methanol, HPLC grade | Fisher Scientific | 10365710 | Solvent. |
| Microcentrifuge tubes, 1.5 mL | Eppendorf | 616201 | Autoclavable tubes. |
| Mini-PROTEAN Tetra Vertical Electrophoresis Cell system | Bio-Rad | 1658035FC | Instrument. 4-gel capacity, for 1.0 mm thick handcast gels, with Mini Trans-Blot Module and PowerPac HC Power Supply. |
| NC-Slide A8 | ChemoMetec | 942 0003 | 8-chamber slide for use with NucleoCounter NC 250. |
| Negative Control DsiRNA, 5 nmol | IDT | 51 01 14 04 | Non-targeting DsiRNA. |
| Nuclease-free duplex buffer | IDT | 11-01-03-01 | Reconstitution buffer for DsiRNA. |
| NucleoCounter NC-250 | Chemometec | contact manufacturer | Instrument. Automated Cell Analyzer |
| Page-Ruler ladder, 10 to 180 kDa | Thermo Fisher Scientific | 26616 | Mixed blue, orange and green protein standards for SDS PAGE and western blotting. |
| PCR tubes | Greiner Bio | 608281 | Autoclavable tubes for DsiRNA aliqouts and glycan preparation. |
| Pipette filter tips sterilised (10, 200, 1000 µL) | Gibson | F171203, F171503, F171703 | Sterile filter tips to avoid RNA contamination. |
| PNGase F enzyme | New England Biolabs | P0704S | Enzymatic cleavage of glycans from glycoproteins. |
| Polypropylene columns, 1 mL | Qiagen | 34924 | Columns for gravity-flow chromatography. |
| Protease Inhibitor Cocktail | Sigma Aldrich | P8340 | Inhibition of serine, cysteine, aspartic proteases and aminopeptidases |
| Protein-A Agarose Beads | Merck-Millipore | 16 125 | For purification of human, mouse and rabbit immunoglobulins. |
| Protein-A biosensor | ForteBio | 18 5010 | Tips functionalised with Protein A for rapid antibody quantification. |
| RNaseZap | Invitrogen | AM9780 | Removes RNAse contamination. |
| Sample dilutent | ForteBio | 18 1104 | Activate Protein A tips. |
| Serological pipets (5, 10, 25 mL) | Corning | 4487, 4488, 4489 | Used for sterile cell culture tecniques. |
| Sodium cyanoborohydride solution 1 M in THF | Sigma Aldrich | 296813 | Reducing agent. |
| Solution 18 | ChemoMetec | 910-3018 | Staining reagent containing acridine orange (AO) and 4',6-diamidino-2-phenylindole (DAPI) |
| Spin-X Centrifuge Tube Filters | Corning | 8161 | 0.22 µm pore, Cellulose Acetate membrane. |
| Suspension plate with lid, 6-well | Greiner Bio | 657 185 | Hydrophobic culture plate for growth of suspension cultures. |
| Syringe filters, 0.22 μm | Sartorius | 514 7011 | Surfactant-free cellulose acetate (SFCA) |
| Syringes with Luer lock tip, 20 mL | Fisher Scientific | 10569215 | For secure connection with syringe filter. |
| Trypan Blue solution | Gibco | 15250061 | Stains dead and dying cells. |
| Vivaspin 500, 3,000 MWCO | Sartorius | VS0191 | Polyethersulfone |
| WesternBreeze Chromogenic Kit, anti-rabbit | Thermo Fisher Scientific | WB7105 | Western blot detection kit, alternative blocking buffers and antibody diluents can be made by the user using recipes available online. |
Références
- Jefferis, R. Glycosylation as a strategy to improve antibody-based therapeutics. Nature Reviews Drug Discovery. 8 (3), 226-234 (2009).
- Rathore, A. S., Winkle, H. Quality by design for biopharmaceuticals. Nature Biotechnology. 27 (1), 26-34 (2009).
- Eon-Duval, A., Broly, H., Gleixner, R. Quality attributes of recombinant therapeutic proteins: An assessment of impact on safety and efficacy as part of a quality by design development approach. Biotechnology Progress. 28 (3), 608-622 (2012).
- Lalonde, M. -. E., Durocher, Y. Therapeutic glycoprotein production in mammalian cells. Journal of Biotechnology. 251, 128-140 (2017).
- Mimura, Y., et al. Glycosylation engineering of therapeutic IgG antibodies: challenges for the safety, functionality and efficacy. Protein & Cell. 9 (1), 47-62 (2018).
- Walsh, G. Biopharmaceutical benchmarks 2018. Nature Biotechnology. 36 (12), 1136-1145 (2018).
- Zhang, M., Koskie, K., Ross, J. S., Kayser, K. J., Caple, M. V. Enhancing glycoprotein sialylation by targeted gene silencing in mammalian cells. Biotechnology and Bioengineering. , (2010).
- Kanda, Y., et al. et al. of a GDP-mannose 4,6-dehydratase (GMD) knockout host cell line: a new strategy for generating completely non-fucosylated recombinant therapeutics. Journal of Biotechnology. 130 (3), 300-310 (2007).
- Umaña, P., Jean-Mairet, J., Moudry, R., Amstutz, H., Bailey, J. E. Engineered glycoforms of an antineuroblastoma IgG1 with optimized antibody-dependent cellular cytotoxic activity. Nature Biotechnology. 17 (2), 176-180 (1999).
- Xu, X., et al. et al. genomic sequence of the Chinese hamster ovary (CHO)-K1 cell line. Nature Biotechnology. 29 (8), 735-741 (2011).
- Chan, K. F., et al. et al. of GDP-fucose transporter gene (Slc35c1) in CHO cells by ZFNs, TALENs and CRISPR-Cas9 for production of fucose-free antibodies. Biotechnology Journal. 11 (3), 399-414 (2016).
- Zhang, P., et al. Identification of functional elements of the GDP-fucose transporter SLC35C1 using a novel Chinese hamster ovary mutant. Glycobiology. 22 (7), 897-911 (2012).
- Amann, T., et al. Genetic engineering approaches to improve posttranslational modification of biopharmaceuticals in different production platforms. Biotechnology and Bioengineering. 116 (10), 2778-2796 (2019).
- Karottki, K. J., et al. Awakening dormant glycosyltransferases in CHO cells with CRISPRa. Biotechnology and Bioengineering. 117 (2), 593-598 (2020).
- Fire, A., et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 391 (6669), 806-811 (1998).
- Hammond, S. M., Bernstein, E., Beach, D., Hannon, G. J. An RNA-directed nuclease mediates post-transcriptional gene silencing in Drosophila cells. Nature. 404 (6775), 293-296 (2000).
- Bernstein, E., Caudy, A. A., Hammond, S. M., Hannon, G. J. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature. 409 (6818), 363-366 (2001).
- Shields, R. L., et al. et al. of fucose on human IgG1 N-linked oligosaccharide improves binding to human FcγRIII and antibody-dependent cellular toxicity. Journal of Biological Chemistry. 277 (30), 26733-26740 (2002).
- Shinkawa, T., et al. et al. Absence of fucose but not the presence of galactose or bisecting N-acetylglucosamine of human IgG1 complex-type oligosaccharides shows the critical role of enhancing antibody-dependent cellular cytotoxicity. Journal of Biological Chemistry. 278 (5), 3466-3473 (2003).
- Mori, K., et al. Engineering Chinese hamster ovary cells to maximize effector function of produced antibodies using FUT8 siRNA. Biotechnology and Bioengineering. 88 (7), 901-908 (2004).
- Imai-Nishiya, H., et al. Double knockdown of alpha1,6-fucosyltransferase (FUT8) and GDP-mannose 4,6-dehydratase (GMD) in antibody-producing cells: a new strategy for generating fully non-fucosylated therapeutic antibodies with enhanced ADCC. BMC Biotechnology. 7, 84 (2007).
- Beuger, V., et al. et al. silencing of fucosyltransferase 8 in Chinese-hamster ovary cells for the production of antibodies with enhanced antibody immune effector function. Biotechnology and Applied Biochemistry. 53 (1), 31 (2009).
- Templeton, N., Dean, J., Reddy, P., Young, J. D. Peak antibody production is associated with increased oxidative metabolism in an industrially relevant fed-batch CHO cell culture. Biotechnology and Bioengineering. 110 (7), 2013-2024 (2013).
- Hussain, H., et al. A comparative analysis of recombinant Fab and full-length antibody production in Chinese hamster ovary cells. Biotechnology and Bioengineering. 118 (12), 4815-4828 (2021).
- Shimoyama, H., et al. Partial silencing of fucosyltransferase 8 gene expression inhibits proliferation of Ishikawa cells, a cell line of endometrial cancer. Biochemistry and Biophysics Reports. 22, 100740 (2020).
- Tummala, S., et al. Evaluation of exogenous siRNA addition as a metabolic engineering tool for modifying biopharmaceuticals. Biotechnology Progress. 29 (2), 415-424 (2013).
- Carillo, S., et al. Comparing different domains of analysis for the characterisation of N-glycans on monoclonal antibodies. Journal of Pharmaceutical Analysis. 10 (1), 23-34 (2020).
- Reusch, D., et al. Comparison of methods for the analysis of therapeutic immunoglobulin G Fc-glycosylation profiles-Part 2: Mass spectrometric methods. mAbs. 7 (4), 732-742 (2015).
- Donini, R., Haslam, S. M., Kontoravdi, C. Glycoengineering Chinese hamster ovary cells: a short history. Biochemical Society Transactions. 49 (2), 915-931 (2021).
- Szigeti, M., Chapman, J., Borza, B., Guttman, A. Quantitative assessment of mAb Fc glycosylation of CQA importance by capillary electrophoresis. ELECTROPHORESIS. 39 (18), 2340-2343 (2018).
- Ruhaak, L. R., et al. Optimized workflow for preparation of APTS-labeled N-glycans allowing high-throughput analysis of human plasma glycomes using 48-channel multiplexed CGE-LIF. Journal of Proteome Research. 9 (12), 6655-6664 (2010).
- Patenaude, A. -. M., et al. N-glycosylation analysis of mouse immunoglobulin G isolated from dried blood spots. Electrophoresis. 42 (24), 2615-2618 (2021).
- Karst, D. J., et al. Modulation and modeling of monoclonal antibody N-linked glycosylation in mammalian cell perfusion reactors. Biotechnology and Bioengineering. 114 (9), 1978-1990 (2017).
- Dadouch, M., Ladner, Y., Perrin, C. Analysis of monoclonal antibodies by capillary electrophoresis: sample preparation, separation, and detection. Separations. 8 (1), 4 (2021).
- Fakhr, E., Zare, F., Teimoori-Toolabi, L. Precise and efficient siRNA design: a key point in competent gene silencing. Cancer Gene Therapy. 23 (4), 73-82 (2016).
- Birmingham, A., et al. A protocol for designing siRNAs with high functionality and specificity. Nature Protocols. 2 (9), 2068-2078 (2007).
- Makrydaki, E., et al. . Immobilised enzyme cascade for targeted glycosylation. , (2022).
- . Bulletin_6201.pdf Available from: https://www.bio-rad.com/webroot/web/pdf/lsr/literature/Bulletin_6201.pdf (2022)
- . Bulletin_6376.pdf Available from: https://www.bio-rad.com/webroot/web/pdf/lsr/literature/Bulletin_6201.pdf (2022)
- . Dot Blot Analysis Available from: https://imagej.nih.gov/ij/docs/examples/dot-blot/index.html (2022)
- Yilmazel, B., et al. Online GESS: prediction of miRNA-like off-target effects in large-scale RNAi screen data by seed region analysis. BMC Bioinformatics. 15 (1), 192 (2014).
- Kittler, R., et al. Genome-wide resources of endoribonuclease-prepared short interfering RNAs for specific loss-of-function studies. Nature Methods. 4 (4), 337-344 (2007).
- Stec, E., et al. A multiplexed siRNA screening strategy to identify genes in the PARP pathway. Journal of Biomolecular Screening. 17 (10), 1316-1328 (2012).
- Brown, J. M., et al. Ligand conjugated multimeric siRNAs enable enhanced uptake and multiplexed gene silencing. Nucleic Acid Therapeutics. 29 (5), 231-244 (2019).
- Parsons, B. D., Schindler, A., Evans, D. H., Foley, E. A direct phenotypic comparison of siRNA pools and multiple individual duplexes in a functional assay. PLoS One. 4 (12), 8471 (2009).
- Martin, S. E., et al. Multiplexing siRNAs to compress RNAi-based screen size in human cells. Nucleic Acids Research. 35 (8), 57 (2007).
- Popp, O., Moser, S., Zielonka, J., Rüger, P., Hansen, S., Plöttner, O. Development of a pre-glycoengineered CHO-K1 host cell line for the expression of antibodies with enhanced Fc mediated effector function. mAbs. 10 (2), 290-303 (2018).
- Yang, P. -. C., Mahmood, T. Western blot: Technique, theory, and trouble shooting. North American Journal of Medical Sciences. 4 (9), 429 (2012).
- Troubleshooting Western Blots with the Western Blot Doctor. Bio-Rad Laboratories Available from: https://www.bio-rad.com/en-uk/applications-technologies/troubleshooting-western-blots-with-western-blot-doctor?ID=MIW4HR15 (2022)
- Steentoft, C., et al. Precision mapping of the human O-GalNAc glycoproteome through SimpleCell technology. EMBO. 32 (10), 1478-1488 (2013).
- . FUT8 - Alpha-(1,6)-fucosyltransferase - Proteomics Available from: https://www.nextprot.org/entry/NX_Q9BYC5/proteomics (2022)
- Kotidis, P., et al. Rapid antibody glycoengineering in CHO cells via RNA interference and CGE-LIF N-glycomics. Methods in Molecular Biology. 2370, 147-167 (2022).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationExplorer plus d’articles
This article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.