Method Article
Demonstration der proteolytischen Aktivierung des epithelialen Natriumkanals (ENaC) durch Kombination von Strommessungen mit Nachweis der Spaltfragmente
In diesem Artikel
Zusammenfassung
Proteolytische Aktivierung des epithelialen Natriumkanals (ENaC) heterolog in Xenopus laevis Oozyten kann durch Kombinieren Strommessungen mit einer Biotinylierung Ansatz, um das Aussehen von Ionenkanal-Spaltprodukte an der Zelloberfläche zu untersuchen nachgewiesen werden. Funktionell wichtigen Schnittstellen können durch ortsgerichtete Mutagenese identifiziert werden.
Zusammenfassung
The described methods can be used to investigate the effect of proteases on ion channels, receptors, and other plasma membrane proteins heterologously expressed in Xenopus laevis oocytes. In combination with site-directed mutagenesis, this approach provides a powerful tool to identify functionally relevant cleavage sites. Proteolytic activation is a characteristic feature of the amiloride-sensitive epithelial sodium channel (ENaC). The final activating step involves cleavage of the channel’s γ-subunit in a critical region potentially targeted by several proteases including chymotrypsin and plasmin. To determine the stimulatory effect of these serine proteases on ENaC, the amiloride-sensitive whole-cell current (ΔIami) was measured twice in the same oocyte before and after exposure to the protease using the two-electrode voltage-clamp technique. In parallel to the electrophysiological experiments, a biotinylation approach was used to monitor the appearance of γENaC cleavage fragments at the cell surface. Using the methods described, it was demonstrated that the time course of proteolytic activation of ENaC-mediated whole-cell currents correlates with the appearance of a γENaC cleavage product at the cell surface. These results suggest a causal link between channel cleavage and channel activation. Moreover, they confirm the concept that a cleavage event in γENaC is required as a final step in proteolytic channel activation. The methods described here may well be applicable to address similar questions for other types of ion channels or membrane proteins.
Einleitung
Proteasen sind Enzyme, die in verschiedenen physiologischen Reaktionen, die von dem bekannten proteolytischen Abbau von Proteinen beteiligt sind, im Zusammenhang mit der Verdauung, um hoch entwickelte Protease-Kaskaden in komplexen regulatorischen Signalwege. Proteasen werden in sieben Gruppen entsprechend ihrer katalytischen aktiven Stelle klassifiziert: Aspartat, Asparagin, Cystein, Glutaminsäure, Metallo-, Serin-und Threonin-Proteasen. Verschiedene Proteasen gezielt verschiedene Schnittstellen, die nicht einfach von der Primärstruktur eines Proteins vorherzusagen sind immer. Die MEROPS Datenbank ( http://merops.sanger.ac.uk/ ) liefert detaillierte Informationen über eine Vielzahl von Proteasen und ihrer bevorzugten Schnittstellen. Funktionell relevante Spaltstellen können durch ortsgerichtete Mutagenese identifiziert werden.
Es ist gut bekannt, daß die proteolytische Prozessierung des ENaC ist ein wichtiger Mechanismus der Aktivierung von thist insbesondere Ionenkanal 1,2. Interessanterweise gibt es Hinweise, dass die Funktion der zugehörigen säureempfindlichen Ionenkanal 1a (ASIC1a) können auch durch Proteasen 3-5 geändert werden. Derzeit bleibt es eine offene Frage, ob proteolytischen Spaltung Kanal spielt eine relevante physiologische Rolle bei der Regulierung der Aktivität anderer Ionenkanäle oder Transporter. 6. Es ist jedoch auch festgestellt, dass die proteolytische Spaltung aktiviert eine Gruppe von G-Protein-gekoppelten Rezeptoren, die Protease-aktivierten Rezeptoren (PARs). Mehrere Serinproteasen (zB Kanal-aktivierenden Proteasen (CAP1-3), Chymotrypsin, Trypsin, Furin, Plasmin, neutrophile Elastase und Kallikrein) haben gezeigt, dass proteolytisch aktivieren ENaC 2. Zusätzlich zu Serinproteasen, können andere Gruppen von Proteasen proteolytische ENaC Aktivierung beteiligt sein. In der Tat, die jüngsten Daten zeigen, dass die Metalloproteinase Meprin-β 7 und die Cystein-Protease Cathepsin-S 8 können auch activate ENaC. Die (patho-) physiologisch relevanten Proteasen für ENaC Aktivierung bleibt jedoch festgelegt werden und kann von Gewebe zu Gewebe unterschiedlich.
Proteasen sind dafür bekannt, bevorzugt an bestimmten Stellen in der Aminosäuresequenz spalten. So zeigt zum Beispiel die Serinprotease Chymotrypsin einer spezifischen Spaltungsmuster Spaltung nach der aromatischen Aminosäurereste Phenylalanin und Tyrosin. Im Gegensatz dazu ist die Serinprotease Trypsin spaltet bevorzugt nach der basischen Reste Lysin oder Arginin. Verwendung von mutierten menschlichen γENaC Konstrukte durch ortsgerichtete Mutagenese erzeugt, könnte funktionell relevante Spaltstellen in ENaC heterolog in der Eizelle Expressionssystem exprimiert 8-13 identifiziert werden.
Durch Einspritzen von cRNA für die drei ENaC-Untereinheiten (αβγ) in Oozyten isoliert, kann ENaC funktionell in diesen Zellen exprimiert werden und die Aktivität der an der Plasmamembran vorhanden Kanäle gemessen werdenVerwendung der Zwei-Elektroden-Voltage-Clamp-Technik. Durch die Verwendung der Diuretikum Amilorid, eine spezifische ENaC-Inhibitor, den Amilorid-sensitiven ENaC-vermittelten Ganzzellstromkomponente (&Dgr; I ami) von unspezifischen Leckströmen oder von Strömen von anderen Ionenkanälen geführt getrennt werden. Somit &Dgr; I ami Werte beziehen Gesamt ENaC-Aktivität und durch Subtrahieren in Gegenwart von Amilorid von den in Abwesenheit von Amilorid aufgezeichnet entsprechenden Ganzzellströme gemessen Ganzzellströme bestimmt werden. Um zu testen, ob eine Protease hat eine stimulierende Wirkung auf ENaC &Dgr; I ami zweimal in derselben Oocyte dh vor und nach der Inkubation der Eizelle in einer Protease enthaltenden Lösung gemessen. Ein Anstieg von &Dgr; I ami von der ersten zur zweiten Messung eine proteolytische Aktivierung ENaC. Chymotrypsin oder Trypsin ist bekannt, maximal stimulieren ENaC in der Eizelle Expressionssystem 2,14 und kann verwendet werden, um Konfirm dass ENaC proteolytische Aktivierung in einer gegebenen Charge von Oozyten nachweisbar.
Parallel Ganzzellstrommessungen wurde eine Biotinylierung Ansatz 9 verwendet, um zu untersuchen, ob die Zunahme &Dgr; AMI detektiert bei der Belichtung von Oozyten Proteasen korreliert mit dem Aussehen des ENaC-Spaltungsfragmente an der Zelloberfläche. Proteine auf der Zelloberfläche mit Biotin markiert sind und von intrazellulären Proteinen durch Bindung der biotinylierten Proteine Neutravidin-Agarose-markierten Perlen abgetrennt werden. Die biotinylierten Proteinen durch Western-Blot analysiert werden. γENaC Spaltungsfragmente an der Zelloberfläche kann unter Verwendung eines spezifisch gegen ein Epitop in der C-Terminus des γENaC gerichteten Antikörper nachgewiesen werden. Funktionell relevant Spaltstelle (n) zu identifizieren, vorhergesagten Spaltstellen können durch ortsgerichtete Mutagenese mutiert werden. Wildtyp-und mutierten Kanäle in parallelen Experimenten unter Verwendung von Eizellen aus den s gegename Charge.
Mit diesem methodischen Ansatz wurde es zum ersten Mal, dass die proteolytische Aktivierung von ENaC-vermittelten Ganzzellströme korreliert mit der zeitabhängigen Erscheinung ENaC-Spaltungsfragmente an der Zelloberfläche nachgewiesen. Diese Ergebnisse legen nahe, einen ursächlichen Zusammenhang zwischen Kanal Spaltung und Kanalaktivierung. Darüber hinaus unter Verwendung von ortsgerichteter Mutagenese von mutmaßlichen Spaltungsstellen in Verbindung mit der Zwei-Elektroden-Voltage-Clamp-Technik, funktionell relevante Spaltstellen für Plasmin, Chymotrypsin und Cathepsin 13-S 8 identifiziert.
Protokoll
1. Isolierung von Xenopus Oozyten und Mikroinjektion von cRNA
- Erhalten Eizellen aus erwachsenen weiblichen Xenopus laevis. Betäuben Tiere in 0,2% MS222 und Eierstock-Resektion Lappen durch einen kleinen Bauchschnitt.
- Isolieren Oozyten aus Eierstocklappen durch enzymatischen Verdau bei 19 ° C für 3-4 h bei 600-700 U / ml Collagenase Typ 2 aus Clostridium histolyticum in calciumfreiem OR2-Lösung (Rezeptur in Tabelle 1) gelöst.
- Für die Auswahl, legen Sie die defollikuliert Eizellen in einer Petrischale unter einem binokularen Mikroskop in einem hohen Natrium-haltigen Lösung (ND96: Rezept in Tabelle 1).
- Wählen Stadium V-VI Oozyten und legen Sie sie in einem anderen Petrischale mit einer Pasteur-Pipette. HINWEIS: Blunt Pasteur-Pipette durch Abflammen zu Eizelle Verletzungen zu vermeiden.
- Inject Oozyten mit cRNA (zB 0,2 ng pro αβγENaC-Untereinheit). Lösen cRNAs in RNase-freiem Wasser. HINWEIS: GesamtVolumen in jede Eizelle injiziert wird, ist 46 nl.
- Shop injizierten Oozyten bei 19 ° C in einem niedrigen Natrium-Lösung für Natrium Belastung der Eizellen zu verhindern (ND9: Rezept in Tabelle 1). Ergänzung der Lösung mit 100 U / ml Penicillin Natrium und 100 ug / ml Streptomycinsulfat, um Bakterienwachstum zu verhindern. Sorgfältig hand die Eizellen, die Menge der beschädigten Eizellen oder tot zu begrenzen und sie in einzelnen kleinen Gruppen in einer 12-Well-Platte Brunnen mit Bad-Lösung während der zwei Tage nach cRNA Injektion gefüllt.
2. Darstellende Zwei-Elektroden-Voltage-Clamp-Experimente
- Messung der Oozyten zwei Tage nach der Injektion.
- Füllen Sie eine Spritze eines Schwerkraft-Perfusions-System mit ND96-Lösung und einer weiteren Spritze mit ND96 Lösung Amilorid (2 uM) mit. Berg Spritzen 50 cm über der Eizelle Bad Kammer. HINWEIS: Die Konzentration des ENaC-Inhibitor Amilorid wurde gewählt, 20-mal höher als die IC 50 (100 nM).
- Schalten Sie eine 150 W Halogen-Kaltlichtquelle und stellen Sie es auf 10 cm über der Eizelle Bad Kammer, die eine gute Visualisierung mit dem binokularen Mikroskop. Dann auf Saug-und justieren das Saugrohr am Ende der Eizelle Bad Kammer. Suchen Saugrohr gegenüber Röhren Superfusion "-Adapter in die Eizelle Badewanne. HINWEIS: Saugleistung muss ausreichen, um kontinuierlichen Fluss der Lösung Superfusion die Eizelle zu unterstützen.
- Superfusion einzustellen Geschwindigkeit jeder Lösung auf 3-5 ml / min unter Verwendung der Schwerkraft iv Strömungssteuereinrichtung. Schließen Sie die Superfusion Rohre mit einem Adapter an der Eizelle Bad Kammer.
- Ziehen Glaskapillaren mit einer Mikropipette Abzieher Spitzendurchmesser von <1 um zu erhalten. Dann füllen Kapillaren ~ 1/4 mit 3 M KCl. HINWEIS: Stellen Sie sicher, dass der Teil des chlorierten Silberdraht des Elektrodenhalter in KCl-Lösung eingetaucht. Auf Luftblasen in der Spitze der Kapillare. Luftblasen beeinträchtigen die Messrement durch zunehmende Resistenz Streukapazität.
- Legen Sie die Kapillaren in den Elektrodenhalter des Stroms und der Spannungselektrode und legen Sie sie in ND96 enthält Amilorid (2 uM)-Lösung unter Verwendung der Mikromanipulatoren.
3. Messung von Amilorid-sensitive Ganzzellströme
- Null das Elektrodenpotential der Spannungselektrode (V m) und die Stromelektrode (V e) durch Einstellen der V m und V e versetzt Tasten. HINWEIS: Der Widerstand sollte 1-2 M &OHgr; für die Elektrode zu V m und 0,5 zu messen -1 MOhm für die aktuelle Injektionselektrode.
- Zeigen die Eizelle in das Bad Kammer in der Nähe der Spannungserfassungselektrode. HINWEIS: Die Eizelle während einer dieser Übertragungsschritte nicht beschädigen. Verwenden Sie eine Pasteur-Pipette, um die Eizelle zu übertragen. Um eine Beschädigung der Eizelle die Kanten der Pipette zu vermeiden, sollte durch brenn abgestumpft werden.
- Impale oozyten vorsichtig mit beiden Mikroelektroden.
- Stellen Sie die Haltepotential an den Verstärker bis -60 mV und schalten Sie den Schreiber. Schalten Sie den Amilorid (2 uM) haltigen Lösung. HINWEIS: Der Strom sollte etwa 0 ± 0,5 uA sein. Größere Leckströme zeigen eine undichte impalement. Daher sollten diese Oozyten zurückzuweisen. Außerdem sollten die in der Gegenwart von Amilorid (2 &mgr; M) gemessenen Leckströme ähnlich αβγ-WT exprimierenden Oozyten denen in αβγ ENaC-Mutante exprimierenden Oozyten gemessen werden. Dies zeigt, dass die Mutationen nicht beeinträchtigen den Amilorid-Empfindlichkeit des Kanals.
- Starten Sie die Aufnahme. Bei Bedarf stellen Sie die Verstärkung.
- Nach der gemessene Strom ein stabiles Plateau erreicht, auf Amilorid-freie Lösung zu ändern. HINWEIS: Abwärts aktuellen Ablenkungen in der aktuellen Spuren entsprechen Einwärtsströme, dh Bewegung der positiven Ladung (Na +) aus der extrazellulären Seite in die Zelle.
- After ein Strom Plateau erreicht ist (nach ~ 60 sec), schalten Sie die Superfusion zurück zum Amilorid haltigen Lösung. Nachdem der Strom der Eizelle die anfängliche Basisstrom erreicht, schalten Sie die Spannungsklemme und die Elektroden vorsichtig zurückziehen.
- Zum Wiederverschließen der Plasmamembran an den Standorten der impalement ermöglichen, legen die Eizelle in eine Vertiefung einer 96-Well-Platte mit 100-150 ul Protease kostenlos ND96 Lösung.
- Nach 5 min übertragen Eizelle einer Protease enthaltenden Lösung oder zu einer Kontrolllösung ohne Protease für 30 min Inkubationszeit. HINWEIS: Die Inkubationszeit ist abhängig von der Protease und der untersuchten Kanals.
- Nach der Inkubation Schritt wiederholen Sie den Strommessung (siehe 3.2 und folgende). HINWEIS: Es ist möglich, nach Inkubation in Protease-Lösung messen> 90% der Eizellen.
4. Biotinylierung Assay
- Wählen und entsorgen Sie defekte Eizellen unter dem Binokular. HINWEIS: Verwenden Sie spritzened Oozyten aus der gleichen Charge für die Strommessung und für die Biotinylierung Experimenten.
- Halten Biotin bei RT für mindestens 20 Minuten vor dessen Verwendung in dem Experiment.
- Bereiten Sie die Lösungen: ND96 und ND96, die die geeignete Protease. Bereiten Pasteur Pipetten durch Markierung und ihnen durch kurzes flamm ihre Tipps, um Verletzungen der Eizellen zu vermeiden. HINWEIS: Hier wird die Protease Chymotrypsin 2 ug / ml in ND96 verwendet. Gönnen Sie jede Gruppe mit einer Pipette, um Kreuzkontamination von Lösungen zu vermeiden.
- Füllen Sie jede Vertiefung einer 6-Well-Platte mit 2,5 ml ND96 oder ND96 Kontrolle, die eine Protease bei RT. Dann hinter 30 Eizellen pro Vertiefung und Inkubation für 30 min bei RT. HINWEIS: Für die anschließenden Verfahren ist es wichtig, Proben auf Eis halten zu allen Zeiten. Alle Zentrifugationsschritte werden bei bei 4 ° C durchgeführt wird
- Füllen Sie jede Vertiefung einer neuen 6-Well-Platte mit 2,5 ml ND96 (jede Gruppe braucht 3 Brunnen für den Waschschritten) und wiegen die Biotin. HINWEIS: 2,5 mg Biotin per gut (1 mg / ml) erforderlich ist. Löse das Biotin in der Biotinylierung Puffer (dh 25 mg Biotin (10 Gruppen) in 25 ml Biotinylierung Puffer (Rezept in Tabelle 1).
- Übertragen Sie jede Gruppe von Eizellen zu einem gut mit 2,5 ml ND96 gefüllt. Über Oozyten nacheinander in zwei weitere Bohrlöcher mit ND96 zu waschen Sie die restlichen Protease. Die Eizellen Inkubation für 5 min in ND96.
- Übertragen Sie die Oozyten in eine gut, die 2,5 ml Biotin-Lösung und Inkubation sie mit sanfter Bewegung ("Shaker") für 15 min. HINWEIS: Minimieren Sie das übertragen ND96 mit der Pipette, um eine Verdünnung des Biotin-Lösung zu vermeiden.
- Übertragen Sie jede Gruppe von Eizellen in einen Brunnen, der 2,5 ml Quench-Puffer (Rezept in Tabelle 1), um die Biotinylierungsreaktion stoppen. Dann übertragen Sie jede Gruppe von Eizellen in eine zweite und auch die 2,5 ml stillen Puffer und Inkubation für 5 min unter leichtem Schütteln.
- Beschädigte oder tot Eizellen. Hinweis:Wählen Sie die gleiche Anzahl von Oozyten pro Gruppe für die folgenden Verfahren.
- Übertragen Sie jede Gruppe von Oozyten in ein 1,5 ml Mikrozentrifugenröhrchen aus Kunststoff. HINWEIS: Minimieren Sie die Anzahl der Quench-Puffer, der übertragen wird.
- Anschließend lysieren die Oozyten, indem sie durch eine 27 G-Nadel in 1 ml Lyse-Puffer (Rezept siehe Tabelle 1) mit Proteaseinhibitoren ergänzt.
- Zentrifugieren der Lysate 10 min bei 1.500 x g.
- Überstand wird abgesaugt und übertragen sie in ein 1,5 ml Mikrozentrifugenröhrchen mit 0,5% Triton-X-100 und 0,5% NP40. Entsorgen Sie die verbleibende Pellet. HINWEIS: Der Überstand enthält biotinylierten Plasmamembranproteinen und nicht-biotinylierte intrazellulären Proteinen.
- Die Reaktionsgefäße für 20 Minuten auf Eis inkubiert. Immer wieder Wirbel die Rohre in diesem Zeitraum vollständig die Proteine in NP40 und Triton-X-100 zu lösen.
- Zentrifuge 100 ul Agarose-Perlen pro Eizelle Gruppe für 3 min bei 1.500 x g. Nachdem derZentrifugation entfernen Stand aus der Perlen-Lösung und dreimal mit Lysepuffer, um die Perlen mit Puffer ins Gleichgewicht.
- Je 100 ul der gewaschenen Kügelchen in jedes Mikrozentrifugenröhrchen, das die Protein-Detergens-Lösung in 4,13 hergestellt Bindung der biotinylierten Proteine an die Kügelchen zu ermöglichen.
- Mikrozentrifugenröhrchen mit Overhead-Dreh O / N bei 4 ° C inkubieren
- Zentrifugieren Sie die Reaktionsgefäße für 3 min bei 1.500 x g. Dann den Überstand in ein neues Röhrchen. HINWEIS: Intrazelluläre Proteine nicht mit Biotin markiert. Überstand bei -20 ° C gelagert werden Saugen Sie nicht die Perlen.
5. Nachweis von ENaC Spaltfragmente an der Zelloberfläche durch Western-Blot-Analyse
- Drei Mal Waschen der Kügelchen mit Lyse-Puffer und 100 ul 2x SDS-PAGE-Probenpuffer. HINWEIS: Die Proben können bei -20 ° C gelagert oder sofort für die Western-Blot-Analyse vorbereitet werden.
- Boil sBeispiele für 5 min bei 95 ° C und dann Platz Röhrchen auf Eis.
- Zentrifuge Proben für 3 min bei 20000 xg und Pipetten den Überstand in ein neues Mikrozentrifugenröhrchen. HINWEIS: Dieser Überstand enthält die biotinylierten Plasmamembran-Proteine von der Zelloberfläche der Eizelle.
- Analysiert 30 &mgr; l dieses Überstands durch Western-Blot auf Spaltungsfragmente an der Zelloberfläche zu untersuchen.
- Trenne die biotinylierten Proteine durch SDS-PAGE (Natriumdodecylsulfat-Polyacrylamid-Gel-Elektrophorese) mit einem geeigneten Gel (8%, 10%, 12%, je nach Molekulargewicht der Spaltfragmente untersucht).
- Transfer der Proteine auf Polyvinylidendifluorid (PVDF)-Membranen mittels semi-dry Blotting.
- Sonde der Membran mit einem spezifischen Antikörper gegen menschliches γENaC gegen ein Epitop in der C-Terminus gerichtet (siehe Fig. 3 und 13).
- Verwenden Meerrettichperoxidase-markierten Ziegen-Anti-Kaninchen-Antikörper als SekundärantiKörper.
- Erkennen Chemilumineszenzsignale.
Ergebnisse
Um zu untersuchen, ob die Serinprotease Plasmin ENaC-vermittelten Ströme aktiviert wurde &Dgr; I ami einzelner ENaC-exprimierenden Oozyten vor und nach 30 min Inkubation der Eizellen in proteasefreien (Kontrolle) (Fig. 2A) oder Plasmin enthaltende Lösung (Abbildung bestimmt 2B) unter Verwendung der Zwei-Elektroden-Voltage-Clamp-Technik (siehe Fig. 1). Die Exposition gegenüber Plasmin erhöht &Dgr; ami in jeder Eizelle gemessen. Im Gegensatz dazu ist in Kontrollexperimenten, 30 min Inkubation von ENaC-exprimierenden Oozyten in Protease-freie Lösung hatte einen vernachlässigbaren Effekt (Abbildung 2 C, D). Somit ist es durch Verwendung dieses Verfahrens eine Stimulation des ENaC-vermittelten Strom durch Plasmin detektiert werden.
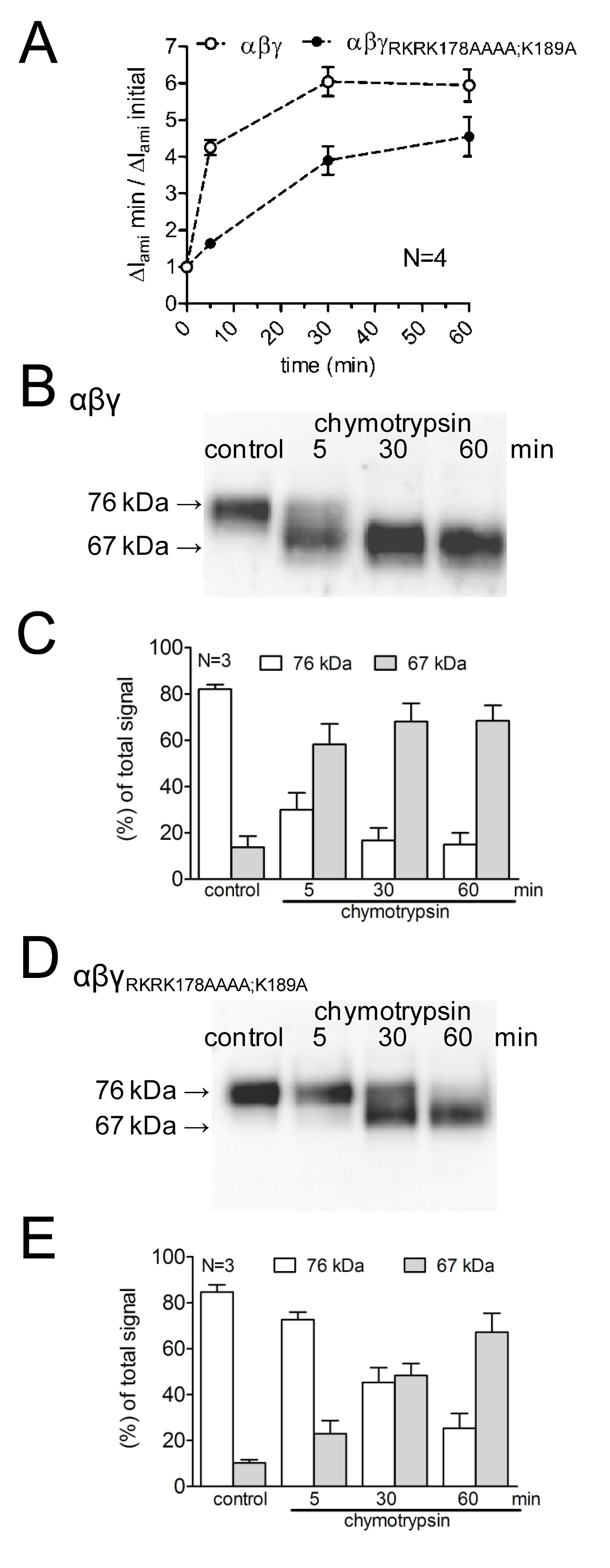
Um die Auswirkungen der Mutation von mutmaßlichen Spaltungsstellen bei der Aktivierung von ENaC-vermittelten Ströme, sowie auf Kanal Spaltung zu studieren, wurde die Wirkung von Chymotrypsin auf WT-ENaC mit diesem Vergleich aufeine Mutante ENaC mit mutiertem Prostasin und Plasmin Spaltstellen (γ RKRK178AAAA; K189A). Der Zeitverlauf der Kanalaktivierung durch Chymotrypsin sowie das Auftreten von ENaC Spaltprodukte an der Zelloberfläche wurde unter Verwendung von verschiedenen Protease Inkubationszeiten (4A) untersucht. Es wurde gezeigt, dass die Mutante Kanalverzögerungen und reduziert die Aktivierung des ENaC-vermittelten Stroms durch Chymotrypsin. Dies wird durch ein verzögertes Auftreten von niedrigerem Molekulargewicht γENaC Spaltfragment von 67 kDa entspricht, der vollständig gespaltene Untereinheit parallel geschaltet. Spaltfragmente wurden unter Verwendung eines γENaC Antikörper gegen ein Epitop in der C-Terminus (Fig. 3) gerichtet sind, detektiert. Dieses methodische Vorgehen zeigt, daß der zeitliche Verlauf der proteolytischen Aktivierung des ENaC-vermittelten Strömen korreliert mit dem Auftreten einer 67-kDa-Spaltprodukt γENaC an der Zelloberfläche (Fig. 4 B, C). Dies unterstützt das Konzept einerKausalzusammenhang zwischen proteolytischen Spaltung Kanal und Kanalaktivierung 13. Darüber hinaus kann durch die Kombination von Strommessungen und die Detektion γENaC Fragmente an der Zelloberfläche wurde gezeigt, dass die mutierten Spaltstellen sind für die proteolytische Kanalaktivierungs funktionell relevant.

1. Verfahren zur Bestimmung des Stimulationswirkung einer Protease auf ENaC heterolog in Xenopus laevis Oozyten. ENaC-Aktivität wird durch Messen der Amilorid-sensitiven Ganzzellstromkomponente &Dgr; I ami geschätzt.

Abbildung 2. Plasmin stimuliert ENaC-vermittelten Währungents in Oozyten, die ENaC. (AD) ENaC menschlichen Oozyten wurden für 30 min in Protease-freie Lösung (Kontrolle) oder in Lösung Plasmin (10 ug / ml) inkubiert. (-), Um &Dgr; I ami vor bestimmen und nach (+) der Inkubation wurden die Oozyten bei einem Haltepotential von -60 mV geklemmt (A, B) vier repräsentativen Ganzzellstrom Spuren von einer Charge von Oozyten.. Amilorid (AMI) war in der Badlösung vorliegenden spezifisch hemmen ENaC durch schwarze Balken dargestellt. (C) Datenpunkte von einem einzelnen Oozyten erhalten werden, die durch eine Linie verbunden. (D) Zusammenfassung der ähnliche Experimente wie in Spalte C gezeigt, darstellen relativen stimulierende Wirkung auf &Dgr; ami berechnet als das Verhältnis von &Dgr; ami nach 30 min Inkubation (&Dgr; I ami 30 min) auf die Anfangs &Dgr; ami (&Dgr; I ami initial), gemessen vor der Inkubation gemessen. Zahlen in den Spalten geben dieAnzahl von einzelnen Oozyten gemessen. N gibt die Anzahl der verschiedenen Chargen von Oocyten. (Diese Zahl hat sich aus geändert worden [Haerteis et al 2012 J Gen Physiol 140, 375-389, doi:. 10.1085/jgp.201110763])

3. Modell der γENaC Einheit zeigt Spaltungsstellen für die proteolytische Aktivierung und der Bindungsstelle des Antikörpers verwendet. Proteolytische Spaltung durch das Golgi-assoziierten Konvertase Furin ist wichtig für ENaC Reifung im Biosyntheseweg, bevor der Kanal die Plasmamembran erreicht. Nach Spaltung durch Furin ein 76 kDa-Fragment an der Zelloberfläche unter Verwendung eines Ansatzes und Biotinylierung eines Antikörpers gegen ein Epitop in der C-Terminus der γ-Untereinheit nachgewiesen werden. Die Schwenk letzte Schritt in proteolytischen ENAC-Aktivierung an der Plasmamembran, wo γENaC wird durch extrazelluläre Proteasen (z. B. Plasmin oder Chymotrypsin) in einem Bereich distal des Furinstelle was zu einem 67 kDa-Spaltfragment gespalten erfolgt wahrscheinlich. (Diese Zahl hat sich aus geändert worden [Haerteis et al 2012 J Gen Physiol 140, 375-389, doi:. 10.1085/jgp.201110763])

Fig. 4:. Mutierende sowohl die Plasmin (K189) und die Prostasin-Spaltstelle (RKRK178) verzögert die Aktivierung des ENaC-vermittelten Ströme und das Aussehen einer 67 kDa-Spaltprodukt von γ-Untereinheit des Kanals Oozyten, WT (offene Symbole) und γ RKRK178AAAA; K189A ENaC mutierten Kanal (geschlossene Symbole) wurden für 30 min in Protease-freie Lösung (Kontrolle) oder für 5, 3 inkubiert0 oder 60 Minuten in einem Chymotrypsin-Lösung (2 &mgr; g / ml) enthält. (A) um &Dgr; I ami vor und nach der Inkubation zu bestimmen, wurden Oozyten bei einem Haltepotential von -60 mV geklemmt. Kreise stellen das Verhältnis von &Dgr; ami gemessen nach 5, 30 oder 60 min Inkubation (&Dgr; I ami min) auf die Anfangs &Dgr; ami (&Dgr; I ami initial) vor der Inkubation gemessen. Jeder Datenpunkt repräsentiert den Mittelwert &Dgr; I ami in 22-24 einzelnen Oozyten von vier verschiedenen Chargen gemessen. (BD) Parallel zur Detektion von AMI &Dgr; I die Expression von biotinylierten γENaC an der Zelloberfläche wurde durch SDS-PAGE analysiert. γENaC wurde mit einem Antikörper gegen ein Epitop in der C-Terminus des humanen γENaC detektiert. Repräsentative Western Blots von einer Charge von Oozyten gezeigt werden. (CE) Densitometrische Analyse der Western-Blots drei ähnlich denen in B oder D gezeigt für jede Bahn, die signals in den Regionen von 76 kD (offene Säulen) und 67 kD (graue Säulen) nachgewiesen wurden bestimmt und normiert auf die Summe der festgestellten Gesamtsignal. N gibt die Anzahl der verschiedenen Chargen von Eizellen. Klicken Sie hier für eine größere Ansicht.
{kind=link}
Diskussion
In diesem Manuskript ein methodischer Ansatz, der erfolgreich angewendet wurde, um die Mechanismen, durch Proteasen zugrunde liegenden die Aktivierung von ENaC studieren beschrieben 8,13. Die bewährte Xenopus laevis Oozyten-Expressionssystem wurde verwendet, um funktionell exprimieren ENaC. ENaC-Funktion wurde mit der konventionellen Zwei-Elektroden-Voltage-Clamp-Technik beurteilt. Ortsspezifische Mutagenese wurde verwendet, um funktionell relevante Protease-Spaltungsstellen zu identifizieren. Biotinylierung Experimente parallel zu den elektrophysiologischen Messungen durchgeführt wurde es möglich, das Auftreten korreliert ENaC Spaltprodukte an der Zelloberfläche proteolytisch Stromaktivierung. Ein Zusammenhang zwischen dem zeitlichen Verlauf der Stromaktivierung und dem Auftreten von proteolytischen Spaltungsfragmente an der Zelloberfläche unterstützt das Konzept der proteolytischen Kanalaktivierung.
Zwei-Elektroden-Voltage-Clamp-Aufnahmen erfordern die Pfählung eines oocyte mit zwei Mikroelektroden. Dieses Verfahren wird in der Regel nur einmal in einer einzelnen Eizelle durchgeführt. Jedoch war es möglich, die Mikroelektroden zu entfernen, nachdem eine anfängliche Ganzzellstrom Aufzeichnungs ohne offensichtliche Beschädigung der Eizelle. Tatsächlich scheint die Plasmamembran an den Stellen der Pfählungen innerhalb weniger Minuten wieder zu verschließen. Somit wird nach Abschluss einer ersten Zwei-Elektroden-Voltage-Clamp-Messung ist es möglich, die Eizelle aus dem experimentellen Strömungskammer des Zwei-Elektroden-Voltage-Clamp-Setup, um ein Mikrozentrifugenröhrchen oder ein Well einer 96-Well-Platte gefüllt übertragen ein kleines Volumen der Test-oder Kontrolllösung. Anschließend kann die gleiche Eizelle zurück zu der Strömungskammer überführt werden und kann wieder bohrt, um eine zweite Zwei-Elektroden-Voltage-Clamp-Messung durchzuführen. Bemerkenswerterweise hat ENaC-vermittelten Ströme nicht sehr zwischen der ersten und der zweiten Messung, wenn die Oocyte wurde in Kontrolllösung gehalten variieren. Im Gegensatz dazu Inkubation der Eizelle in einer Protease enthält Solution nach der ersten Messung zu erhöhten ENaC-vermittelten Stroms in der zweiten Messung (Fig. 2). Diese Feststellung zeigt proteolytischen Kanalaktivierung.
Durchführung von zwei separaten Strommessungen in einer einzigen Eizelle bietet den Vorteil, dass die Eizelle kann Proteasen oder anderen pharmakologischen Mitteln zwischen den zwei Messungen für eine variable Zeitdauer in einer kleinen Menge Testlösung ausgesetzt werden. Dies ist wichtig bei der Verwendung von Mitteln, die teuer und / oder in großen Mengen verfügbar sind, z. B. gereinigte Protease-Zubereitungen. Die begrenzte Verfügbarkeit von Mitteln kann es unmöglich machen (oder unbezahlbar), um sie im Dauer Zwei-Elektroden-Voltage-Clamp-Aufnahmen verwenden, da der große Mengen von für die kontinuierliche Superfusion der Oozyten mit Durchflussraten von mehreren Millilitern pro Minute erforderlich Testlösung. Außerdem werden kontinuierlich zwei Elektrodenspannungs-Clamp-Messungen von dem bekannten Phänomen begrenztnomen der spontanen Kanal heruntergekommenen auch für ENaC 15 beschrieben. Im Gegensatz dazu Belichten Oozyten Lösungen zwischen zwei separaten Messungen für bis zu einer Stunde oder mehr Test nicht allgemein ein Problem darstellen (siehe Fig. 4A). Schließlich zwei aufeinanderfolgenden Messungen in der gleichen Eizelle durchgeführt, damit gepaart Beobachtungen der Arzneimittelwirkungen. Dies hat einen Vorteil gegenüber ungepaarten Messungen von zwei verschiedenen Gruppen von Oocyten (Protease-behandelten und Vehikel-behandelten), da es das Problem der hohen Variabilität zwischen den Oozyten, in der Regel in der Ionenkanalexpression beobachtet reduziert. Gepaart mit Beobachtungen und der Möglichkeit, die Daten in der ersten Messung zu normalisieren, werden weniger Oozyten pro Versuchsgruppe benötigt, um eine signifikante Wirkung eines pharmakologischen Wirkstoffs zeigen. Die Normalisierung der Daten macht es auch einfach, Daten aus verschiedenen Chargen von Oozyten mit unterschiedlichen Ionenkanal-Expressionsniveaus und damit unterschiedliche Ausgangsströme (2D zusammenfassen). Offensichtlich sind Kontrollversuche notwendig, dieses Konzept zu demonstrieren, dass der Ionenkanal-Aktivität von Interesse bleibt in Vehikel-behandelten Kontroll Oozyten stabil von der ersten zur zweiten Messung (siehe 2).
Um zu zeigen, daß proteolytische Stromaktivierung korreliert mit dem Auftreten von ENaC Spaltprodukte an der Zelloberfläche, eine Biotinylierung Ansatz ursprünglich von Harris et al. 9 verwendet werden kann. Dieses Verfahren (wie im Protokoll Abschnitt beschrieben und in Fig. 4 gezeigt) wurde angepaßt, daß die Belastung der Kanäle, Proteasen und nachfolgende Aktivierung ENaC-vermittelten Ströme wird durch die zeitabhängige Auftreten von Spaltfragmente parallel zu demonstrieren. Die Biotinylierung Verfahren erlaubt auch die Analyse von einer allgemeinen Zunahme oder Abnahme der Membranproteine auf der Zelloberfläche. Somit ist dieses Verfahren geeignet, die Wirkung von Proteasen und anderen phar untersuchenmacological Agenten auf Kanal Einsetzen in die Plasmamembran oder beim Kanal Abruf. Außerdem Western-Blot-Analyse der biotinylierten Plasmamembranproteinen erlaubt die Detektion von Protein-Fragmenten (z. B. proteolytische Fragmente ENaC) oder Änderungen im Glykosylierungsmuster, die funktionell relevant sein können.
Im Ergebnis ist die Kombination der Verfahren verwendet werden, um die stimulierende Wirkung von Proteasen auf ENaC-vermittelten Ganzzellströme zu untersuchen und zeigen eine Korrelation mit dem Auftreten des ENaC Spaltprodukte an der Zelloberfläche kann für einen breiten Bereich von Anwendungen. Insbesondere kann diese Methoden geeignet zu ähnlichen Fragen hinsichtlich der Regulierung der andere Ionenkanäle, Transporter oder Transmembran-Rezeptoren (zB Protease-aktivierten Rezeptoren PAR) zu adressieren.
Offenlegungen
The authors have nothing to disclose.
Danksagungen
The expert technical assistance of Céline Grüninger, Christina Lang, Sonja Mayer, and Ralf Rinke is gratefully acknowledged. We thank Dr. Morag K. Mansley for carefully reading the manuscript. This project was supported by a grant of the Deutsche Forschungsgemeinschaft (Grant SFB 423: Kidney Injury: Pathogenesis and Regenerative Mechanisms, to C. Korbmacher), grants of the Interdisziplinäres Zentrum für Klinische Forschung (to S. Haerteis and M. Krappitz), the ELAN program (to S. Haerteis) of the Friedrich-Alexander-Universität Erlangen-Nürnberg, and the University Library of Erlangen-Nürnberg.
Materialien
| Name | Company | Catalog Number | Comments |
| Bath Clamp Headstage for OC-725C-V | Warner Instrument Corporation | - | |
| Cold light source - Schott KL 1500 LCD | Schott | #SCOC150200EU | brightness 4; mechanical aperture: D; color temperature: 3000 K |
| E Series Electrode Holder (Str, Vent, Ag Wire, 1.2 mm) | ADinstruments | #ESW-F10v | |
| left micromanipulator; MM-33L | Warner Instrument Corporation | #64-0055 | |
| LIH 1600 - computer interface | HEKA | - | |
| magnetic valve system (ALA BPS-8) in combination with a TIB14 interface (HEKA) | ALA Scientific Instruments, HEKA | - | |
| OC-725C amplifier for two-electrode voltage-clamp recordings | Warner Instrument Corporation | - | |
| P-97 FLAMING/BROWN Micropipette Puller | Sutter Instruments | - | heat=550; velocity=22; time=200 |
| right micromanipulator; MM-33R | Warner Instrument Corporation | #64-0056 | |
| Series Electrode Holder (45°, Vent, Handle, Ag Wire, 1.2 mm) | ADinstruments | #E45w-f10vh | |
| STAT 2 IV Gravity Flow Controller | Conmed | #P-S2V-60 | |
| vacuum generator ejector SEG - for suction to remove bath solution | Schmalz | - | |
| Material | |||
| INFUJECT 60ml pump syringes for solutions | Braun | #22050 | |
| Injekt-F for lysing the oocytes | Braun | #9166033V | |
| standard wall borosilicate tubing with filament | Sutter Instruments | #BF150-86-10 | outside diameter: 1.50 mm; inside diameter: 0.86 mm; length: 10 cm |
| Reagent | |||
| complete, Mini, EDTA-free protease inhibitor cocktail tablets | Roche Applied Science | #11836170001 | |
| EZ-Link Sulfo-NHS-Biotin | Thermo Scientific | #21217 | |
| Horseradish peroxidase-labeled secondary goat anti-rabbit antibody | Santa Cruz Biotechnology | #sc-2004 | |
| NeutrAvidin Agarose | Thermo Scientific | #29200 | Neutravidin-labeled agarose beads |
| NP40 (Nonidet P-40) | Sigma-Aldrich | #I8896 | |
| Roti-Load 1 (2× SDS-PAGE sample buffer) | Carl Roth | #K929.2 | |
| SuperSignal West Femto Chemiluminescent Substrate for detection of chemiluminescent signals | Thermo Scientific | #34095 | |
| Triton-X-100 | Sigma-Aldrich | #T8787 | |
Referenzen
- Kleyman, T. R., Carattino, M. D., Hughey, R. P. ENaC at the cutting edge: regulation of epithelial sodium channels by proteases. The Journal of Biological Chemistry. 284, 20447-20451 (2009).
- Rossier, B. C., Stutts, M. J. Activation of the epithelial sodium channel (ENaC) by serine proteases. Annu Rev Physiol. 71, 361-379 (2009).
- Poirot, O., Vukicevic, M., Boesch, A., Kellenberger, S. Selective regulation of acid-sensing ion channel 1 by serine proteases. The Journal of Biological Chemistry. 279, 38448-38457 (2004).
- Vukicevic, M., Weder, G., Boillat, A., Boesch, A., Kellenberger, S. Trypsin cleaves acid-sensing ion channel 1a in a domain that is critical for channel gating. The Journal of Biological Chemistry. 281, 714-722 (2006).
- Clark, E. B., Jovov, B., Rooj, A. K., Fuller, C. M., Benos, D. J. Proteolytic cleavage of human acid-sensing ion channel 1 by the serine protease matriptase. The Journal of Biological Chemistry. 285, 27130-27143 (2010).
- Ossovskaya, V. S., Bunnett, N. W. Protease-activated receptors: contribution to physiology and disease. Physiological reviews. 84, 579-621 (2004).
- Garcia-Caballero, A., et al. Activation of the epithelial sodium channel by the metalloprotease meprin β-subunit. Channels (Austin. 5, 14-22 (2011).
- Haerteis, S., et al. Proteolytic activation of the epithelial sodium channel (ENaC) by the cysteine protease cathepsin-S. Pflügers Archiv: European Journal of Physiology. 464, 353-365 (2012).
- Harris, M., Firsov, D., Vuagniaux, G., Stutts, M. J., Rossier, B. C. A novel neutrophil elastase inhibitor prevents elastase activation and surface cleavage of the epithelial sodium channel expressed in Xenopus laevis oocytes. The Journal of Biological Chemistry. 282, 58-64 (2007).
- Passero, C. J., Mueller, G. M., Rondon-Berrios, H., Tofovic, S. P., Hughey, R. P., Kleyman, T. R. Plasmin activates epithelial Na+ channels by cleaving the γ-subunit. The Journal of Biological Chemistry. 283, 36586-36591 (2008).
- Svenningsen, P., et al. Plasmin in nephrotic urine activates the epithelial sodium channel. Journal of the American Society of Nephrology : JASN. 20, 299-310 (2009).
- Patel, A. B., Chao, J., Palmer, L. G. Tissue kallikrein activation of the epithelial Na channel. American Journal of Physiology. Renal Physiology. 303, (2012).
- Haerteis, S., Krappitz, M., Diakov, A., Krappitz, A., Rauh, R., Korbmacher, C. Plasmin and chymotrypsin have distinct preferences for channel activating cleavage sites in the γ-subunit of the human epithelial sodium channel. The Journal of General Physiology. 140, 375-389 (2012).
- Chraibi, A., Vallet, V., Firsov, D., Hess, S. K., Horisberger, J. D. Protease modulation of the activity of the epithelial sodium channel expressed in Xenopus oocytes. The Journal of General Physiology. 111, 127-138 (1998).
- Volk, T., Konstas, A. A., Bassalay, P., Ehmke, H., Korbmacher, C. Extracellular Na+ removal attenuates rundown of the epithelial Na+-channel (ENaC) by reducing the rate of channel retrieval. Pflügers Archiv: European Journal of Physiology. 447, 884-894 (2004).
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten