Method Article
Demonstração de Proteolytic ativação do canal de sódio epitelial (ENAC) combinando Medidas atuais com Detecção de clivagem Fragments
Neste Artigo
Resumo
Activação proteolítica do canal de sódio epitelial (ENaC) heterologamente expressos em oócitos de Xenopus laevis pode ser demonstrada através da combinação de medições de corrente com uma abordagem de biotinilação para investigar o aparecimento de produtos de clivagem do canal de iões na superfície da célula. Locais de clivagem funcionalmente importantes podem ser identificados através de mutagénese dirigida ao local.
Resumo
The described methods can be used to investigate the effect of proteases on ion channels, receptors, and other plasma membrane proteins heterologously expressed in Xenopus laevis oocytes. In combination with site-directed mutagenesis, this approach provides a powerful tool to identify functionally relevant cleavage sites. Proteolytic activation is a characteristic feature of the amiloride-sensitive epithelial sodium channel (ENaC). The final activating step involves cleavage of the channel’s γ-subunit in a critical region potentially targeted by several proteases including chymotrypsin and plasmin. To determine the stimulatory effect of these serine proteases on ENaC, the amiloride-sensitive whole-cell current (ΔIami) was measured twice in the same oocyte before and after exposure to the protease using the two-electrode voltage-clamp technique. In parallel to the electrophysiological experiments, a biotinylation approach was used to monitor the appearance of γENaC cleavage fragments at the cell surface. Using the methods described, it was demonstrated that the time course of proteolytic activation of ENaC-mediated whole-cell currents correlates with the appearance of a γENaC cleavage product at the cell surface. These results suggest a causal link between channel cleavage and channel activation. Moreover, they confirm the concept that a cleavage event in γENaC is required as a final step in proteolytic channel activation. The methods described here may well be applicable to address similar questions for other types of ion channels or membrane proteins.
Introdução
As proteases são enzimas que estão envolvidas em várias respostas fisiológicas que vão desde a degradação proteolítica bem conhecida de proteínas, no contexto da digestão, a cascata de protease altamente sofisticados envolvidas em vias de sinalização complexas reguladora. As proteases são classificados em sete grupos de acordo com o seu sítio ativo catalítico: aspartato, asparagina, cisteína, ácido glutâmico, metalo, serina, treonina e proteases. Proteases diferentes locais alvo de clivagem distintas que não são sempre fáceis de prever a partir da estrutura primária de uma proteína. O banco de dados MEROPS ( http://merops.sanger.ac.uk/ ) fornece informações detalhadas sobre uma ampla gama de proteases e seus sítios de clivagem preferenciais. Locais de clivagem funcionalmente relevantes podem ser identificados utilizando a mutagénese dirigida ao local.
Está bem estabelecido que o processamento proteolítico de ENaC é um importante mecanismo de activação do thé especial 1,2 canal iônico. Curiosamente, não há evidências de que a função do canal de iões de 1a-detecção de ácido correspondente (ASIC1a) pode também ser modificada por proteases 3-5. Actualmente, continua a ser uma questão em aberto se canal proteolítica clivagem desempenha um papel fisiológico importante na regulação da atividade de outros canais iônicos ou transportadores. No entanto, está bem estabelecido que a clivagem proteolítica activa um grupo de receptores acoplados a proteínas, os receptores activados por proteases (PARs) 6. Várias proteases de serina (por exemplo, proteases de activação do canal (CAP1-3), quimotripsina, tripsina, furina, plasmina, elastase de neutrófilos, e calicreína) têm mostrado activar proteoliticamente ENaC 2. Além de proteases de serina, proteases de outros grupos podem estar envolvidos na activação proteolítica ENaC. De facto, dados recentes mostram que a metaloproteinase meprin-β 7 e a protease de cisteína Catepsina S-8 também pode activaçãote ENaC. No entanto, os (pato-proteases) fisiologicamente relevantes para a ativação ENaC continuam a ser determinado e pode variar de tecido para tecido.
As proteases são conhecidas por clivar preferencialmente nos locais específicos na sequência de aminoácidos. Por exemplo, a quimotripsina de serina-protease mostra um padrão de clivagem específico de clivagem após o amino-ácido aromático resíduos de fenilalanina e tirosina. Em contraste, a protease tripsina serina preferencialmente cliva após a resíduos de lisina ou arginina de base. Utilizando construções γENaC humana mutantes gerados por mutagénese dirigida ao local, locais de clivagem funcionalmente relevantes em ENaC heterologamente expressos no sistema de expressão do oócito pode ser identificado 8-13.
Por injecção de ARNc para as três subunidades de ENaC (αβγ) em oócitos isolados, ENaC pode ser funcionalmente expressa nestas células e a actividade de canais presentes na membrana do plasma pode ser medida pelautilizando a técnica de dois eletrodos de tensão-clamp. Ao utilizar o diurético amilorida, um inibidor específico de ENaC, o ENaC mediada componente da corrente de célula inteira sensível à amilorida (ΔI ami) pode ser separado a partir de correntes de fuga não específicos ou a partir de correntes conduzidas por outros canais iónicos. Assim, os valores ΔI ami reflectem a actividade global do ENaC e pode ser determinada subtraindo as correntes de células completas medidos na presença de amiloride a partir das correntes de células completas correspondentes gravadas na ausência de amilorida. Para testar se uma protease tem um efeito estimulador sobre ENaC, ΔI ami é medida duas vezes no mesmo oócito, isto é, antes e depois da incubação do oócito em uma solução contendo protease. Um aumento de ΔI ami da primeira para a segunda medição indica a activação proteolítica ENaC. Quimotripsina ou tripsina são conhecidos por estimular maximamente ENaC no sistema de expressão de oócitos 2,14 e pode ser usado para confirm que a activação proteolítica ENaC é detectável em um determinado lote de oócitos.
Em paralelo com as medições de corrente de células completas, uma abordagem biotinilação 9 foi utilizado para investigar se o aumento de ΔI ami detectadas após exposição dos oócitos para proteases se correlaciona com o aparecimento de fragmentos de clivagem ENaC na superfície da célula. As proteínas na superfície da célula são marcados com biotina e pode ser separado a partir de proteínas intracelulares de ligação das proteínas biotiniladas a esferas de agarose neutravidina-rotulados. As proteínas biotiniladas podem ser analisados por western blot. fragmentos de clivagem γENaC na superfície da célula pode ser detectada utilizando um anticorpo específico dirigido contra um epitopo no terminal C do γENaC. Para identificar o local de clivagem funcionalmente relevante (s), previu locais de clivagem pode ser mutado utilizando mutagénese dirigida ao local. Canais de tipo selvagem e mutantes são comparadas em experiências paralelas utilizando oócitos dos slote ame.
Com esta abordagem metodológica foi demonstrado pela primeira vez que a activação proteolítica de correntes de células completas ENaC mediadas correlaciona-se com o aparecimento em função do tempo de ENaC fragmentos de clivagem na superfície da célula. Estes resultados sugerem uma relação causal entre a clivagem canal ea ativação do canal. Além disso, utilizando a mutagénese dirigida ao local de locais de clivagem putativos em combinação com a técnica de dois eléctrodos de voltagem-grampo, locais de clivagem funcionalmente relevantes para a plasmina, quimotripsina e 13 de catepsina S-8 foram identificados.
Protocolo
1. Isolamento de Xenopus Os oócitos e Microinjecção de ARNc
- Obter oócitos de Xenopus laevis fêmea adulta. Anestesiar os animais em 0,2% MS222, e ressecção de lóbulos do ovário, através de uma pequena incisão abdominal.
- Isolar oócitos dos lóbulos ovarianos por digestão enzimática a 19 ° C durante 3-4 horas com 600-700 U / ml de colagenase de tipo 2 a partir de Clostridium histolyticum dissolvidos em solução livre de cálcio OR2 (receita na Tabela 1).
- Para a selecção, colocar os oócitos desfoliculados em uma placa de Petri com um microscópio binocular de um alto teor de sódio contendo uma solução (ND96: Receita na Tabela 1).
- Selecione estágio V-VI oócitos e colocá-los em outra placa de Petri com uma pipeta Pasteur. NOTA: Blunt Pasteur pipeta por chamas para evitar ferimentos oócito.
- Injectar oócitos com cRNA (por exemplo, 0,2 ng por αβγENaC subunidade). Dissolver cRNAs em água livre de RNase. NOTA: Totalvolume injetado em cada ovócito é de 46 nl.
- Loja oócitos injectados a 19 ° C numa solução de baixo teor de sódio para evitar o carregamento de sódio dos oócitos (ND9: Receita na Tabela 1). Complementar a solução com penicilina 100 U / ml de sódio e 100 ug / ml de sulfato de estreptomicina para prevenir crescimento bacteriano. Tratar com cuidado os ovócitos para limitar a quantidade de oócitos danificadas ou mortas e mantê-los em pequenos grupos individuais em uma poços de 12 bem-prato cheio de solução de banho durante os dois dias após a injecção cRNA.
2. Realizar dois eletrodos Experimentos Voltage-clamp
- Medir a oócitos de dois dias após a injecção.
- Encher uma seringa de um sistema de perfusão alimentada por gravidade com uma solução ND96 e outra seringa com solução ND96 contendo amiloride (2 uM). Monte seringas 50 centímetros acima da câmara de banho oócito. NOTA: A concentração de inibidor a amilorida ENaC foi escolhido ser 20 vezes maior do que a IC 50 (100 nM).
- Ligue uma fonte de luz fria halógena de 150 W e ajustá-lo a 10 cm acima da câmara de banho oócito permitindo boa visualização com o microscópio binocular. Em seguida, ligar sucção e ajustar o tubo de sucção no final da câmara do banho de oócito. Localize sucção tubo oposto ao superfusão tubos 'adaptador de entrar no banho oócito. NOTA: A potência de aspiração deve ser suficiente para suportar o fluxo contínuo da solução superfusing o oócito.
- Ajuste a velocidade de superfusão de cada solução de 3-5 ml / min, utilizando o dispositivo de controlo de fluxo iv gravidade. Ligar os tubos de superfusão com um adaptador à câmara de banho de oócito.
- Puxe capilares de vidro com um puxador de micropipeta para obter diâmetros de ponta <1 mícron. Em seguida, preencha capilares para ~ 1/4 com 3 M KCl. NOTA: Certifique-se de que a parte clorada do fio de prata do suporte do eletrodo é imerso em solução de KCl. Verifique se há bolhas de ar na ponta do capilar. As bolhas de ar prejudica o measusentadoria, aumentando a resistência capacitância parasita.
- Insira os capilares para os titulares dos eléctrodos da corrente e do eletrodo de tensão e colocá-los em solução (2 mM) ND96 contendo amilorida usando os micro-manipuladores.
3. Medição das correntes Whole-cell Amiloride sensíveis
- Zero o potencial de eletrodo do eletrodo de tensão (V m) eo eletrodo de corrente (V e), ajustando a V m e V e compensar botões NOTA:. A resistência deve ser de 1-2 mohms para o eletrodo para medir V e 0,5 m -1 mohms para o eletrodo de injeção de corrente.
- Coloque o oócito na câmara do banho em estreita proximidade com o eléctrodo de detecção de tensão. NOTA: Não danifique o oócito durante qualquer uma dessas etapas de transferência. Usar uma pipeta de Pasteur, para transferir o oócito. Para evitar danificar o oócito as bordas da pipeta deve ser anulado pela chama.
- Empalar oocitos suavemente com ambas microeletrodos.
- Definir o potencial de retenção no amplificador para -60 mV e ligar o gravador gráfico. Ligue o amilorida (2 mM) contendo solução. NOTA: A corrente deve ser de cerca de 0 ± 0,5 mA. Correntes de fuga maiores indicam um empalamento gotejante. Portanto, esses óvulos devem ser rejeitados. Além disso, as correntes de fuga medida na presença de amilorida (2 ^ M) deve ser semelhante em αβγ-WT oócitos expressando daqueles medidos em αβγ mutante ENaC expressando oócitos. Isto indica que as mutações não afectam a amilorida-sensibilidade do canal.
- Inicie a gravação. Se necessário, ajustar o ganho.
- Após a corrente medida atinge um patamar estável, alterar a solução livre de amilorida. NOTA: os desvios de corrente descendente nos traços atuais correspondem a correntes de entrada, ou seja, o movimento de carga positiva (Na +) do lado extracelular para dentro da célula.
- After um patamar de corrente for atingido (após ~ 60 seg), mude o superfusão de volta para a solução contendo amilorida. Depois de a corrente do oócito atinge a corrente de linha de base inicial, desligue a braçadeira de tensão e gentilmente retirar os eletrodos.
- Para permitir que a nova selagem da membrana de plasma nos locais da empalação, colocar o oócito em um poço de uma placa de 96 poços, contendo 100-150 mL de solução ND96 protease livre.
- Após 5 min, a transferência do oócito de uma protease contendo uma solução ou uma solução de controlo sem protease para um tempo de incubação de 30 min. NOTA: O tempo de incubação depende da protease e o canal estudado.
- Após a etapa de incubação repetir a medição de corrente (ver 3.2 e seguintes). Nota: é possível medir> 90% dos oócitos após incubação em solução de protease.
4. Biotinilação Ensaio
- Selecione e descartar oócitos defeituosos sob o microscópio binocular. NOTA: Utilize injetared oócitos, a partir do mesmo lote para as medições de corrente e para as experiências de biotinilação.
- Manter biotina à TA durante pelo menos 20 min antes da sua utilização no ensaio.
- Preparar as soluções: ND96 e ND96 contendo a protease apropriada. Prepare pipetas Pasteur, rotulando-os e por flamejante brevemente as suas dicas para evitar lesões dos oócitos. NOTA: Aqui, a protease quimotripsina 2 ug / ml em ND96 é usado. Trate cada grupo com uma pipeta separado para evitar a contaminação cruzada das soluções.
- Encher cada poço de uma placa de 6 poços com 2,5 ml de controlo ou ND96 ND96 contendo uma protease à TA. Em seguida, o depósito de 30 oócitos por poço e incubar durante 30 min à temperatura ambiente. NOTA: Para os processos subsequentes, é importante para manter as amostras em gelo em todos os momentos. Todos os passos de centrifugação a são realizados a 4 ° C.
- Encher cada poço de uma nova placa de 6 poços com 2,5 ml de ND96 (cada grupo tem três poços para os passos de lavagem) e pesar a biotina. NOTA: 2,5 mg biotina per bem (1 mg / ml) é necessário. Dissolve-se a biotina no buffer de biotinilação (ou seja, 25 mg de biotina (para 10 grupos) em 25 ml de tampão de biotinilação (receita na Tabela 1).
- Transferir cada grupo de oócitos de um poço cheio com ND96 2,5 ml. Transferência de ovócitos sequencialmente em dois poços adicionais ND96 para lavar qualquer protease restante. Incubar os oócitos durante 5 min em ND96.
- Transferir os ovócitos para um 2,5 ml de solução de biotina contendo bem e incubar com agitação suave ("shaker") durante 15 min. NOTA: Minimizar o ND96 transferida com pipeta para evitar a diluição da solução de biotina.
- Transferir cada grupo de oócitos dentro de um poço que continha 2,5 ml de tampão de paragem (receita na Tabela 1) para parar a reacção de biotinilação. Em seguida, transferir cada grupo de oócitos em um segundo poço contendo também 2,5 ml de tampão de têmpera e incubar durante 5 min com agitação suave.
- Remover oócitos danificadas ou mortas. Nota:Escolha o mesmo número de oócitos por grupo para o seguinte procedimento.
- Transferir cada grupo de oócitos para um tubo de microcentrífuga de 1,5 mL de plástico. NOTA: Minimizar a quantidade de tampão de têmpera que são transferidos.
- Subsequentemente, lisar os oócitos, passando-os através de uma agulha de 27 G em 1 ml de tampão de lise (receita ver Quadro 1) suplementado com inibidores de protease.
- Centrifugar os lisados durante 10 minutos a 1500 x g.
- Sobrenadante aspirado e transferi-lo para um tubo de microcentrífuga de 1,5 ml contendo 0,5% de Triton-X-100 e 0,5% de NP40. Desprezar o pelete remanescente. NOTA: O sobrenadante contendo proteínas de membrana de plasma biotinilada e as proteínas intracelulares não biotinilados.
- Incubar os tubos de microcentrífuga durante 20 minutos em gelo. Repetidamente vórtice dos tubos durante este período para dissolver completamente as proteínas em NP40 e Triton-X-100.
- Centrifugar 100 ml de pérolas de agarose por grupo de oócitos durante 3 min a 1500 x g. Após oremover o sobrenadante a partir de centrifugação a solução de grânulos e lavar três vezes com tampão de lise para equilibrar as contas com tampão.
- Pipetar 100 l de as esferas lavadas em cada tubo de microcentrífuga contendo o detergente para a solução de proteína preparada de 4.13 para permitir a ligação das proteínas biotinilados às esferas.
- Incubar os tubos de microcentrífuga com sobrecarga de rotação O / N a 4 ° C.
- Centrifugar os tubos de microcentrífuga de 3 min a 1500 x g. Em seguida, transferir o sobrenadante para um novo tubo. NOTA: proteínas intracelulares não são marcados com biotina. O sobrenadante pode ser armazenada a -20 ° C. Não aspirar as contas.
5. Detecção de Fragmentos de ENaC clivagem na superfície celular por Western Blot Análise
- Lavar as pérolas três vezes com tampão de lise e adicionar 100 ul de tampão de amostra de SDS-PAGE 2x. NOTA: As amostras podem ser armazenadas a -20 ° C ou imediatamente preparados para análise de transferência de western.
- Ferva splos de 5 min a 95 ° C e em seguida, coloque os tubos no gelo.
- Centrifugue as amostras durante 3 minutos a 20000 xg e pipeta-se o sobrenadante para um novo tubo de microcentrifugação. NOTA: Este sobrenadante contém as proteínas de membrana de plasma biotinilados a partir da superfície celular do ovócito.
- Analise 30 ul desta sobrenadante por western blot para investigar os fragmentos de clivagem na superfície da célula.
- Separam-se as proteínas biotiniladas por SDS-PAGE (electroforese em gel de dodecil sulfato de sódio poliacrilamida) utilizando um gel apropriado (8%, 10%, 12%, dependendo do peso molecular dos fragmentos de clivagem investigados).
- Transferir as proteínas para difluoreto de polivinilideno (PVDF) por membranas de transferência semi-seco.
- Sondar a membrana com um anticorpo específico contra γENaC humano dirigido contra um epitopo no terminal C (ver a Figura 3 e 13).
- Use de rábano de cabra marcado com peroxidase de anticorpo anti-coelho como anti secundáriocorpo.
- Detectar sinais quimioluminescentes.
Resultados
Para investigar se a plasmina protease de serina pode activar as correntes mediadas por ENaC, ΔI ami de oócitos ENaC expressam individuais foi determinado antes e depois de 30 minutos de incubação dos oócitos em isenta de proteases (controlo) (Figura 2A) ou solução contendo plasmina (figura 2B) utilizando a técnica de dois eléctrodos de tensão-grampo (ver Figura 1). A exposição a plasmina aumento ΔI ami em cada ovócito medido. Em contraste, nas experiências de controlo, 30 minutos de incubação do ENaC expressam oócitos em solução isenta de protease tinha um efeito negligenciável (Figura 2 C, D). Assim, usando este método de uma estimulação de corrente ENaC mediada por plasmina pode ser detectada.
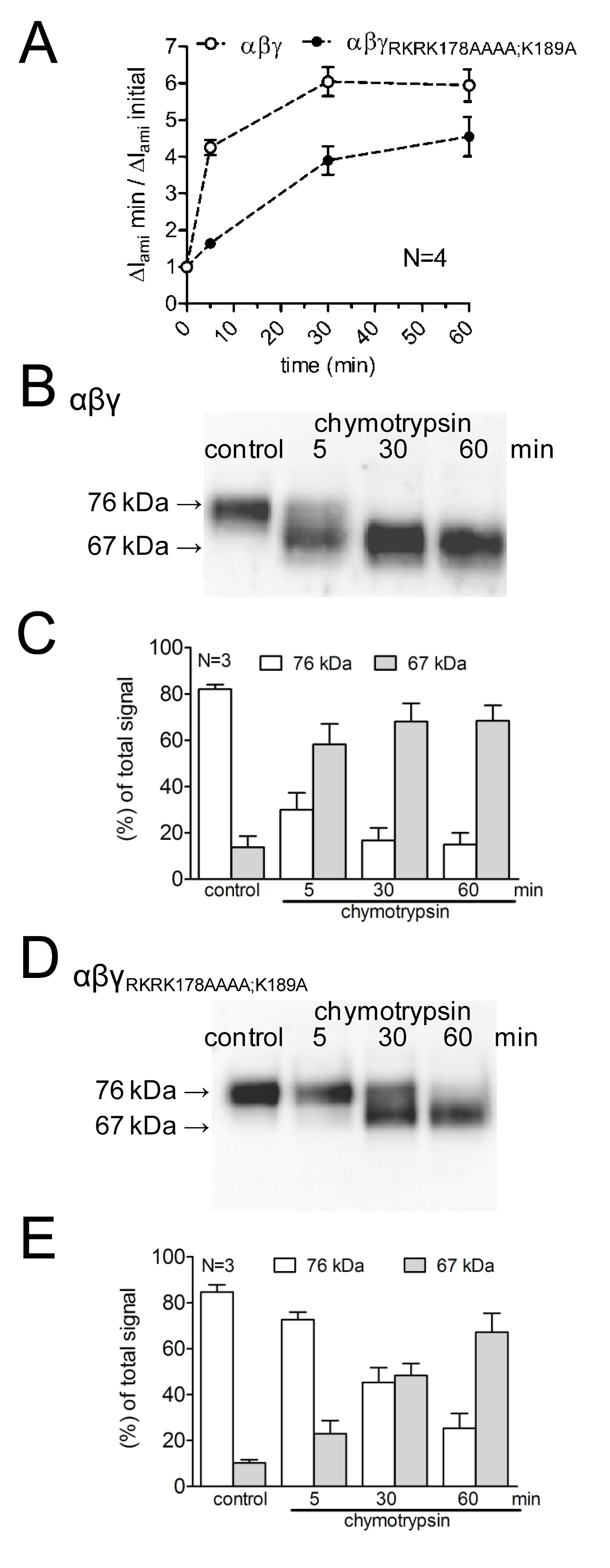
Para estudar os efeitos da mutação de locais de clivagem putativos sobre a activação de correntes mediadas por ENaC, bem como em cima do canal de clivagem, o efeito de quimotripsina em WT-ENaC foi comparada com a dasum mutante com ENaC prostasina mutado e locais de clivagem de plasmina (γ RKRK178AAAA; K189A). O curso de tempo da activação do canal por quimotripsina, assim como o aparecimento de produtos de clivagem de ENaC na superfície celular foi investigada por meio de diferentes tempos de incubação da protease (Figura 4A). Demonstrou-se que os atrasos e canal mutantes reduz a ativação do atual ENaC mediada por quimotripsina. Isto é acompanhado por um aparecimento retardado de um fragmento de clivagem inferior molecular γENaC peso de 67 kDa correspondente à subunidade totalmente clivado. Fragmentos de clivagem foram detectadas utilizando um anticorpo γENaC dirigido contra um epitopo no terminal C (Figura 3). Esta abordagem metodológica demonstra que o curso de tempo da activação proteolítica de correntes mediadas por ENaC correlaciona-se com a aparência de um produto de clivagem de 67 kDa γENaC na superfície celular (Figura 4 B, C). Isso apóia o conceito de umanexo de causalidade entre a clivagem proteolítica canal ea ativação do canal 13. Além disso, através da combinação de medições de corrente e a detecção de fragmentos γENaC na superfície da célula que se demonstrou que os locais de clivagem mutantes são funcionalmente relevantes para a activação proteolítica de canal.

Figura 1. Processo de determinação do efeito estimulante de uma protease em ENaC heterologamente expressos em oócitos de Xenopus laevis. Actividade ENaC é estimada através da medição da corrente de componente de célula inteira sensível à amilorida ΔI ami.

Figura 2. Plasmin estimula moedas ENaC mediadasts em oócitos expressando ENaC. (AD) oócitos expressando ENaC humana foram incubadas durante 30 min em solução isenta de proteases (controlo) ou na solução contendo plasmina (10 ug / ml). Para determinar ΔI ami antes (-) e depois (+) de incubação, os oócitos foram presa a um potencial de retenção de -60 mV (A, B) Quatro representativos de células inteiras a partir de vestígios de corrente um lote de oócitos.. Amiloride (ami) estava presente na solução de banho para inibir especificamente ENaC como indicado por barras pretas. (C) Os pontos de dados obtidos a partir de um ovócito individuais são ligados por uma linha (D). Sumário de experiências similares, como mostrado em C. As colunas representam efeito estimulador em relação ΔI ami calculado como a relação de ΔI ami medida após uma incubação de 30 minutos (ΔI ami 30 min) para o ΔI ami inicial (ΔI ami inicial), medida antes da incubação. Os números dentro das colunas indicam onúmero de oócitos individuais medidos. N indica o número de lotes diferentes de oócitos. (Este valor foi modificado de [Haerteis et al 2012 J Gen Physiol 140, 375-389, doi:. 10.1085/jgp.201110763])

Figura 3. Modelo das subunidades que mostra os locais de clivagem para a activação proteolítica γENaC e o local de ligação do anticorpo usado. Clivagem proteolítica pela furina convertase Golgi-associado é importante para a maturação do ENaC na via biossintética antes do canal atinge a membrana plasmática. Após a clivagem por furina um fragmento de 76 kDa pode ser detectada na superfície da célula através de uma abordagem de biotinilação e um anticorpo contra um epitopo no terminal C da subunidade γ. O passo final crucial na proteolítica PTactivação aC provavelmente ocorre na membrana do plasma onde γENaC é clivada por proteases extracelulares (por exemplo, plasmina ou quimotripsina) em uma região distai para o local de furina, resultando num fragmento de clivagem de 67 kDa. (Este valor foi modificado de [Haerteis et al 2012 J Gen Physiol 140, 375-389, doi:. 10.1085/jgp.201110763])

Figura 4:. Mutação tanto a plasmina (K189) e o local de clivagem de prostasina (RKRK178) atrasa a activação de correntes de ENaC-mediadas e o aparecimento de um produto de clivagem de 67 kDa de γ da subunidade do canal de oócitos que expressam WT (símbolos abertos) e γ RKRK178AAAA; K189A ENaC canal mutante (símbolos a cheio) foram incubadas durante 30 min em solução isenta de proteases (controlo) ou de 5, 30, ou 60 minutos em uma solução contendo quimotripsina (2 ug / ml). (A) Para determinar ΔI ami antes e depois da incubação, os oócitos foram presa a um potencial de retenção de -60 mV. Os círculos representam a proporção de ΔI ami medida após 5, 30, ou 60 minutos de incubação (ΔI ami min) para o ΔI ami inicial (ΔI ami inicial), medida antes da incubação. Cada ponto de dados representa a média ΔI ami medida em 22-24 oócitos individuais de quatro lotes diferentes. (BD) em paralelo para a detecção de ΔI ami, expressão de γENaC biotinilado na superfície celular foi analisada por SDS-PAGE. γENaC foi detectado com um anticorpo contra um epitopo no terminal C de γENaC humano. Western blots representativos de um lote de oócitos são mostrados. (CE) A análise densitométrica das três transferências Western semelhantes aos mostrados em B ou D. Para cada pista, as signals detectados na região de 76 kD (colunas a branco) e 67 kD (colunas cinzentas) foram determinadas e normalizadas para a soma do sinal total detectada. N indica o número de lotes diferentes de oócitos. clique aqui para ampliar.
{kind=link}
Discussão
Neste manuscrito uma abordagem metodológica que foi aplicado com sucesso para estudar os mecanismos subjacentes a ativação de ENaC por proteases é descrito 8,13. O sistema de expressão laevis oócito Xenopus bem estabelecida foi usado para funcionalmente expressa ENaC. Função ENaC foi avaliada com a técnica de dois eletrodos de tensão-clamp convencional. A mutagénese dirigida foi utilizada para identificar os locais de clivagem da protease funcionalmente relevantes. Biotinilação experiências realizadas em paralelo com as medições electrofisiológicas tornou possível correlacionar o aparecimento de ENaC clivagem produtos na superfície da célula, com a activação proteolítica de corrente. Uma correlação entre o curso de tempo da activação actual e o aparecimento de fragmentos de clivagem proteolitica na superfície da célula suporta o conceito de activação do canal proteolítica.
Dois eléctrodos de gravação voltagem-grampo requerem a empalação de um oocyte com dois microeletrodos. Este procedimento é geralmente realizado apenas uma vez em um ovócito individual. No entanto, foi possível remover os microeléctrodos após uma gravação de corrente inicial de células inteiras sem dano aparente ao oócito. Com efeito, a membrana de plasma nos locais da impalements parece selar dentro de poucos minutos. Assim, após a conclusão de uma medição de tensão-grampo primeiro de dois eléctrodos, é possível transferir o ovócito da câmara de fluxo experimental da configuração de tensão-grampo de dois eléctrodos para um tubo de microcentrífuga ou um poço de uma placa de 96 poços preenchidos com um pequeno volume de solução de teste ou de controlo. Em seguida, o mesmo oócito pode ser transferido de volta para a câmara de fluxo e pode ser empalado novamente para executar uma segunda medição de tensão-grampo de dois eléctrodos. Notavelmente, as correntes mediadas por ENaC não variar muito entre a primeira e segunda medição, quando o oócito foi mantido em solução de controlo. Em contraste, a incubação do oócito em uma protease contendo sollução após a primeira medição, resultou num aumento da corrente de ENaC mediada na segunda medição (Figura 2). Este achado indica a ativação do canal proteolítica.
Realizando duas medições de correntes separadas em um único oócito, oferece a vantagem de que o oócito pode ser exposta a proteases ou outros agentes farmacológicos entre as duas medições para um comprimento variável de tempo, num pequeno volume de solução de teste. Isto é importante quando se utiliza agentes que são caros e / ou disponível em grandes quantidades, por exemplo, preparações de protease purificadas. A disponibilidade limitada de agentes pode tornar impossível (ou inviável) para usá-los em contínuas de dois eletrodos gravações tensão-clamp por causa dos grandes volumes de solução de teste necessário para superfusing continuamente os oócitos com vazões de vários mililitros por minuto. Por outro lado, de dois eléctrodos de medição de tensão contínua de braçadeira estão limitados pelo fenó conhecidomeno de resumo canal espontânea também descrito para ENaC 15. Em contraste, a exposição oócitos para testar soluções entre duas medições separadas de até uma hora ou mais geralmente não constitui um problema (ver Figura 4A). Finalmente, duas medidas seqüenciais realizados na mesma oócito permitir observações pareadas de efeitos de drogas. Isto tem uma vantagem sobre as medições desemparelhados de dois grupos distintos de oócitos (-tratados e tratados com protease do veículo), uma vez que reduz o problema da alta variabilidade entre oócitos, normalmente observadas na expressão de canal iónico. Com pares de observação e a possibilidade de normalizar os dados para a primeira medição, são necessários menos oócitos por grupo experimental para demonstrar um efeito significativo de um agente farmacológico. A normalização dos dados também faz com que seja fácil de sintetizar os dados a partir de diferentes lotes de oócitos com diferentes níveis de expressão de canais de iões e, consequentemente, diferentes correntes de linha de base (Figura 2D). Obviamente, as experiências de controlo são necessários para esta abordagem para demonstrar que a actividade de canal iónico de interesse permanece estável em oócitos de controlo tratado com veículo a partir da primeira para a segunda medição (ver Figura 2).
Para demonstrar que a activação proteolítica corrente correlaciona com o aparecimento de produtos de clivagem de ENaC na superfície da célula, uma abordagem biotinilação originalmente descrita por Harris et al. Nove pode ser utilizada. Este procedimento (como detalhado na secção de protocolo e mostrado na Figura 4) foi adaptado para demonstrar que a exposição de canais de proteases e subsequente activação de correntes mediadas por ENaC é paralelo com o aparecimento em função do tempo de fragmentos de clivagem. O método de biotinilação também permite a análise de um aumento ou diminuição global das proteínas de membrana na superfície da célula. Assim, este método é adequado para investigar o efeito de proteases e outras farmaagentes farmacológicas sobre a inserção do canal na membrana plasmática ou sobre a recuperação do canal. Além disso, a análise Western blot das proteínas da membrana de plasma biotiniladas permite a detecção de fragmentos de proteína (por exemplo, fragmentos proteolíticos ENaC) ou alterações no padrão de glicosilação, que pode ser funcionalmente relevantes.
Em conclusão, a combinação de métodos utilizados para investigar o efeito estimulador de proteases sobre correntes de células completas ENaC-mediadas e para demonstrar uma correlação com a ocorrência de clivagem ENaC produtos na superfície da célula pode ser útil para uma ampla gama de aplicações. Em particular, esses métodos podem ser adequados para tratar de questões semelhantes no que respeita à regulação de outros canais iônicos, transportadores ou receptores transmembrana (por exemplo, receptores ativados por protease pars).
Divulgações
The authors have nothing to disclose.
Agradecimentos
The expert technical assistance of Céline Grüninger, Christina Lang, Sonja Mayer, and Ralf Rinke is gratefully acknowledged. We thank Dr. Morag K. Mansley for carefully reading the manuscript. This project was supported by a grant of the Deutsche Forschungsgemeinschaft (Grant SFB 423: Kidney Injury: Pathogenesis and Regenerative Mechanisms, to C. Korbmacher), grants of the Interdisziplinäres Zentrum für Klinische Forschung (to S. Haerteis and M. Krappitz), the ELAN program (to S. Haerteis) of the Friedrich-Alexander-Universität Erlangen-Nürnberg, and the University Library of Erlangen-Nürnberg.
Materiais
| Name | Company | Catalog Number | Comments |
| Bath Clamp Headstage for OC-725C-V | Warner Instrument Corporation | - | |
| Cold light source - Schott KL 1500 LCD | Schott | #SCOC150200EU | brightness 4; mechanical aperture: D; color temperature: 3000 K |
| E Series Electrode Holder (Str, Vent, Ag Wire, 1.2 mm) | ADinstruments | #ESW-F10v | |
| left micromanipulator; MM-33L | Warner Instrument Corporation | #64-0055 | |
| LIH 1600 - computer interface | HEKA | - | |
| magnetic valve system (ALA BPS-8) in combination with a TIB14 interface (HEKA) | ALA Scientific Instruments, HEKA | - | |
| OC-725C amplifier for two-electrode voltage-clamp recordings | Warner Instrument Corporation | - | |
| P-97 FLAMING/BROWN Micropipette Puller | Sutter Instruments | - | heat=550; velocity=22; time=200 |
| right micromanipulator; MM-33R | Warner Instrument Corporation | #64-0056 | |
| Series Electrode Holder (45°, Vent, Handle, Ag Wire, 1.2 mm) | ADinstruments | #E45w-f10vh | |
| STAT 2 IV Gravity Flow Controller | Conmed | #P-S2V-60 | |
| vacuum generator ejector SEG - for suction to remove bath solution | Schmalz | - | |
| Material | |||
| INFUJECT 60ml pump syringes for solutions | Braun | #22050 | |
| Injekt-F for lysing the oocytes | Braun | #9166033V | |
| standard wall borosilicate tubing with filament | Sutter Instruments | #BF150-86-10 | outside diameter: 1.50 mm; inside diameter: 0.86 mm; length: 10 cm |
| Reagent | |||
| complete, Mini, EDTA-free protease inhibitor cocktail tablets | Roche Applied Science | #11836170001 | |
| EZ-Link Sulfo-NHS-Biotin | Thermo Scientific | #21217 | |
| Horseradish peroxidase-labeled secondary goat anti-rabbit antibody | Santa Cruz Biotechnology | #sc-2004 | |
| NeutrAvidin Agarose | Thermo Scientific | #29200 | Neutravidin-labeled agarose beads |
| NP40 (Nonidet P-40) | Sigma-Aldrich | #I8896 | |
| Roti-Load 1 (2× SDS-PAGE sample buffer) | Carl Roth | #K929.2 | |
| SuperSignal West Femto Chemiluminescent Substrate for detection of chemiluminescent signals | Thermo Scientific | #34095 | |
| Triton-X-100 | Sigma-Aldrich | #T8787 | |
Referências
- Kleyman, T. R., Carattino, M. D., Hughey, R. P. ENaC at the cutting edge: regulation of epithelial sodium channels by proteases. The Journal of Biological Chemistry. 284, 20447-20451 (2009).
- Rossier, B. C., Stutts, M. J. Activation of the epithelial sodium channel (ENaC) by serine proteases. Annu Rev Physiol. 71, 361-379 (2009).
- Poirot, O., Vukicevic, M., Boesch, A., Kellenberger, S. Selective regulation of acid-sensing ion channel 1 by serine proteases. The Journal of Biological Chemistry. 279, 38448-38457 (2004).
- Vukicevic, M., Weder, G., Boillat, A., Boesch, A., Kellenberger, S. Trypsin cleaves acid-sensing ion channel 1a in a domain that is critical for channel gating. The Journal of Biological Chemistry. 281, 714-722 (2006).
- Clark, E. B., Jovov, B., Rooj, A. K., Fuller, C. M., Benos, D. J. Proteolytic cleavage of human acid-sensing ion channel 1 by the serine protease matriptase. The Journal of Biological Chemistry. 285, 27130-27143 (2010).
- Ossovskaya, V. S., Bunnett, N. W. Protease-activated receptors: contribution to physiology and disease. Physiological reviews. 84, 579-621 (2004).
- Garcia-Caballero, A., et al. Activation of the epithelial sodium channel by the metalloprotease meprin β-subunit. Channels (Austin. 5, 14-22 (2011).
- Haerteis, S., et al. Proteolytic activation of the epithelial sodium channel (ENaC) by the cysteine protease cathepsin-S. Pflügers Archiv: European Journal of Physiology. 464, 353-365 (2012).
- Harris, M., Firsov, D., Vuagniaux, G., Stutts, M. J., Rossier, B. C. A novel neutrophil elastase inhibitor prevents elastase activation and surface cleavage of the epithelial sodium channel expressed in Xenopus laevis oocytes. The Journal of Biological Chemistry. 282, 58-64 (2007).
- Passero, C. J., Mueller, G. M., Rondon-Berrios, H., Tofovic, S. P., Hughey, R. P., Kleyman, T. R. Plasmin activates epithelial Na+ channels by cleaving the γ-subunit. The Journal of Biological Chemistry. 283, 36586-36591 (2008).
- Svenningsen, P., et al. Plasmin in nephrotic urine activates the epithelial sodium channel. Journal of the American Society of Nephrology : JASN. 20, 299-310 (2009).
- Patel, A. B., Chao, J., Palmer, L. G. Tissue kallikrein activation of the epithelial Na channel. American Journal of Physiology. Renal Physiology. 303, (2012).
- Haerteis, S., Krappitz, M., Diakov, A., Krappitz, A., Rauh, R., Korbmacher, C. Plasmin and chymotrypsin have distinct preferences for channel activating cleavage sites in the γ-subunit of the human epithelial sodium channel. The Journal of General Physiology. 140, 375-389 (2012).
- Chraibi, A., Vallet, V., Firsov, D., Hess, S. K., Horisberger, J. D. Protease modulation of the activity of the epithelial sodium channel expressed in Xenopus oocytes. The Journal of General Physiology. 111, 127-138 (1998).
- Volk, T., Konstas, A. A., Bassalay, P., Ehmke, H., Korbmacher, C. Extracellular Na+ removal attenuates rundown of the epithelial Na+-channel (ENaC) by reducing the rate of channel retrieval. Pflügers Archiv: European Journal of Physiology. 447, 884-894 (2004).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados