Method Article
Dimostrazione di Proteolytic attivazione della epiteliale di sodio Channel (ENAC), combinando misure di corrente con rilevamento di Decolleté Frammenti
In questo articolo
Riepilogo
L'attivazione proteolitica del canale epiteliale del sodio (ENaC) eterologhi espressi in oociti di Xenopus laevis sono evidenziabili combinando misure di corrente con un approccio biotinylation per indagare la comparsa di prodotti di scissione del canale ionico sulla superficie cellulare. Funzionalmente importanti siti di taglio possono essere identificati usando mutagenesi sito-diretta.
Abstract
The described methods can be used to investigate the effect of proteases on ion channels, receptors, and other plasma membrane proteins heterologously expressed in Xenopus laevis oocytes. In combination with site-directed mutagenesis, this approach provides a powerful tool to identify functionally relevant cleavage sites. Proteolytic activation is a characteristic feature of the amiloride-sensitive epithelial sodium channel (ENaC). The final activating step involves cleavage of the channel’s γ-subunit in a critical region potentially targeted by several proteases including chymotrypsin and plasmin. To determine the stimulatory effect of these serine proteases on ENaC, the amiloride-sensitive whole-cell current (ΔIami) was measured twice in the same oocyte before and after exposure to the protease using the two-electrode voltage-clamp technique. In parallel to the electrophysiological experiments, a biotinylation approach was used to monitor the appearance of γENaC cleavage fragments at the cell surface. Using the methods described, it was demonstrated that the time course of proteolytic activation of ENaC-mediated whole-cell currents correlates with the appearance of a γENaC cleavage product at the cell surface. These results suggest a causal link between channel cleavage and channel activation. Moreover, they confirm the concept that a cleavage event in γENaC is required as a final step in proteolytic channel activation. The methods described here may well be applicable to address similar questions for other types of ion channels or membrane proteins.
Introduzione
Le proteasi sono enzimi che sono coinvolti in varie reazioni fisiologiche che vanno dal noto degradazione proteolitica delle proteine, nel contesto della digestione, alle sofisticate cascate proteasi implicate in vie di segnalazione regolamentazione complesse. Le proteasi sono classificati in sette gruppi in base al loro sito attivo catalitico: aspartato, asparagina, cisteina, acido glutammico, metallo, serina, treonina e proteasi. Diverse proteasi bersaglio siti di taglio distinte non sempre facile prevedere dalla struttura primaria di una proteina. Il database MEROPS ( http://merops.sanger.ac.uk/ ) fornisce informazioni dettagliate su una vasta gamma di proteasi e dei loro siti di taglio preferenziali. Siti di clivaggio funzionalmente rilevanti possono essere identificati mediante mutagenesi sito-diretta.
E 'ben noto che l'elaborazione proteolitica dell'Enac è un importante meccanismo di attivazione del secoloè particolare 1,2 canale ionico. È interessante notare, è evidente che la funzione del relativo acido-sensing canale ionico 1a (ASIC1a) può anche essere modificato da proteasi 3-5. Al momento resta una questione aperta se la scissione proteolitica canale gioca un ruolo fisiologico rilevante nel regolare l'attività di altri canali ionici e trasportatori. Tuttavia, è ben noto che clivaggio proteolitico attiva un gruppo di G recettori accoppiati alla proteina, i recettori proteasi-attivato (PARs) 6. Diversi serina-proteasi (ad esempio proteasi canale attivazione (CAP1-3), chimotripsina, tripsina, Furin, plasmina, neutrofili elastasi e callicreina) sono stati indicati per attivare proteoliticamente ENaC 2. Oltre alla serina proteasi, altri gruppi di proteasi possono essere coinvolti nella attivazione proteolitica ENaC. Infatti, dati recenti mostrano che la metalloproteinasi meprin-β 7 e la cisteina proteasi catepsina S-8 può anche activaTE ENaC. Tuttavia, le (pato-proteasi) fisiologicamente rilevanti per l'attivazione ENaC restano da definire e possono differire da tessuto a tessuto.
Le proteasi sono noti per scindere preferenzialmente in particolari siti nella sequenza aminoacidica. Per esempio, la chimotripsina serina proteasi mostra un modello specifico di clivaggio clivaggio dopo il aminoacidi aromatici residui fenilalanina e tirosina. Al contrario, la serina proteasi tripsina preferenzialmente scinde dopo la residui di lisina o arginina base. Utilizzando mutanti costrutti γENaC umano generati da mutagenesi sito-diretta, siti di clivaggio funzionalmente pertinenti dell'ENaC eterologhi espressi nel sistema di espressione ovocita potrebbero essere identificati 8-13.
Iniettando cRNA per i tre subunità dell'ENaC (αβγ) in oociti isolati, ENaC può esprimere funzionalmente in queste cellule e l'attività dei canali presenti sulla membrana plasmatica può essere misurata medianteusando la tecnica voltage-clamp a due elettrodi. Utilizzando il amiloride diuretico, un inibitore ENaC specifico, il ENaC-mediata componente corrente whole-cell amiloride-sensibile (ΔI ami) possono essere separati dalle correnti di fuga aspecifiche o da correnti condotte da altri canali ionici. Così, i valori ΔI ami riflettono attività complessiva ENaC e possono essere determinati sottraendo correnti cellule intere misurati in presenza di amiloride dalle correnti di cellule intere corrispondenti registrate in assenza di amiloride. Per verificare se una proteasi ha un effetto stimolante sulla ENaC, ΔI ami viene misurato due volte nello stesso ovocita, cioè prima e dopo incubazione della ovocita in una soluzione contenente proteasi. Un aumento di ΔI ami dalla prima alla seconda misura indica l'attivazione proteolitica ENaC. Chimotripsina tripsina o sono noti per stimolare massimo dell'ENaC nel sistema di espressione ovocita 2,14 e possono essere usati per confirm che l'attivazione proteolitica ENaC è rilevabile in un determinato lotto di ovociti.
In parallelo a misure di corrente di cellule intere, un approccio biotinilazione 9 è stato utilizzato per studiare se l'aumento ΔI ami rilevata dopo esposizione degli ovociti alle proteasi correla con la comparsa di dell'ENaC clivaggio frammenti sulla superficie cellulare. Proteine sulla superficie cellulare sono etichettati con biotina e possono essere separati dalle proteine intracellulari legando le proteine biotinilati di agarosio NeutrAvidin-etichettati. Le proteine biotinilati possono essere analizzati mediante western blot. frammenti di scissione γENaC sulla superficie cellulare possono essere rilevate utilizzando un anticorpo specifico diretto contro un epitopo nel C-terminale della γENaC. Per identificare sito di scissione funzionalmente rilevanti (s), previsto siti di taglio possono essere mutati mediante mutagenesi sito-diretta. Canali di tipo selvatico e mutante sono confrontate in esperimenti paralleli utilizzando ovociti dalle slotto ame.
Con questo approccio metodologico è stato dimostrato per la prima volta che l'attivazione proteolitica di correnti di cellule intere dell'ENaC-mediate correla con la comparsa dipendente dal tempo di dell'ENaC clivaggio frammenti sulla superficie cellulare. Questi risultati suggeriscono un nesso di causalità tra la scissione del canale e l'attivazione del canale. Inoltre, mediante mutagenesi sito-diretta di putativi siti di taglio in combinazione con la tecnica a due elettrodi voltage-clamp, siti di clivaggio funzionalmente rilevanti per plasmina, chimotripsina e catepsina 13-S 8 sono stati identificati.
Protocollo
1. Isolamento di Xenopus ovociti e microiniezione di cRNA
- Ottenere ovociti da adulto di sesso femminile Xenopus laevis. Anestetizzare gli animali nel 0,2% MS222, e resecare lobi ovarici attraverso una piccola incisione addominale.
- Isolare ovociti dai lobi ovarici mediante digestione enzimatica a 19 ° C per 3-4 ore con 600-700 U / ml di collagenasi di tipo 2 Clostridium histolyticum disciolti in soluzione OR2 calcio libero (ricetta in Tabella 1).
- Per la selezione, posizionare gli ovociti defolliculated in una capsula di Petri sotto un microscopio binoculare in una soluzione contenente sodio alta (ND96: ricetta in Tabella 1).
- Selezionare fase V-VI ovociti e metterli in un altro piatto di Petri con una pipetta Pasteur. NOTA: Blunt pipetta Pasteur da fiamme per evitare lesioni ovocita.
- Iniettare ovociti con cRNA (ad esempio 0,2 ng per αβγENaC subunità). Sciogliere cRNA in acqua RNase-free. NOTA: Totalvolume iniettato in ciascun ovocita è di 46 nl.
- Conservare iniettato ovociti a 19 ° C in una soluzione di sodio bassa da evitare il caricamento di sodio degli ovociti (ND9: ricetta in Tabella 1). Integrare la soluzione con penicillina 100 U / ml di sodio e 100 pg / ml di solfato di streptomicina per prevenire la crescita batterica. Maneggiare con cura gli ovociti per limitare la quantità di ovociti danneggiati o morti e mantenerli in singoli piccoli gruppi in un pozzo di 12 pozzetti riempiti con soluzione del bagno durante i due giorni dopo l'iniezione cRNA.
2. Esecuzione a due elettrodi Esperimenti di tensione-clamp
- Misurare la ovociti due giorni dopo l'iniezione.
- Riempire una siringa di un sistema di perfusione gravità alimentata con soluzione ND96 e un'altra siringa con soluzione ND96 contenente amiloride (2 mM). Mount siringhe 50 centimetri sopra la camera da bagno ovocita. NOTA: La concentrazione del amiloride inibitore ENaC è stato scelto per essere 20 volte superiore alla IC50 (100 nm).
- Accendere una sorgente di luce fredda alogena 150 W e regolarlo a 10 cm al di sopra della camera di bagno ovocita che consente una buona visualizzazione con il microscopio binoculare. Quindi accendere aspirante e regolare il tubo di aspirazione alla fine della camera di bagno ovocita. Individuare tubo di aspirazione opposta a superfusione tubi 'adattatore di entrare nel bagno ovocita. NOTA: Potenza di aspirazione deve essere sufficiente a sostenere il flusso continuo della soluzione superfusing l'ovocita.
- Regolare la velocità superfusione di ciascuna soluzione di 3-5 ml / min utilizzando il dispositivo di controllo del flusso iv gravità. Collegare i tubi superfusione con un adattatore alla camera bagno ovocita.
- Tirare capillari di vetro con un estrattore micropipetta per ottenere diametri di punta di <1 micron. Poi riempire capillari a ~ 1/4 con 3 M KCl. NOTA: Assicurarsi che la parte clorurato del filo d'argento del titolare elettrodo è immerso in soluzione KCl. Verificare la presenza di bolle d'aria nella punta del capillare. Le bolle d'aria compromettono la misurement aumentando la resistenza capacità parassite.
- Inserire i capillari nei supporti elettrodi della corrente e l'elettrodo di tensione e metterli in ND96 contenente amiloride soluzione (2 micron) utilizzando i micro-manipolatori.
3. Misura delle Correnti Whole-cell Amiloride-sensibili
- Zero potenziale dell'elettrodo dell'elettrodo tensione (V m) e l'elettrodo di corrente (V e) regolando la V m e V e compensati pulsanti. NOTA: La resistenza deve essere 1-2 mW per l'elettrodo per misurare V e 0,5 m -1 mW per l'elettrodo di iniezione di corrente.
- Posizionare l'ovocita nella camera bagno in prossimità dell'elettrodo rilevamento della tensione. NOTA: Non danneggiare l'ovocita durante una di queste operazioni di trasferimento. Utilizzare una pipetta Pasteur per trasferire l'ovocita. Per evitare di danneggiare l'ovocita i bordi della pipetta deve essere smussati mediante esposizione alla fiamma.
- Impale oociti delicatamente con entrambe le microelettrodi.
- Impostare la potenziale partecipazione al amplificatore a -60 mV e accendere il registratore grafico. Accendere il amiloride (2 micron) contenente soluzione. NOTA: La corrente dovrebbe essere di circa 0 ± 0,5 μA. Correnti di fuga più grandi indicano un impalamento perde. Pertanto, questi ovociti devono essere respinti. Inoltre, le correnti di fuga misurata in presenza di amiloride (2 mM) dovrebbero essere simili in αβγ-WT esprimendo ovociti a quelle misurate in αβγ-mutante ENaC esprimere oociti. Ciò indica che le mutazioni non influiscono sulla amiloride-sensibilità del canale.
- Avviare la registrazione. Se necessario regolare il guadagno.
- Dopo la corrente misurata raggiunge un plateau stabile, passare alla soluzione senza amiloride. NOTA: deviazioni in corso al ribasso le tracce attuali corrispondono a correnti interiori, vale a dire il movimento di carica positiva (Na +) dal lato extracellulare nella cellula.
- After un plateau di corrente viene raggiunto (dopo ~ 60 sec), accendere il superfusione torna alla soluzione contenente amiloride. Dopo la corrente del ovocita raggiunge la corrente di linea di base iniziale, spegnere il morsetto di tensione e delicatamente ritirare gli elettrodi.
- Per consentire richiusura della membrana plasmatica nei siti di impalamento, posizionare l'ovocita in un pozzetto di una piastra a 96 pozzetti contenente 100-150 ml di soluzione ND96 proteasi gratuito.
- Dopo 5 min, trasferire oocita una proteasi contenente soluzione o di una soluzione di controllo senza proteasi per un tempo di incubazione di 30 min. NOTA: Il tempo di incubazione dipende dalla proteasi e il canale studiato.
- Dopo la fase di incubazione ripetere la misura della corrente (vedere 3.2 e seguenti). NOTA: È possibile misurare> 90% degli ovociti dopo incubazione in soluzione di proteasi.
4. Biotinilazione Assay
- Selezionare e scartare ovociti difettosi sotto il microscopio binoculare. NOTA: Utilizzare InjectEd ovociti dallo stesso lotto per le misure di corrente e per gli esperimenti biotinilazione.
- Mantenere biotina a TA per almeno 20 minuti prima del suo uso nell'esperimento.
- Preparare le soluzioni: ND96 e ND96 contenente la proteasi appropriato. Preparare pipette Pasteur assegnando un'etichetta e brevemente fiammeggiante loro consigli per evitare lesioni degli ovociti. NOTA: Qui, viene utilizzata la proteasi chimotripsina 2 ug / ml in ND96. Trattare ogni gruppo con una pipetta separato per evitare la contaminazione incrociata di soluzioni.
- Riempire ogni pozzetto di una piastra da 6 pozzetti con 2,5 ml controllo ND96 ND96 o contenente una proteasi a RT. Poi depositare 30 ovociti per bene e li incubare per 30 minuti a temperatura ambiente. NOTA: Per le procedure successive è importante mantenere i campioni in ghiaccio in ogni momento. Tutte le fasi di centrifugazione sono al eseguite a 4 ° C.
- Riempire ogni pozzetto di una nuova piastra da 6 pozzetti con 2,5 ml ND96 (ogni gruppo ha bisogno di 3 pozzi per le fasi di lavaggio) e pesare la biotina. NOTA: 2,5 mg biotina per bene è necessaria (1 mg / ml). Sciogliere la biotina nel buffer biotinylation (cioè 25 mg biotina (per 10 gruppi) in 25 ml di tampone biotinylation (ricetta in Tabella 1).
- Trasferire ciascun gruppo di oociti di un pozzo riempito con 2,5 ml ND96. Trasferimento ovociti sequenziale in due pozzi aggiuntivi con ND96 per lavare via ogni residuo della proteasi. Incubare gli ovociti per 5 min in ND96.
- Trasferire gli ovociti in un pozzo contenente 2,5 ml di soluzione di biotina e incubare con leggera agitazione ('shaker') per 15 min. NOTA: Ridurre al minimo l'ND96 trasferita con la pipetta per evitare la diluizione della soluzione di biotina.
- Trasferire ciascun gruppo di oociti in un pozzo contenente 2,5 ml di tampone quench (ricetta in Tabella 1) per arrestare la reazione biotinilazione. Poi, il trasferimento di ciascun gruppo di ovociti in un secondo pozzo anche contenente 2,5 ml di placare il buffer e incubare per 5 minuti con agitazione.
- Rimuovere ovociti danneggiati o morti. Nota:Scegliere lo stesso numero di ovociti per gruppo per la seguente procedura.
- Trasferire ciascun gruppo di oociti in una provetta di plastica da 1,5 ml. NOTA: Ridurre al minimo la quantità di buffer di tempra che viene trasferito.
- Successivamente, lisare gli ovociti facendoli passare attraverso un ago 27 G in 1 ml di tampone di lisi (ricetta vedi Tabella 1) integrato con inibitori della proteasi.
- Centrifugare i lisati per 10 min a 1500 x g.
- Aspirare il surnatante e trasferirlo in una provetta da 1,5 ml microcentrifuga contenente 0,5% Triton-X-100 e 0,5% NP40. Eliminare il pellet rimanente. NOTA: Il surnatante contiene proteine di membrana del plasma biotinilato e proteine intracellulari non biotinilato.
- Incubare le provette da microcentrifuga per 20 min in ghiaccio. Ripetutamente vortex le provette durante questo periodo per sciogliere completamente le proteine NP40 e Triton-X-100.
- Centrifuga 100 ml di agarosio per gruppo di ovociti per 3 minuti a 1500 x g. Dopo l'centrifugazione Rimuovere il surnatante dalla soluzione perline e lavare tre volte con tampone di lisi per equilibrare le perline con tampone.
- Pipettare 100 ml di perline lavati in ciascuna provetta contenente la proteina-detergente-soluzione preparata in 4.13 per consentire il legame delle proteine biotinilati verso le perline.
- Incubare le provette per microcentrifuga con testa di rotazione O / N a 4 ° C.
- Centrifugare le provette da microcentrifuga per 3 minuti a 1500 x g. Poi trasferire il surnatante in una nuova provetta. NOTA: le proteine intracellulari non sono etichettati con biotina. Il supernatante può essere conservato a -20 ° C. Non aspirare le perline.
5. Rilevazione dell'Enac Decolleté frammenti sulla superficie cellulare tramite Western Blot analisi
- Lavare le perline tre volte con tampone di lisi e aggiungere 100 ml di 2x tampone campione SDS-PAGE. NOTA: I campioni possono essere conservati a -20 ° C o immediatamente preparati per l'analisi Western Blot.
- Bollire samples per 5 min a 95 ° C e posizionare i tubi su ghiaccio.
- Centrifugare i campioni per 3 min a 20.000 xg e pipetta il surnatante in una nuova provetta. NOTA: Questo surnatante contiene le proteine della membrana plasmatica biotinilati dalla superficie cellulare dell'ovocita.
- Analizzare 30 ml di questa supernatante mediante western blot per indagare frammenti scissione sulla superficie cellulare.
- Separare le proteine biotinilati mediante SDS-PAGE (sodio dodecil solfato elettroforesi su gel di poliacrilammide) utilizzando un gel appropriato (8%, 10%, 12% a seconda del peso molecolare dei frammenti di scissione indagati).
- Trasferire le proteine di polivinilidene (PVDF), membrane per blotting semi-asciutto.
- Sondare la membrana con un anticorpo specifico contro γENaC umano diretto contro un epitopo nel C-terminale (vedi figura 3 e 13).
- Utilizzare anticorpo anti-coniglio perossidasi di rafano-marcato capra come anti secondariocorpo.
- Rilevare i segnali chemioluminescenti.
Risultati
Per verificare se la plasmina serina proteasi può attivare correnti ENAC-mediate, ΔI ami di singoli ovociti ENAC-esprimono è stata determinata prima e dopo 30 min di incubazione degli ovociti in libera-proteasi (controllo) (Figura 2A) o una soluzione contenente plasmina (Figura 2B) mediante la tecnica voltage-clamp a due elettrodi (vedere Figura 1). L'esposizione a plasmina è aumentata ΔI ami in ogni ovocita misurato. Al contrario, in esperimenti di controllo, 30 min di incubazione di ovociti dell'ENaC-esprimenti in soluzione priva di proteasi hanno un effetto trascurabile (Figura 2 C, D). Così, utilizzando questo metodo una stimolazione di corrente dell'ENaC mediata da plasmina può essere rilevato.
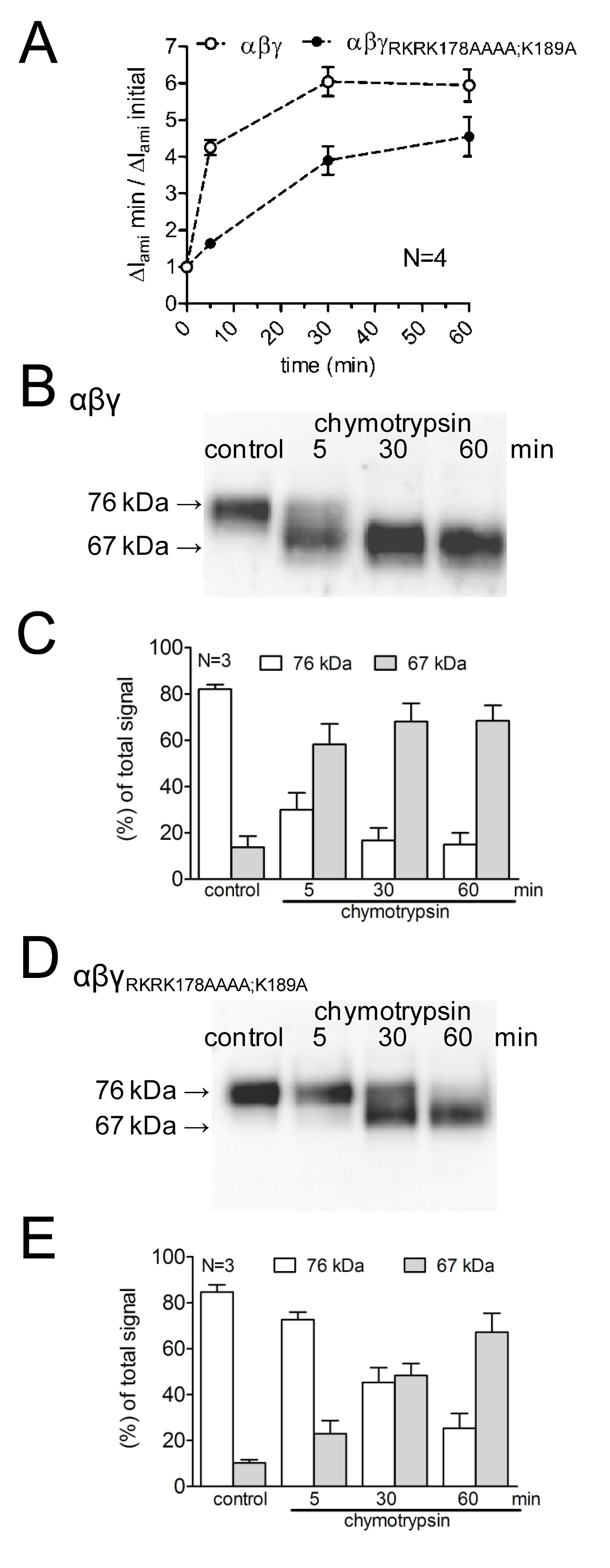
Per studiare gli effetti di mutanti putativi siti di clivaggio dall'attivazione delle correnti ENaC-mediata, così come su clivaggio canale, l'effetto della chimotripsina su WT-ENaC è stato confrontato con quello relativoun ENaC mutante con prostasin mutato e siti di taglio plasmina (γ RKRK178AAAA; K189A). La durata di attivazione del canale da chimotripsina e la comparsa di dell'ENaC clivaggio prodotti sulla superficie cellulare è stata studiata utilizzando diversi tempi di incubazione proteasi (Figura 4A). È stato dimostrato che i ritardi canali mutanti e riduce l'attivazione di corrente dell'ENaC mediata da chimotripsina. Parallelamente si ha una comparsa ritardata di un peso γENaC scissione frammento molecolare inferiore di 67 kDa corrispondente alla subunità completamente scissa. Frammenti di scissione sono state rilevate utilizzando un anticorpo γENaC diretto contro un epitopo nel C-terminale (Figura 3). Questo approccio metodologico dimostra che l'andamento nel tempo di attivazione proteolitica delle correnti dell'ENaC-mediate correla con la comparsa di un prodotto di scissione 67 kDa γENaC sulla superficie cellulare (Figura 4 B, C). Questo supporta il concetto di unnesso di causalità tra scissione proteolitica canale e l'attivazione del canale 13. Inoltre, combinando misure di corrente e la rivelazione di frammenti γENaC alla superficie delle cellule è stato dimostrato che i siti di clivaggio mutati sono funzionalmente rilevante per l'attivazione del canale proteolitica.

Figura 1. Procedura di determinare l'effetto stimolatorio di una proteasi su dell'ENaC eterologhi espressi in oociti di Xenopus laevis. Attività dell'ENaC è stimata misurando la cellula intera componente corrente amiloride sensibile ΔI ami.

Figura 2. Plasmina stimola valute ENAC-mediatets in oociti che esprimono ENaC. (AD) Gli oociti esprimenti dell'ENaC umano sono state incubate per 30 min in soluzione priva di proteasi (controllo) o in soluzione contenente plasmina (10 mcg / ml). Per determinare ΔI ami prima (-) e dopo (+) incubazione, oociti sono stati fissati ad un potenziale di -60 mV (A, B) Quattro rappresentativi cellule intere tracce corrente da un lotto di ovociti.. Amiloride (AMI) era presente nella soluzione del bagno di inibire specificamente dell'ENaC come indicato dalle barre nere. (C) punti dati ottenuti da un singolo ovocita sono collegati da una linea. (D) Sintesi di esperimenti simili, come mostrato in C. colonne rappresentano relativo effetto stimolante sulla ΔI ami calcolato come rapporto tra ΔI ami misurata dopo una incubazione di 30 min (ΔI ami 30 min) a quella iniziale ΔI ami (ΔI ami iniziale) misurata prima dell'incubazione. Numeri nelle colonne indicano lanumero di singoli ovociti misurati. N indica il numero di diversi lotti di oociti. (Questa cifra è stata modificata da [Haerteis et al 2012 J Gen Physiol 140, 375-389, doi:. 10.1085/jgp.201110763])

Figura 3. Modello dei γENaC subunità mostrando siti di taglio per l'attivazione proteolitica e il sito di legame dell'anticorpo utilizzato. Clivaggio proteolitico della convertasi furina Golgi-associata è importante per dell'ENaC maturazione della via biosintetica prima che il canale raggiunge la membrana plasmatica. Dopo clivaggio da Furin un frammento di 76 kDa può essere rilevata sulla superficie cellulare utilizzando un approccio biotinylation e un anticorpo contro un epitopo nel C-terminale della subunità γ. L'ultimo passo fondamentale nel proteolitici ENattivazione aC probabilmente prende posto a livello della membrana plasmatica dove γENaC viene scissa da proteasi extracellulari (es. plasmina o chimotripsina) in una regione distale al sito furin risultante in un frammento di clivaggio 67 kDa. (Questa cifra è stata modificata da [Haerteis et al 2012 J Gen Physiol 140, 375-389, doi:. 10.1085/jgp.201110763])

Figura 4:. Mutazione sia la plasmina (K189) e il sito di clivaggio prostasina (RKRK178) ritarda l'attivazione di correnti ENaC-mediata e l'aspetto di un prodotto di scissione 67 kDa di γ-subunità del canale oociti esprimere WT (simboli aperti) e γ RKRK178AAAA; canale mutante K189A dell'ENaC (simboli chiusi) sono state incubate per 30 min in soluzione priva di proteasi (controllo) o per 5, 30, o 60 min in una soluzione contenente chimotripsina (2 mg / ml). (A) Per determinare ΔI ami prima e dopo incubazione, gli ovociti sono stati fissata ad un potenziale di -60 mV. Cerchi rappresentano il rapporto di ΔI ami misurata dopo 5, 30, o 60 min di incubazione (ΔI ami min) a quella iniziale ΔI ami (ΔI ami iniziale) misurata prima dell'incubazione. Ogni punto rappresenta la media ΔI ami misurata in 22-24 oociti individuali di quattro gruppi differenti. (BD) Parallelamente alla rilevazione di ΔI ami, espressione di γENaC biotinilato alla superficie cellulare è stata analizzata mediante SDS-PAGE. γENaC stato rilevato con un anticorpo contro un epitopo nel C-terminale della γENaC umana. Sono mostrati western blot rappresentativi di un lotto di ovociti. (CE) analisi densitometrica dei tre western blot simili a quelli mostrati in B o D. Per ogni corsia, le signals rilevati nelle regioni di 76 kD (colonne aperte) e 67 kD (colonne grigie) sono stati determinati e normalizzato alla somma totale del segnale rilevato. N indica il numero di diversi lotti di oociti. Clicca qui per vedere l'immagine ingrandita.
{kind=link}
Discussione
In questo manoscritto un approccio metodologico che è stato applicato con successo per studiare i meccanismi alla base l'attivazione dell'Enac da proteasi è descritto 8,13. Il laevis sistema di espressione consolidata Xenopus ovocita è stato usato per funzionalmente espresso ENaC. Funzione ENaC è stata valutata con i due elettrodi tecnica voltage-clamp convenzionale. Mutagenesi sito-diretta è stato impiegato per identificare funzionalmente pertinenti siti di clivaggio della proteasi. Biotinylation esperimenti effettuati in parallelo con le misure elettrofisiologiche hanno permesso di correlare l'aspetto di dell'ENaC clivaggio prodotti sulla superficie cellulare con l'attivazione proteolitica corrente. Una correlazione tra l'andamento nel tempo di attivazione corrente e la comparsa di frammenti clivaggio proteolitico alla superficie cellulare supporta il concetto di attivazione del canale proteolitica.
Due elettrodi di voltage-clamp richiedono l'impalamento di un oocyte con due microelettrodi. Questa procedura viene di solito eseguita solo una volta in un individuo ovocita. Tuttavia, era possibile eliminare i microelettrodi dopo una registrazione corrente whole-cell iniziale senza danno apparente al ovocita. Infatti, la membrana plasmatica nei siti di impalements sembra richiudere in pochi minuti. Così, dopo aver completato una prima misura di tensione-clamp a due elettrodi, è possibile trasferire l'ovocita dalla camera di flusso sperimentale della configurazione voltage-clamp a due elettrodi in una provetta da microcentrifuga o un pozzetto di una piastra da 96 pozzetti riempiti con un piccolo volume di soluzione di prova o di controllo. Successivamente, lo stesso ovocita può essere trasferito di nuovo alla camera di flusso e può essere nuovamente impalata per eseguire una seconda misurazione di tensione-clamp a due elettrodi. Sorprendentemente, le correnti ENaC-mediate non variano molto tra la prima e la seconda misurazione quando l'ovocita è stato mantenuto in soluzione di controllo. Al contrario, l'incubazione del ovocita in una proteasi contenente solution dopo la prima misurazione comportato un aumento corrente ENaC-mediata nella seconda misurazione (Figura 2). Questa scoperta indica l'attivazione del canale proteolitica.
Esecuzione di due misure di corrente separati in un unico ovocita offre il vantaggio che l'ovocita può essere esposta a proteasi o altri agenti farmacologici tra le due misurazioni per un tempo variabile in un piccolo volume di soluzione di prova. Questo è importante quando si usano agenti che sono costosi e / o non disponibile in grandi quantità, per esempio proteasi preparazioni purificate. La limitata disponibilità di farmaci può rendere impossibile (o sono troppo costosi) usarli in continuo due elettrodi voltage-clamp a causa dei grandi volumi di soluzione da esaminare per superfusing continuamente gli ovociti con portate di diversi millilitri al minuto. Inoltre, continui a due elettrodi misurazioni di tensione-clamp sono limitate dal fenomeno notoenon di fatiscente canale spontanea descritti anche per ENaC 15. Al contrario, esponendo gli ovociti di testare soluzioni tra due misurazioni distinte per un massimo di un'ora o più in genere non rappresenta un problema (vedi Figura 4A). Infine, due misure sequenziali eseguite nello stesso ovocita permettono osservazioni accoppiate degli effetti del farmaco. Questo ha un vantaggio rispetto misurazioni spaiati da due distinti gruppi di ovociti (proteasi trattati e trattati con veicolo), perché riduce il problema della elevata variabilità tra ovociti, generalmente osservati in canali ionici. Con osservazioni accoppiati e la possibilità di normalizzare i dati per la prima misura, meno ovociti sono necessari per gruppo sperimentale di dimostrare un effetto significativo di un agente farmacologico. Normalizzazione dei dati rende anche più facile da riassumere i dati provenienti da diversi lotti di ovociti con diversi livelli di espressione del canale ionico e, quindi, diverse correnti di base (Figura 2D). Ovviamente, esperimenti di controllo sono necessari per questo approccio per dimostrare che l'attività del canale ionico di interesse rimane stabile in oociti di controllo trattati con veicolo dalla prima alla seconda misura (vedi figura 2).
Per dimostrare che l'attivazione proteolitica corrente correla con la comparsa di dell'ENaC clivaggio prodotti sulla superficie cellulare, un approccio biotinilazione originariamente descritto da Harris et al. 9 può essere utilizzato. Questa procedura (come descritto nella sezione del protocollo e mostrato in Figura 4) è stato adattato per dimostrare che l'esposizione dei canali di proteasi e successiva attivazione di correnti dell'ENaC-mediate è accompagnato dalla comparsa dipendente dal tempo di frammenti di scissione. Il metodo biotinylation permette anche l'analisi di un aumento o una diminuzione complessiva di proteine di membrana sulla superficie cellulare. Pertanto, questo metodo è adatto per studiare l'effetto di proteasi e altri pharAgenti same momento dell'inserimento canale nella membrana plasmatica o su recupero canale. Inoltre, l'analisi Western blot delle proteine della membrana plasmatica biotinilati permette di individuare i frammenti proteici (ad esempio frammenti proteolitici dell'ENaC) o cambiamenti nel modello di glicosilazione che può essere funzionalmente rilevante.
In conclusione, la combinazione dei metodi usati per studiare l'effetto stimolatorio di proteasi sulle correnti di cellule intere ENaC-mediate e dimostrare una correlazione con il verificarsi di dell'ENaC clivaggio prodotti sulla superficie cellulare può essere utile per una vasta gamma di applicazioni. In particolare, questi metodi possono essere adatti ad affrontare questioni analoghe per quanto riguarda la regolamentazione di altri canali ionici, trasportatori o recettori transmembrana (ad esempio proteasi-activated recettori PARs).
Divulgazioni
The authors have nothing to disclose.
Riconoscimenti
The expert technical assistance of Céline Grüninger, Christina Lang, Sonja Mayer, and Ralf Rinke is gratefully acknowledged. We thank Dr. Morag K. Mansley for carefully reading the manuscript. This project was supported by a grant of the Deutsche Forschungsgemeinschaft (Grant SFB 423: Kidney Injury: Pathogenesis and Regenerative Mechanisms, to C. Korbmacher), grants of the Interdisziplinäres Zentrum für Klinische Forschung (to S. Haerteis and M. Krappitz), the ELAN program (to S. Haerteis) of the Friedrich-Alexander-Universität Erlangen-Nürnberg, and the University Library of Erlangen-Nürnberg.
Materiali
| Name | Company | Catalog Number | Comments |
| Bath Clamp Headstage for OC-725C-V | Warner Instrument Corporation | - | |
| Cold light source - Schott KL 1500 LCD | Schott | #SCOC150200EU | brightness 4; mechanical aperture: D; color temperature: 3000 K |
| E Series Electrode Holder (Str, Vent, Ag Wire, 1.2 mm) | ADinstruments | #ESW-F10v | |
| left micromanipulator; MM-33L | Warner Instrument Corporation | #64-0055 | |
| LIH 1600 - computer interface | HEKA | - | |
| magnetic valve system (ALA BPS-8) in combination with a TIB14 interface (HEKA) | ALA Scientific Instruments, HEKA | - | |
| OC-725C amplifier for two-electrode voltage-clamp recordings | Warner Instrument Corporation | - | |
| P-97 FLAMING/BROWN Micropipette Puller | Sutter Instruments | - | heat=550; velocity=22; time=200 |
| right micromanipulator; MM-33R | Warner Instrument Corporation | #64-0056 | |
| Series Electrode Holder (45°, Vent, Handle, Ag Wire, 1.2 mm) | ADinstruments | #E45w-f10vh | |
| STAT 2 IV Gravity Flow Controller | Conmed | #P-S2V-60 | |
| vacuum generator ejector SEG - for suction to remove bath solution | Schmalz | - | |
| Material | |||
| INFUJECT 60ml pump syringes for solutions | Braun | #22050 | |
| Injekt-F for lysing the oocytes | Braun | #9166033V | |
| standard wall borosilicate tubing with filament | Sutter Instruments | #BF150-86-10 | outside diameter: 1.50 mm; inside diameter: 0.86 mm; length: 10 cm |
| Reagent | |||
| complete, Mini, EDTA-free protease inhibitor cocktail tablets | Roche Applied Science | #11836170001 | |
| EZ-Link Sulfo-NHS-Biotin | Thermo Scientific | #21217 | |
| Horseradish peroxidase-labeled secondary goat anti-rabbit antibody | Santa Cruz Biotechnology | #sc-2004 | |
| NeutrAvidin Agarose | Thermo Scientific | #29200 | Neutravidin-labeled agarose beads |
| NP40 (Nonidet P-40) | Sigma-Aldrich | #I8896 | |
| Roti-Load 1 (2× SDS-PAGE sample buffer) | Carl Roth | #K929.2 | |
| SuperSignal West Femto Chemiluminescent Substrate for detection of chemiluminescent signals | Thermo Scientific | #34095 | |
| Triton-X-100 | Sigma-Aldrich | #T8787 | |
Riferimenti
- Kleyman, T. R., Carattino, M. D., Hughey, R. P. ENaC at the cutting edge: regulation of epithelial sodium channels by proteases. The Journal of Biological Chemistry. 284, 20447-20451 (2009).
- Rossier, B. C., Stutts, M. J. Activation of the epithelial sodium channel (ENaC) by serine proteases. Annu Rev Physiol. 71, 361-379 (2009).
- Poirot, O., Vukicevic, M., Boesch, A., Kellenberger, S. Selective regulation of acid-sensing ion channel 1 by serine proteases. The Journal of Biological Chemistry. 279, 38448-38457 (2004).
- Vukicevic, M., Weder, G., Boillat, A., Boesch, A., Kellenberger, S. Trypsin cleaves acid-sensing ion channel 1a in a domain that is critical for channel gating. The Journal of Biological Chemistry. 281, 714-722 (2006).
- Clark, E. B., Jovov, B., Rooj, A. K., Fuller, C. M., Benos, D. J. Proteolytic cleavage of human acid-sensing ion channel 1 by the serine protease matriptase. The Journal of Biological Chemistry. 285, 27130-27143 (2010).
- Ossovskaya, V. S., Bunnett, N. W. Protease-activated receptors: contribution to physiology and disease. Physiological reviews. 84, 579-621 (2004).
- Garcia-Caballero, A., et al. Activation of the epithelial sodium channel by the metalloprotease meprin β-subunit. Channels (Austin. 5, 14-22 (2011).
- Haerteis, S., et al. Proteolytic activation of the epithelial sodium channel (ENaC) by the cysteine protease cathepsin-S. Pflügers Archiv: European Journal of Physiology. 464, 353-365 (2012).
- Harris, M., Firsov, D., Vuagniaux, G., Stutts, M. J., Rossier, B. C. A novel neutrophil elastase inhibitor prevents elastase activation and surface cleavage of the epithelial sodium channel expressed in Xenopus laevis oocytes. The Journal of Biological Chemistry. 282, 58-64 (2007).
- Passero, C. J., Mueller, G. M., Rondon-Berrios, H., Tofovic, S. P., Hughey, R. P., Kleyman, T. R. Plasmin activates epithelial Na+ channels by cleaving the γ-subunit. The Journal of Biological Chemistry. 283, 36586-36591 (2008).
- Svenningsen, P., et al. Plasmin in nephrotic urine activates the epithelial sodium channel. Journal of the American Society of Nephrology : JASN. 20, 299-310 (2009).
- Patel, A. B., Chao, J., Palmer, L. G. Tissue kallikrein activation of the epithelial Na channel. American Journal of Physiology. Renal Physiology. 303, (2012).
- Haerteis, S., Krappitz, M., Diakov, A., Krappitz, A., Rauh, R., Korbmacher, C. Plasmin and chymotrypsin have distinct preferences for channel activating cleavage sites in the γ-subunit of the human epithelial sodium channel. The Journal of General Physiology. 140, 375-389 (2012).
- Chraibi, A., Vallet, V., Firsov, D., Hess, S. K., Horisberger, J. D. Protease modulation of the activity of the epithelial sodium channel expressed in Xenopus oocytes. The Journal of General Physiology. 111, 127-138 (1998).
- Volk, T., Konstas, A. A., Bassalay, P., Ehmke, H., Korbmacher, C. Extracellular Na+ removal attenuates rundown of the epithelial Na+-channel (ENaC) by reducing the rate of channel retrieval. Pflügers Archiv: European Journal of Physiology. 447, 884-894 (2004).
Ristampe e Autorizzazioni
Richiedi autorizzazione per utilizzare il testo o le figure di questo articolo JoVE
Richiedi AutorizzazioneThis article has been published
Video Coming Soon
Personale delle biblioteche
Copyright © 2025 MyJoVE Corporation. Tutti i diritti riservati