
Method Article
Generación de monómero Exon1 huntingtina nativo, sin etiquetar y fibrillas utilizando una estrategia de fusión de SUMO
En este artículo
Resumen
Aquí, presentamos un protocolo robusto y optimizado para la producción de cantidades de miligramos de monómeros nativos, libre de etiqueta y las fibrillas de la exon1 de la proteína huntingtina (Httex1) basado en la fusión transitoria de ubiquitina pequeños modificadores relacionados (SUMO).
Resumen
Enfermedad de Huntington (EH) es una enfermedad hereditaria neurodegenerativa fatal causada por una expansión CAG (≥36) en el primer exón del gen HD, dando como resultado la expresión de la proteína huntingtina (Htt) o N-terminal sus fragmentos con una ampliada del polyglutamine ( estiramiento de la poliQ).
Exon1 de la proteína huntingtina (Httex1) es el más pequeño fragmento de Htt que recoge muchas de las características de HD en celulares y modelos de animales y es uno de los fragmentos más ampliamente estudiados de Htt. El tamaño pequeño del Httex1 hace experimentalmente más favorable a la caracterización biofísica, utilizando técnicas estándar y de alta resolución en comparación con fragmentos más largos o Htt de larga duración. Sin embargo, la propensión de la alta agregación de mutante Httex1 (mHttex1) con contenido mayor poliQ (≥42) ha hecho más difícil desarrollar expresión eficiente y sistemas de purificación para producir estas proteínas en cantidades suficientes y hacerlos accesibles a científicos de diferentes disciplinas sin el uso de otras estrategias que alteran la secuencia nativa de la proteína o proteínas de fusión. Aquí presentamos un método robusto y optimizado para la producción de cantidades del miligramo de nativo, etiqueta libre de Httex1 basado en la fusión transitoria de ubiquitina pequeños modificadores relacionados (SUMO). La simplicidad y la eficacia de la estrategia eliminará la necesidad de utilizar secuencias de no nativos de Httex1, así que esta proteína más accesible a los investigadores y mejorar la reproducibilidad de los experimentos en diferentes laboratorios. Creemos que estos avances también facilitará futuros estudios dirigidos a dilucidar la relación estructura-función de Htt, así como en desarrollo nuevas herramientas de diagnóstico y terapias para tratar o retardar la progresión de la EH.
Introducción
HTT es una proteína del 348 kDa y se ha implicado en varias funciones fisiológicas1. Cuando Htt contiene una región de poliQ ampliado de más de 36 residuos en su N-terminal, causa HD2,3. Patología de la EH se caracteriza por las inclusiones celulares en el cuerpo estriado y la corteza, que conduce a la muerte neuronal y atrofia de los tejidos afectados4,5. Varios fragmentos N-terminal Htt que contienen las vías repetición poliQ han sido detectados en cerebros post mortem de pacientes con EH y se piensan para ser generadas por el proceso proteolítico de la proteína huntingtina del6. Estudios recientes sugieren que Httex1 también podría ser formado debido a empalmar de mRNA aberrante. Httex1 contiene la mutación patológica poliQ y su sobreexpresión en los animales puede recapitular muchas de las características claves de HD7, resaltando así un posible papel central de este fragmento en HD patología y enfermedad progresión6, 8,9.
Debido a la agregación alta propensión del mutante Httex1 (mHttex1) en vías de expansión poliQ, la mayoría de los sistemas de expresión existentes se basa en la fusión transitoria de Httex1 a las proteínas (como la glutatión-S-transferasa (GST), tiorredoxina (TRX) o proteína de unión a maltosa (MBP) o péptidos (poly-histidina) que diferencialmente mejoran su solubilidad, estabilidad, purificación o expresiones10,11,12,13,14 ,15,16,17,18,19,20,21,22,23 ,24,25,26,27,28. El socio de fusión es Httex1 con una corta secuencia que contiene un sitio de la hendidura para proteasas como tripsina, tabaco etch virus (TEV) proteasa o PreScission para el escote y la liberación de Httex1 antes de la iniciación de la agregación o purificación. Limitaciones de estos métodos incluyen la posibilidad de dejar residuos adicionales debido a la hendidura no sin dejar huellas y la creación de fragmentos truncados debido a la miscleavage dentro de la secuencia de Httex1, además de heterogeneidad debido a la hendidura incompleta ( Ver Vieweg et al. , para una discusión más detallada sobre las ventajas y limitaciones de este enfoque)10. Para hacer frente a estas limitaciones, hemos desarrollado recientemente una estrategia de expresión que permite la generación de Httex1 libre de etiqueta nativo por primera vez utilizando una fusión transitoria del N-terminal de Synechocystis SP. (Ssp) DnaB intein a Httex110. Mientras que el escote intein es traceless y específica y produce la cantidad de mg de proteínas, aún sufre dos inconvenientes que podrían reducir el rendimiento: es decir, prematura ruptura de la inteína que puede ocurrir durante la expresión y el hecho de que la hendidura ocurre en varias horas, que podrían conducir a la pérdida de proteínas debido a la agregación, especialmente para Httex1 con repeticiones de expansión poliQ.
Para enfrentar estas limitaciones y para afinar nuestra estrategia para la producción de nativo, Httex1 libre de etiqueta, hemos desarrollado un nuevo sistema de expresión basado en la fusión transitoria de SUMO, más exactamente el homólogo de levadura Smt3 a Httex1. La aplicación del sistema para la producción de proteínas recombinantes SUMO fue publicado por primera vez en 200429, donde se demostraron una mayor tasa de expresión y solubilidad de la proteína de la fusión de SUMO. La etiqueta SUMO puede ser dividida por la ubiquitina como proteasa específica de proteína 1 (ULP1), que no requiere un sitio de reconocimiento, pero reconoce la estructura terciaria de SUMO y prácticamente elimina la posibilidad de miscleavage30. Por otra parte, el clivaje mediado por ULP1 es rápido y sin dejar huellas y no deja residuos adicionales. La ruptura prematura de la etiqueta de fusión, como se observa con el10del intein autocatalíticas, es evitar totalmente por el requisito de una proteasa externo. Mientras que la estrategia SUMO es hoy en día ampliamente utilizada para la producción de proteína recombinante31,32,33, demostramos en este trabajo que es especialmente útil para la generación de un intrínsecamente desordenadas, proteína de amyloidogenic proclive a la agregación, como Httex1. Creemos que la simplicidad, eficiencia y robustez de nuestro método basado en la fusión de SUMO hacen más accesible a investigadores de diferentes disciplinas Httex1 nativa, etiqueta-libre y eliminar la necesidad de utilizar secuencias de no nativos de Httex1 en vitro . Este es un importante avance que facilitará futuros estudios para aclarar la relación estructura-función de Httex1.
El protocolo describe la purificación de Httex1 de 12 L de cultivo bacteriano, pero el protocolo podría adaptarse fácilmente para producciones de menor o mayor escala. El protocolo describe la producción de tipo salvaje Httex1 (wtHttex1) con una longitud de la repetición poliQ debajo (23Q) y mutante Httex1 (mHttex1) con un poliQ repetir longitud arriba (43Q) el umbral patógeno (36Q).
Protocolo
1. expresión de Httex1 recombinante 23Q y 43Q
- Preparar las soluciones y buffers requeridos. Preparar 1000 x solución ampicilina (AMP, 100 mg/mL), filtro (0.2 μm), alícuota y almacenar a-20 ° C. Preparación de medio de caldo (LB) Lisogenia (25 g LB Miller por 1 L H2O), autoclave. Preparar solución stock de 1 M isopropílico ß-D-1thiogalactopyranoside (IPTG), (0,2 μm), alícuota y tienda a-20 ° C.
- Transformación química competente e. coli B ER 2566 con un vector de pTWIN1, que contiene Httex1 humanos fusionados con una etiqueta de His6-SUMO del N-terminal con el método de choque de calor34.
Nota: La cepa de e. coli BL21 DE3 también se ha utilizado. Sin embargo, en este caso se observó un aumento en la cantidad de truncamientos. - Inocular con 1 200 mL de medio LB x AMP en un erlenmeyer de 1 litro mediante la adición de una sola Colonia de la placa de agar con una pipeta estéril. Incubar el cultivo a 30 º C y 180 rpm durante 20 h (noche) en una incubadora de bacteria.
- Tomar una muestra de la cultura con una pipeta estéril 1 mL. Medir la densidad óptica a 600 nm (OD600) de la muestra con una cubeta de plástico desechable y un fotómetro (respeto la medida comprendida entre 0.1 y 1, diluir con medio LB si es necesario). Calcular la cantidad de planas que dan lugar a una partida de OD600 de 0.05 en una cultura de 3 L (con un planas de OD600 = 3 que significa 50 mL).
- Inocular cuatro culturas (cada 3 L de medio LB con 1 x AMP en un matraz de 5 L), añadiendo la cantidad calculada de planas con una pipeta estéril. Incubar los cultivos a 37 ° C y 180 rpm en una incubadora de bacteria.
- Cada 30 minutos, tomar una muestra de la cultura con una pipeta estéril 1 mL. Medir el600 del OD de la muestra con una cubeta de plástico desechable y un fotómetro. Cuando ha llegado a OD600 0.1 (típicamente después de 1-2 h), ajuste la temperatura de la incubadora bacteriana a 14 ° C y continuar la incubación mientras se enfría. Cada 30 minutos, tomar una muestra de la cultura con una pipeta estéril 1 mL. Medir el600 del OD de la muestra con una cubeta de plástico desechable y un fotómetro.
Nota: El tiempo para enfriar las culturas puede variar con la incubadora usada, por lo que el momento de comenzar el enfriamiento tendría que ser adaptado según el tipo de incubadora usado. Sin embargo, cambiar el gradiente de temperatura sólo debe tener un pequeño impacto en el rendimiento como la proteína de la fusión de SUMO parece ser bastante estable. - Cuando OD600 llega a 0.3-0.4 (normalmente después de 1-2 h), tome una muestra de la inducción de la cultura para el análisis de SDS-PAGE de la sobreexpresión. Calcular el tamaño de la muestra que da una cantidad similar de células y una buena señal de Coomassie manchado SDS-PAGE: gel bien para un 10: volumen = 0,2 mL/OD600; tomar la mitad para un gel bien 15.
- Para un cultivo bacteriano con un OD600= 0.4, tomar 500 μl. Tomar el volumen calculado de cultivo bacteriano con una pipeta estéril. Desactivación de la muestra (18000 x g, 4 º C, 2 min.) y descarte el sobrenadante. Mantener el diábolo a-20 ° C hasta que esté listo para usar para el análisis (paso 1.11).
- Inducir la expresión de la proteína por pipeteo 1,2 mL de una solución madre del IPTG de 1 M a cada solución de cultura 3 L (concentración final 0.4 mM). Continúe incubando la cultura a 14 ° C durante 16 h (noche).
Nota: La temperatura por lo general habrá alcanzado ~ 20 ° C por el momento que se agrega el IPTG, dependiendo el desempeño de la incubadora. - Tome una muestra de la inducción de la cultura para el análisis de SDS-PAGE de la sobreexpresión, siguiendo el procedimiento descrito en el paso 1.7.
- Recoger las células por centrifugación en tubos de 1 L (3993 x g, 4 ° C, 10 minutos). Eliminar el sobrenadante, mantener el pellet celular en hielo y proceder directamente a la purificación.
- Análisis de la sobreexpresión por SDS-PAGE35,36. Resuspender las muestras previas y post inducción en 20 μl de corriente buffer y 20 μl de 2 x carga del tinte. Calentar las muestras por 5 min a 95 ° C en un bloque de calor y carga 20 μl en un gel 15% mientras esté todavía caliente. Correr el gel durante 90 minutos a 180 V. tinción del gel con Coomassie tinte siguiendo las instrucciones del fabricante. Comparar los resultados con resultados representativos en la figura 1C.
Nota: El protocolo se puede detener aquí, el precipitado de células puede ser congelado y almacenado a-80 ° C durante varias semanas. Para obtener resultados óptimos, se recomienda utilizar el pellet bacteriano fresco y evitar la congelación. Congelación y descongelación puede producir lisis de las células y la degradación de Httex1. Esto podría reducir el rendimiento y la calidad de la proteína.
2. célula de la lisis y la purificación de su6-proteína de fusión Httex1 SUMO inmovilizados por cromatografía de afinidad metálica (IMAC)
- Preparar 2 L de tampón A en un frasco de vidrio (50 mM Tris (hidroximetil)-aminomethan (Tris), 500 mM NaCl, imidazol de 15 mM). Preparar 1 L de tampón B (50 mM Tris, 500 mM NaCl, pH de 500 mM imidazol 7,4) en una botella de vidrio. Después de disolver las sales, ajustar el pH con HCl N 10 y filtrar las soluciones en frascos frescos con un filtro de botella (0.65 μm). Preparar un 1000 x phenylmethylsulfonyl fluoruro (PMSF, 0,3 M) stock solución, alícuota de 100 μl y almacenar a-20 ° C.
Nota: El protocolo está diseñado para permitir completar todos los pasos de lisis de los pellets bacterianos a purificación cromatografía líquida (HPLC-RP) de alto rendimiento de fase inversa y liofilización en 8-9 h. Para limitar la agregación y la proteólisis, es aconsejable trabajar rápidamente sin detenerse y realizar todos los pasos a 4 ° C o en hielo. - Añadir 100 μl de solución stock de PMSF y cinco tabletas (1 por cada 30 mL de volumen final) de inhibidor de la proteasa a 100 mL de buffer previamente enfriada A. agregar el sedimento bacteriano en el búfer y homogeneizar la suspensión por agitación con una barra de agitación magnética y mediante pipeteo arriba y abajo de wi TH una pipeta estéril de 10 mL (~ 30 min).
- La suspensión de las bacterias se dividen en alícuotas de 40 mL en tubos de plástico desechables de 50 mL. Someter a ultrasonidos cada alícuota en un lote de agua/hielo de lisis celular (70% de amplitud, tiempo de sonicación total 5 minutos, intervalos de sonicación de 30 s, pausa s 30).
Nota: Es importante que la muestra no calienta para arriba durante el paso de sonicación. Se recomienda añadir algo de agua para el baño de hielo para mejorar la disipación de calor durante la sonicación. El procedimiento de sonicación tendría que adaptarse si se utiliza un instrumento diferente. Otros métodos de lisis una prensa francesa como un microfluidizer deberían funcionar así y podrían ser beneficiosos para evitar el calentamiento de la agregación de la muestra y de la proteína. Estos dispositivos no estaban disponibles en nuestro laboratorio y obtuvimos buenos resultados con nuestro protocolo de sonicación. - Tome una muestra de 50 μl del lisado p.a. sodio dodecil sulfato poliacrilamida gel electroforesis (SDS-PAGE). Centrifugue la muestra (18000 x g, 4 º C, 2 min.) y pipetear la fracción soluble en un nuevo tubo de ensayo. Resuspender la fracción insoluble en 50 μl de tampón A con una pipeta. Mantener las muestras en hielo hasta análisis de SDS-PAGE (paso 2.6).
- Aclarar el lisado por centrifugación (39191 x g, 4 º C, 60 min).
- Durante la etapa de centrifugación, analizar el paso de lisis celular por SDS-PAGE. Añada 50 μl de 2 x carga del tinte a la fracción soluble e insoluble del lisado respectivamente. Calentar por 5 min a 95 ° C y carga 2 μL en un gel 15% mientras esté todavía caliente. Correr el gel durante 90 minutos a 180 V. tinción del gel con Coomassie tinte siguiendo las instrucciones del fabricante. Comparar los resultados con resultados representativos en la figura 1C.
- Filtrar el sobrenadante (0,45 μm, filtros de la jeringuilla). Tome una muestra de 20 μl del sobrenadante filtrado para el análisis de SDS-PAGE (paso 2.11).
Nota: Por lo general, se obtiene un volumen de 90 a 100 mL de sobrenadante clarificado y filtrado. Por lo general, son suficientes 3 filtros de la jeringuilla. Si la filtración es engorrosa, intente aumentar la velocidad de centrifugación y tiempo. - Aislar la proteína de la fusión de His6-SUMO Httex1 de la bacteriana clarificada lisado mediante cromatografía de afinidad metálica inmovilizados (IMAC) en un sistema de cromatografía líquida (FPLC) rendimiento rápido a 4 º C37.
- Llenar el clarificado lisado superloop y carga en la columna de Ni-NTA (despojado, limpiar y volver a cargar según el manual del fabricante, se recomienda un espacio en blanco anterior ejecutar) en volúmenes de columna de 2 mL/min 10 pasar (CV, 200 mL) de tampón A 10 mL/min para lavar a las proteínas no Unidas.
- Eluir la proteína de fusión con 2.5 CV (50 mL) del 100% de tampón B en 2 mL/min usar un tamaño de fracción de 50 mL para la carga y de lavado y 5 mL para la elución. Comparar los resultados con resultados representativos enD, figura 1.
- Tomar una muestra de cada fracción para el análisis de SDS-PAGE (20 μl) y las fracciones que contiene la proteína de fusión según el pico del cromatograma IMAC de la piscina. Añadir (2S, 3S) - 1,4 - Bis (sulfanyl) butano-2, 3-diol (DTT) y L-cisteína (concentración final de 100 m m) como polvo y disolver suavemente invirtiendo el tubo.
Nota: En nuestra experiencia la pureza de la proteína de la fusión de las diferentes fracciones es comparable. Como medida de precaución, las fracciones de la proteína de fusión purificada deben combinarse rápidamente después de IMAC para prevenir la agregación de las fracciones altamente concentradas. Además, se recomienda a proceder directamente a la ruptura de la etiqueta SUMO y purificación de HPLC. Si es necesario, el protocolo se puede detener aquí. La solución diluida de la proteína de fusión fue congelada en nitrógeno líquido, almacenada a-80 ° C y purificada después de descongelar sin una reducción significativa en el rendimiento. Almacenamiento de la solución diluida de la proteína de fusión a 4 ° C por 24 h también da resultados similares. - Analizar el IMAC por SDS-PAGE. Añadir 20 μl de colorante a cada muestra de carga. 2 μl del material crudo (2.7), la fracción no unida, la fracción de lavado y cada fracción del pico de elución en un gel de 15% de la carga. Correr el gel durante 90 minutos a 180 V. tinción del gel con Coomassie tinte siguiendo las instrucciones del fabricante. Comparar los resultados con resultados representativos enD, figura 1.
3. hendidura de su6-SUMO-etiqueta y purificación de HPLC
PRECAUCIÓN: El ácido trifluoroacético (TFA) es un líquido volátil y pueden causa quemaduras graves así que manejar con cuidado. Llevar a cabo toda la gestión en una campana de humos y usar equipo de protección personal adecuado (es decir, guantes de nitrilo desechables, gafas de seguridad y una bata de laboratorio).
- En una botella de 5 L, añadir 5 mL de TFA con una jeringa de plástico de 5 L de agua (solvente A: H2O, 0.1% (TFA). Añadir 2,5 mL de TFA con una jeringa de plástico de una botella de 2,5 L de acetonitrilo (solvente B: acetonitrilo, 0.1% TFA).
- Preparar el sistema HPLC según lo sugerido por el fabricante. Realizar una corrida en blanco para asegurar una columna limpia.
- Tome una muestra de 100 μl de la proteína de fusión antes de la adición de ULP1 para controlar la reacción de hendidura por UPLC (paso 3.5).
- Transferir 20 mL de la proteína de fusión a un nuevo tubo de 50 mL y añadir 0,4 mL de solución stock ULP1, incubar en hielo. Mantener la proteína de fusión restantes en el hielo.
Nota: El su etiquetado catalítico fragmento 403-621 de la proteasa de ubiquitina-como-específica 1 (aquí denominado "ULP1") fue utilizado para hender la etiqueta SUMO. La proteína de fusión es más estable que los Httex1 troceados. Es aconsejable no hiende la etiqueta de SUMO de toda la masa. Por el contrario, continuar con alícuotas de un tamaño que puede aplicarse directamente y completamente a la columna HPLC. - Cada 10 minutos, tomar una muestra de 100 μl de la reacción de hendidura para supervisar el progreso por ultra-cromatografía líquida (UPLC). Centrifugar las muestras (18000 rpm, 4 ° C, 2 min.) y analizar 2 μl del sobrenadante por UPLC (gradiente de 10% a 90% solvente B en el A 0.25 a 3 min, 10% B durante 1 min, consulte las instrucciones del fabricante para el uso del instrumento). Comparar los cromatogramas obtenidos para la muestra antes de la adición de ULP1 y las muestras tomadas. Comparar los resultados con resultados representativos en la figura 2B.
- Una vez que el SUMO-escote (el pico de la proteína de fusión ha desaparecido en el cromatograma UPLC y completamente se convierte en el pico que aparece recién SUMO y Httex1), filtrar la muestra con un filtro de jeringa (0,22 μm).
Nota: La separación SUMO es típicamente muy rápida (10-20 min a 4 ° C) por lo tanto el análisis UPLC con un tiempo de 4 minutos es una herramienta valiosa para controlar la reacción. Filtrado de la muestra antes de HPLC purificación es principalmente una medida preventiva para aumentar la vida útil de la columna. La muestra no debe resultar turbia. - Purificar la muestra filtrada por RP-HPLC (gradiente de 25-35% de disolvente B en A solvente, más de 40 min a 15 mL/min (0-10 minutos: 5%; 10-12.5 min: 5 a 25%, 12.5-52.5 min.: 25 a 35%, 52.5-57.5 min 35 a 95%; refiérase a las instrucciones del fabricante para el uso del instrumento). Comparar los resultados con resultados representativos enC figura 2.
Nota: Httex1 y su6-SUMO separado bien por RP-HPLC. Sin embargo, puede haber pequeñas cantidades de Httex1 truncada en el principio y el final del pico. Recogen pequeñas fracciones para obtener la máxima cantidad de material puro.
PRECAUCIÓN: Use el equipo de seguridad apropiado (es decir, bata de laboratorio, guantes aislados y un protector de cara) cuando manejo fluidos criogénicos. - Analizar las fracciones de HPLC por espectrometría de masas de ionización electro spray (ESI-MS, muestreadores, inyectar 10 μl, flujo de 0.6 mL/min, solvente: 20% B en A, ninguna columna, consulte las instrucciones del fabricante para el uso del instrumento) y UPLC (pendiente de 10% a 90% solvente B en el A 0.25 a 3 min, 10% B durante 1 min, consulte las instrucciones del fabricante para el uso del instrumento). Piscina fracciones de pureza en tubos de plástico de 50 mL, congelar en nitrógeno líquido y liofilizar. Pesar y transferir la proteína liofilizada en tubos de plástico de 2 mL y almacenar a-20 ° C.
- Caracterizar el material purificado por UPLC, ESI-MS y SDS-PAGE. Disolver 100 μg de Httex1 liofilizado en 8 μl de aseado TFA en un tubo de 1,5 mL e incubar durante 20 min a temperatura ambiente en el tubo de ensayo cerrado. Cuidadosamente se evaporan el TFA bajo una campana de humos con una corriente de nitrógeno o argón. Utilizar baja presión de nitrógeno/argón para evitar la pérdida de la muestra.
- Disolver la proteína en 100 μl de H2O. analizar 2 μl por UPLC y 5 μl de ESI-MS como en el paso 3.8. Mezcla 20 μl de solución de proteína con 20 μl de 2 x carga del tinte.
- Analizar cantidades de 1 μg a 10 μg por SDS-PAGE. Correr el gel durante 90 minutos a 180 V. tinción del gel con Coomassie tinte siguiendo las instrucciones del fabricante. Comparar los resultados con resultados representativos enD figura 2.
4. desagregación y resolubilización de proteínas Httex1
PRECAUCIÓN: TFA es un líquido volátil y pueden causa quemaduras graves así que manejar con cuidado. Llevar a cabo toda la gestión en una campana de humos y usar equipo de protección personal adecuado (es decir, guantes de nitrilo desechables, gafas de seguridad y una bata de laboratorio).
- Preparar 10 mL de solución salina amortiguada de fosfatos de Dulbecco (DPBS) (137 mM NaCl, 2,7 mM KCl, 10 mM Na2HPO4, 2 mM KH2PO4, pH 7.4) del polvo premezclado en un tubo de 50 mL. Filtrar la solución DPBS a través de un filtro de 0.2 μm antes de cada uso.
- Disolver 150 μg de Httex1 liofilizado en 12 μl de aseado TFA en un tubo de 1,5 mL e incubar durante 20 min a temperatura ambiente en el tubo de ensayo cerrado. Cuidadosamente se evaporan el TFA bajo una campana de humos con una corriente de nitrógeno o argón. Utilizar baja presión de nitrógeno/argón para evitar la pérdida de la muestra38.
Nota: en general, utilice 4 μL TFA para disolver y separar 50 μg de proteína. Este procedimiento creará una película de proteína en el interior las paredes de la probeta. Para evitar la agregación inmediata de Httex1 en los siguientes pasos, trabajar con reservas previamente enfriados, mantener la proteína siempre en el hielo y evitar altas concentraciones. - Disolver la proteína desglosada en 1 mL de DPBS previamente enfriado y ajustar el pH a 7.2-7.4 con 1 M NaOH. Filtrar la solución de la proteína a través de un filtro centrífugo de 100 kDa en tubos de plástico de 1,5 mL (20000 x g, 4 ° C, 20 minutos).
Nota: La concentración teórica calculada de Httex1 es mayor que la concentración final deseada para tener en cuenta posibles pérdidas. El paso de filtración es necesario quitar cualquier agregados que puedan haberse formado durante la disolución de la proteína. - Determinar la concentración de Httex1 utilizando una curva de calibración de UPLC basada en el análisis del aminoácido (detección en λ214) o 2 μg de la proteína y aminoácidos análisis para validar la concentración de10. Calcular la cantidad de DPBS que debe añadirse para obtener una concentración de 3 μm Httex1.
- Diluir la proteína a 3 μm por agregar la cantidad calculada de DPBS al tubo de ensayo. Mantenga el tubo en hielo hasta la iniciación del Protocolo de agregación.
Nota: Httex1 43Q no debe ser almacenado en solución. Siempre prepare una solución de proteína fresca basada en el protocolo anterior. Las proteínas Httex1 se conservan mejor en un polvo liofilizado a-20 ° C.
5. seguimiento de la cinética de agregación de Httex1 43Q utilizando UPLC espectroscopia circular del dicroísmo (CD) y caracterización de los agregados por microscopia electrónica de transmisión (TEM)
- Preparar solución de formato de uranilo para TEM como previamente divulgados39.
- Iniciar la agregación de Httex1 43Q por incubación en una solución de 3 μm en DPBS a 37 ° C (uso 1 mL de la solución, preparada como se describe en el protocolo de desagregación).
Nota: La agregación de Httex1 puede realizarse en concentraciones más altas dependiendo de las necesidades y objetivos del experimento. - Cuantificar la cantidad de proteína soluble utilizando UPLC en tiempo indicado puntos (0, 1, 2, 4, 6, 8, 12, 24, 48 y 120 h). Para ello, tomar una alícuota de 35 μl y eliminar los agregados insolubles por centrifugación (20000 x g, 4 ° C, 20 min). Inyectar 4 μL del sobrenadante en el UPLC. Calcular el porcentaje de monómero soluble basado en el cambio de la zona de pico con el instrumento software 40. Comparar los resultados con resultados representativos en la figura 3A.
- Caracterizar los cambios en la estructura secundaria usando la espectroscopia del CD a las 0 y 48 h. Tomar una alícuota de 100 μl y medir la elipticidad (cubeta de cuarzo de 1 mm, 195 nm a 250 nm, 20 ° C, datos de puntos de cada 0,2 nm, velocidad 10 nm/min, tiempo 2 de integración digital s, ancho de banda de 1.0 nm). Adquirir 6 espectros de la muestra y la media y suave utilizando un filtro de binomio con la anchura de la convolución de 99. Trazar los espectros como el residuo medio elipticidad molar (θMRE)41. Comparar los resultados con resultados representativos en la figura 3B.
- Caracterizar las propiedades estructurales y morfológicas de los agregados por TEM. Colocar 3 μl de la solución de la proteína en una Formvar /-recubiertas de carbón malla 200, resplandor-descargado rejilla de cobre durante 1 minuto lavar la rejilla dos veces con 15 μL agua, una vez 15 μL de formato de uranilo 0,7% (w/v) y la mancha de 30 s con 15 μL de formato de uranilo 0,7% w/v. Realizar un análisis TEM de las rejillas. Comparar los resultados con resultados representativos en la figura 3C.
Resultados
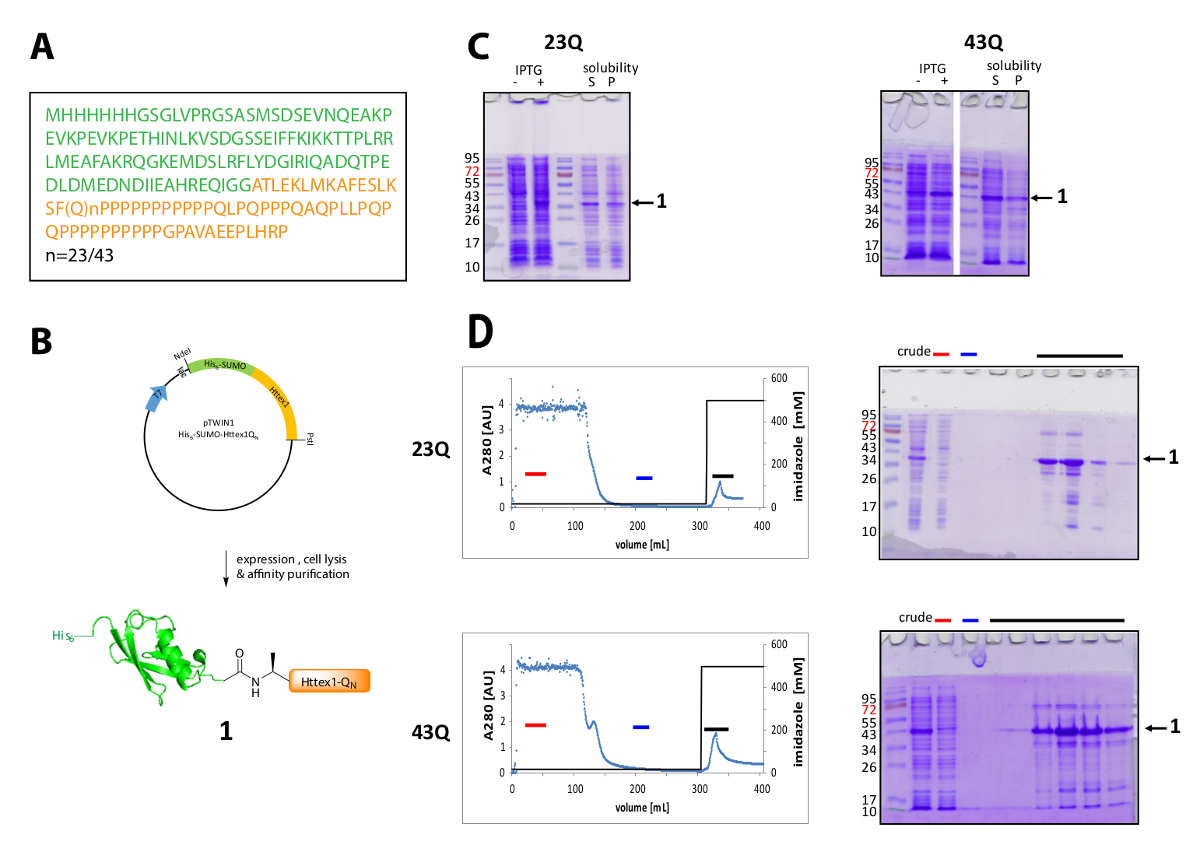
Httex1 se expresa en e. coli con un N-terminal su6-etiqueta SUMO. Los resultados representativos de la expresión y purificación de la proteína de fusión se resumen en la figura 1. La secuencia de Httex1 consiste en los residuos 2-90 de Htt y empieza por Ala2, porque Met1 es totalmente exfoliados en vivo42. La numeración de los aminoácidos se refiere a la variante 23Q, la secuencia completa de la proteína de fusión expresada se muestra en la figura 1A. La plásmidos se depositará en el Addgene en un futuro cercano para compartir con la comunidad. Se muestra un esquema del plásmido y la proteína de fusión expresado en la figura 1B. His6-SUMO Httex1 expresa en un nivel medio (figura 1C) y la mayoría de la proteína de fusión está presente en la fracción soluble después de la lisis, tanto para la variante 43Q y la 23Q. La proteína de fusión migra más de lo esperado, basado en el peso molecular. Esto es en parte debido al fuerte repliegue de SUMO pero sobre todo debido a la composición de la secuencia inusual de Httex1, que contiene principalmente residuos de glutamina y prolina. El tipo salvaje (23Q) y el mutante (43Q) proteína de fusión puede ser enriquecida a ~ 80% de pureza por IMAC (figura 1D) la presencia de co purificar proteína huésped puede explicarse por el nivel comparativamente bajo de la expresión de Httex1 y la gran muestra volumen aplicado a la columna.
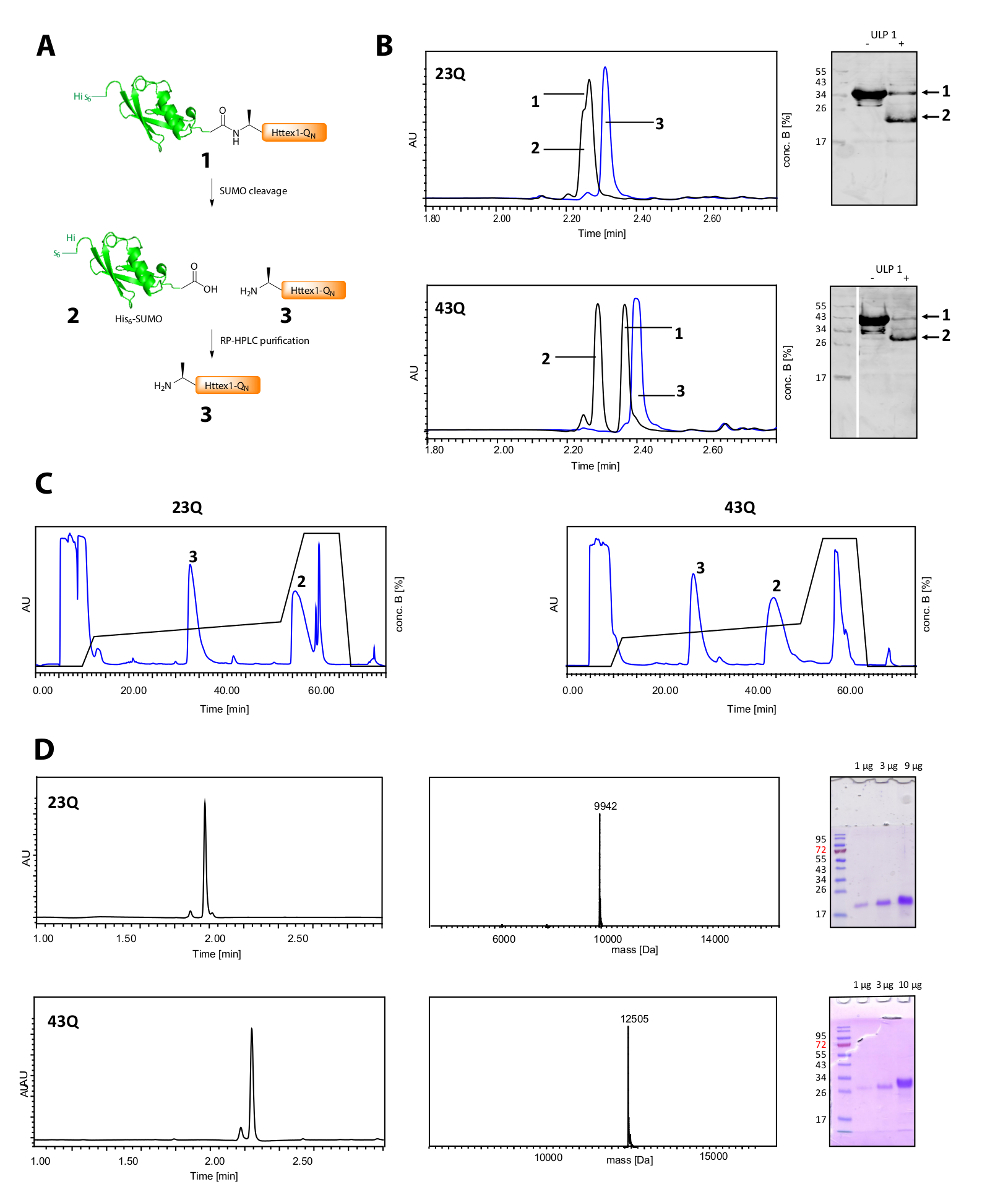
El escote de su6-etiqueta SUMO y la purificación del Httex1 se muestra en la figura 2A. UPLC es una herramienta eficaz para controlar la ruptura de su6-etiqueta SUMO (figura 2B). Se consume el pico original de la proteína de fusión y dos picos nuevos y bien separados, correspondientes a su6-etiqueta SUMO y Httex1 aparecen. La reacción de hendidura está acabada en 10-20 minutos. El Western Blot (WB) es demasiado lento para controlar eficazmente la reacción de hendidura, pero ha sido incluida en la figura de referencia y demostrar la integridad de la división SUMO. Ambos Httex1 23Q y 43Q pueden ser separados de su6-SUMO de la etiqueta por RP-HPLC (figura 2C) y fueron obtenidos en alta pureza como se muestra por el análisis de UPLC, MS y SDS-PAGE (figura 2D).
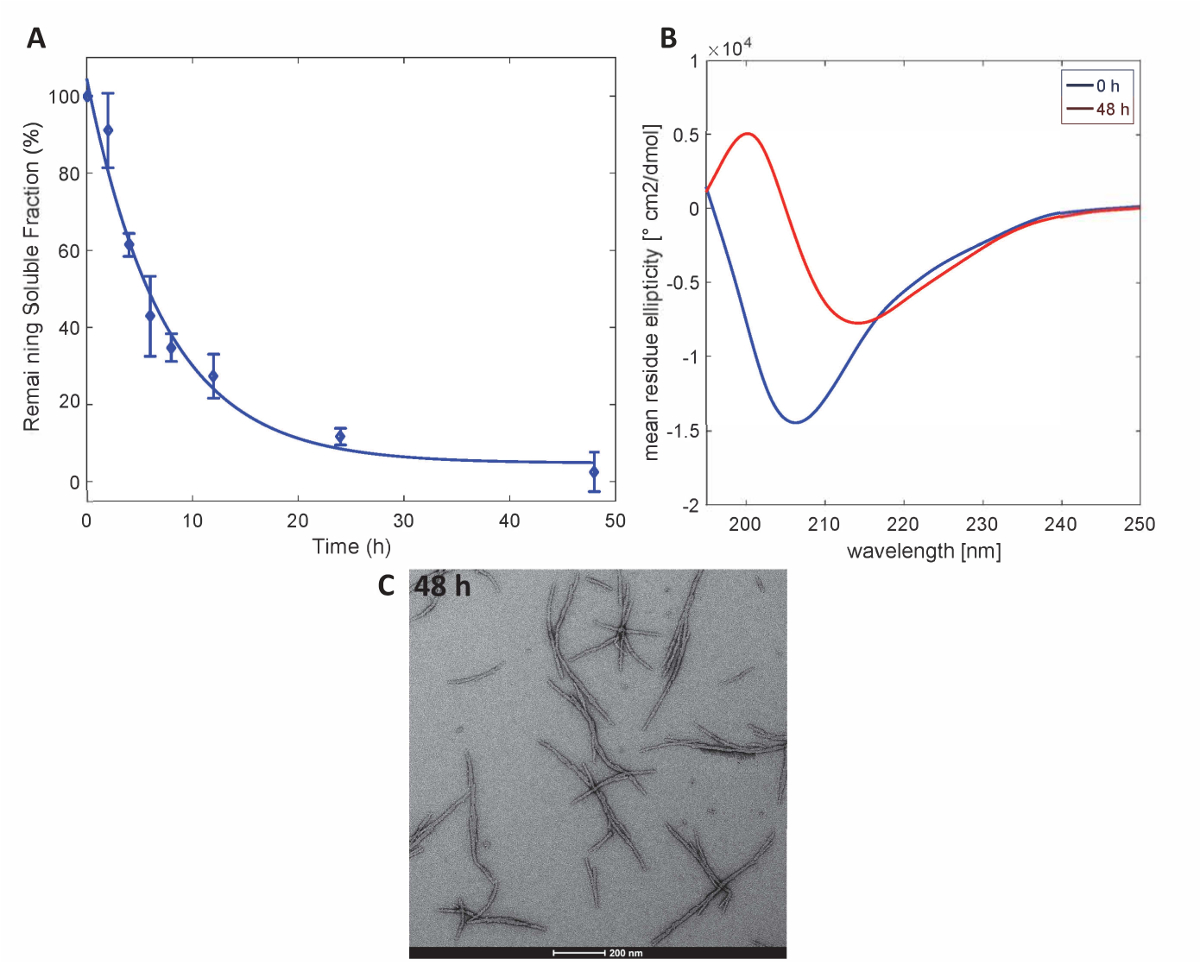
Para ilustrar que las proteínas de Httex1 preparadas por este método de conservan las propiedades de agregación esperado de Httex1, se evaluó la cinética del fibrillization de la mutante Httex1 a 37 ° C por un ensayo de sedimentación, controlar los cambios en la estructura secundaria por el CD Espectroscopia y caracteriza la morfología de los agregados por TEM. Un conjunto de datos representativo de la cinética de agregación de mHttex1 de formación de fibrilas según lo determinado por un ensayo de sedimentación se muestra en lafigura 3A. La pérdida de Httex1 soluble 43Q con el tiempo, debido a la formación de fibrilas se cuantificó por UPLC. Se observa un agotamiento total de proteína soluble después de 48 horas de incubación. Además, se determinó la estructura secundaria de la proteína por espectroscopia de CD (figura 3B). Httex1 43Q turnos de no estructurados (λmin 205 nm) a conformación rica principalmente β-hoja (λmin 215 nm) después de 48 horas de incubación. Este cambio estructural es acompañado por la formación de agregados fibrilares largos como observables por TEM en 48 horas (figura 3C).

Figura 1 . Expresión y purificación de su6-proteína de fusión del SUMO Httex1.
(A) la secuencia de aminoácidos de su6-SUMO-Httex1-QN fusión construcciones (su6-SUMO en verde y Httex1 QN en color naranja); (B) Descripción esquemática de la expresión y purificación de la proteína de fusión; (C) análisis de SDS-PAGE de la expresión y la solubilidad de la proteína de fusión después de lisis; (D) cromatograma del análisis de las fracciones por SDS-PAGE y la purificación de la IMAC de la proteína de fusión (roja de la barra: barra de fracción no unida, azul: fracción de lavado, negro de la barra: fracciones que contienen el pico de elución); Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 2 . Escote de su etiqueta SUMO6 y purificación de proteínas Q Httex1N libre de etiqueta.
(A) Descripción esquemática; (B) análisis de la ruptura de la etiqueta SUMO con ULP1 por UPLC (azul: antes de la adición de ULP1; negro: 20 min (23Q), respectivamente de 10 min (43Q) después de la adición de ULP1) y WB (MAB5492 1: 2000, secundario cabra anti anticuerpo de ratón 1: 5000); (C) cromatograma de la purificación de RP-HPLC preparativa de Httex1; D: Análisis de la Httex1 purificada por UPLC, SDS-PAGE y ESI-MS; el peso molecular esperado es Da 9943 (23Q) y Da 12506 (43Q) respectivamente. Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}

Figura 3 . Agregación de 43Q Httex1: ensayo de sedimentación (A) basa en UPLC. (B) espectro de CD de la estructura secundaria en 0 h y micrografías TEM 48 h. (C) de los agregados a las 48 h (las barras de escala son 200 nm). Haga clic aquí para ver una versión más grande de esta figura.
{kind=link}
Discusión
En este protocolo, hemos descrito un procedimiento eficaz para obtener cantidades de miligramos de nativos, untagged Httex1 que contienen residuos de glutamina 23 o 43. Esto se logró mediante la expresión Httex1 como una fusión de la terminal C a su6-etiqueta de SUMO, que se utiliza para aislar la proteína de la fusión de la célula de lisado de IMAC y troceado antes de purificación de HPLC de Httex1. Mientras que la estrategia SUMO se ha utilizado en la producción de varias otras proteínas, nuestro método muestra que las propiedades únicas de que sumo también podría ser utilizado para generar intrínsecamente desorden, propenso a la agregación, proteína amyloidogenic que previamente han demostrado ser extremadamente difícil de manejar y producir43,44. Presentamos un protocolo que es sencillo, fácil de usar y comparable a un protocolo para la generación de una proteína "bien-comportada". La fusión de SUMO solubiliza y estabiliza la Httex1 durante el paso de la purificación de IMAC y la expresión. Ruptura prematura de la etiqueta, como se observa con la agregación y intein estrategia10 eran no más un problema.
Las proteínas intrínsecamente desordenadas son especialmente vulnerables a la degradación. Mientras que la degradación del N-terminal en la región de N17 no es un problema con este protocolo, truncamientos en el PRD de Httex1 pueden ocurrir. Como las proteínas truncadas son muy similares a Httex1 de hidrofobicidad, carga y tamaño, quitar por medios cromatográficos es un reto, así es mejor prevenir su formación en primer lugar. Pegarse de cerca en el protocolo, siempre trabajando en el hielo y usando una cantidad suficiente de inhibidor de la proteasa deberían ayudar a mantener muy bajo el nivel de truncamiento observado. Aplicar una etiqueta de fusión en el C-terminal de Httex1 podría quitar truncamientos en el PRD fácilmente como la proteína truncada perdería así la etiqueta de afinidad. Sin embargo, si la secuencia nativa necesita mantener que esta opción no puede aplicarse como Httex1 termina con prolina y al mejor de nuestro conocimiento no hay no hay etiquetas de fusión C-terminal que son conocidos por inducir escote sin dejar huellas y eficiente después de prolina.
La parte más crítica del protocolo es el manejo de la Httex1 liberada después de la ruptura de la etiqueta SUMO por ULP1. La proteína debe ser purificada inmediatamente por RP-HPLC. Afortunadamente, se trata de una reacción rápida y eficaz que generalmente se completa en 10-20 min a 4 ° C. En contraste, la estrategia intein requiere varias horas de ruptura completa de las inteinas, así requiriendo un compromiso entre la hendidura incompleta y agregación de principio con el fin de maximizar el rendimiento. Un workup rápido es necesario para la mutante Httex1, comienza a agregar en la comparativamente alta concentración presente en la reacción de hendidura, mientras que la variante 23Q es estable por más tiempo. Durante la purificación de RP-HPLC, otra ventaja de SUMO se hace evidente: mientras que la Ssp DnaB intein es hidrófobo y se adhiere fuertemente a la columna, SUMO es más hidrofílico y elutes totalmente de la columna de fase inversa de C4. Aunque ULP1 comercial es bastante costosa, la proteína se puede producir fácilmente en alto rendimiento siguiendo protocolos previamente publicados29.
La importancia de la aplicación de un protocolo de desagregación antes de utilizar Httex1 no puede ser tensionado bastantes. Proteínas poliQ liofilizado como Httex1 son estables y pueden ser almacenados largos periodos, pero no son totalmente solubles en agua y en tampones. La presencia oligómeros preformados o fibrillas podría tener un impacto significativo sobre la cinética de agregación y propiedades biofísicas de la proteína45. El protocolo de desagregación se describe aquí permite la desagregación de la proteína, eliminación de los agregados preformados y generación de una solución de Httex1 monoméricas a partir de una muestra liofilizada. Se observó morfología similar de cinética y fibrilla de agregación de Httex1 obtenidos con el SUMO y la estrategia del intein.
En comparación con los métodos anteriores para la producción de Httex1, la estrategia SUMO aquí descrita ofrece varias ventajas y amplía la gama de posibles estudios para investigar la estructura y propiedades funcionales de esta proteína en salud y enfermedad. La proteína de la fusión de SUMO Httex1 es fácil de manejar, puede ser congelado y almacenado o mantenido en solución por 24 h a temperatura ambiente, mientras que el mHttex1 gratis agregado rápidamente. La estabilidad y la alta solubilidad de las proteínas de fusión de SUMO-Httex1 proporcionan una mayor flexibilidad para manipular la proteína o introducir modificaciones enzimáticas y químicas en mHttex1 que de lo contrario no sería posible después de la hendidura. Esto incluye la introducción de modificaciones post-traduccionales, fluoróforos, spin etiquetas, etiquetas de biotina, etc. los avances presentados aquí deben 1) facilitar futuros estudios para elucidar relaciones estructura-función de Httex1; 2) generan nuevas herramientas para investigar Htt agregación y difusión de la patología; 3) permiten el desarrollo de nuevos ensayos para identificar moléculas que estabilicen a mutante Httex1 y evitar su agregación; y 4) alentar a los científicos de otros campos para llevar a trabajar en esta proteína y Únete a nuestra búsqueda para encontrar curas para la enfermedad de Huntington.
Divulgaciones
Los autores declaran que tienen no hay conflictos de intereses con el contenido de este artículo.
Agradecimientos
Este trabajo fue financiado principalmente por donaciones de la Fundación CHDI y la Swiss National Science Foundation. Agradecemos Dr. Sophie Vieweg discusiones útiles durante el desarrollo de este nuevo sistema de expresión y otros miembros del grupo Lashuel para compartir su experiencia con este sistema de expresión y para su entrada y valiosa retroalimentación. También agradecemos a Prof. Oliver Hantschel para proporcionar el plásmido ULP1. Los autores agradecen Dr. John B. Warner y Dr. Senthil K. Thangaraj revisión crítica del manuscrito
Materiales
| Name | Company | Catalog Number | Comments |
| Uranyl formate (UO2(CHO2)2) | EMS | 22450 | |
| Formvar/carbon 200 mesh, Cu 50 grids | EMS | FCF200-Cu-50 | |
| High Precision Cell made of Quartz SUPRASIL 1 mm light path from Hellma Analytics | HellmaAnalytics | 110-1-40 | |
| Buffer Substance Dulbecco's (PBS w/o Ca and Mg) ancinne ref 47302 (RT) SERVA | Witech | SVA4730203 | |
| Ampicillin | AxonLab | A0839.0100 | |
| Luria Broth (Miller's LB Broth) | Chemie Brunschwig | 1551.05 | |
| Isopropyl β-D-1-thiogalactopyranoside (IPTG) | AxonLab | A1008.0025 | |
| E. coli B ER2566 | NEB | NEB# E4130 | |
| Imidazole | Sigma | 56750-500G | |
| cOmplete Protease Inhibitor Cocktail | Roche | 4693116001 | |
| Anti-Huntingtin Antibody, a.a. 1-82 | Merck Millipore Corporation | MAB5492 | |
| IRDye 680RD Goat anti-Mouse IgG (H + L) | Licor | 925-68070 | |
| PMSF | AxonLab | A0999.0005 | |
| HisPrep 16/10 column | GE Healthcare | 28936551 | |
| C4 HPLC column | Phenomenex | 00G-4168.P0 | 10 µm C4 300 Å, LC Column 250 x 21.2 mm, Phenomenex, 19x10 mm guard column, not temperature jacketed |
| Acetonitrile HPLC | MachereyNagel | C2502 | |
| Filtre seringue Filtropur S 0,45 ul sans prefiltre sterile | Sarstedt AG | 83.1826 | |
| Spectrophotometer semi-micro cuvette | Reactolab S.A. | 2534 | |
| Superloop, 1/16" fittings (ÄKTAdesign), 50 ml | GE Healthcare | 18111382 | |
| Trifluoroacetic acid | Sigma | 302031 | |
| GREINER Tubes fo FPLC 16 x 100 mm, cap. 12.0 ml | Greiner Bio-One | 7.160 102 | |
| 100 kD Microcon fast flow filters | Merck Millipore Corporation | MRCF0R100 | |
| Vibra-cell VCX130 ultrasonic liquid processor | Sonics | ||
| Äkta 900 equipped with a fraction collector | GE Healthcare | ||
| Jasco J-815 Circular Dichroism | Jasco | ||
| Waters UPLC system | Waters | C8 BEH acquity 2.1x100 mm 1.7 micron column , preheated column (40 °C), flow rate of 0.6 mL/min, injection volume of 4 μL | |
| waters HPLC system | Waters | 2489 UV detector and 2535 quaternary gradient module, 20 mL loop in a FlexInject housing | |
| ESI-MS: Finnigan LTQ | Thermo Fisher Scientific | ||
| lyophylizer instrument | FreeZone 2.5 Plus | ||
| Tecnai Spirit BioTWIN | FEI | electron microscope equipped with a LaB6 gun and a 4K x 4K FEI Eagle CCD camera (FEI) and operated at 80 kV | |
| 37 °C shaking incubator | Infors HT multitron Standard | ||
| Biophotometer plus | Eppendorf |
Referencias
- Saudou, F., Humbert, S. The Biology of Huntingtin. Neuron. 89 (5), 910-926 (2016).
- MacDonald, M. E., Gines, S., Gusella, J. F., Wheeler, V. C. Huntington's disease. Neuromolecular Medicine. 4 (1-2), 7-20 (2003).
- Li, S., Li, X. J. Multiple pathways contribute to the pathogenesis of Huntington disease. Molecular Neurodegeneration. 1, 19 (2006).
- DiFiglia, M. Aggregation of Huntingtin in Neuronal Intranuclear Inclusions and Dystrophic Neurites in Brain. Science. 277 (5334), 1990-1993 (1997).
- Atwal, R. S., et al. Huntingtin has a membrane association signal that can modulate huntingtin aggregation, nuclear entry and toxicity. Human Molecular Genetics. 16 (21), 2600-2615 (2007).
- Sathasivam, K., et al. Aberrant splicing of HTT generates the pathogenic exon 1 protein in Huntington disease. Proceedings of the National Academy of Sciences U S A. 110 (6), 2366-2370 (2013).
- Mangiarini, L., et al. Exon 1 of the HD gene with an expanded CAG repeat is sufficient to cause a progressive neurological phenotype in transgenic mice. Cell. 87 (3), 493-506 (1996).
- El-Daher, M. T., et al. Huntingtin proteolysis releases non-polyQ fragments that cause toxicity through dynamin 1 dysregulation. EMBO Journal. 34 (17), 2255-2271 (2015).
- Lunkes, A., et al. Proteases acting on mutant Huntingtin generate cleaved products that differentially build up cytoplasmic and nuclear inclusions. Molecular Cell. 10 (2), 259-269 (2002).
- Vieweg, S., Ansaloni, A., Wang, Z. M., Warner, J. B., Lashuel, H. A. An Intein-based Strategy for the Production of Tag-free Huntingtin Exon 1 Proteins Enables New Insights into the Polyglutamine Dependence of Httex1 Aggregation and Fibril Formation. Journal of Biological Chemistry. 291 (23), 12074-12086 (2016).
- Georgalis, Y., et al. Huntingtin aggregation monitored by dynamic light scattering. Proceedings of the National Academy of Sciences U S A. 95 (11), 6118-6121 (1998).
- Scherzinger, E., et al. Huntingtin-encoded polyglutamine expansions form amyloid-like protein aggregates in vitro and in vivo. Cell. 90 (3), 549-558 (1997).
- Scherzinger, E., et al. Self-assembly of polyglutamine-containing huntingtin fragments into amyloid-like fibrils: implications for Huntington's disease pathology. Proceedings of the National Academy of Sciences U S A. 96 (8), 4604-4609 (1999).
- Muchowski, P. J., et al. Hsp70 and hsp40 chaperones can inhibit self-assembly of polyglutamine proteins into amyloid-like fibrils. Proceedings of the National Academy of Sciences U S A. 97 (14), 7841-7846 (2000).
- Heiser, V., et al. Inhibition of huntingtin fibrillogenesis by specific antibodies and small molecules: implications for Huntington's disease therapy. Proceedings of the National Academy of Sciences U S A. 97 (12), 6739-6744 (2000).
- Bennett, E. J., Bence, N. F., Jayakumar, R., Kopito, R. R. Global impairment of the ubiquitin-proteasome system by nuclear or cytoplasmic protein aggregates precedes inclusion body formation. Molecular Cell. 17 (3), 351-365 (2005).
- Tam, S., et al. The chaperonin TRiC blocks a huntingtin sequence element that promotes the conformational switch to aggregation. Nature Structural & Molecular Biology. 16 (12), 1279-1285 (2009).
- Nekooki-Machida, Y., et al. Distinct conformations of in vitro and in vivo amyloids of huntingtin-exon1 show different cytotoxicity. Proceedings of the National Academy of Sciences U S A. 106 (24), 9679-9684 (2009).
- Wacker, J. L., Zareie, M. H., Fong, H., Sarikaya, M., Muchowski, P. J. Hsp70 and Hsp40 attenuate formation of spherical and annular polyglutamine oligomers by partitioning monomer. Nature Structural & Molecular Biology. 11 (12), 1215-1222 (2004).
- Legleiter, J., et al. Monoclonal antibodies recognize distinct conformational epitopes formed by polyglutamine in a mutant huntingtin fragment. Journal of Biological Chemistry. 284 (32), 21647-21658 (2009).
- Legleiter, J., et al. Mutant huntingtin fragments form oligomers in a polyglutamine length-dependent manner in vitro and in vivo. Journal of Biological Chemistry. 285 (19), 14777-14790 (2010).
- Nucifora, L. G., et al. Identification of novel potentially toxic oligomers formed in vitro. from mammalian-derived expanded huntingtin exon-1 protein. Journal of Biological Chemistry. 287 (19), 16017-16028 (2012).
- Dahlgren, P. R., et al. Atomic force microscopy analysis of the Huntington protein nanofibril formation. Nanomedicine. 1 (1), 52-57 (2005).
- Poirier, M. A., et al. Huntingtin spheroids and protofibrils as precursors in polyglutamine fibrilization. Journal of Biological Chemistry. 277 (43), 41032-41037 (2002).
- Duim, W. C., Chen, B., Frydman, J., Moerner, W. E. Sub-diffraction imaging of huntingtin protein aggregates by fluorescence blink-microscopy and atomic force microscopy. Chemphyschem. 12 (13), 2387-2390 (2011).
- Pieri, L., Madiona, K., Bousset, L., Melki, R. Fibrillar alpha-synuclein and huntingtin exon 1 assemblies are toxic to the cells. Biophysical Journal. 102 (12), 2894-2905 (2012).
- Monsellier, E., Redeker, V., Ruiz-Arlandis, G., Bousset, L., Melki, R. Molecular interaction between the chaperone Hsc70 and the N-terminal flank of huntingtin exon 1 modulates aggregation. Journal of Biological Chemistry. 290 (5), 2560-2576 (2015).
- Isas, J. M., Langen, R., Siemer, A. B. Solid-State Nuclear Magnetic Resonance on the Static and Dynamic Domains of Huntingtin Exon-1 Fibrils. Biochemistry. 54 (25), 3942-3949 (2015).
- Malakhov, M. P., et al. SUMO fusions and SUMO-specific protease for efficient expression and purification of proteins. Journal of Structural Function Genomics. 5 (1-2), 75-86 (2004).
- Mossessova, E., Lima, C. D. Ulp1-SUMO crystal structure and genetic analysis reveal conserved interactions and a regulatory element essential for cell growth in yeast. Molecular Cell. 5 (5), 865-876 (2000).
- Kumari, S., Pal, R. K., Gupta, R., Goel, M. High Resolution X-ray Diffraction Dataset for Bacillus licheniformis Gamma Glutamyl Transpeptidase-acivicin complex: SUMO-Tag Renders High Expression and Solubility. Protein Journakl. 36 (1), 7-16 (2017).
- Zhang, J., Sun, A., Dong, Y., Wei, D. Recombinant Production and Characterization of SAC, the Core Domain of Par-4, by SUMO Fusion System. Applied Biochemistry and Biotechnology. , (2017).
- Reif, A., et al. Semisynthesis of biologically active glycoforms of the human cytokine interleukin 6. Angewandte Chemie International Edition English. 53 (45), 12125-12131 (2014).
- Froger, A., Hall, J. E. Transformation of plasmid DNA into E. coli using the heat shock method. Journal of Visualized Experiments. (6), 253 (2007).
- Smith, B. J. SDS Polyacrylamide Gel Electrophoresis of Proteins. Methods in Molecular Biology. 1, 41-55 (1984).
- Lawrence, A. M., Besir, H. U. Staining of proteins in gels with Coomassie G-250 without organic solvent and acetic acid. Journal of Visualized Experiments. (30), (2009).
- Block, H., et al. Immobilized-metal affinity chromatography (IMAC): a review. Methods in Enzymology. 463, 439-473 (2009).
- Chen, S. M., Wetzel, R. Solubilization and disaggregation of polyglutamine peptides. Protein Science. 10 (4), 887-891 (2001).
- Booth, D. S., Avila-Sakar, A., Cheng, Y. Visualizing proteins and macromolecular complexes by negative stain EM: from grid preparation to image acquisition. Journal of Visualized Experiments. (58), (2011).
- O'Nuallain, B., et al. Kinetics and thermodynamics of amyloid assembly using a high-performance liquid chromatography-based sedimentation assay. Amyloid, Prions, and Other Protein Aggregates, Pt C. 413, 34-74 (2006).
- Greenfield, N. J. Analysis of circular dichroism data. Methods in Enzymology. 383, 282-317 (2004).
- Aiken, C. T., et al. Phosphorylation of threonine 3: implications for Huntingtin aggregation and neurotoxicity. Journal of Biological Chemistry. 284 (43), 29427-29436 (2009).
- Satakarni, M., Curtis, R. Production of recombinant peptides as fusions with SUMO. Protein Expression and Purification. 78 (2), 113-119 (2011).
- Davies, H. A., Wilkinson, M. C., Gibson, R. P., Middleton, D. A. Expression and purification of the aortic amyloid polypeptide medin. Protein Expression and Purification. 98, 32-37 (2014).
- Chen, S., Wetzel, R. Solubilization and disaggregation of polyglutamine peptides. Protein Science. 10 (4), 887-891 (2001).
Reimpresiones y Permisos
Solicitar permiso para reutilizar el texto o las figuras de este JoVE artículos
Solicitar permisoThis article has been published
Video Coming Soon
ACERCA DE JoVE
Copyright © 2025 MyJoVE Corporation. Todos los derechos reservados