
Method Article
דור של Huntingtin Exon1 טבעית, לא מתויגות מונומר, הסיבים משתמש באסטרטגיה פיוז'ן סומו
In This Article
Summary
כאן, אנו מציגים פרוטוקול ממוטבת ועמיד לייצור כמויות מיליגרם של מונומרים מקורי, ללא תג, הסיבים של exon1 של החלבון Huntingtin (Httex1) המבוסס על הפיוז'ן ארעית של צירוף קשורים אוביקוויטין קטן (סומו).
Abstract
מחלת הנטינגטון (HD) היא מחלה תורשתית ניווניות קטלנית הנגרמת על ידי הרחבה החטיבה (≥36) ב אקסון הראשון של הגן HD, והתוצאה היא הביטוי של החלבון Huntingtin (Htt) או N-מסוף קטעים הימנו עם (polyglutamine המורחב מתיחה polyQ).
Exon1 של החלבון Huntingtin (Httex1) נמצא השבר Htt הקטן ביותר זה recapitulates רבות מהתכונות של HD בסלולר, חיה מודלים, נמנה שברי למדה הנפוצים Htt. גודל קטן של Httex1 הופך השפעול יותר נוטה biophysical אפיון באמצעות טכניקות סטנדרטי, רזולוציה גבוהה לעומת קטעים ארוכים או Htt באורך מלא. עם זאת, צבירת גבוהה נטיה של מוטציה Httex1 (mHttex1) עם תוכן מוגבר polyQ (≥42) הפכה אותו קשה לפתח ביטוי יעילה ומערכות טיהור לייצר חלבונים אלה בכמויות מספיקות ולגרום להם נגיש מדענים מתחומים שונים ללא שימוש פיוז'ן חלבונים או שיטות אחרות שמשנות את רצף מקורית של החלבון. אנו מציגים כאן שיטה חזקה ממוטב הייצור של כמויות מיליגרם של ילידים, תג בחינם Httex1 מבוסס על הפיוז'ן ארעית של צירוף קשורים אוביקוויטין קטן (סומו). הפשטות והיעילות של האסטרטגיה יבטל את הצורך להשתמש שנוצרה רצפים של Httex1, ובכך הפיכת חלבון זה יותר נגיש לחוקרים ושיפור הפארמצבטית את הניסויים בין מעבדות שונות. אנו מאמינים כי ההתפתחויות הללו גם יקל על מחקרים עתידיים מכוון שחקרתי את הקשר מבנה פונקציה של Htt וכן בפיתוח כלי אבחון הרומן של טיפולים לטיפול או להאט את ההתקדמות של HD.
Introduction
Htt הוא חלבון 348 kDa, היה מעורב פונקציות פיזיולוגיים מספר1. Htt כוללת אזור polyQ המורחב של שאריות יותר מ 36 אמיני שלה, היא גורמת HD2,3. פתולוגיה HD מאופיין תכלילים הסלולר סטריאטום ואת קליפת המוח, אשר מוביל למוות עצביים, ניוון של רקמות המושפעת4,5. מספר קטעים N-מסוף Htt המכילים את דרכי חזרה polyQ זוהו במוח פוסט-מורטם מחולים HD, הם חשבו להיות שנוצר על-ידי עיבוד הפרוטאוליטי של חלבון huntingtin6. מחקרים שנעשו לאחרונה מראים כי Httex1 גם יכול להיווצר עקב החדרת ה-mRNA חריגה. Httex1 מכיל את המוטציה polyQ פתולוגית, שלה ביטוי אצל בעלי חיים יכולים לסכם רבים של התכונות העיקריות של HD7, ובכך הארת תפקיד מרכזי אפשרי של קטע זה ב HD פתולוגיה ומחלות התקדמות6, 8,9.
עקב צבירת גבוהה נטיה של מוטציה Httex1 (mHttex1) עם polyQ המורחב בדרכי, הרוב המכריע של מערכות קיימות ביטוי מבוססים על הפיוז'ן ארעית של Httex1 חלבונים (כגון גלוטתיון-S-טרנספראז (GST), thioredoxin (TRX) או מלטוז-מחייב-חלבון (MBP) ו/או פפטידים (פולי-היסטידין) באופן שונה לשפר את הביטוי, יציבות, טיהור ו/או מסיסות10,11,12,13,14 ,15,16,17,18,19,20,21,22,23 ,24,25,26,27,28. הזוג פיוז'ן מקושר Httex1 עם רצף קצר המכיל אתר המחשוף פרוטאזות כמו טריפסין, tabacco לחרוט פרוטאז וירוס (אס) או PreScission כדי לאפשר המחשוף ושחרור של Httex1 לפני אתחול של צבירת או טיהור. החסרונות של שיטות אלה כוללים את האפשרות להשאיר משקעים נוספים בשל מחשוף traceless ויצירת של קטעים שנחתכו עקב miscleavage בתוך הרצף של Httex1, בנוסף הטרוגניות עקב המחשוף לא שלם ( ראה Vieweg et al. . לדיון מעמיק יותר על היתרונות ועל המגבלות של גישה זו)10. כדי לטפל מגבלות אלה, שפיתחנו לאחרונה אסטרטגיית הבעה המאפשרת את הדור של Httex1 ללא תג מקורית בפעם הראשונה על-ידי ניצול פיוז'ן N-מסוף ארעית של Synechocystis sp. (Ssp) Intein DnaB Httex110. בעוד המחשוף intein הוא traceless, מדויקת, התשואות mg בכמות החלבונים, היא עדיין סובלת שני החסרונות יכול להפחית את התשואה: כלומר, מוקדמת המחשוף של intein אשר יכול להתרחש במהלך הביטוי, ואת העובדה המחשוף מתרחשת מעל שעות מספר, אשר יכול להוביל לאובדן חלבון עקב צבירת, במיוחד עבור Httex1 עם polyQ המורחב חזרה.
כדי לטפל מגבלות אלה וכדי לחדד את האסטרטגיה שלנו לייצור של מקורי, ללא תג Httex1, פיתחנו מערכת ביטוי חדשה המבוססת על הפיוז'ן ארעית של הסומו, יותר בדיוק השמרים homolog Smt3 כדי Httex1. היישום של מערכת סומו לייצור של חומרים פורסם לראשונה ב 200429, שבה הודגם שיעור מוגבר של הביטוי, המסיסות של חלבון כימרי סומו. יכול להיות ביקע התג סומו מאת אוביקוויטין כמו חלבון ספציפי פרוטאז 1 (ULP1), אשר אינו דורש אתר הכרה, אבל מזהה את מבנה שלישוני של סומו, למעשה מבטלת את האפשרות של miscleavage30. יתר על כן, המחשוף בתיווך ULP1 מהיר, traceless, אינו משאיר משקעים נוספים. המחשוף מוקדמת של התג פיוז'ן, כפי שנצפה עם autocatalytic intein10, לחלוטין נמנעת על ידי הדרישה פרוטאז חיצוניים. ואילו האסטרטגיה סומו משמש כיום נרחב חלבון רקומביננטי ייצור31,32,33, נדגים בעיתון הזה שזה שימושי במיוחד עבור הדור של המתוסבכים מיסודה, חלבון amyloidogenic נוטה צבירה, כגון Httex1. אנו מאמינים כי פשטות, יעילות ועמידות בשיטה שלנו סומו-פיוז'ן מבוססי להפוך Httex1 מקורי, ללא תג יותר נגיש חוקרים מדיסציפלינות שונות מסלק את הצורך להשתמש שנוצרה רצפים של Httex1 במבחנה . זה מקדמה חשובה זה יקל על מחקרים עתידיים להבהיר את הקשר מבנה פונקציה של Httex1.
הפרוטוקול מתאר הטיהור של Httex1 מ L 12 של התרבות חיידקי, אך הפרוטוקול יכול להתאים בקלות להפקות בקנה מידה קטן או גדול יותר. הפרוטוקול מתאר את הייצור של פראי סוג Httex1 (wtHttex1) עם polyQ אני חוזר אורך מתחת (23Q), מוטציה Httex1 (mHttex1) עם polyQ חוזר אורך לעיל (43Q) הסף פתוגניים (36Q).
Protocol
1. ביטוי של Recombinant Httex1 23Q, 43Q
- להכין את מאגרי נדרשים פתרונות. להכין 1000 x פתרון מניות אמפיצילין (כח, 100 מ"ג/מ"ל), מסנן (0.2 µm), aliquot, חנות ב-20 ° C. להכין lysogeny מרק (LB) בינוני (25 גרם ליברות מילר לכל 1 ליטר H2O), תא לחץ. להכין פתרון מניות של 1 מ' איזופרופיל ß-D-1thiogalactopyranoside (IPTG), מסנן (0.2 µm), aliquot, חנות ב-20 ° C.
- המרה כימית המוסמכת e. coli B ER 2566 עם וקטור pTWIN1, המכיל Httex1 האנושי דבוקה תג N-מסוף His6-סומו עם שיטה הלם חום34.
הערה: המתח BL21 DE3 e. coli שימש גם. עם זאת, במקרה זה כמות מוגברת של truncations נצפתה. - לחסן 200 מ ל LB-בינוני, 1 x אמפר בבקבוקון 1 ליטר חרוט על-ידי הוספת שמושבה בודדת מהצלחת אגר עם טיפ פיפטה סטרילי. דגירה התרבות של 30 מעלות צלזיוס, 180 סל ד עבור 20 ש ח (לילה) בחודש חממה חיידקי.
- . קח דגימה 1 מ"ל של התרבות עם פיפטה סטרילי מודדים את צפיפות אופטית על 600 nm (OD600) של המדגם cuvette פלסטיק חד פעמיות, של photometer (כבוד המדידה נעות בין 0.1 ל- 1, שתדללו LB-בינונית במידת הצורך). לחשב את כמות preculture כי תגרום ההתחלה OD600 של 0.05 בתרבות 3 L (עם preculture של יתר600 = 3 זה אומר 50 מ ל).
- לחסן ארבע תרבויות (כל L 3 ליברות-בינוני עם 1 x אמפר בבקבוקון 5 L), על-ידי הוספת כמות preculture עם פיפטה סטרילי מחושב. דגירה התרבויות-37 ° C ו 180 סל ד ב חממה חיידקי.
- בכל 30 דקות, לקחת דגימה 1 מ"ל של התרבות עם פיפטה סטרילי. למדוד את יתר600 של המדגם cuvette פלסטיק חד פעמיות, photometer. כאשר יתר600 הגיעה 0.1 (בדרך כלל לאחר 1-2 h), לקבוע את הטמפרטורה של החממה חיידקי 14 מעלות ולהמשיך הדגירה תוך קירור. בכל 30 דקות, לקחת דגימה 1 מ"ל של התרבות עם פיפטה סטרילי. למדוד את יתר600 של המדגם cuvette פלסטיק חד פעמיות, photometer.
הערה: הזמן לקרר את התרבויות עשויים להשתנות עם החממה משמש, אז הזמן להתחיל קירור ייתכן שיהיה עליך להתאים בהתאם לסוג של החממה משמש. עם זאת, שינוי טמפרטורת מעבר הצבע רק צריך השפעה קטנה על התשואה כמו החלבון פיוז'ן סומו שנראה די יציב. - כאשר יתר600 הגיעה 0.3-0.4 (בדרך כלל לאחר 1-2 h), לקחת דגימה אינדוקציה מראש של התרבות לניתוח מרחביות-דף של ביטוי. חישוב גודל המדגם שנותן כמות דומה של תאים והמוכתמות איתות טוב על Coomassie מרחביות-דף: ג'ל טוב עבור 10: נפח = 0.2 מ"ל/כמנת600; . קח חצי ג'ל טוב 15.
- על תרבות חיידקי עם יתר600= 0.4, לקחת 500 µL. קח את הנפח מחושב של התרבות חיידקי עם פיפטה סטרילי. בדוגמה (18000 x g, 4 ° C, 2 דקות) וזורקים את תגובת שיקוע. לשמור את צניפה ב-20 ° C עד מוכנים לשימוש עבור ניתוח (שלב 1.11).
- זירוז ביטוי חלבונים על-ידי pipetting 1.2 מ של 1 מ' IPTG מניות פתרון כל פתרון תרבות 3 L (הריכוז הסופי 0.4 מ מ). המשך המקננת התרבות ב 14 מעלות צלזיוס במשך 16 h (לילה).
הערה: הטמפרטורה בדרך כלל הגיעו ~ 20 ° C עד שנוסף IPTG, בהתאם הביצועים של החממה. - . קח דגימה שלאחר אינדוקציה של התרבות לניתוח מרחביות-דף של ביטוי, בעקבות ההליך שמתואר בשלב 1.7
- לקצור את התאים על ידי צנטריפוגה צינורות 1 ליטר (3993 x g, 4 ° C, 10 דקות). למחוק את תגובת שיקוע, לשמור בגדר תא בקרח, להמשיך ישירות הטיהור.
- לנתח את ביטוי ידי מרחביות-עמוד35,36. Resuspend הדגימות קדם ומעגל שלאחר ב 20 µL של הפעלת מאגר ו- µL 20 של 2 x טוען לצבוע. מחממים את הדגימות למשך 5 דקות ב 95 מעלות צלזיוס בתוך גוש חום וטען µL 20 על 15% ג'ל בזמן עדיין חם. הפעל את הג'ל במשך 90 דקות ב- 180 (פ') כתם הג'ל עם Coomassie לצבוע לפי ההוראות של היצרן. להשוות את התוצאות עם תוצאות נציג איור 1C.
הערה: הפרוטוקול ניתן לעצור כאן, בגדר התא יכול להיות קפוא, המאוחסנים ב-80 מעלות צלזיוס למשך מספר שבועות. לקבלת תוצאות מיטביות, מומלץ להשתמש בגדר חיידקי טריים ולהימנע לקפוא. הפשרת ההקפאה עלול להוביל פירוק של תאים והשפלות של Httex1. דבר זה עשוי להפחית את התשואה ואת איכות החלבון.
2. התא פירוק וטיהור של שלו6-סומו Httex1 חלבון כימרי ותשמרו על ידי כרומטוגרפיית זיקה מתכת (IMAC)
- להכין 2 L מאגר A בבקבוק זכוכית (50 מ מ טריס (hydroxymethyl)-aminomethan (טריס), 500 מ מ NaCl, imidazole 15 מ מ). להכין 1 ליטר של מאגר B (50 מ"מ טריס, 500 מ מ NaCl, 500 מ מ imidazole pH 7.4) בבקבוק זכוכית. לאחר המסת המלחים, להתאים את רמת ה-pH עם 10 N HCl ולסנן את הפתרונות לתוך בקבוקים טריים עם מסנן העליון בקבוק (0.65 מיקרומטר). הכן של 1000 x phenylmethylsulfonyl פלואוריד (PMSF, 0.3 M) במלאי פתרון, aliquot 100 µL, חנות ב-20 ° C.
הערה: הפרוטוקול נועד לאפשר לכל השלבים של פירוק של כדורי חיידקי ביצועים גבוהים הפוך-פאזי כרומטוגרפיה נוזלית (RP-HPLC) טיהור, lyophilization בתוך 8-9 שעות. כדי להגביל את צבירת ו- proteolysis, מומלץ לעבוד במהירות ללא הפסקה, לבצע את כל השלבים ב 4 ° C או על קרח. - להוסיף 100 µL של פתרון מניות PMSF 5 טבליות (1 לכל 30 מ של נפח סופי) של מעכב פרוטאז 100 מ של טרום מקורר מאגר א הוסף בגדר חיידקי למאגר, homogenize התליה על ידי ערבוב עם פס מגנטי מערבבים ועל ידי pipetting למעלה ולמטה אינטרנט th פיפטה 10 מ"ל סטרילי (~ 30 דקות).
- לחלק את המתלים חיידקים aliquots של 40 מ ל 50 מ ל צינורות פלסטיק חד פעמיות. Sonicate כל aliquot באצווה קרח/מים עבור פירוק התא (70% משרעת, זמן sonication סה כ 5 דקות, במרווחים של 30 s sonication, השהה s 30).
הערה: חשוב כי המדגם לא לחמם במהלך השלב sonication. מומלץ להוסיף מעט מים לאמבט קרח כדי לשפר את פליטת חום במהלך sonication. ההליך sonication ייתכן שיהיה עליך להתאים אם משמש כלי נגינה שונים. שיטות פירוק אחרות כמו מכבש צרפתי או microfluidizer צריך לעבוד גם, יכול להיות מועיל למנוע חימום של צבירה מדגם וחלבון. התקנים אלה היו זמינות במעבדה שלנו, השגנו תוצאות טובות עם פרוטוקול sonication שלנו. - לקחת דגימה של 50 µL של lysate נתרן dodecyl סולפט לזיהוי ג'ל אלקטרופורזה (מרחביות-עמוד) ניתוח. Centrifuge המדגם (18000 x g, 4 ° C, 2 דקות), pipette השבר מסיסים במבחנה חדש. Resuspend השבר לא מסיסים ב- 50 µL מאגר A עם פיפטה. שומרים את הדגימות על הקרח עד מרחביות-דף ניתוח (שלב 2.6).
- להבהיר את lysate על ידי צנטריפוגה (39191 x g, 4 ° C, 60 דקות).
- במהלך השלב צנטריפוגה, לנתח את הצעד פירוק התא על ידי עמודים מרחביות. להוסיף 50 µL של 2 x טוען לצבוע את השבר מסיסים ובחלקם לא מסיסים של lysate בהתאמה. חום במשך 5 דקות ב 95 ° C וטען µL 2 על ג'ל 15% בעוד עדיין חם. הפעל את הג'ל במשך 90 דקות ב- 180 (פ') כתם הג'ל עם Coomassie לצבוע לפי ההוראות של היצרן. להשוות את התוצאות עם תוצאות נציג איור 1C.
- לסנן את תגובת שיקוע (0.45 מיקרומטר, סביבונים). לקחת דגימה של 20 µL של תגובת שיקוע מסוננים לניתוח עמוד מרחביות (שלב 2.11).
הערה: בדרך כלל, נפח של 90 ל- 100 מ"ל של תגובת שיקוע מובהר ומסוננות מתקבל. בדרך כלל, 3 סביבונים מספיקים. אם הסינון היא מסורבלת, נסה להגדיל את מהירות צנטריפוגה ו/או זמן. - לבודד את חלבון כימרי His6-סומו Httex1 מבקטריאלי מובהר lysate על ידי קיבוע המתכת כרומטוגרפיית זיקה (IMAC) במערכת כרומטוגרפיה נוזלית (FPLC) ביצועים מהירים ב 4 ° C37.
- למלא את שקופה lysate לתוך superloop עומס על העמודה Ni-נ (המושפל, ניקה, נטען מחדש לפי ספר ההוראות של היצרן, ריק הקודם לרוץ מומלץ) בעוצמות עמודה 2 מ"ל לדקה לעבור 10 (CV, 200 מ ל) של מאגר A ב- 10 מ ל/דקה לשטוף את החלבונים לא מאוגד.
- Elute החלבון פיוז'ן עם 2.5 CV (50 מ"ל) של 100% של מאגר B-2 מ ל לדקה לשימוש בגודל השבר של 50 mL עבור טעינת ושטיפת ו- 5 מ"ל • תנאי. להשוות את התוצאות עם תוצאות נציג איור 1D.
- לקחת דגימה של כל שבר לניתוח מרחביות-דף (20 µL), מאגר שברים המכיל את חלבון כימרי לפי לשיא chromatogram ה-IMAC. להוסיף (2S, 3S) - 1,4 - Bis (sulfanyl) בוטאן-2, 3-diol (DTT) ו- L-ציסטאין (ריכוז סופי 100 מ מ) האבקה ו להמיס על-ידי היפוך בעדינות את הצינור.
הערה: מניסיוננו הטוהר של החלבון פיוז'ן של שברים שונים היא דומה. כאמצעי זהירות, השברים של החלבון פיוז'ן מטוהרים צריך להיות איחדו במהירות לאחר ה-IMAC כדי למנוע צבירה של שברים מרוכז. בנוסף, מומלץ להמשיך ישירות המחשוף של תג סומו, טיהור HPLC. במידת הצורך, הפרוטוקול ניתן לעצור כאן. הפתרון מדולל של החלבון פיוז'ן היה קפוא חנקן נוזלי, המאוחסנים ב- 80 ° C, מטוהרים לאחר מפשיר ללא ירידה משמעותית התשואה. אחסון של הפתרון מדולל של החלבון פיוז'ן ב 4 מעלות צלזיוס במשך 24 שעות ביממה גם נותן תוצאות דומות. - לנתח IMAC מאת מרחביות-דף. הוסף 20 µL של טעינת לצבוע את כל דגימה. לטעון את 2 µL של החומר הגולמי (2.7), השבר לא מאוגד, השבר לשטוף כל שבר הפסגה • תנאי ג'ל 15%. הפעל את הג'ל במשך 90 דקות ב- 180 (פ') כתם הג'ל עם Coomassie לצבוע לפי ההוראות של היצרן. להשוות את התוצאות עם תוצאות נציג איור 1D.
3. המחשוף שלו6-סומו-תג וטיהור HPLC
התראה: חומצה Trifluoroacetic (TFA) הוא נוזל נדיף, יכול לגרום לכוויות קשות כל כך לטפל. לבצע כל טיפול בשכונה fume וללבוש מספקת ציוד מגן אישי (קרי, חד פעמיות nitrile כפפות, בטיחות משקפיים, חלוק מעבדה).
- בבקבוק 5 L, הוסף 5 מ של TFA עם מזרק פלסטיק 5 לליטר מים (הממס H ת2O, 0.1% (TFA). להוסיף 2.5 מ של TFA עם מזרק פלסטיק בקבוק 2.5 L acetonitrile (הממס ב': acetonitrile, 0.1% TFA).
- הכן מערכת HPLC כפי שהוצע על ידי היצרן. לבצע ריצה ריק כדי להבטיח עמודה נקי.
- לקחת דגימה של 100 µL של החלבון פיוז'ן לפני התוספת של ULP1 כדי לנטר את התגובה המחשוף מאת UPLC (שלב 3.5).
- 20 מ של החלבון פיוז'ן להעביר צינור 50 מ ל, להוסיף 0.4 מ של פתרון מניות ULP1, דגירה על קרח. שמור את חלבון כימרי הנותרים על קרח.
הערה: שלו מתויג קטליטי השבר 403-621 הפרוטאז אוביקוויטין-כמו-ספציפי 1 (המכונה כאן "ULP1") שימש כדי לבקע את התג סומו. החלבון היתוך יציב יותר Httex1 cleaved. מומלץ לא כדי לדבוק התג סומו של אצוות שלם. במקום זאת, להמשיך עם aliquots של גודל שניתן באופן ישיר לחלוטין להחיל על העמודה HPLC. - כל 10 דקות, לקחת דגימה של 100 µL של התגובה המחשוף מעקב אחר ההתקדמות על-ידי ביצועים אולטרה כרומטוגרפיה נוזלית (UPLC). Centrifuge את הדגימות (18000 סל ד, 4 ° C, 2 דקות) וניתוח µL 2 של תגובת שיקוע על-ידי UPLC (ההדרגתי מ 10% כדי 90% ממס B ב- A עבור 0.25 עד 3 דקות, 10% B עבור 1 דקות, עיין בהוראות היצרן לשימוש מכשיר). להשוות את chromatograms השיג עבור המדגם לפני תוספת של ULP1, את הדגימות נלקחה. להשוות את התוצאות עם תוצאות נציג איור 2B.
- לאחר סיום הסומו-המחשוף (הפסגה של החלבון פיוז'ן נעלם בתוך chromatogram UPLC, מומרת באופן מלא אל פסגת סומו, Httex1 לאחרונה הופעת), לסנן את הדגימה עם מסנן מזרק (0.22 מיקרומטר).
הערה: המחשוף סומו הוא בדרך כלל מהיר מאוד (10-20 דקות ב 4 ° C) ולכן ניתוח UPLC עם זמן ריצה של 4 דקות הוא כלי חשוב כדי לפקח על התגובה. טיהור, סינון המדגם לפני HPLC הוא בעיקר אמצעי מניעה כדי להגדיל את אורך החיים של העמודה. המדגם לא מומלץ לבטל עכורים. - לטהר את הדגימה מסוננת על-ידי RP-HPLC (הדרגתי של 25-35% ממס B a הממס, מעל 40 דקות-15 מ"ל לדקה (0-10 דקות: 5%; 10-12.5 דקות: 5 עד 25%; 12.5-52.5 מין: 25 ל-35%; 52.5-57.5 מינימום 35 עד 95%; עיין בהוראות היצרן לשימוש מכשיר). להשוות את התוצאות עם תוצאות נציג איור 2C.
הערה: Httex1 ושלו6-סומו נפרד היטב על ידי RP-HPLC. עם זאת, לא יכול להיות כמויות קטנות של Httex1 קטום ב ההתחלה ואת הסוף של הפסגה. לאסוף שברים קטנים כדי להשיג את הכמות המרבית של חומר טהור.
התראה: להשתמש בציוד בטיחות המתאים (קרי, חלוק, כפפות מבודדות, מגן הפנים) כאשר טיפול קריוגני נוזלים. - לנתח HPLC-השברים ידי אלקטרו-תרסיס יינון ספקטרומטר מסה (ESI-MS, תעשיה, להזריק 10 µL, זרימה 0.6 מ"ל/min, הממס: 20% B a, אין עמודת, עיין בהוראות היצרן לשימוש מכשיר) ו- UPLC (מעבר מ 10% ל- 90% ממס B ב- A עבור 0.25 עד 3 דקות, 10% B עבור 1 דקות, עיין בהוראות היצרן לשימוש כלי). בריכה שברים של טוהר דומה צינורות פלסטיק 50 מל, להקפיא במצב חנקן נוזלי, lyophilize. שוקל ולאחר להעביר את החלבון lyophilized לתוך צינורות פלסטיק 2 מ"ל ולאחסן ב-20 ° C.
- לאפיין את החומר מטוהרת על ידי UPLC, ESI-MS מרחביות-דף. להמיס 100 µg של Httex1 lyophilized ב µL 8 של TFA מסודר צינור 1.5 מ ל, תקופת דגירה של 20 דקות בטמפרטורת החדר בו מום סגור. בזהירות להתנדף TFA ברדס fume עם זרם של חנקן או ארגון. השתמש לחץ נמוך של חנקן/ארגון כדי למנוע אובדן של המדגם.
- להמיס את החלבון µL 100 של µL2O. לנתח 2 H על-ידי UPLC, 5 µL על ידי ESI-MS כמו שלב 3.8. מיקס 20 µL של חלבון פתרון עם 20 µL של 2 x טוען לצבוע.
- לנתח כמויות של µg 1 כדי µg 10 מאת מרחביות-דף. הפעל את הג'ל במשך 90 דקות ב- 180 (פ') כתם הג'ל עם Coomassie לצבוע לפי ההוראות של היצרן. להשוות את התוצאות עם תוצאות נציג איור 2D.
4. disaggregation ו- Resolubilization של חלבונים Httex1
התראה: TFA נמצא נוזל נדיף, יכול לגרום לכוויות קשות כל כך לטפל. לבצע כל טיפול בשכונה fume וללבוש מספקת ציוד מגן אישי (דהיינו חד פעמיות nitrile כפפות, בטיחות משקפיים, חלוק מעבדה).
- הכינו 10 מ ל תמיסת פוספט באגירה של Dulbecco (DPBS) (137 מ מ NaCl, מ מ 2.7 אשלגן כלורי, 10 מ מ נה2HPO4, 2 מ מ ח'2PO4, pH 7.4) מן האבקה מעורבבים מראש בצינור 50 מ. לסנן את הפתרון DPBS דרך מסנן 0.2 µm לפני כל שימוש.
- להמיס 150 µg של Httex1 lyophilized ב µL 12 של TFA מסודר צינור 1.5 מ ל, תקופת דגירה של 20 דקות בטמפרטורת החדר בו מום סגור. בזהירות להתנדף TFA ברדס fume עם זרם של חנקן או ארגון. השתמש לחץ נמוך של חנקן/ארגון כדי למנוע אובדן מדגם38.
הערה: באופן כללי, השתמש 4 µL TFA לפרק ו disaggregate 50 µg של חלבון. הליך זה יהיה ליצור סרט של חלבון בתוך כותלי בו מום. כדי למנוע צבירה מיידית של Httex1 בשלבים הבאים, לעבוד עם אנשים רגילים טרום מקורר, לשמור את החלבון תמיד על הקרח ולמנוע ריכוזים גבוהים. - להמיס את החלבון disaggregated של 1 מ"ל של טרום מקורר DPBS ולהתאים את ה-pH ל 7.2-7.4 עם 1 M NaOH. לסנן את הפתרון חלבון דרך מסנן צנטריפוגלי 100 kDa לתוך צינורות פלסטיק 1.5 mL (20000 x g, 4 ° C, 20 דקות).
הערה: הריכוז התיאורטי המחושב של Httex1 הוא גבוה יותר מאשר הריכוז הסופי הרצוי עבור אובדן אפשרי. השלב הסינון הוא צורך להסיר כל אגרגטים זה אולי יצרו במהלך פירוק החלבון. - לקבוע את הריכוז של Httex1 באמצעות עקומת כיול UPLC מבוסס על חומצת אמינו ניתוח (זיהוי-λ214) ונשלח µg 2 של החלבון חומצת אמינו ניתוח כדי לאמת את ריכוז10. לחשב את כמות DPBS שצריכה להתווסף בריכוז מיקרומטר 3 Httex1.
- לדלל את החלבון מיקרומטר 3 על-ידי הוספת הסכום המחושב של DPBS בו מום. שמור את הצינורית על הקרח עד החניכה של פרוטוקול צבירה.
הערה: Httex1 43Q אין לאחסן בפתרון. תמיד הכינו פתרון חלבון טרי המבוסס על פרוטוקול לעיל. Httex1 חלבונים מאוחסנים בצורה הטובה ביותר כאבקה lyophilized ב-20 ° C.
5. ניטור של צבירת קינטיקה של Httex1 43Q באמצעות UPLC, ספקטרוסקופיה Dichroism (CD) מעגלית, אפיון מצרפי על ידי שידור מיקרוסקופ אלקטרונים (TEM)
- להכין פתרון formate uranyl TEM כפי שדווחה בעבר39.
- ליזום את המצבור של Httex1 43Q על ידי המקננת פתרון מיקרומטר 3 ב- DPBS ב 37 מעלות צלזיוס (שימוש 1 מ"ל של פתרון מוכן כמתואר לעיל בפרוטוקול disaggregation).
הערה: ניתן לבצע הצבירה של Httex1 בריכוזים גבוהים בהתאם הצרכים ואת המטרות של הניסוי. - לכמת את כמות חלבון מסיס באמצעות UPLC בנקודות הזמן המצוין (ב- 0, 1, 2, 4, 6, 8, 12, 24, 48 ו 120 h). כדי לעשות זאת, לקחת את aliquot של 35 µL ולהסיר מצרפי לא מסיסים על ידי צנטריפוגה (20000 x g, 4 ° C, 20 דקות). מזריקים µL 4 של תגובת שיקוע לתוך UPLC. לחשב את הפרופורציה של מונומר מסיסים בהתבסס על השינוי של אזור הפסגה באמצעות כלי תוכנה 40. להשוות את התוצאות עם תוצאות נציג איור 3א.
- לאפיין את שינויי מבנה שניוני באמצעות תקליטור ספקטרוסקופיה-0 ל- 48 שעות. . קח את aliquot של 100 µL ולמדוד את ellipticity (1 מ מ קוורץ cuvette, 195 nm 250 ננומטר, 20 ° C, הנתונים מצביע כל 0.2 ננומטר, מהירות 10 ננומטר/min, אינטגרציה הדיגיטלי זמן 2 s, רוחב הפס של 1.0 ננומטר). לרכוש 6 ספקטרום של הדגימה של הממוצע וחלק באמצעות מסנן בינומי עם רוחב קונבולוציה של 99. מגרש הספקטרום כמו שאריות רשע ellipticity טוחנת (θ באמצעותMRE)41. להשוות את התוצאות עם תוצאות נציג איור 3B.
- מאפיינים מורפולוגיים מבניים המאפיינים של מצרפי מאת TEM. את µL 3 של הפתרון חלבון אל Formvar/פחמן-מצופה 200-רשת, משוחררים-זוהר נחושת רשת 1 מינימלית שטיפת הרשת פעמיים עם מים μL 15, פעם אחת עם 15 μL של-0.7% (w/v) uranyl formate, הכתם ב-30 s עם 15 μL כ- 0.7% w/v uranyl formate. לבצע ניתוח TEM של הרשתות. להשוות את התוצאות עם תוצאות נציג איור 3ג.
תוצאות
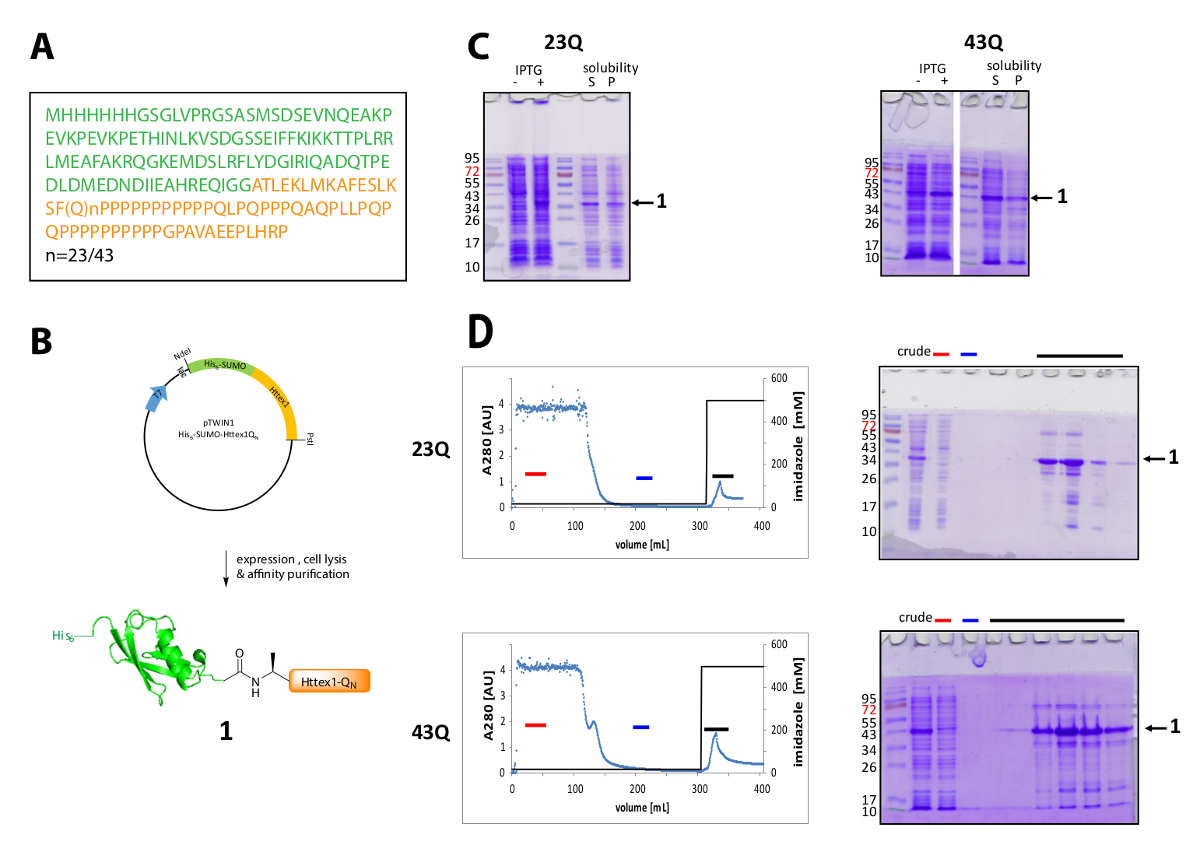
Httex1 מתבטאת e. coli עם N-מסוף שלו6-סומו תג. התוצאות נציג של הביטוי, טיהור של החלבון פיוז'ן מסוכמות באיור1. רצף Httex1 מורכב שאריות 2-90 Htt ומתחיל עם Ala2, כי Met1 מלא cleaved ויוו42. המספור חומצות אמינו מתייחסת 23Q variant, הסדרה מלאה של החלבון פיוז'ן ביטוי מוצג איור 1א'. פלסמידים יופקד אצל Addgene בעתיד הקרוב לשיתוף עם הקהילה. תיאור סכמטי של פלסמיד ואת החלבון פיוז'ן ביטוי מוצג באיור 1ב'. Httex1 His6-סומו מבטאת ברמה בינונית (איור 1C), רוב החלבון היתוך הוא נוכח השבר מסיסים לאחר פירוק, גם את 23Q וגם את הווריאציה 43Q. החלבון פיוז'ן נודד גבוה מהצפוי, מבוסס על משקל מולקולרי. זה בין השאר בשל העדר חזקה של סומו אלא בעיקר בגלל ההרכב רצף חריג של Httex1, המכיל בעיקר שאריות וגם גלוטמין פרולין. את wildtype (23Q) והן החשבונאי (43Q) חלבון כימרי יכול להיות מועשר לטוהר ~ 80% על-ידי IMAC (איור 1D) הנוכחות של שיתוף טיהור מארח חלבון יכולה להיות מוסברת על-ידי רמת ביטוי נמוך יחסית של Httex1, המדגם גדול נפח חלה על העמודה.
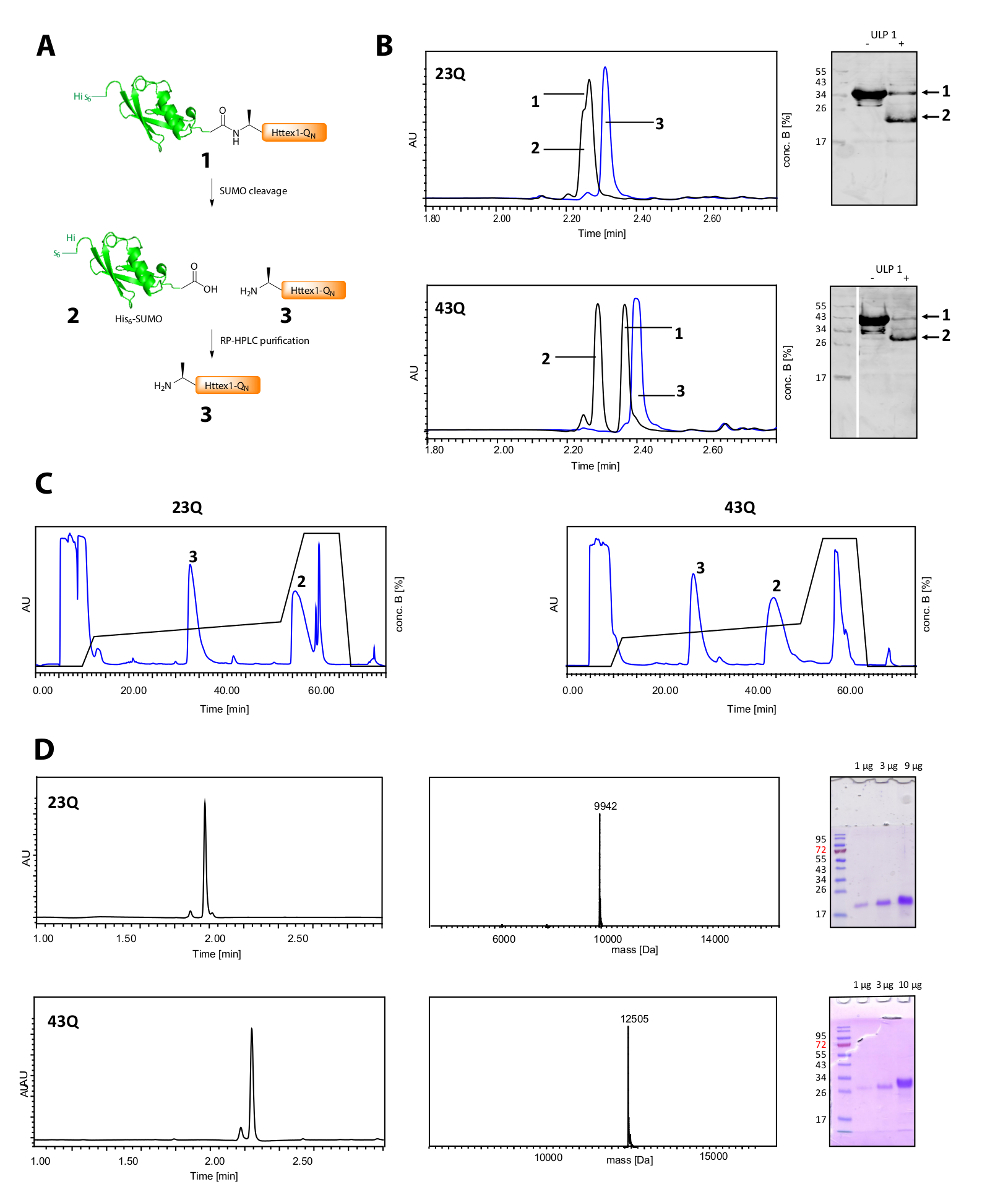
המחשוף שלו6-סומו תג ולטיהור Httex1 מוצג באיור 2א. UPLC הוא כלי יעיל כדי לנטר את המחשוף שלו6-סומו תג (איור 2B). הפסגה המקורי של החלבון היתוך הוא נצרך ושל פסגות חדשות, מופרדים היטב שני המקביל שלו6-תג סומו, Httex1 מופיעים. התגובה המחשוף מסתיים בעוד 10-20 דקות. המערבי כתם (WB) הוא איטי מדי לפקח ביעילות את התגובה המחשוף, אבל זה כבר נכלל באיור לעיון וכדי להדגים את שלמות התרגום של המחשוף סומו. שני Httex1 23Q ו- 43Q ניתן להפריד מנקודת מבט שלו6-סומו תג על ידי RP-HPLC (איור 2C), התקבלו אצל טוהר גבוהה, כפי שהראה UPLC, MS וניתוח מרחביות-דף (איור 2D).
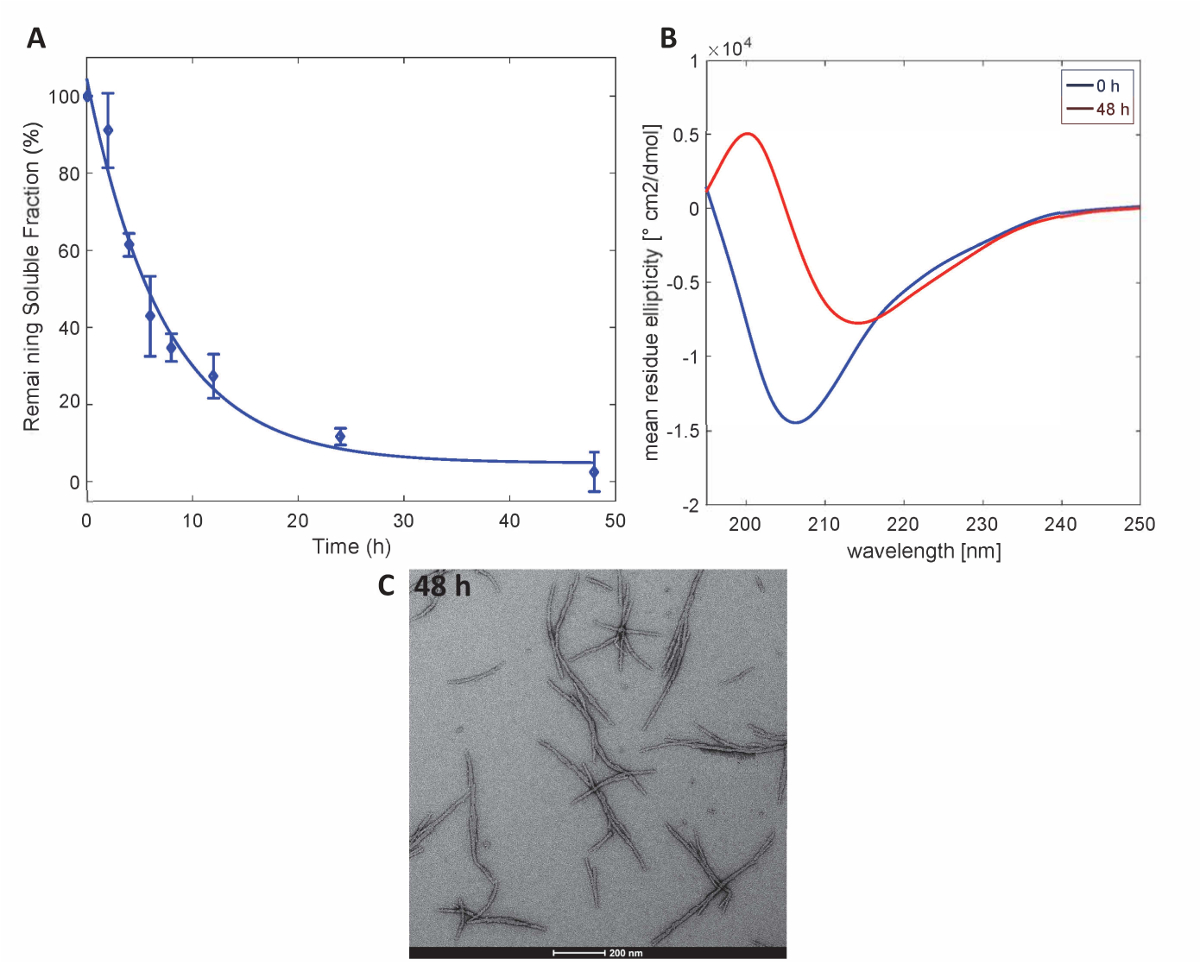
כדי להמחיש כי החלבונים Httex1 שהוכנו על ידי שיטה זו לשמור על המאפיינים צבירת הצפוי של Httex1, אנחנו לאומדן של קינטיקה fibrillization של מוטציה Httex1 ב 37 מעלות צלזיוס וזמינותו שיקוע, פיקוח שינויי מבנה שניוני באמצעות תקליטור ספקטרוסקופיה, מאופיין המורפולוגיה של מצרפי מאת TEM. ערכת נתונים נציג של קינטיקה צבירת היווצרות fibril mHttex1 כפי שנקבע על ידי וזמינותו שקיעת מוצגבאיור 3א. אובדן מסיסים Httex1 43Q לאורך זמן, עקב היווצרות fibril היה לכמת על-ידי UPLC. אנו מבחינים של דלדול מלא של חלבון מסיס לאחר 48 שעות של דגירה. בנוסף, אנחנו נקבע את מבנה שניוני של החלבון על ידי תקליטור ספקטרוסקופיה (איור 3ב). Httex1 43Q משמרות החל לא מובנים (λmin 205 nm) כדי בעיקר β-גיליון עשיר קונפורמציה (λmin 215 ננומטר) לאחר 48 שעות של דגירה. השינוי המבני הזה מלווה היווצרות של אגרגטים זמן fibrillar כמו הנצפה על ידי TEM ב 48 שעות (איור 3ג').

איור 1 . הביטוי ולטיהור שלו6-חלבון כימרי סומו Httex1.
(א) שלו על רצף חומצות אמיניות6-סומו-Httex1-QN פיוז'ן בונה (שלו6-סומו ירוק ו- Httex1-QN ב orange); (B) סקירה סכמטי של הביטוי, טיהור של החלבון פיוז'ן; (ג) אנליזה מרחביות-דף של הביטוי, המסיסות החלבון פיוז'ן לאחר פירוק; (ד) Chromatogram של הטיהור IMAC של החלבון פיוז'ן וניתוח של שברים מאת מרחביות-דף (הבר באדום: שבר לא מאוגד, בלו בר: שטיפת שבר, שחור בר: שברים המכיל הפסגה • תנאי); אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 2 . המחשוף שלו6 טיהור החלבונים Httex1-QN ללא תגית ותגית סומו.
סקירה סכימטי (א); (B) ניתוח של המחשוף של התג סומו עם ULP1 על ידי UPLC (כחול: לפני תוספת של ULP1; שחור: 20 דקות (23Q), בהתאמה 10 דקות (43Q) לאחר תוספת של ULP1) ו WB (MAB5492 1:2000, עז משני אנטי עכבר נוגדן 1:5000); (ג) Chromatogram של הטיהור RP-HPLC מפוח של Httex1; D: ניתוח Httex1 מטוהרת על-ידי UPLC, למען חברה דמוקרטית-דף ו- ESI-MS; משקל מולקולרי הצפוי הוא 9943 Da (23Q) ודה 12506 (43Q) בהתאמה. אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}

איור 3 . צבירה של Httex1-43Q: assay שיקוע (A) מבוסס על UPLC. (B) CD ספקטרום של המבנה משני הו ו 48 ה (ג) micrographs TEM של מצרפי-48 שעות (סולם ברים הם 200 ננומטר). אנא לחץ כאן כדי להציג גירסה גדולה יותר של הדמות הזאת.
{kind=link}
Discussion
ב פרוטוקול זה, אנחנו המחולקות הליך יעיל עבור להשיג כמויות מיליגרם של מקורי, לא מתויגות Httex1 המכילים שאריות גלוטמין 23 או 43. דבר זה הושג על ידי הבעת Httex1 כמו פיוז'ן C-מסוף כדי שלו6-סומו תג, אשר משמש כדי לבודד את חלבון כימרי מהתא lysate על ידי ה-IMAC הוא ביקע לפני טיהור HPLC של Httex1. ואילו האסטרטגיה סומו שימש בייצור של מספר חלבונים אחרים, השיטה שלנו מראה כי תכונותיו הייחודיות שסומו יכול לשמש גם כדי ליצור ממהותם מגרה, צבירת-מועדים, חלבון amyloidogenic הוכחו בעבר קשה מאוד לטפל ולהפיק43,44. אנו מציגים פרוטוקול זה פשוט, קל לשימוש, להשוות פרוטוקול לדור של חלבון "נאותה". פיוז'ן סומו solubilizes ומייצב Httex1 במהלך השלב טיהור IMAC וביטוי. פצילות מוקדמת של התג, כפי שנצפה עם אסטרטגיה intein10 , צבירת היו כבר לא בעיה.
חלבונים ממהותם המתוסבכים פגיעים במיוחד השפלה. בעוד N-מסוף השפלה באזור N17 אינה בעיה באמצעות פרוטוקול זה, truncations של ההקלטה של Httex1 יכול להתרחש. החלבונים קטום דומים מאוד Httex1 hydrophobicity, תשלום, בגודל, הסרתם באמצעים כרומטוגרפי הוא מאתגר, ולכן עדיף למנוע היווצרות שלהם מלכתחילה. נצמדים היטב הפרוטוקול, תמיד עובד על הקרח ושימוש כמות מספקת של מעכבי פרוטאז אמור לעזור לשמור על הרמה של החיתוך שנצפה נמוך מאוד. החלת תג פיוז'ן ב C-הסופית של Httex1 יכול להסיר truncations ההקלטה בקלות כמו החלבון קטום יפסיד תג קירבה. עם זאת, אם הרצף המקורי צריך להיות מתוחזק שאין אפשרות להחיל את אפשרות זו כפי Httex1 מסתיים עם פרולין, לפי מיטב ידיעתנו ישנם אין תגיות פיוז'ן C-מסוף ידועים לזירוז פצילות traceless ויעילה לאחר פרולין.
החלק הקריטי ביותר של הפרוטוקול הוא טיפול Httex1 משוחררת לאחר המחשוף של התג סומו ע י ULP1. החלבון צריך להיטהר באופן מיידי על-ידי RP-HPLC. למרבה המזל, זה לתגובה, מהיר ויעיל, בדרך כלל שהושלמו ב- 10-20 דקות ב 4 º C. לעומת זאת, האסטרטגיה intein נדרש מספר שעות למחשוף מלאה של intein, וכך מצריכים פשרה בין תחילת צבירת המחשוף לא שלם על מנת למקסם את התשואה. בדיקה מהירה נדרשת עבור מוטציה Httex1, כמו זה יהיה להתחיל לצבור על הריכוז הגבוה יחסית נוכח התגובה המחשוף, ואילו הגרסה 23Q יציב למשך זמן ארוך יותר. במהלך הטיהור RP HPLC, יתרון נוסף של סומו הופך לכאורה: בעוד intein DnaB Ssp דוחה מים נדבק בחריפות העמודה, סומו הוא יותר הידרופיליות, elutes לחלוטין מן העמודה הפוך-פאזי C4. ULP1 מסחרי אמנם קצת יותר יקר, החלבון יכול להיות בקלות מיוצר תשואה גבוהה בעקבות פרוטוקולים שפורסמו בעבר29.
חשיבות קריטית החלת נוהל disaggregation לפני שימוש Httex1 לא יכול להיות מתוח מספיק. Lyophilized polyQ חלבונים כגון Httex1 יציבים, יכולים להיות מאוחסנים תקופות ארוכות, אך אינם לגמרי מסיסי מים, מאגרי. הנוכחות של preformed oligomers או הסיבים יכול להיות השפעה משמעותית על צבירת קינטיקה ומאפיינים ביופיזיקלי חלבון45. פרוטוקול disaggregation המתוארים כאן מאפשר את disaggregation של החלבון, הסרה של אגרגטים preformed, דור של פתרון של Httex1 monomeric מתוך מדגם lyophilized. הבחנו מורפולוגיה קינטיקה, fibril צבירת דומה עבור Httex1 שהושג עם בגלל הסומו ואסטרטגיה intein.
לעומת שיטות קודמות לייצור Httex1, האסטרטגיה סומו המתוארים כאן מציעה מספר יתרונות, מרחיבה את מגוון מחקרים ניתן לחקור את מבנה ומאפייני פונקציונלי של חלבון זה על בריאות ומחלה. החלבון פיוז'ן סומו-Httex1 קל לטפל, שניתן יהיה קפוא, לאחסן או שמרו על פתרון עבור 24 שעות בטמפרטורת החדר, בעוד mHttex1 חינם צבירה במהירות. יציבות, המסיסות גבוהה החלבונים פיוז'ן סומו-Httex1 מספקים גמישות רבה יותר לתמרן החלבון ו/או להציג את השינויים אנזימטי והכימיות לתוך mHttex1 שאחרת לא היה אפשרי לאחר המחשוף. זה כולל המבוא של שינויים post-translational, fluorophores, ספין תוויות, ביוטין תגיות, וכו ' ההתקדמות המובאת כאן צריך 1) לקדם מחקרים עתידיים התירי קשרי גומלין מבנה פונקציה של Httex1; 2) ליצור כלים חדשים לחקור Htt צבירת והפצת פתולוגיה; 3) לאפשר הפיתוח של מבחני חדש לזיהוי מולקולות לייצב את המוטציה Httex1 ולמנוע צבירת שלה; . ו-4) לעודד מדענים מתחומים אחרים להביא לעבוד על חלבון זה ולהצטרף החיפוש שלנו כדי למצוא תרופות עבור מחלת הנטינגטון.
Disclosures
המחברים מצהירים כי יש להם שאין ניגודי אינטרסים עם התוכן של מאמר זה.
Acknowledgements
עבודה זו מומן בעיקר על ידי מענקים קרן CHDI, הקרן הלאומית למדע שוויצרי. אנו מודים ד ר סופי Vieweg לדיונים שימושי במהלך הפיתוח של מערכת חדשה זו הביטוי ואת שאר חברי הקבוצה Lashuel לשיתוף ניסיונם עם מערכת זו ביטוי ועל המשוב שלהם קלט ובעל ערך. אנו מודים גם פרופסור אוליבר Hantschel עבור מספקים את פלסמיד ULP1. המחברים תודה ד ר ג'ון B. וורנר, ד ר יצחק ק' Thangaraj על סקירה ביקורתית של כתב היד
Materials
| Name | Company | Catalog Number | Comments |
| Uranyl formate (UO2(CHO2)2) | EMS | 22450 | |
| Formvar/carbon 200 mesh, Cu 50 grids | EMS | FCF200-Cu-50 | |
| High Precision Cell made of Quartz SUPRASIL 1 mm light path from Hellma Analytics | HellmaAnalytics | 110-1-40 | |
| Buffer Substance Dulbecco's (PBS w/o Ca and Mg) ancinne ref 47302 (RT) SERVA | Witech | SVA4730203 | |
| Ampicillin | AxonLab | A0839.0100 | |
| Luria Broth (Miller's LB Broth) | Chemie Brunschwig | 1551.05 | |
| Isopropyl β-D-1-thiogalactopyranoside (IPTG) | AxonLab | A1008.0025 | |
| E. coli B ER2566 | NEB | NEB# E4130 | |
| Imidazole | Sigma | 56750-500G | |
| cOmplete Protease Inhibitor Cocktail | Roche | 4693116001 | |
| Anti-Huntingtin Antibody, a.a. 1-82 | Merck Millipore Corporation | MAB5492 | |
| IRDye 680RD Goat anti-Mouse IgG (H + L) | Licor | 925-68070 | |
| PMSF | AxonLab | A0999.0005 | |
| HisPrep 16/10 column | GE Healthcare | 28936551 | |
| C4 HPLC column | Phenomenex | 00G-4168.P0 | 10 µm C4 300 Å, LC Column 250 x 21.2 mm, Phenomenex, 19x10 mm guard column, not temperature jacketed |
| Acetonitrile HPLC | MachereyNagel | C2502 | |
| Filtre seringue Filtropur S 0,45 ul sans prefiltre sterile | Sarstedt AG | 83.1826 | |
| Spectrophotometer semi-micro cuvette | Reactolab S.A. | 2534 | |
| Superloop, 1/16" fittings (ÄKTAdesign), 50 ml | GE Healthcare | 18111382 | |
| Trifluoroacetic acid | Sigma | 302031 | |
| GREINER Tubes fo FPLC 16 x 100 mm, cap. 12.0 ml | Greiner Bio-One | 7.160 102 | |
| 100 kD Microcon fast flow filters | Merck Millipore Corporation | MRCF0R100 | |
| Vibra-cell VCX130 ultrasonic liquid processor | Sonics | ||
| Äkta 900 equipped with a fraction collector | GE Healthcare | ||
| Jasco J-815 Circular Dichroism | Jasco | ||
| Waters UPLC system | Waters | C8 BEH acquity 2.1x100 mm 1.7 micron column , preheated column (40 °C), flow rate of 0.6 mL/min, injection volume of 4 μL | |
| waters HPLC system | Waters | 2489 UV detector and 2535 quaternary gradient module, 20 mL loop in a FlexInject housing | |
| ESI-MS: Finnigan LTQ | Thermo Fisher Scientific | ||
| lyophylizer instrument | FreeZone 2.5 Plus | ||
| Tecnai Spirit BioTWIN | FEI | electron microscope equipped with a LaB6 gun and a 4K x 4K FEI Eagle CCD camera (FEI) and operated at 80 kV | |
| 37 °C shaking incubator | Infors HT multitron Standard | ||
| Biophotometer plus | Eppendorf |
References
- Saudou, F., Humbert, S. The Biology of Huntingtin. Neuron. 89 (5), 910-926 (2016).
- MacDonald, M. E., Gines, S., Gusella, J. F., Wheeler, V. C. Huntington's disease. Neuromolecular Medicine. 4 (1-2), 7-20 (2003).
- Li, S., Li, X. J. Multiple pathways contribute to the pathogenesis of Huntington disease. Molecular Neurodegeneration. 1, 19 (2006).
- DiFiglia, M. Aggregation of Huntingtin in Neuronal Intranuclear Inclusions and Dystrophic Neurites in Brain. Science. 277 (5334), 1990-1993 (1997).
- Atwal, R. S., et al. Huntingtin has a membrane association signal that can modulate huntingtin aggregation, nuclear entry and toxicity. Human Molecular Genetics. 16 (21), 2600-2615 (2007).
- Sathasivam, K., et al. Aberrant splicing of HTT generates the pathogenic exon 1 protein in Huntington disease. Proceedings of the National Academy of Sciences U S A. 110 (6), 2366-2370 (2013).
- Mangiarini, L., et al. Exon 1 of the HD gene with an expanded CAG repeat is sufficient to cause a progressive neurological phenotype in transgenic mice. Cell. 87 (3), 493-506 (1996).
- El-Daher, M. T., et al. Huntingtin proteolysis releases non-polyQ fragments that cause toxicity through dynamin 1 dysregulation. EMBO Journal. 34 (17), 2255-2271 (2015).
- Lunkes, A., et al. Proteases acting on mutant Huntingtin generate cleaved products that differentially build up cytoplasmic and nuclear inclusions. Molecular Cell. 10 (2), 259-269 (2002).
- Vieweg, S., Ansaloni, A., Wang, Z. M., Warner, J. B., Lashuel, H. A. An Intein-based Strategy for the Production of Tag-free Huntingtin Exon 1 Proteins Enables New Insights into the Polyglutamine Dependence of Httex1 Aggregation and Fibril Formation. Journal of Biological Chemistry. 291 (23), 12074-12086 (2016).
- Georgalis, Y., et al. Huntingtin aggregation monitored by dynamic light scattering. Proceedings of the National Academy of Sciences U S A. 95 (11), 6118-6121 (1998).
- Scherzinger, E., et al. Huntingtin-encoded polyglutamine expansions form amyloid-like protein aggregates in vitro and in vivo. Cell. 90 (3), 549-558 (1997).
- Scherzinger, E., et al. Self-assembly of polyglutamine-containing huntingtin fragments into amyloid-like fibrils: implications for Huntington's disease pathology. Proceedings of the National Academy of Sciences U S A. 96 (8), 4604-4609 (1999).
- Muchowski, P. J., et al. Hsp70 and hsp40 chaperones can inhibit self-assembly of polyglutamine proteins into amyloid-like fibrils. Proceedings of the National Academy of Sciences U S A. 97 (14), 7841-7846 (2000).
- Heiser, V., et al. Inhibition of huntingtin fibrillogenesis by specific antibodies and small molecules: implications for Huntington's disease therapy. Proceedings of the National Academy of Sciences U S A. 97 (12), 6739-6744 (2000).
- Bennett, E. J., Bence, N. F., Jayakumar, R., Kopito, R. R. Global impairment of the ubiquitin-proteasome system by nuclear or cytoplasmic protein aggregates precedes inclusion body formation. Molecular Cell. 17 (3), 351-365 (2005).
- Tam, S., et al. The chaperonin TRiC blocks a huntingtin sequence element that promotes the conformational switch to aggregation. Nature Structural & Molecular Biology. 16 (12), 1279-1285 (2009).
- Nekooki-Machida, Y., et al. Distinct conformations of in vitro and in vivo amyloids of huntingtin-exon1 show different cytotoxicity. Proceedings of the National Academy of Sciences U S A. 106 (24), 9679-9684 (2009).
- Wacker, J. L., Zareie, M. H., Fong, H., Sarikaya, M., Muchowski, P. J. Hsp70 and Hsp40 attenuate formation of spherical and annular polyglutamine oligomers by partitioning monomer. Nature Structural & Molecular Biology. 11 (12), 1215-1222 (2004).
- Legleiter, J., et al. Monoclonal antibodies recognize distinct conformational epitopes formed by polyglutamine in a mutant huntingtin fragment. Journal of Biological Chemistry. 284 (32), 21647-21658 (2009).
- Legleiter, J., et al. Mutant huntingtin fragments form oligomers in a polyglutamine length-dependent manner in vitro and in vivo. Journal of Biological Chemistry. 285 (19), 14777-14790 (2010).
- Nucifora, L. G., et al. Identification of novel potentially toxic oligomers formed in vitro. from mammalian-derived expanded huntingtin exon-1 protein. Journal of Biological Chemistry. 287 (19), 16017-16028 (2012).
- Dahlgren, P. R., et al. Atomic force microscopy analysis of the Huntington protein nanofibril formation. Nanomedicine. 1 (1), 52-57 (2005).
- Poirier, M. A., et al. Huntingtin spheroids and protofibrils as precursors in polyglutamine fibrilization. Journal of Biological Chemistry. 277 (43), 41032-41037 (2002).
- Duim, W. C., Chen, B., Frydman, J., Moerner, W. E. Sub-diffraction imaging of huntingtin protein aggregates by fluorescence blink-microscopy and atomic force microscopy. Chemphyschem. 12 (13), 2387-2390 (2011).
- Pieri, L., Madiona, K., Bousset, L., Melki, R. Fibrillar alpha-synuclein and huntingtin exon 1 assemblies are toxic to the cells. Biophysical Journal. 102 (12), 2894-2905 (2012).
- Monsellier, E., Redeker, V., Ruiz-Arlandis, G., Bousset, L., Melki, R. Molecular interaction between the chaperone Hsc70 and the N-terminal flank of huntingtin exon 1 modulates aggregation. Journal of Biological Chemistry. 290 (5), 2560-2576 (2015).
- Isas, J. M., Langen, R., Siemer, A. B. Solid-State Nuclear Magnetic Resonance on the Static and Dynamic Domains of Huntingtin Exon-1 Fibrils. Biochemistry. 54 (25), 3942-3949 (2015).
- Malakhov, M. P., et al. SUMO fusions and SUMO-specific protease for efficient expression and purification of proteins. Journal of Structural Function Genomics. 5 (1-2), 75-86 (2004).
- Mossessova, E., Lima, C. D. Ulp1-SUMO crystal structure and genetic analysis reveal conserved interactions and a regulatory element essential for cell growth in yeast. Molecular Cell. 5 (5), 865-876 (2000).
- Kumari, S., Pal, R. K., Gupta, R., Goel, M. High Resolution X-ray Diffraction Dataset for Bacillus licheniformis Gamma Glutamyl Transpeptidase-acivicin complex: SUMO-Tag Renders High Expression and Solubility. Protein Journakl. 36 (1), 7-16 (2017).
- Zhang, J., Sun, A., Dong, Y., Wei, D. Recombinant Production and Characterization of SAC, the Core Domain of Par-4, by SUMO Fusion System. Applied Biochemistry and Biotechnology. , (2017).
- Reif, A., et al. Semisynthesis of biologically active glycoforms of the human cytokine interleukin 6. Angewandte Chemie International Edition English. 53 (45), 12125-12131 (2014).
- Froger, A., Hall, J. E. Transformation of plasmid DNA into E. coli using the heat shock method. Journal of Visualized Experiments. (6), 253 (2007).
- Smith, B. J. SDS Polyacrylamide Gel Electrophoresis of Proteins. Methods in Molecular Biology. 1, 41-55 (1984).
- Lawrence, A. M., Besir, H. U. Staining of proteins in gels with Coomassie G-250 without organic solvent and acetic acid. Journal of Visualized Experiments. (30), (2009).
- Block, H., et al. Immobilized-metal affinity chromatography (IMAC): a review. Methods in Enzymology. 463, 439-473 (2009).
- Chen, S. M., Wetzel, R. Solubilization and disaggregation of polyglutamine peptides. Protein Science. 10 (4), 887-891 (2001).
- Booth, D. S., Avila-Sakar, A., Cheng, Y. Visualizing proteins and macromolecular complexes by negative stain EM: from grid preparation to image acquisition. Journal of Visualized Experiments. (58), (2011).
- O'Nuallain, B., et al. Kinetics and thermodynamics of amyloid assembly using a high-performance liquid chromatography-based sedimentation assay. Amyloid, Prions, and Other Protein Aggregates, Pt C. 413, 34-74 (2006).
- Greenfield, N. J. Analysis of circular dichroism data. Methods in Enzymology. 383, 282-317 (2004).
- Aiken, C. T., et al. Phosphorylation of threonine 3: implications for Huntingtin aggregation and neurotoxicity. Journal of Biological Chemistry. 284 (43), 29427-29436 (2009).
- Satakarni, M., Curtis, R. Production of recombinant peptides as fusions with SUMO. Protein Expression and Purification. 78 (2), 113-119 (2011).
- Davies, H. A., Wilkinson, M. C., Gibson, R. P., Middleton, D. A. Expression and purification of the aortic amyloid polypeptide medin. Protein Expression and Purification. 98, 32-37 (2014).
- Chen, S., Wetzel, R. Solubilization and disaggregation of polyglutamine peptides. Protein Science. 10 (4), 887-891 (2001).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved