
Method Article
Conception expérimentale à l’intérieur du sujet à l’aide d’une tâche d’emplacement d’objet chez le rat
* Ces auteurs ont contribué à parts égales
Dans cet article
Résumé
Ce protocole fournit des étapes détaillées pour une tâche de localisation d’objet avec quatre répétitions utilisant la même cohorte de rats. Un encodage faible et fort peut produire des souvenirs à court et à long terme. La flexibilité du protocole avec répétition peut être bénéfique pour les études impliquant des opérations chirurgicales en économisant du temps et de la main-d’œuvre.
Résumé
La reconnaissance de la place des objets est une méthode importante utilisée pour étudier la mémoire spatiale chez les rongeurs. Cette mémoire de reconnaissance de lieu d’objet constitue la base de la tâche d’emplacement d’objet. Cet article fournit un protocole complet pour guider l’établissement d’une tâche de localisation d’objet avec l’option de jusqu’à quatre répétitions utilisant la même cohorte de rats. Les protocoles d’encodage faibles et forts peuvent être utilisés pour étudier les mémoires spatiales à court et à long terme de force variable et pour permettre la mise en œuvre de manipulations pertinentes inhibant ou améliorant la mémoire. De plus, la répétition du test avec le contrepoids présenté ici permet la combinaison des résultats de deux tests ou plus pour la comparaison à l’intérieur du sujet afin de réduire la variabilité entre les rats. Cette méthode aide à augmenter la puissance statistique et est fortement recommandée, en particulier lors de l’exécution d’expériences qui produisent une forte variation dans le comportement individuel. Cela affine directement l’étude en augmentant les données obtenues auprès de chaque animal et en réduisant le nombre total d’animaux nécessaires. Enfin, la mise en œuvre de la tâche répétée de localisation d’objets augmente l’efficacité des études impliquant des procédures chirurgicales en économisant du temps et de la main-d’œuvre.
Introduction
Les tâches de reconnaissance spontanée(p. ex., reconnaissance d’objets, reconnaissance de lieux d’objets) ont été utilisées dans une large mesure dans l’étude de la mémoire chez les rongeurs. Ces tests sont différents de la variété de tests utilisés pour évaluer la mémoire qui sont basés sur le conditionnement de la peur ou la motivation de récompense, en ce sens que les tâches de reconnaissance spontanée sont basées uniquement sur un comportement exploratoire spontané envers de nouveaux stimuli. Ce comportement, appelé « préférence néotique »1,est inhérent aux rongeurs ainsi qu’à d’autres espèces de mammifères et à certains non-mammifères tels que les oiseaux et les poissons2. La reconnaissance de place d’objet, qui dépend de la mémoire spatiale, peut être observée à l’aide de la tâche d’emplacement d’objet (également appelée tâche de reconnaissance d’objet spatial)3. Des études de lésions ont montré que la reconnaissance du lieu de l’objet nécessite un hippocampe intact4,5. En raison du protocole d’entraînement relativement simple et de l’absence de tout renforcement, cette tâche est préférable dans de nombreuses études. L’absence de renforcement positif et négatif minimise les paramètres supplémentaires et les régions du cerveau qui pourraient conduire le comportement. Par conséquent, le comportement ici est neutre et est basé sur la curiosité et la mémoire spatiale, permettant l’étude des mécanismes impliqués dans l’encodage, la consolidation et la récupération de la mémoire spatiale.
Le protocole pour la tâche de localisation d’objet se compose généralement de sessions d’accoutumance suivies d’une seule session d’encodage et d’essais de test, séparées par une période de retard, qui varie de plusieurs minutes à quelques heures. Il est fortement recommandé de manipuler les rats au préalable pour minimiser le niveau de stress des animaux et, par conséquent, les comportements qui pourraient affecter la mémoire de reconnaissance, tels que l’aversion pour la nouveauté. De même, un protocole d’accoutumant bien conçu joue un rôle essentiel dans la prévention du stress qui pourrait entraver le comportement naturel du rat pendant la tâche. Cependant, l’étendue de la manipulation et de l’accoutumant varie largement entre les laboratoires et les expérimentateurs, ce qui peut contribuer à une faible reproductibilité6,7,8. Dans l’essai d’encodage, le rat a le temps d’explorer une arène avec deux objets identiques situés dans deux coins désignés. Dans l’essai test, qui est retardé d’une période, le rat a le temps d’explorer l’arène avec la même paire d’objets, mais cette fois l’un d’eux a été déplacé vers un nouvel endroit. La préférence spontanée manifestée par les rats et l’augmentation du temps passé à explorer l’objet à l’emplacement nouveau qui en résulte sont révélatrices de la reconnaissance spatiale et de la mémoire des emplacements de l’objet3. La modification de l’essai d’encodage (durée et nombre de répétitions) influence la force de la mémoire.
Selon l’objectif de l’étude, la durée du délai entre l’encodage et les essais peut être modifiée pour modéliser la mémoire à court terme indépendante de la synthèse des protéines ou la mémoire à long terme dépendante de la synthèse des protéines. Par conséquent, la tâche de localisation de l’objet peut être utilisée pour une grande variété d’études en adaptant le protocole au besoin. En outre, la mise en œuvre de manipulations expérimentales, telles que des interventions pharmacologiques et optogénétiques, est également possible entre ces essais, tout comme l’imagerie in vivo. Il existe plusieurs études9,10 qui rapportent des itérations répétées de la tâche de localisation de l’objet au sein de la même cohorte de rats. Cela contraste avec l’utilisation traditionnelle dans laquelle un animal a une séance sans répétitions. Cependant, l’efficacité de ces paradigmes n’a pas été étudiée de manière approfondie, et il n’existe aucun document de méthode les décrivant. À notre connaissance, il s’agit de la première description rapportée d’un protocole qui décrit en détail une tâche de localisation d’objet avec jusqu’à quatre répétitions utilisant la même cohorte de rats, qui compare également systématiquement les résultats de chaque répétition. Les répétitions peuvent être utilisées pour contrebalancer les conditions expérimentales afin de permettre une comparaison à l’intérieur du sujet avec une variabilité réduite entre les tests. La répétition fiable de la tâche permet de mettre en commun les données, ce qui signifie qu’une quantité suffisante de données peut être générée à l’aide d’un nombre relativement faible de rats. Enfin, les répétitions utilisant le même rat peuvent être bénéfiques dans les expériences impliquant des opérations chirurgicales et des implantations en réduisant le nombre de rats requis qui, par conséquent, permet d’économiser du temps et des coûts de main-d’œuvre.
Cette étude présente un protocole détaillé détaillant comment effectuer une tâche de localisation d’objet chez des rats adultes en utilisant des essais de codage forts et faibles suivis d’essais avec des retards de 1 h et 24 heures. Le protocole d’encodage puissant produit une mémoire de reconnaissance statistiquement significative lorsqu’il est testé avec des retards de 1 h et 24 heures et peut donc être utilisé pour étudier les souvenirs à court et à long terme lors de la mise en œuvre de manipulations pour inhiber ces souvenirs11. En revanche, le protocole d’encodage faible ne produit une mémoire à court terme significative que lorsqu’il est testé avec un délai de 1 h. L’absence de mémoire à long terme peut être utilisée pour étudier les manipulations visant à améliorer la rétention de la mémoire11,12. Ce protocole inclut également des sessions de manipulation et d’accoutumage détaillées, qui visent à augmenter la reproductibilité de la tâche d’emplacement de l’objet. Cet article démontre également la répétition de la tâche dans quatre contextes distincts avec la même cohorte de rats en utilisant le protocole de codage faible, qui est confirmé pour produire des résultats reproductibles et cohérents à chaque fois.
Protocole
Toutes les méthodes décrites ici ont été approuvées par les autorités nationales danoises (numéro de licence: 2018-15-0201-01405) conformément à la législation danoise et européenne sur le bien-être animal.
1. Mise en place expérimentale et préparation de contextes distincts
- Arène d’emplacement d’objet avec contexte
REMARQUE: La configuration ci-dessous est illustrée dans une boîte insonorisée fermée (Figure 1B) avec la source lumineuse située le long des bords du plafond et la caméra située au centre du plafond de la boîte. L’arène, de 60 cm x 60 cm avec des murs de 100 cm de haut(Figure 1B),est placée à l’intérieur de la boîte et est entièrement isolée de la pièce environnante. Tous les indices spatiaux sont à l’intérieur de l’arène. Cela simplifie le processus de création de contextes distincts. Un niveau similaire d’isolement de la pièce environnante peut être atteint en enfermant une arène normale en plein champ avec un rideau uniforme autour des murs.- Obtenez une arène carrée en plastique dur opaque et non poreux d’une largeur minimale de 60 cm et d’une hauteur minimale de 50 cm. Choisissez une couleur pour le sol contrastant avec la couleur du rat pour un enregistrement réussi des mouvements du rat par le logiciel automatisé (le cas échéant). Placez l’aréna soit à l’intérieur d’une boîte(Figure 1B),soit sur une plate-forme fermée par un rideau.
- Pour créer un contexte, procurez-vous une deuxième couche de murs insérables(p. ex., un revêtement mural fait du même matériau que l’arène ou du papier peint en plastique qui peut être facilement nettoyé) de différentes couleurs et/ou motifs(p. ex., noir, blanc, rayures ou points). Insérez la deuxième couche de murs dans l’arène de manière à ce qu’ils soient distincts les uns des autres.
- Obtenez des repères spatiaux tridimensionnels (3D) (1-2 par contexte) avec des dimensions variant entre 10 cm x 10 cm x 5 cm et 20 cm x 15 cm x 15 cm (largeur x longueur x hauteur) et avoir (i) des formes géométriques distinctes et (ii) des couleurs qui contrastent avec la couleur du mur. Accrochez-les aux murs assez haut pour que les rats ne puissent pas atteindre ces indices.
- Obtenez différentes paires d’objets (autant que le numéro de contexte) qui sont non poreux, non à croquer et faciles à nettoyer. Essayez d’avoir des formes géométriques et des textures distinctes pour chaque nouvel objet. Choisissez des objets de 5 à 15 cm de largeur et de hauteur (évitez les objets plus hauts). Voir la figure 1D pour des exemples de quatre objets distincts (cônes, ballons de football, prismes rectangulaires et prismes triangulaires).
REMARQUE: Chaque objet doit présenter un intérêt similaire à celui des rats, de sorte que les temps d’exploration totaux pour tous les objets soient comparables. - Trouvez la meilleure solution pour fixer les objets sur le sol de l’arène(par exemple, en utilisant des tapis collants, du ruban adhésif double face, en attachant une plaque métallique sous l’objet et un aimant d’appariement sous l’arène, etc.).
- Lors de la création d’un autre contexte, recréant les murs de manière à ce qu’ils contrastent la distribution de la couleur et du motif des murs par rapport au(x) contexte(s) précédent(s). Utilisez de nouveaux repères spatiaux 3D qui sont différents de tous les repères précédents et contrastent avec eux. Voir la figure 1C pour des exemples de quatre contextes distincts.
- Obtenez une source de lumière qui assurera un éclairage diffus et égal dans l’arène et qui a une option de gradation. Ajustez l’intensité lumineuse à environ 100-120 lux aux coins de l’arène après avoir créé chaque contexte. Procurez-vous une caméra et placez-la au centre du plafond de la boîte.
REMARQUE: L’intensité lumineuse peut être ajustée à un niveau inférieur si le logiciel de notation automatisé n’est pas utilisé.
- Compartiment d’objets
- Procurez-vous un seau (>50 cm de diamètre). Ne choisissez pas une forme carrée pour éviter toute ressemblance avec l’arène expérimentale. Remplissez-le de matériel de literie.
- Obtenez 5 à 10 objets de formes et de tailles différentes (différents des objets qui vont être utilisés dans l’expérience) et placez-les tous au hasard dans le compartiment (Figure 1A).

Figure 1: La configuration expérimentale, comprenant quatre contextes et objets distincts. (A) Compartiment d’objets pour l’accoutumage d’objets. (B) L’appareil expérimental (à gauche), enfermant l’arène de localisation de l’objet, la caméra et la source lumineuse. La boîte expérimentale et l’arène avant la configuration du contexte (au milieu) et l’arène avec la configuration du contexte (à droite). (C) Quatre contextes (1-4) avec des couleurs et des motifs muraux distincts, ainsi que des repères spatiaux tridimensionnels. (D) Quatre objets utilisés dans les contextes 1 à 4, respectivement. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
- Caméra et logiciel de suivi (en option)
- Procurez-vous un logiciel qui peut être utilisé pour contrôler à distance l’enregistreur de la caméra et qui peut suivre les nez de rat. Effectuez les ajustements logiciels pour chaque contexte spécifique et chaque souche de rat avant chaque expérience.
- Contrepoids des emplacements d’objets et des groupes expérimentaux
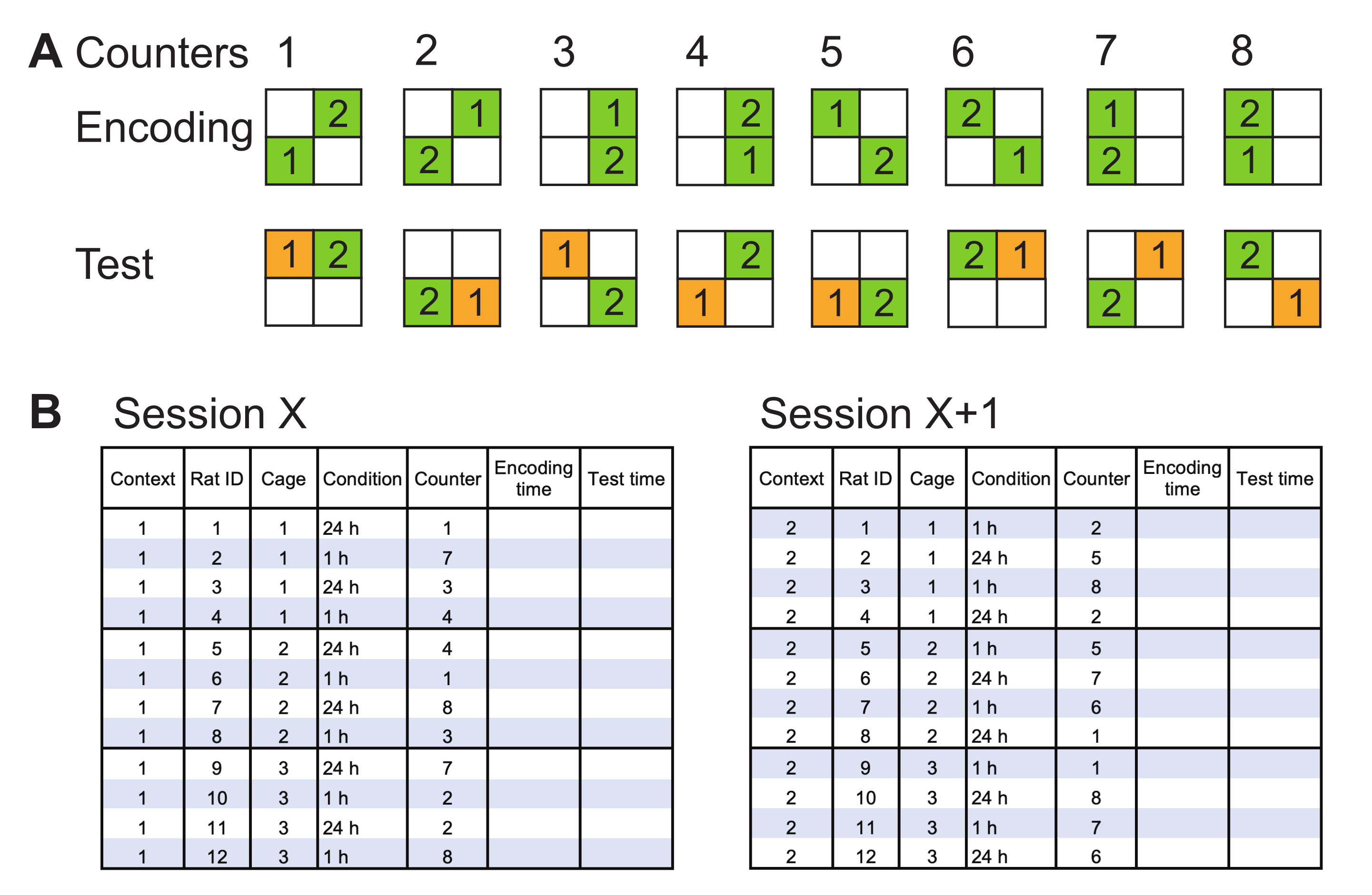
- Préparez les combinaisons possibles d’emplacements d’objets pour l’encodage et les essais de test, et nommez-les comme compteurs. Créez les combinaisons de manière à couvrir tous les coins en tant qu’emplacements d’objet et le mouvement de l’objet des coins adjacents aux coins diagonals et vice versa (Figure 2A).
- Préparez un calendrier pour l’expérience spécifique, en faisant correspondre chaque rat d’un groupe expérimental avec un compteur. Utilisez chaque paire des deux compteurs appariés (Figure 2A) au sein d’un groupe, s’il y a suffisamment de rats. Utilisez le même ensemble de compteurs pour les deux groupes expérimentaux en une seule session d’encodage/test (Figure 2B). Réaffecter des compteurs pour les sessions suivantes(c.-à-d. chaque nouveau contexte).
REMARQUE: Exécutez les rats dans un ordre mixte pendant les sessions d’encodage / test(par exemple,ne faites pas courir tous les rats dans une cage l’un après l’autre; faites plutôt pivoter les cages pour assurer un environnement calme dans une cage de plus d’un rat). - Lorsque vous utilisez deux contextes ou plus pour contrebalancer les groupes expérimentaux(par exemple, des groupes de mémoire de 1 h par rapport à des groupes de mémoire de 24 heures), assignez des rats à chaque groupe et modifiez les groupes dans les contextes suivants (Figure 2B).

Figure 2: Méthodes représentatives de contrepoids. (A) Les orientations possibles des objets dans l’arène lors des essais d’encodage et d’essai sont appelées compteurs. L’objet 1 est toujours l’objet en mouvement. Tous les deux compteurs sont contrebalancés de telle sorte que l’emplacement de l’objet en mouvement change. Chaque coin est occupé deux fois, et l’objet 1 est déplacé de la diagonale vers l’adjacent et vice versa pour un nombre égal de fois. (B) Exemple de calendrier d’encodage/test pour deux sessions contrebalancées(par exemple, les contextes 1 et 2). Les rats sont assignés à des conditions expérimentales dans le contexte 1 (session X, à gauche). Un ensemble de paires de compteurs(c.-à-d. 1-2, 3-4, 5-6 et 7-8) est sélectionné et attribué à chaque rat dans un groupe expérimental. Le même ensemble de compteurs est attribué aux rats dans les deux groupes expérimentaux. Dans la session suivante dans le contexte 2 (session X + 1; à droite), les rats des groupes expérimentaux sont changés pour le contrepoids, et un nouvel ensemble de paires de compteurs est attribué. L’heure au début de l’encodage et des essais d’essai doit être notée. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
REMARQUE: Toutes les sessions de manipulation, d’accoutumation et d’encodage / test de ce protocole ont été optimisées pendant la phase de lumière d’un cycle lumière/obscurité de 12 heures, et il est donc recommandé d’effectuer des expériences pendant la phase de lumière.
2. Manipulation et accoutumant
- Commencez à manipuler les rats en commençant soit par le sevrage (si les rats sont élevés dans l’installation d’origine), soit 2 à 3 semaines avant le début des expériences (dans le cas où les rats sont commandés dans une installation externe, après leur avoir permis de s’acclimater pendant une semaine après leur arrivée).
- Passez au moins 10 à 15 minutes sur chaque cage de 4 rats pendant 2 ou 3 jours par semaine jusqu’à ce que les rats soient à l’aise d’être touchés et ramassés par l’expérimentateur. Ajustez le temps alloué par cage en fonction du nombre de rats dans une cage.
REMARQUE: Il est important que tous les expérimentateurs qui s’attendent à travailler avec les rats soient présents lors de la manipulation. - Dans les cas où la manipulation commence au sevrage, réduisez la manipulation au minimum (facultatif) une fois ce niveau atteint. Si vous commencez 2-3 semaines avant les expériences, continuez la manipulation jusqu’au début des séances d’accoutumant.
- Amenez les rats dans leurs cages à la salle d’expérimentation pour habituer les rats au transport ainsi qu’à la salle d’expérimentation. Laissez les rats s’asseoir pendant au moins 30 minutes pour leur donner le temps de se calmer et de s’habituer. Passé ce temps, retournez les rats/cages dans la salle d’habitation.
REMARQUE: L’étape 2.4 peut être combinée avec la manipulation et répétée autant de fois que nécessaire. Une accoutumabilité supplémentaire peut être mise en œuvre à cette étape si le protocole comprend d’autres manipulations(par exemple, la manipulation pour la procédure d’injections, etc.). - Effectuer l’accoutumabilité des objets pour habituer les rats à interagir avec les objets et pour réduire les niveaux de stress généraux découlant de l’expérience de nouveaux environnements.
- Pour la séance 1, apportez toutes les cages de la maison dans la salle d’expérimentation et laissez les rats s’habituer à la pièce et installez-vous pendant au moins 30 minutes. Mettez les rats (2-4 rats) de la même cage ensemble dans le seau pendant 20 min. Nettoyez le seau en enlevant toute matière fécale entre chaque groupe de rats. Répétez la procédure pour toutes les cages. Mettez tous les rats dans leurs cages et retournez dans la pièce d’habitation.
- Pour la séance 2, un jour séparé, apportez toutes les cages dans la salle d’expérimentation et laissez agir pendant au moins 30 minutes. Mettez chaque rat individuellement dans le seau pendant 10 min. Replacez le rat dans la cage de la maison et nettoyez le seau après chaque rat. Retournez toutes les cages dans la salle de logement.
- Pour la session 3, répétez l’étape 2.5.2 un jour distinct.
- Si l’appareil expérimental est une boîte fermée(figure 1B),choisissez d’effectuer une accoutumante de boîte vide pour habituer les rats au nouvel appareil expérimental. Dans la session 4, apportez toutes les cages dans la salle d’expérimentation et laissez pendant au moins 30 minutes. Placez les rats de la même cage ensemble (2-4 rats) dans l’arène vide sans contexte ni repère spatial(Figure 1B,milieu) pendant 20 min. Replacez tous les rats dans la cage de la maison et essuyez l’arène avec 70% d’éthanol après chaque groupe de rats.
REMARQUE : Les étapes 2.5 et 2.6 doivent être effectuées en une seule semaine, précédant la semaine d’accoutumage du contexte (étape 2.7 ; voir la figure 3). Une pause de quelques jours pendant ces étapes est acceptable. Toutefois, après avoir commencé l’étape 2.7, chaque étape doit être effectuée sur des jours consécutifs comme spécifié, jusqu’à la fin de l’essai (étape 2.9). - Effectuer une accoutumabilité contextuelle pour habituer les rats au contexte et aux indices 3D, pour réduire les niveaux de stress généraux et pour soutenir l’apprentissage spatial de l’environnement.
- Modifiez l’arène vide pour créer le premier contexte comme décrit à la section 1.1, mais ne placez pas les objets dans l’arène. Préparez l’appareil de contrôle.
- Pour la séance 1, apportez toutes les cages dans la salle d’expérimentation et laissez agir pendant au moins 30 minutes. Démarrez l’enregistreur si vous le faites manuellement. Placez le premier rat au centre de l’arène et laissez le rat explorer l’arène pendant 10 minutes. Ensuite, arrêtez l’enregistreur (s’il est manuel) et replacez le rat dans la cage de la maison. Essuyez soigneusement l’arène avec 70% d’éthanol après chaque rat et retournez toutes les cages dans la salle de logement une fois terminée.
- Pour les séances 2 et 3, répétez l’étape 2.7.2 pour chaque rat sur deux jours consécutifs de sorte qu’il y ait 3 séances d’accoutumabilité du contexte par rat au total.
REMARQUE: Envisagez de mélanger l’ordre dans lequel les rats vont dans l’arène, en particulier lorsqu’il s’agit d’un grand groupe. Cela évite de faire courir des rats spécifiques à plusieurs reprises à la même heure de la journée.

Figure 3: La conception de l’expérience comportementale, y compris les protocoles de manipulation, d’accoutumabilité et de localisation des objets. Les rats doivent être manipulés régulièrement à partir de quelques semaines avant la semaine d’accoutumant. Au cours de la semaine 0, les habitudes d’objets et de boîtes expérimentales sont effectuées sur 4 séances avec des intervalles d’au moins 24 heures entre les deux. Au cours de la semaine 1, l’accoutumation du contexte est effectuée sur 3 sessions consécutives avec des intervalles de 24 heures entre les deux, suivies d’essais d’encodage et de test. Il devrait y avoir un intervalle minimum de 48 h et jusqu’à 1 semaine avant de procéder à la séance suivante(p. ex., commencer l’accoutumation au contexte suivant à la semaine 2 ou 3). Abréviation : Hab., accoutumant. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
- Essai d’encodage (session 4)
REMARQUE : Dans le cas de manipulations pharmacologiques, un délai raisonnable pour administrer un agent peut être avant ou immédiatement après le ou les essais de codage et/ou avant l’essai d’essai, selon la nature de l’agent pharmacologique.- Apportez toutes les cages dans la salle d’expérimentation et laissez agir pendant au moins 30 minutes. À l’aide de l’horaire établi à l’avance(Figure 2B),placez la première paire d’objets identiques aux endroits désignés (à 2 coins et à une distance de >10 cm de chaque mur respectif; un morceau de carton en forme de L peut être utilisé pour maintenir la même distance à chaque fois) en utilisant des tapis collants ou du ruban adhésif double face.
- Démarrez l’enregistreur (s’il est manuel). Placez le premier rat dans l’arène face à un mur ou à un coin qui n’est occupé par aucun objet (distance égale à chaque objet).
REMARQUE: Suivez les étapes ci-dessous pour un encodage faible ou fort. - Pour un encodage faible (1 essai),laissez le rat explorer l’arène et les objets pendant 20 min. Ensuite, arrêtez l’enregistreur (s’il est manuel) et replacez le rat dans la cage de la maison. Retirez les objets et essuyez soigneusement les objets et l’arène avec de l’éthanol à 70%.
- Répétez l’étape 2.8.3 pour tous les rats afin que chaque rat reçoive 1 essai de codage de 20 min.
- Pour un encodage fort (3 essais),laissez le rat explorer l’arène et les objets pendant 5 min. Ensuite, arrêtez l’enregistreur (s’il est manuel) et replacez le rat dans la cage de la maison. Ne supprimez pas les objets. Essuyez l’arène et les objets avec 70% d’éthanol.
- Répétez l’étape 2.8.5 deux fois de plus avec le même rat de sorte qu’il y ait 3 essais au total. Replacez le rat dans la cage de la maison lorsque le temps est passé. Retirez les objets pour un nettoyage en profondeur et essuyez les objets et l’arène avec de l’éthanol à 70%.
REMARQUE: L’intervalle entre les essais pour un rat doit être d’environ 1 à 2 minutes. - Répétez les étapes 2.8.5 à 2.8.6 pour chaque rat.
- Si le délai est inférieur à 24 h, gardez les cages dans la salle d’expérience jusqu’à l’essai. Si ce n’est pas le cas, retournez toutes les cages dans la salle d’habitation une fois terminées.
- Essai d’essai (session 4)
REMARQUE : La période de retard doit être comptée à partir du début de la version d’évaluation de l’encodage.- En cas de retard de 24 heures (ou de tout retard nécessitant que l’essai soit effectué le lendemain), apportez toutes les cages à la salle d’expérience, en laissant suffisamment de temps avant le premier test pour que les rats puissent être laissés pendant au moins 30 minutes. Selon le calendrier, placez les objets dans les emplacements désignés (l’un des objets à un nouvel emplacement).
- Quand il est temps, démarrez l’enregistreur (s’il est manuel). Placez le premier rat dans l’arène face à un mur ou à un coin qui n’est occupé par aucun objet (distance égale à chaque objet).
- Laissez le rat explorer l’arène et les objets pendant 5 min. Ensuite, arrêtez l’enregistreur (s’il est manuel). Replacez le rat dans la cage de la maison. Retirez les objets et essuyez soigneusement les objets et l’arène avec de l’éthanol à 70%.
- Répétez les étapes 2.9.2 à 2.9.3 pour chaque rat. Retournez toutes les cages dans la pièce d’habitation.
REMARQUE: Dans chaque session d’encodage/test suivante, démarrez le protocole d’accoutumière à partir de l’étape 2.7 (accoutumation contextuelle) après un intervalle d’au moins 48 h et jusqu’à 1 semaine.
3. Analyse des données
- Pour chaque rat, notez le temps d’exploration de chaque objet dans les essais d’encodage et de test à l’aide d’un logiciel conçu à cet effet ou à l’aide d’une configuration manuelle. Score des essais d’encodage pour toute la durée. Score des essais de test pendant 2 min pour la meilleure performance de discrimination3. Si vous utilisez la notation automatisée en ligne, exportez les données de notation à partir du logiciel.
- Comptez le temps d’exploration lorsque le rat est en contact avec l’objet, reniflant l’objet ou faisant face à l’objet à une distance inférieure à 2 cm. Inclure l’escalade et l’assise sur l’objet comme exploration à moins que l’attention du rat ne semble être ailleurs que l’objet(par exemple, détourner le regard de l’objet).
- Calculez le temps total d’exploration pour les deux objets pour chaque rat. Envisagez d’exclure de ce test tout rat dont le temps d’exploration total est inférieur à 10 s dans l’essai (pour une notation de 2 minutes), car cela peut refléter une exploration peu fiable.
- Calculez le pourcentage d’exploration pour chaque objet (équation 1) ou l’indice de discrimination (DI) pour chaque rat (équation 2), et calculez les valeurs moyennes pour les groupes.
 (1)
(1)
REMARQUE: Si % d’exploration est de 50 % ou si l’ID est de 0, cela signifie que la performance est au niveau du hasard et que le rat n’a aucune préférence pour l’un ou l’autre objet. Le pourcentage moyen d’exploration et d’ID pendant les essais de codage devrait être d’environ 50 % ou 0, respectivement. Tout rat présentant une préférence supérieure à [moyenne ± (2 × SD)] pour l’un ou l’autre objet de l’essai de codage doit être exclu de l’analyse du test respectif. Cela permet une interprétation fiable de la préférence dans l’essai de test en tant que mémoire de l’emplacement de l’objet stable. Cette valeur peut être calculée pour un test individuel ou pour des données de codage combinées provenant de plusieurs tests. - Analysez les données par la méthode qui convient le mieux à la configuration expérimentale. Utilisez un test tà un échantillon pour détecter une préférence significative au-dessus du niveau de chance.
- Tout en utilisant plus d’un contexte avec contrepoids, combinez les résultats de la même condition expérimentale dans tous les contextes.
REMARQUE: Il en résultera des groupes composés des mêmes rats, ce qui permettra une comparaison à l’intérieur du sujet à l’aide d’un test tapparié pour deux groupes et en utilisant des mesures répétées d’analyse de variance (ANOVA) pour plus de deux groupes.
Résultats
Voici les résultats représentatifs des protocoles de codage fort et faible décrits à l’aide de rats transgéniques tyrosine hydroxylase (Th)-Cre mâles13 avec une souche Long-Evans croisée quatre fois avec une souche Lister Hooded et des rats Lister Hooded de type sauvage. Des rats transgéniques Th-Cre ont été utilisés car cette lignée de rats sera utilisée dans de futures études impliquant l’optogénétique. En utilisant chaque protocole, la mémoire a été testée avec des délais de 1 et 24 h. Les tests à 1 h démontrent une mémoire à court terme, tandis que les tests de 24 heures démontrent une mémoire à long terme. La valeur d’exclusion pour la préférence de codage a été calculée comme décrit dans le protocole, en utilisant les données combinées de cinq tests (protocoles de codage forts et faibles) comme [50,8 % ± (2×10,8 %)]. Les rats qui avaient une préférence de codage au-dessus et en dessous de ces valeurs ont été exclus des analyses des tests respectifs.
Pour les expériences de codage fort, 16 rats ont été utilisés, et pour les expériences de codage faible, 19 rats ont été utilisés. Pendant les essais d’encodage fort (3 × 5 min d’encodage; Figure 4A), il n’y avait pas de préférence significative pour l’un ou l’autre objet (52,0 ± 1,9 %, n = 16, t15 = 1,1, p = 0,29 ; test tà un échantillon par rapport au niveau de hasard). Ce protocole de codage fort a conduit à une préférence pour l’objet à l’emplacement nouveau, comme le montre le pourcentage moyen d’exploration, qui était significativement plus élevé que le niveau de chance (50%) dans les tests avec des retards de 1 h et 24 heures (mémoire de 1 h, 77,9 ± 2,4%, n = 8, t7 = 11,8, p < 0,001; mémoire de 24 heures, 65,2 ± 5,3 %, n = 8, t7 = 2,8, p = 0,025; test tà un échantillon par rapport au niveau de chance). Il n’y avait pas de différence significative entre la mémoire de 1 h et 24 h (p = 0,056 ; test tde Welch non appaire).
Au cours des essais de codage faible (20 min d’encodage; résultats regroupés dans quatre contextes; Figure 4B), il n’y avait pas de préférence significative pour l’un ou l’autre objet (51,1 ± 1,0 %, n =66, t 65 = 1,2, p = 0,24 ; test tà un échantillon par rapport au niveau de hasard). Ce protocole de codage faible a produit une augmentation significative de la préférence pour l’objet à l’emplacement nouveau par rapport au niveau de hasard dans les tests avec un délai de 1 h, mais pas un délai de 24 heures (données combinées des quatre contextes; mémoire de 1 h, 66,7 ± 2,0%, n = 32, t31 = 8,2, p < 0,001; mémoire de 24 heures, 49,6 ± 2,6 %, n = 34, t33 = 0,16, p = 0,87; test tà un échantillon par rapport au niveau de chance). Il y avait une différence significative entre les performances dans les tests avec des retards de 1 h et 24 h (mémoire de 1 h : n = 32, mémoire de 24 h : n = 34, t61,5 = 5,2, p < 0,001 ; test tde Welch non appacré).
La mémoire au niveau du groupe n’a pas été observée dans le test de retard de 24 heures indexée par la performance au niveau du hasard, mais a montré des variations individuelles. Cette variation plus élevée pour les conditions faibles à sans mémoire(par exemple,test de 24 heures) a été couramment observée en raison d’une exploration plus aléatoire des objets. Par conséquent, il est important de ne pas interpréter les performances des rats individuellement. Au lieu de cela, la distribution des points de données individuels peut être utilisée avec la moyenne du groupe comme résultat fiable du test. Plus l’encodage est fort, plus le comportement des rats devient uniforme et moins le nombre de rats nécessaires pour atteindre la signification statistique est élevé, comme on peut l’observer à la figure 4A pour le protocole de codage fort. En revanche, des groupes plus importants sont nécessaires pour obtenir des résultats fiables pour les conditions faibles(Figure 4B).

Figure 4: Performances de la mémoire après codage fort et faible. (A) Essai de codage fort (3 × 5 min d’encodage) suivi d’essais de test de 1 h ou de 24 h. Il n’y avait pas de préférence significative pour l’un ou l’autre objet lors des essais de codage (n = 16). Le fort encodage a produit une préférence significativement accrue pour l’objet au nouvel emplacement dans les tests avec des retards de 1 h et de 24 heures par rapport au niveau de hasard (mémoire de 1 h et 24 heures: n = 8 dans chaque groupe). Il n’y avait pas de différence significative entre les groupes. (B) L’essai de codage faible (codage de 20 min) suivi d’essais de test de 1 h ou de 24 heures. Il n’y avait pas de préférence significative pour l’un ou l’autre objet en tant que groupe lors des essais de codage (n = 66). Le faible codage a produit une préférence significativement accrue pour l’objet à l’emplacement nouveau dans le test avec un délai de 1 h, mais pas de 24 heures, par rapport au niveau de hasard (mémoire de 1 h: n = 32; mémoire de 24 heures: n = 34). Il y avait une différence significative entre les performances dans les tests avec des retards de 1 h et de 24 heures. Les résultats ont été regroupés dans quatre contextes. Les points de données individuels sont présentés sous forme de points. Toutes les barres indiquent le pourcentage d’exploration de l’objet à un nouvel emplacement en moyenne ± SEM. *p < 0,05, ***p < 0,001; test tà un échantillon par rapport au niveau de hasard (50 %, ligne pointillée). ###p < 0,001; ns, non significatif; test tde Welch non appaéri. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Un avantage significatif de ce protocole établi est qu’il peut être effectué quatre fois en utilisant quatre contextes distincts(Figure 1C)avec la même cohorte de rats. Les résultats présentés à la figure 5 illustrent une façon possible d’utiliser le contrepoids avec deux groupes expérimentaux (groupes mémoire 1-h et 24-h). Les deux groupes ont été contrebalancés sur deux contextes (contextes 1 et 2), et cela a été répété dans deux contextes supplémentaires (contextes 3 et 4; Figure 5A). Les résultats des quatre contextes sont présentés individuellement à la figure 5B,D, où la mémoire de chaque groupe expérimental a été évaluée en comparant la préférence au niveau de hasard dans chaque contexte (mémoire de 1 h: contexte 1, 69,9 ± 3,6%, n = 9, t8 = 5,5, p < 0,001; Contexte 2, 65,6 ± 3,9 %, n = 9, t8 = 4,0, p = 0,004; Contexte 3, 65,2 ± 3,8 %, n = 7, t6 = 4,0, p = 0,007; Contexte 4, 65,3 ± 5,6 %, n = 7, t6 = 2,7, p = 0,035; Mémoire 24 heures sur 24 : Contexte 1, 45,1 ± 6,4 %, n = 9, t8 = 0,77, p = 0,46 ; Contexte 2, 49,1 ± 4,9 %, n = 9, t8 = 0,18, p = 0,86; Contexte 3, 57,2 ± 4,1 %, n = 8, t7 = 1,7, p = 0,12; Contexte 4, 47,6 ± 4,7 %, n = 8, t7 = 0,52, p = 0,62; test tà un échantillon par rapport au niveau de chance).
Dans les contextes 1, 2 et 4, la comparaison entre les sujets des groupes a révélé des différences significatives entre la mémoire de 1 h et de 24 h (mémoire de 1 h par rapport à la mémoire de 24 heures : contexte1, t 12,7 = 3,4, p = 0,005 ; Contexte 2, t15,2 = 2,6, p = 0,019; Contexte 3, t13,0 = 1,4, p = 0,17; Contexte 4, t12,2 = 2,4, p = 0,032; t-testde Welch non appairé). Pour une meilleure représentation et une comparaison des données à l’intérieur du sujet, les résultats de deux contextes contrebalancés ont été combinés (Figure 5C,E). Les groupes expérimentaux combinés ont été comparés au niveau de hasard individuellement (Contextes 1 et 2 combinés : mémoire 1 h, 67,8 ± 2,6 %, n = 18, t17 = 6,7, p < 0,001 ; mémoire 24 heures, 47,1 ± 3,9 %, n = 18, t17 = 0,74, p = 0,47 ; Contextes 3 et 4 combinés : 1 h de mémoire, 65,3 ± 3,3 %, n = 14, t13 = 4,7, p < 0,001 ; Mémoire 24 heures sur 24, 52,4 ± 3,2 %, n = 16, t15 = 0,73, p = 0,48 ; test tà un échantillon par rapport au niveau de chance). Ensuite, les groupes expérimentaux ont été comparés les uns aux autres.
Dans les deux paires de contextes, il y avait des différences significatives entre les groupes, comme le révèlent les comparaisons à l’intérieur du sujet (mémoire de 1 h par rapport à la mémoire de 24 heures : contextes 1 et 2 combinés, t16 = 3,5, p = 0,003 ; Contextes 3 et 4 combinés, t13 = 2,4, p = 0,032; couplé t-test). Des résultats comparables ont également été obtenus avec des rats à capuchon Lister de type sauvage dans le protocole de codage faible en utilisant les contextes 1 et 4 pour les deux sessions contrebalancées (données non présentées). La reproductibilité et la fiabilité des résultats ont été validées en comparant chaque ensemble de données à l’aide d’ANOVA unidirectionnelle. Aucune différence significative n’a été détectée entre les quatre contextes (mémoire 1 h : F3,28 = 0,31, p = 0,81 ; mémoire 24 heures : F3,30 = 0,99, p = 0,41). Par conséquent, le test de localisation de l’objet peut être répété de manière fiable avec une influence minimale des répétitions, étant donné que les instructions de ce protocole sont suivies.

Figure 5: Différentes façons de présenter et d’analyser les résultats du protocole de codage faible avec deux groupes expérimentaux contrebalancés sur deux sessions. (A) La conception expérimentale du contrepoids avec deux groupes expérimentaux (groupes de mémoire de 1 h et 24 heures) sur deux sessions (contextes 1 et 2). Le contrepoids a été répété lors de deux sessions supplémentaires (contextes 3 et 4). (B et D) Les résultats de chaque contexte et des groupes expérimentaux ont été comparés individuellement au niveau de hasard et les uns aux autres. Dans les quatre contextes, la préférence pour l’objet à l’emplacement nouveau dans les tests avec un délai de 1 h a été significativement augmentée par rapport au niveau de hasard [Contexte 1 et 2: n = 9 par groupe (B); Contexte 3 et 4 : n = 7 par groupe (D)]. Dans les tests de retard de 24 heures, la préférence pour l’objet à l’emplacement nouveau ne différait pas du hasard (Contexte 1 et 2: n = 9 par groupe; Contexte 3 et 4 : n = 8 par groupe). Il y avait une différence significative entre les préférences des groupes expérimentaux dans les contextes 1, 2 et 4, mais pas le contexte 3, comme le révèle la comparaison entre les sujets. *p < 0,05, **p < 0,01, ***p < 0,001; test tà un échantillon par rapport au niveau de hasard (50 %, ligne pointillée). #p < 0,05; ##p < 0,01; ns, non significatif; test tde Welch non appaéri. (C et E) Les résultats sont présentés après avoir combiné les groupes expérimentaux des deux contextes contrebalancés [Contextes 1 et 2 combinés, n = 17 par groupe(C); Contextes 3 et 4 combinés, n = 14 par groupe (E)]. La préférence pour l’objet à l’emplacement nouveau a été significativement augmentée par rapport au niveau de hasard dans les tests avec un délai de 1 h, mais pas de 24 heures, dans les deux paires de contextes. La comparaison des groupes expérimentaux à l’intérieur du sujet a révélé des différences significatives entre les préférences pour l’objet au nouvel emplacement dans les tests avec des retards de 1 h et de 24 heures dans les deux paires de contextes. p < 0,001; test tà un échantillon par rapport au niveau de hasard (50 %, ligne pointillée). #p < 0,05, ##p < 0,01; t-testapparié. Les points de données individuels sont présentés sous forme de points. Toutes les barres indiquent le pourcentage d’exploration de l’objet à l’emplacement du roman en tant que moyenne ± SEM. Veuillez cliquer ici pour voir une version plus grande de cette figure.
{kind=link}
Discussion
La tâche de localisation d’objet peut être utilisée dans diverses études pour étudier la mémoire spatiale comme décrit précédemment. La flexibilité de la configuration permet la modélisation de la mémoire à court et à long terme de différentes forces, et elle peut être facilement mise en œuvre à faible coût. Cependant, comme il existe de nombreux paramètres dans le protocole qui peuvent influencer les résultats, et que différentes études varient légèrement dans ces paramètres6, on peut rencontrer des difficultés à mettre en œuvre la tâche avec succès pour la première fois. Le protocole ci-dessus est destiné à guider les lecteurs tout au long de ce processus en douceur. D’autres étapes cruciales qui pourraient être importantes dans la mise en œuvre réussie de la tâche avec une grande reproductibilité seront discutées ci-dessous.
Bien que la session d’encodage / test soit souvent au centre de l’exécution d’expériences de localisation d’objets, les protocoles de manipulation et d’accoutumant ont un effet profond sur le résultat de ce type de tests comportementaux où le résultat dépend du comportement naturel non perturbé du rat14,15. En tant que telles, les étapes précédant la session d’encodage / test doivent être conçues avec prudence, car elles peuvent influencer le comportement et la mémoire du rat et, par conséquent, influencer les résultats finaux. Un bon niveau de manipulation et d’accoutumant de sorte que les rats se familiarisent avec l’expérimentateur et la tâche minimisera l’effet des facteurs de stress tout en augmentant la probabilité de présenter un comportement naturel8. Comme mentionné dans le protocole, la manipulation peut commencer dès le sevrage des petits si la souche de rat est maintenue dans l’établissement à domicile. Sur la base de l’expérience antérieure (données non présentées) et de celle de plusieurs études antérieures16,17, cette manipulation précoce entraîne une faible anxiété et une curiosité accrue dans les mois qui suivent.
Comme la tâche de localisation de l’objet dépend uniquement de l’entraînement exploratoire intrinsèque des rats, le comportement attendu peut être facilement entravé si les rats ne sont pas désireux d’explorer ou réticents à approcher la nouveauté, ce que l’on appelle le « comportement néophobe »1. En tant que tel, il est fortement recommandé d’inclure un protocole de manipulation et d’accoutumant approfondi en fonction des besoins spécifiques de l’étude. Ce protocole peut être utilisé comme guide des exigences minimales et d’autres étapes peuvent être mises en œuvre(par exemple,si l’étude doit inclure des injections à un stade ultérieur, une accoutumée aux procédures d’injection et une position de maintien spécifique sont requises). La souche et l’âge des rats expérimentaux sont deux autres facteurs influents et doivent être pris en compte avant de planifier une expérience pour éviter des résultats sous-optimaux. Différentes souches de rats peuvent avoir des comportements et des niveaux d’anxiété de basedifférents 18,19,20 et, par conséquent, un ajustement spécifique au protocole peut être nécessaire en fonction de la souche utilisée.
Il est confirmé que ce protocole fonctionne bien avec les rats transgéniques Th-Cre avec la souche Long-Evans croisée quatre fois avec la souche Lister Hooded et les rats Lister Hooded de type sauvage. Un âge de départ logiquement idéal pour les rats dans les expériences comportementales est d’environ 12 semaines20, mais la variabilité entre les souches et les exigences spécifiques de la tâche doivent être prises en compte. Il pourrait également être possible d’utiliser des rats en développement si cela présente un intérêt pour l’étude, bien que des ajustements au protocole puissent être nécessaires et ne soient pas couverts ici. Cependant, il est important de déterminer si le rat à un âge donné a développé les fonctions cognitives nécessaires pour effectuer avec succès cette tâche. Une étude21 portant sur ce sujet a rapporté que seuls les rats adolescents au 38e jour postnatal et non avant, ont montré une mémoire spatiale allocentrique reflétée dans la préférence pour l’objet au nouvel emplacement, comme observé chez les rats adultes. Le protocole présenté ici a été couronné de succès en utilisant des rats âgés de 15 à 16 semaines au début de la première session d’encodage / test. Auparavant, le même protocole d’encodage fort produisait des résultats sous-optimaux à négatifs lors de l’utilisation de rats âgés de 23 semaines qui n’avaient pas atteint le niveau optimal d’accoutumant en raison du manque de manipulation et d’accoutumant à un âge suffisamment jeune. Ces rats n’ont pas réussi à performer différemment du niveau du hasard ou, en fait, ont montré une aversion pour la nouveauté observée en termes de préférence pour les objets stables au lieu des objets déplacés (données non montrées). Ces résultats fournissent la preuve que l’âge et le moment de l’accoutumance à la manipulation peuvent avoir un impact sur l’efficacité de l’accoutumance et, par conséquent, contribuer à l’observation d’un comportement anxieux et néophobe dans les tests.
Ici, deux protocoles différents sont décrits, assurant un encodage fort ou faible dans la tâche d’emplacement de l’objet. Au cours de l’établissement de ces protocoles, il a été observé que l’intérêt pour les objets a diminué après 5 à 10 minutes d’exploration au cours d’essais longs uniques(par exemple, 20 minutes d’encodage), et les rats ont finalement cessé d’explorer. Il en résulte une mémoire plus faible des emplacements des objets. Un protocole d’encodage qui entrelace les essais de codage avec de courtes périodes de repos(par exemple, un codage de 3 x 5 minutes) surmonte ce problème et conduit à une exploration élevée tout au long des essais. Ainsi, le temps d’exploration actif et la disposition différente de ces deux protocoles d’encodage influencent la force de la mémoire, qui est plus forte après 3 x 5 min d’encodage qu’après des protocoles d’encodage de 20 min. Des résultats similaires peuvent également être obtenus en utilisant des durées légèrement différentes avec des protocoles d’essai uniques par rapport à des protocoles d’essai entrelacés, et des ajustements peuvent être apportés pour répondre aux besoins de l’étude et de la souche de rat.
Contrairement aux protocoles utilisant un champ ouvert blanc uni avec seulement des signaux externes dans la pièce, le protocole présenté ici utilise une arène avec des contextes distincts et des indices intra-labyrinthe qui nécessite probablement plus de temps pour apprendre. Par conséquent, l’ajout d’une étape d’accoutumabilité du contexte dans le protocole avant l’essai de codage est recommandé. Cela permettra aux rats de former une carte spatiale de chaque contexte pendant l’accoutumant et de réduire la durée de l’essai de codage suivant, car les rats n’auront qu’à coder les emplacements des objets par rapport à cette carte. De plus, l’accoutumation du contexte permettra aux rats de s’habituer à tout distracteur possible dans chaque contexte, comme les signaux spatiaux 3D, minimisant ainsi les comportements autres que l’exploration d’objets dans la session d’encodage / test à suivre. Avec la mise en œuvre d’une méthode de contrepoids approfondie composée de plusieurs niveaux(c’est-à-dire un large éventail de combinaisons d’emplacement d’objet (compteurs) et de direction du déplacement de l’objet), les préférences indésirables qui peuvent augmenter en raison des variations de l’intensité lumineuse et des couleurs / motifs muraux aux coins de l’arène sont minimisées.
Plusieurs facteurs doivent être pris en compte lors de la répétition de la tâche pour augmenter la reproductibilité entre les sessions d’encodage / test et minimiser l’influence de la répétition. Tout d’abord, des contextes distincts (jusqu’au nombre de répétitions de sessions d’encodage/test) doivent être conçus pour éviter l’accumulation de mémoire spatiale qui pourrait être causée par l’exécution des sessions répétées en utilisant le même contexte. Pour ce faire, un appareil avec des parois remplaçables de différentes couleurs et motifs a été utilisé (Figure 1B, C). Les murs distincts et les objets 3D (tels que des jouets ou de petits objets du quotidien de couleurs et de formes distinctes, voir protocole et figure 1C)accrochés aux murs sont les repères spatiaux et les points de repère que les rats utilisent potentiellement pour apprendre l’emplacement des objets par rapport à leurs contextes. Dans le cas où un test ne parvient pas à produire une préférence pour l’objet déplacé, la modification de ces paramètres du contexte (conception du mur et repères spatiaux) peut être envisagée. Alternativement, une arène de forme rectangulaire ou circulaire peut être utilisée pour des tâches de localisation d’objets au lieu d’une arène carrée comme dans ce protocole. Les arènes circulaires élimineraient les préférences de coin22 qui sont souvent observées dans les arènes avec des coins, et par conséquent, cela peut être bénéfique lorsqu’il s’agit d’une souche de rat ou de souris particulièrement anxieuse. Bien que les exigences de création de quatre contextes distincts dans ce protocole fonctionnent de manière optimale avec une forme quadrangulaire, une arène circulaire peut également être rendue fonctionnelle après quelques ajustements.
Deuxièmement, les intervalles entre chaque session d’encodage/test doivent être déterminés de manière à ce que les rats conservent le même niveau d’intérêt à chaque fois, tout en évitant le risque d’apprentissage cumulatif résultant d’un calendrier dense de répétitions. Habituellement, un intervalle d’au moins le double de la durée du délai entre l’encodage et les essais est suffisant, des intervalles plus longs étant plus favorables pour plus de deux répétitions. Cela signifie que, bien qu’un intervalle minimum de 48 heures après un test de 24 heures soit suffisant pour une ou deux répétitions, l’utilisation d’un intervalle de 1 semaine est recommandée pour quatre répétitions. Comme le montrent les résultats de la figure 5 et la comparaison à l’aide de l’ANOVA, la tâche peut être répétée avec succès quatre fois. Sur cette base, le protocole établi peut être utilisé pour contrebalancer jusqu’à quatre conditions expérimentales. Le nombre de groupes expérimentaux détermine le nombre de répétitions de sessions d’encodage/d’essai dans des contextes distincts. Les résultats de la figure 5 représentent une façon possible d’utiliser le protocole avec deux groupes expérimentaux. Les groupes ont été contrebalancés en deux sessions, et les mêmes conditions ont été répétées en deux sessions supplémentaires (à des fins de validation). La deuxième série de sessions contrebalancées pourrait également être utilisée pour contrebalancer les nouvelles conditions. De même, trois ou quatre conditions expérimentales peuvent être comparées à l’aide de trois ou quatre séances contrebalancées, respectivement.
Dans ces cas, les contextes doivent être conçus pour tenir compte des caractéristiques contrastées décrites dans le protocole. Il convient de noter que la conception contrebalancée peut ne pas convenir aux expériences dans lesquelles des manipulations supplémentaires, telles qu’une intervention pharmacologique qui pourrait laisser un effet ou des dommages durables, doivent être utilisées. Pour maintenir l’efficacité et la reproductibilité des essais, l’expérience doit être conçue en conséquence. Les données des tests répétés peuvent être présentées et analysées de plusieurs manières, comme le montre la figure 5. Pour une analyse initiale, les groupes expérimentaux dans chaque contexte peuvent être comparés individuellement au niveau de hasard à l’aide d’un test tà un échantillon pour déterminer toute préférence significative(Figure 5B,D). Cela peut être utile pour obtenir une compréhension rapide des données, mais cela ne garantit qu’une comparaison indirecte des groupes. Ainsi, pour comparer deux groupes ou plus, les données doivent être analysées à l’aide de tests tà deux échantillons (appariés ou non appariés) ou d’ANOVA, respectivement. Cela peut se faire sous la forme d’une comparaison entre les sujets des groupes au cours d’une seule session de codage/test(Figure 4A et Figure 5B,D)ou d’une comparaison à l’intérieur du sujet des groupes de deux (ou plusieurs) contextes contrebalancés(Figure 5C,E). Cette dernière méthode est fortement recommandée, en particulier lorsqu’il s’agit de conditions de mémoire faibles, ce qui, comme expliqué précédemment, entraîne une variance élevée en raison du comportement aléatoire.
La combinaison des contextes contrebalancés conduit à des groupes plus importants qui sont nécessaires pour visualiser de manière fiable le comportement du groupe avec une variation minimale. En utilisant un protocole avec des répétitions dans des sessions contrebalancées, on peut s’attendre à une diminution du nombre de rats à environ un tiers du nombre qui serait nécessaire en utilisant un seul test avec la même puissance statistique. Habituellement, la taille des échantillons dans une plage de 7 à 15 rats (total) pour les séances contrebalancées et dans une gamme de 20 à 50 rats (10 à 25 par groupe) pour une seule session avec une taille d’effet et une puissance supérieures à 0,8 sont suffisantes. La diminution du nombre d’animaux nécessaires et l’augmentation de l’information que nous obtenons de chaque animal utilisant ce protocole affinent l’étude et servent les principes 3R d’utilisations éthiques des animaux dans la recherche. Il est important à cette étape de garder à l’esprit que le comportement aléatoire du rat, qui n’est pas accompagné d’une forte mémoire, peut entraîner des préférences individuelles fortes à la fois inférieures et supérieures au hasard, mais la moyenne du groupe devrait donner une préférence qui n’est pas significativement différente du hasard. Les données individuelles doivent être interprétées avec prudence. La distribution des points de données individuels au sein d’un groupe peut également être informative pour interpréter les résultats. Comme le illustrent les figures 4 et 5,la distribution change en fonction de la force de la mémoire. Dans l’ensemble, le protocole présenté ici peut être suivi facilement pour implémenter la tâche de localisation de l’objet avec des répétitions pour modéliser la mémoire spatiale à court terme et / ou à long terme. Le protocole de formation simple et flexible et la possibilité de mettre en œuvre d’autres manipulations font de cette tâche un choix populaire. Ces modifications apportées au protocole permettent d’enquêter sur des étapes particulières telles que l’acquisition, la consolidation et le rappel de la mémoire.
Déclarations de divulgation
Les auteurs n’ont rien à divulguer.
Remerciements
Nous tenons à remercier Antonios Asiminas, Dorothy Tse, Kiichi O’Hara et David Bett pour leurs commentaires et suggestions perspicaces. Cette étude a été soutenue par Erasmus+ (à G.B. et L.N.); la Graduate School of Health, Université d’Aarhus (à K.H.); Novo Nordisk Foundation Young Investigator Award 2017 (NNF17OC0026774), Lundbeckfonden (DANDRITE-R248-2016-2518) et PROMEMO - Center for Proteins in Memory, un centre d’excellence financé par la Fondation nationale danoise de la recherche (DNRF133) (à T.T.).
matériels
| Name | Company | Catalog Number | Comments |
| Open-field/experimental box | O'Hara & Co (Japan) | OF-3001 | Open-field box for the object location task |
| Object 1: cones | O'Hara & Co (Japan) | ORO-RR | |
| Object 2: footballs | O'Hara & Co (Japan) | ORO-RB | |
| Object 3: rectangular blocks | O'Hara & Co (Japan) | ORO-RC | Rectangular blocks were modified after purchase |
| Object location task apparatus | O'Hara & Co (Japan) | SPP-4501 | Sound attenuating box that contains the open-field box for the object location task |
| Tracking software | O'Hara & Co (Japan) | TimeSSI | For movement tracking and automated camera functions |
| Wild-type Lister Hooded rats | Charles River | 603 |
Références
- Hughes, R. N. Neotic preferences in laboratory rodents: issues, assessment and substrates. Neuroscience and Biobehavioral Reviews. 31 (3), 441-464 (2007).
- Blaser, R., Heyser, C. Spontaneous object recognition: a promising approach to the comparative study of memory. Frontiers in Behavioral Neuroscience. 9, 183 (2015).
- Dix, S. L., Aggleton, J. P. Extending the spontaneous preference test of recognition: evidence of object-location and object-context recognition. Behavioral Brain Research. 99 (2), 191-200 (1999).
- Barker, G. R., Warburton, E. C. When is the hippocampus involved in recognition memory. Journal of Neuroscience. 31 (29), 10721-10731 (2011).
- Mumby, D. G., Gaskin, S., Glenn, M. J., Schramek, T. E., Lehmann, H. Hippocampal damage and exploratory preferences in rats: memory for objects, places, and contexts. Learning & Memory. 9 (2), 49-57 (2002).
- Gulinello, M., et al. Rigor and reproducibility in rodent behavioral research. Neurobiology of Learning and Memory. 165, 106780 (2019).
- Rudeck, J., Vogl, S., Banneke, S., Schonfelder, G., Lewejohann, L. Repeatability analysis improves the reliability of behavioral data. PLoS One. 15 (4), 0230900 (2020).
- Gouveia, K., Hurst, J. L. Optimising reliability of mouse performance in behavioural testing: the major role of non-aversive handling. Scientific Reports. 7, 44999 (2017).
- Migues, P. V., et al. Blocking synaptic removal of GluA2-containing AMPA receptors prevents the natural forgetting of long-term memories. Journal of Neuroscience. 36 (12), 3481-3494 (2016).
- Maingret, N., Girardeau, G., Todorova, R., Goutierre, M., Zugaro, M. Hippocampo-cortical coupling mediates memory consolidation during sleep. Nature Neuroscience. 19 (7), 959-964 (2016).
- Chao, O. Y., de Souza Silva, M. A., Yang, Y. M., Huston, J. P. The medial prefrontal cortex - hippocampus circuit that integrates information of object, place and time to construct episodic memory in rodents: Behavioral, anatomical and neurochemical properties. Neuroscience and Biobehavioral Reviews. 113, 373-407 (2020).
- Takeuchi, T., et al. Locus coeruleus and dopaminergic consolidation of everyday memory. Nature. 537 (7620), 357-362 (2016).
- Witten, I. B., et al. Recombinase-driver rat lines: tools, techniques, and optogenetic application to dopamine-mediated reinforcement. Neuron. 72 (5), 721-733 (2011).
- Costa, R., Tamascia, M. L., Nogueira, M. D., Casarini, D. E., Marcondes, F. K. Handling of adolescent rats improves learning and memory and decreases anxiety. Journal of the American Association for Labaratory Animal Science. 51 (5), 548-553 (2012).
- Schmitt, U., Hiemke, C. Strain differences in open-field and elevated plus-maze behavior of rats without and with pretest handling. Pharmacology Biochemistry and Behavior. 59 (4), 807-811 (1998).
- Kosten, T. A., Kim, J. J., Lee, H. J. Early life manipulations alter learning and memory in rats. Neuroscience and Biobehavioral Reviews. 36 (9), 1985-2006 (2012).
- Denenberg, V. H., Grota, L. J. Social-seeking and novelty-seeking behavior as a function of differential rearing histories. Journal of Abnormal and Social Psychology. 69 (4), 453-456 (1964).
- Clemens, L. E., Jansson, E. K., Portal, E., Riess, O., Nguyen, H. P. A behavioral comparison of the common laboratory rat strains Lister Hooded, Lewis, Fischer 344 and Wistar in an automated homecage system. Genes, Brain, and Behavior. 13 (3), 305-321 (2014).
- Ennaceur, A., Michalikova, S., Bradford, A., Ahmed, S. Detailed analysis of the behavior of Lister and Wistar rats in anxiety, object recognition and object location tasks. Behavioral Brain Research. 159 (2), 247-266 (2005).
- Deacon, R. M. Housing, husbandry and handling of rodents for behavioral experiments. Nature Protocols. 1 (2), 936-946 (2006).
- Contreras, M. P., Born, J., Inostroza, M. The expression of allocentric object-place recognition memory during development. Behavioral Brain Research. 372, 112013 (2019).
- Yaski, O., Eilam, D. How do global and local geometries shape exploratory behavior in rats. Behavioral Brain Research. 187 (2), 334-342 (2008).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.