
Method Article
Collecte de routine d’ensembles de données cryo-EM haute résolution à l’aide d’un microscope électronique à transmission de 200 KV
* Ces auteurs ont contribué à parts égales
Dans cet article
Résumé
Des cartes cryo-EM à haute résolution de macromolécules peuvent également être obtenues à l’aide de microscopes TEM de 200 kV. Ce protocole montre les meilleures pratiques pour définir des alignements optiques précis, des schémas d’acquisition de données et la sélection de zones d’imagerie qui sont tous essentiels à la collecte réussie d’ensembles de données haute résolution à l’aide d’un TEM de 200 kV.
Résumé
La cryo-microscopie électronique (cryo-EM) a été établie comme une méthode de routine pour la détermination de la structure des protéines au cours de la dernière décennie, prenant une part toujours croissante des données structurelles publiées. Les progrès récents de la technologie TEM et de l’automatisation ont augmenté à la fois la vitesse de collecte des données et la qualité des images acquises tout en diminuant simultanément le niveau d’expertise requis pour obtenir des cartes cryo-EM à des résolutions inférieures à 3 Å. Alors que la plupart de ces structures à haute résolution ont été obtenues à l’aide de systèmes cryo-TEM de 300 kV à la pointe de la technologie, des structures à haute résolution peuvent également être obtenues avec des systèmes cryo-TEM de 200 kV, en particulier lorsqu’elles sont équipées d’un filtre à énergie. De plus, l’automatisation des alignements de microscopes et de la collecte de données avec une évaluation de la qualité d’image en temps réel réduit la complexité du système et assure des réglages optimaux du microscope, ce qui se traduit par un rendement accru d’images de haute qualité et un débit global de collecte de données. Ce protocole démontre la mise en œuvre des avancées technologiques récentes et des fonctionnalités d’automatisation sur un microscope électronique à cryo-transmission de 200 kV et montre comment collecter des données pour la reconstruction de cartes 3D suffisantes pour la construction de modèles atomiques de novo . Nous nous concentrons sur les meilleures pratiques, les variables critiques et les problèmes courants qui doivent être pris en compte pour permettre la collecte de routine de tels ensembles de données cryo-EM à haute résolution. En particulier, les sujets essentiels suivants sont examinés en détail: i) automatisation des alignements de microscopes, ii) sélection de zones appropriées pour l’acquisition de données, iii) paramètres optiques optimaux pour la collecte de données de haute qualité et à haut débit, iv) réglage du filtre d’énergie pour l’imagerie sans perte, et v) gestion des données et évaluation de la qualité. L’application des meilleures pratiques et l’amélioration de la résolution réalisable à l’aide d’un filtre à énergie seront démontrées sur l’exemple de l’apo-ferritine qui a été reconstruite à 1,6 Å, et du protéasome Thermoplasma acidophilum 20S reconstruit à une résolution de 2,1-Å à l’aide d’un TEM de 200 kV équipé d’un filtre d’énergie et d’un détecteur d’électrons direct.
Introduction
La détermination de la structure des protéines est essentielle pour comprendre l’architecture moléculaire, la fonction et la régulation des complexes protéiques impliqués dans les processus cellulaires clés, tels que le métabolisme cellulaire, la transduction du signal ou les interactions hôte-pathogène. La cryo-microscopie électronique (cryo-EM) est apparue comme une technique puissante capable de résoudre la structure 3D de nombreuses protéines et de leurs complexes qui étaient trop difficiles pour les techniques structurelles traditionnelles, telles que la diffraction des rayons X et la spectroscopie RMN. En particulier, la cryo-EM a été prouvée comme la méthode de choix pour les protéines membranaires, qui ne peuvent pas être facilement cristallisées ou préparées en quantités suffisantes pour les techniques structurelles traditionnelles, et a fourni de nouvelles informations sur la structure et la fonction des récepteurs cellulaires importants et des canaux ioniques 1,2,3,4,5 . Plus récemment, cryo-EM a joué un rôle important dans la lutte contre la pandémie de Covid-19 en déterminant le mécanisme de l’infection par le SARS-CoV-2 au niveau moléculaire, ce qui a élucidé les origines de la maladie Covid-19 et a fourni la base pour le développement rapide de vaccins et de thérapies efficaces 6,7,8,9,10.
En règle générale, les microscopes électroniques à transmission (TEM) haut de gamme de 300 kV sont utilisés pour la détermination de la structure à haute résolution des biomolécules par analyse monoparticulaire (SPA) cryo-EM afin de révéler leur conformation et leurs interactions. Récemment, la technique SPA a atteint une nouvelle frontière lorsque l’échantillon de référence commun cryo-EM apo-ferritine a été reconstruit à une résolution atomique (1,2 Å)11,12 à l’aide d’un TEM de 300 kV équipé du pistolet d’émission de champ froid (E-CFEG), d’un détecteur d’électrons direct et d’un filtre à énergie. Lors de cette résolution, il a été possible de résoudre sans ambiguïté les positions des atomes individuels dans la structure, la conformation des chaînes latérales d’acides aminés individuelles ainsi que la liaison hydrogène et d’autres interactions, ce qui ouvre de nouvelles possibilités pour la découverte de nouvelles cibles par des médicaments basés sur la structure et l’optimisation de candidats médicaments existants.
Les microscopes TEM de milieu de gamme de 200 kV sont souvent utilisés pour le criblage des échantillons et l’optimisation des échantillons avant la collecte finale de données à haute résolution à l’aide des microscopes TEM haut de gamme, en particulier dans les grandes installations cryo-EM. En règle générale, les échantillons imagés peuvent être résolus dans la plage de résolution 3-4 Å qui est suffisante pour passer à un TEM haut de gamme de 300 kV pour la collecte finale des données. Par conséquent, la collecte de données à l’aide du TEM 200 kV n’est souvent pas optimisée davantage pour obtenir les résultats de résolution les plus élevés possibles. De plus, de nombreuses questions biologiques intéressantes peuvent déjà être répondues et publiées à ces résolutions, car toutes les chaînes latérales d’acides aminés sont déjà résolues et l’occupation des sites de liaison des ligands peut également être déterminée de manière fiable13. Il a déjà été démontré que les TEM 200 kV peuvent atteindre des résolutions supérieures à 3 Å pour de nombreux échantillons 14,15,16,17,18. Les images prises à 200 kV présentent un contraste intrinsèquement plus élevé des particules imagées, ce qui peut même faciliter un alignement initial plus précis des particules malgré un signal plus atténué à haute résolution par rapport aux images TEM de 300 kV. Il est important de noter que la résolution obtenue des cartes cryo-EM reconstruites est également limitée par la flexibilité structurelle et l’hétérogénéité conformationnelle des échantillons imagés, ce qui a un impact sur les reconstructions de 200 kV et de 300 kV. En fait, beaucoup plus de reconstructions cryo-EM obtenues à l’aide des systèmes 300 kV ont été résolues dans la plage de résolution 3-4 Å qu’à des résolutions plus élevées19. Comme les microscopes TEM de 200 kV sont moins complexes et s’intègrent dans des pièces plus petites, ces microscopes représentent une bonne option moins coûteuse pour la détermination de la structure des macromolécules biologiques par cryo-EM tout en préservant l’automatisation de longues collections de données à partir de plusieurs échantillons stockés dans le système de chargeur automatique de microscope.
La collecte d’ensembles de données cryo-EM pour la détermination de la structure à haute résolution nécessite un alignement précis de l’optique du microscope. Les alignements de colonnes vont systématiquement de la source d’électrons jusqu’au système de lentilles du condensateur, à la lentille de l’objectif et au filtre d’énergie avec un détecteur d’électrons. La séquence complète des alignements n’est généralement pas requise. En cas de besoin, l’utilisateur est guidé via des procédures semi-automatisées avec une description appropriée de chaque étape dans une fenêtre d’aide contextuelle tout au long de la procédure d’alignement dans l’interface utilisateur du microscope (panneau de commande Alignements directs). Une fois que le microscope est complètement aligné, l’optique électronique reste stable et les alignements n’ont pas besoin d’être modifiés pendant au moins quelques mois. Seuls les alignements les plus sensibles, tels que l’éclairage parallèle du plan de l’échantillon, l’astigmatisme objectif et l’alignement sans coma, doivent être affinés juste avant de commencer la collecte de chaque ensemble de données. La qualité des données collectées peut ensuite être surveillée lors de la collecte de données à l’aide de différents logiciels, tels que EPU Quality Monitor, cryoSPARC Live20, Relion21, Scipion22, WARP23 ou Appion24.
Outre les alignements précis du microscope, la haute qualité d’échantillons bien purifiés avec une hétérogénéité conformationnelle et compositionnelle minimale est également une condition préalable à la collecte d’ensembles de données à haute résolution et à la résolution de structures à haute résolution. Plus de détails sur les protocoles typiques, les défis fréquents et les remèdes possibles peuvent être trouvés dans d’autres revues consacrées à ce sujet 25,26,27. Essentiellement, il est essentiel de trouver des zones sur une grille cryo-EM donnée qui a une glace suffisamment mince pour préserver les informations à haute résolution, et les particules individuelles sont densément réparties à des orientations aléatoires sans chevauchement. Cependant, les grilles cryo-EM typiques ont une épaisseur de glace non uniforme, et il est donc important de trouver et de sélectionner les zones optimales pour l’imagerie. Différents moyens d’estimation de l’épaisseur de glace sur la grille sont disponibles dans des progiciels dédiés à la collecte automatisée d’ensembles de données cryo-EM, tels que EPU 2, Leginon28 ou SerialEM29.
L’avènement des détecteurs d’électrons directs rapides et sensibles a permis la collecte d’images en de nombreuses fractions sous forme de films qui ont permis la compensation des mouvements induits par le faisceau et ont entraîné une augmentation substantielle de la qualité et de la quantité de données utilisées dans le traitement d’images et la reconstruction 3D finale30. Dans le même temps, l’automatisation et la collecte de données à haut débit fournissent d’énormes ensembles de données avec des milliers d’images / films qui représentent des défis pour le stockage et l’accès aux données. Le modèle adopté avec de grandes installations cryo-EM desservant des dizaines à des centaines d’utilisateurs nécessite en particulier une gestion organisée des données avec un suivi et un partage appropriés des données dans les pipelines cryo-EM établis31,32.
Cette étude décrit un protocole de collecte de routine d’ensembles de données cryo-EM à haute résolution à l’aide du microscope TEM Glacios de 200 kV. Les alignements nécessaires de l’optique du microscope sont décrits ainsi que les procédures d’évaluation des échantillons cryo-EM et la sélection des zones appropriées pour la collecte de données à haute résolution. L’organisation des données collectées et des métadonnées connexes avec des exemples d’informations est démontrée dans Athena - une plate-forme de gestion des données qui facilite l’examen des informations sur les échantillons et des données collectées. En utilisant l’échantillon d’apo-ferritine de souris, il a été possible de réaliser une reconstruction 3D à une résolution de 1,6 Å13. En utilisant le protocole décrit, nous avons également reconstruit la carte de densité 3D du protéasome 20S de Thermoplasma acidophilum à une résolution de 2,1 Å.
Protocole
Toutes les étapes du protocole sont décrites pour le système TEM Glacios 200 kV (ci-après dénommé TEM 200 kV) équipé du filtre d’énergie Selectris-X (ci-après dénommé filtre d’énergie) et du détecteur Falcon 4 (ci-après dénommé détecteur d’électrons directs). Les étapes du protocole sont spécifiques à l’application EPU, qui est le logiciel de collecte de données SPA par défaut préinstallé sur chaque système Glacios. Les étapes du protocole ci-dessous correspondent à la version 2.14 de l’EPU et de petites modifications sont attendues lors de l’utilisation d’une version différente de l’EPU. Les conditions préalables à ce protocole sont les suivantes : i) les alignements du canon et de la colonne sont bien alignés, ii) les étalonnages EM sont corrects et iii) les fonctions automatiques de l’EPU sont correctement étalonnées.
1. Chargement des grilles dans le microscope
REMARQUE: Le TEM de 200 kV utilisé dans cette expérience peut contenir jusqu’à 12 grilles automatiques (c’est-à-dire des grilles TEM conventionnelles clipsées dans une cartouche spéciale) dans une cassette chargée à l’intérieur du chargeur automatique du microscope et constamment maintenue à des températures inférieures à -170 ° C pour empêcher la dévitrification de l’échantillon.
- Insérez des grilles automatiques dans la cassette autoloader dans des conditions d’azote liquide.
- Insérez la cassette avec des grilles automatiques dans une capsule de transfert refroidie à l’azote liquide.
- Insérez la capsule dans le microscope et cliquez sur le bouton Dock dans l’interface utilisateur du microscope pour charger la cassette de la capsule dans le chargeur automatique du microscope.
- Cliquez sur le bouton Inventaire pour vérifier la présence de grilles automatiques dans la cassette chargée.
- Cliquez sur les boutons Charger et Décharger pour insérer des grilles automatiques dans la colonne pour l’imagerie TEM.
2. Définition d’un projet dans une plateforme de gestion de données (facultatif)
REMARQUE: Les exemples d’informations et les données collectées peuvent être organisés dans la plate-forme de gestion des données fournie qui permet le stockage de données structurées pour tous les instruments connectés. Un projet peut être créé pour lequel toutes les étapes du flux de travail peuvent être définies pour capturer les images et les métadonnées de manière organisée pour révision et exportation.
- Démarrez l’application du portail de gestion des données et connectez-vous avec un nom d’utilisateur et un mot de passe.
- Dans le panneau de gauche de l’interface utilisateur du portail, cliquez sur le bouton Ajouter un projet pour créer un nouveau projet ou cliquez sur un bouton d’un projet existant dans la liste ci-dessous pour ouvrir un projet.
- Cliquez sur le bouton Ajouter une expérience pour créer une nouvelle expérience dans le projet ouvert.
- Remplissez la description dans le panneau Métadonnées de la nouvelle expérience et cliquez sur le bouton Ajouter un flux de travail pour créer un nouveau flux de travail dans l’expérience (par exemple, Analyse de particules uniques).
- Si vous le souhaitez, cliquez sur le bouton Ajouter une étape dans le panneau central pour créer un flux de travail personnalisé ou ajouter des étapes supplémentaires au flux de travail SPA prédéfini (Figure 1 et Figure 2).
REMARQUE: Les étapes peuvent représenter la préparation des échantillons, la caractérisation des échantillons par des techniques biochimiques, la vitrification des échantillons, le criblage des grilles cryo-EM, les sessions de collecte de données et l’analyse des données. - Si vous le souhaitez, remplissez les descriptions dans les notes de chaque étape, qui peuvent inclure des images et des photos.
- Créez une étape de jeu de données dans le flux de travail et sélectionnez le type de jeu de données comme EPU.
REMARQUE: Cela permettra au logiciel d’analyse de placer automatiquement toutes les données / métadonnées de résultats au bon endroit dans le portail de gestion des données lors de l’acquisition des données et d’exporter automatiquement les données vers la destination préférée tout en conservant un enregistrement complet de toutes les étapes et du transfert de données. - Remplissez des modèles sur les grilles Échantillon et EM à l’étape Biochimie et associez chaque grille à un échantillon (notez qu’un échantillon peut être associé à plusieurs grilles).
- Associez des combinaisons exemple-grilles dans la section métadonnées de l’étape Dataset .
3. Configurer des préréglages d’imagerie et des étalonnages de décalage d’image dans le logiciel d’analyse
- Définissez les paramètres d’imagerie pour les modes d’imagerie individuels, comme indiqué dans le tableau 1. Les considérations relatives à la sélection de paramètres spécifiques sont décrites en détail dans la section Discussion.
- Réglez l’éclairage parallèle pour le grossissement choisi dans le préréglage Acquisition de données du logiciel d’analyse
REMARQUE : Il n’y a qu’un seul réglage de la lentille C2 pour l’éclairage parallèle de l’échantillon à la taille SPOT donnée (c.-à-d. les forces prédéfinies de la lentille C1) sur les systèmes TEM à 2 condensateurs, tels que le TEM 200 kV utilisé dans cette étude. Le débit de dose d’électrons par unité de surface (e-/Å2/s) ne peut donc être ajusté qu’en modifiant les réglages de taille SPOT et en réajustant la force C2 (intensité du faisceau) en fonction des étapes ci-dessous.- Déplacez-vous vers une zone avec une feuille de carbone de support et de la glace mince ou nulle.
- Sélectionnez l’ouverture de l’objectif de 100 μm ou 70 μm dans le menu Ouverture de l’interface utilisateur TEM.
- Appuyez sur le bouton Diffraction des panneaux de commande pour passer en mode diffraction.
- Insérez l’écran fluorescent et changez le mode FluCam en haute résolution.
- Déplacez le point de diffraction central vers un bord de l’ouverture. Si le bord d’ouverture n’est pas visible, augmentez la longueur de l’appareil photo (à l’aide du bouton Grossissement).
- Tournez délicatement le bouton de mise au point du panneau de commande pour mettre au point le plan focal arrière. Le plan focal arrière est mis au point lorsque le bord d’ouverture est visiblement net.
- Réduisez la sensibilité de la FluCam au niveau le plus bas et déplacez le point de diffraction central au centre de la FluCam.
- Réduisez la taille du point central à l’aide du bouton Intensité du panneau de commande.
- Retirez l’ouverture de l’objectif dans l’interface utilisateur TEM et revenez au mode d’imagerie.
- Cliquez sur le bouton Obtenir dans le logiciel d’analyse pour enregistrer les paramètres dans le préréglage Acquisition de données.
- Ajustez le débit de dose d’électrons dans le préréglage d’acquisition de données qui est optimal pour le détecteur utilisé :
- Se déplacer vers une zone vide de la grille (par exemple, un carré de grille avec une feuille de carbone cassée)
- Cliquez sur le bouton Mesurer la dose dans le logiciel d’analyse pour mesurer le débit de dose d’électrons.
- Modifiez la taille SPOT pour obtenir le débit de dose optimal. Dans le cas du détecteur d’électrons directs utilisé dans ce protocole, un débit de dose de 4-5 e-/pixel/s est généralement utilisé pour l’acquisition de données à haute résolution. Cela correspond généralement à la taille SPOT 4-6.
- Vérifiez et ajustez l’éclairage parallèle au nouveau réglage de taille SPOT comme décrit ci-dessus.
- Sélectionnez l’option EER sous Fractions pour utiliser le mode EER sur le détecteur d’électrons direct33.
- Cliquez sur le bouton Obtenir dans le logiciel d’analyse pour enregistrer les paramètres dans le préréglage Acquisition de données.
- Basculez vers le paramètre prédéfini Autofocus et appuyez sur le bouton Obtenir pour enregistrer les paramètres d’éclairage pour la mise au point. Modifiez manuellement le temps d’exposition à 1 s et le mode de binning 2.
- Passez au préréglage Thon Rings et appuyez sur le bouton Obtenir pour enregistrer les paramètres d’éclairage de ce préréglage. Modifiez manuellement le temps d’exposition à 1-2 s et le mode de binning 2.
- Passez au préréglage Zero Loss et appuyez sur le bouton Obtenir pour enregistrer les paramètres d’éclairage de ce préréglage. Modifiez manuellement le temps d’exposition à 0,5 s et le mode de binning 4.
- Calibrer les décalages d’image entre les paramètres optiques définis, comme décrit dans le manuel du logiciel d’analyse, à l’aide de la tâche Calibration du décalage d’image de l’onglet Préparation (Figure supplémentaire 1).
Tableau 1 : Paramètres d’imagerie typiques pour l’acquisition de données à haute résolution à l’aide d’un cryo-TEM de 200 kV équipé d’un filtre à énergie et d’un détecteur d’électrons direct. Les paramètres sont affichés pour chaque préréglage optique utilisé dans la configuration de la collecte automatisée de données (section 3 du Protocole). Ces paramètres sont spécifiques au microscope TEM de 200 kV et au détecteur d’électrons directs utilisés dans cette étude. Veuillez cliquer ici pour télécharger ce tableau.
4. Cartographie de la grille et sélection des meilleures grilles cryo-EM pour la collecte de données
- Sélectionnez l’onglet Atlas et cliquez sur le bouton Nouvelle session pour ouvrir une nouvelle session.
- Remplissez des détails tels que le nom de la session et l’emplacement de stockage des données et cliquez sur le bouton Appliquer pour ouvrir une nouvelle fenêtre de tâche de filtrage , qui affiche une liste de toutes les grilles de l’inventaire du chargeur automatique. Si vous le souhaitez, modifiez les noms des grilles.
- Sélectionnez les grilles d’intérêt en cochant une case en regard du numéro de grille correspondant.
- Cliquez sur le bouton Démarrer pour démarrer une collection entièrement automatisée d’atlas de toutes les grilles sélectionnées.
- Une fois la collection terminée, cliquez sur les étiquettes de grille pour consulter les atlas acquis.
REMARQUE: Les carrés de grille individuels sont classés en fonction de leur épaisseur relative de glace, qui est basée sur l’évaluation de la valeur relative des niveaux de gris dans chaque atlas. Les carrés de grille classés sont représentés dans différents schémas de couleurs qui peuvent être utilisés pour guider la sélection des zones d’acquisition de données. Une grille produite avec un Vitrobot Mk IV affichera généralement un gradient d’épaisseur de glace sur l’ensemble de la grille, ce qui pourrait aider à identifier l’épaisseur de glace idéale pour la collecte de données. Une grille optimale doit contenir le moins de contamination possible par la glace de transfert et présenter suffisamment de carrés de grille ininterrompus et exempts de fissures (figure 3). Les grilles avec une distribution de glace appropriée peuvent être étudiées plus en détail pour évaluer la distribution des particules dans la glace à fort grossissement (c.-à-d. la densité et l’orientation des particules individuelles). - Cliquez sur le bouton Charger l’échantillon dans le menu supérieur pour insérer une grille choisie avec une distribution de glace appropriée dans la colonne du microscope.
- Sélectionnez l’onglet Atlas , cliquez avec le bouton droit de la souris sur un carré de grille approprié dans l’image de l’atlas et sélectionnez l’option Déplacer l’étape ici dans le menu déroulant pour déplacer la scène vers le carré de la grille.
- Sélectionnez l’onglet Fonctions automatiques pour définir le carré de la grille sur la hauteur eucentrique. Passez au préréglage Hauteur eucentrique , cliquez sur la fonction automatique Auto-Eucentric by Beam Tilt au milieu du carré de grille choisi et cliquez sur le bouton Démarrer .
- Basculez vers le préréglage Grid Square et acquérez une image. Déplacez la scène vers un trou avec de la glace et aucune contamination ou une contamination minimale par un clic droit et l’option Déplacer la scène ici .
- Basculez vers le préréglage Trou/Hauteur eucentrique et obtenez une image. Déplacez la scène vers une zone de carbone à côté du trou d’intérêt par un clic droit et l’option Déplacer la scène ici .
- Cliquez sur la fonction auto-Focus et définissez les valeurs de la mise au point souhaitée sur 0 μm et de l’itération sur -2 μm. Passez au préréglage Autofocus et cliquez sur le bouton Démarrer .
- Sélectionnez à nouveau l’onglet Préparation et passez au préréglage Trou/Hauteur eucentrique . Sur l’image de trou précédemment acquise, sélectionnez une zone d’intérêt dans le trou et déplacez la scène vers cette position en cliquant avec le bouton droit de la souris et en sélectionnant l’option Déplacer la scène ici .
- Passez au préréglage Acquisition de données et réglez le temps d’attente après le décalage du faisceau sur 0,5 s et le temps d’attente après le déplacement de l’étape sur 20 s. Acquérir une image en utilisant la mise au point entre -3 μm et -5 μm pour augmenter le contraste à basse résolution pour une meilleure visualisation des particules individuelles.
- En option, répétez les étapes 4.7 à 4.13 pour imager les particules dans différents trous et différents carrés de grille avec différentes épaisseurs de glace selon les besoins.
- Une fois que la grille la plus appropriée pour la collecte de données haute résolution est identifiée et sélectionnée, cliquez sur le bouton Charger l’échantillon dans le menu supérieur pour charger la grille sélectionnée dans le TEM.
REMARQUE: Sinon, si plus d’informations sont nécessaires et / ou l’automatisation de ce processus de dépistage est souhaitée, effectuez la section 5.
5. Mise en place d’une session de collecte de données dans le logiciel d’analyse de particules uniques
REMARQUE: Si vous utilisez les grilles de feuilles d’or, le raffinement de l’astigmatisme objectif et des alignements de coma (section 6) peut ne pas fonctionner de manière fiable. Il est conseillé de charger une grille de feuille de carbone ou une grille EM à réseau croisé et d’effectuer ces alignements finaux avant de mettre en place la collecte de données.
- Sélectionnez l’onglet EPU et cliquez sur le bouton Création de session pour créer une nouvelle session dans le panneau de gauche. Sélectionnez l’option Nouvelle session pour utiliser les préréglages optiques actuels ou l’option Nouveau à partir des préférences pour charger la configuration de session précédemment exportée.
- Renseignez le nom de la session et l’emplacement de stockage des données.
REMARQUE : Cet emplacement sera utilisé pour enregistrer des images et des métadonnées intégrées à partir de la session de collecte de données dans un sous-dossier avec le nom de la session. Bien que ces données n’occupent pas une quantité importante d’espace de stockage, il est recommandé d’enregistrer ces données sur le stockage de données de déchargement Falcon du serveur DMP car les images de caméra EER sont toujours stockées dans le répertoire racine de ce stockage de données dans un répertoire avec le nom de session. - Sélectionnez le type Manuel de la session pour contrôler la sélection des trous individuels dans les carrés de grille sélectionnés pour la collecte de données plus tard dans le protocole.
- Sélectionnez le mode d’acquisition plus rapide pour utiliser le décalage d’image sans aberration (AFIS) pour la collecte de données afin de réduire les mouvements de scène entre les trous individuels, de réduire la dérive globale de l’échantillon et d’augmenter le débit de collecte de données sans détérioration de la qualité de l’image.
- Sélectionnez le format mrc par défaut des images intégrées enregistrées.
- Spécifiez la grille utilisée et son type. Ce protocole utilisait le R-1.2/1.3 UltraAufoil pour l’apo-ferritine et la grille R-2/1 Quantifoil pour le protéasome 20S. Sélectionnez Quantifoil sous Spécimen porteur et R1.2/1.3 ou 2/1 sous Type quantifoil.
- FACULTATIF : Cliquez sur le bouton Athena Login dans le coin inférieur droit pour utiliser EPU Quality Monitor (EQM).
- Entrez les informations de connexion dans la fenêtre contextuelle du navigateur pour activer la section des paramètres dans la vue d’ensemble de la configuration de session .
- Cliquez sur le bouton Sélectionner dans la section des paramètres et recherchez le jeu de données créé précédemment (section 2 du protocole) pour l’associer à la collecte de données en cours. Activez la case à cocher Activer le Moniteur de qualité .
- Cliquez sur le bouton Appliquer pour créer une nouvelle session.
REMARQUE : cette action ouvrira de nouvelles tâches dans le menu de la colonne de gauche. À tout moment de la session, si certains détails sont incorrects, il est possible de revenir à la tâche de configuration de la session , de modifier / mettre à jour les détails et de cliquer à nouveau sur Appliquer pour mettre à jour la session. - Sélectionnez la tâche Sélection carrée dans le panneau de gauche pour afficher l’atlas collecté de la grille.
- En option, double-cliquez sur n’importe quelle tuile de l’atlas pour voir une image de meilleure qualité afin de mieux juger de la qualité de la glace dans les carrés de la grille. Double-cliquez à nouveau sur l’image pour revenir à l’atlas de la grille.
- Identifiez les carrés de grille présentant les caractéristiques suivantes (Figure 4) : (i) La feuille de support dans le carré de la grille est intacte sans dommage, (ii) Glace mince vitrée dans les trous de feuille (les trous semblent plus brillants que la feuille de carbone de support), (iii) Aussi peu de contamination cristalline que possible par la glace cristalline (taches noires) dans le carré de la grille, (iv) Gradient de luminosité minimal à travers le carré de la grille et dans les trous de feuille individuels.
REMARQUE: Avec les préréglages choisis et avec une bonne grille, le cryo TEM 200 kV utilisé dans cette étude peut imager à un taux d’environ 200-300 films / h. Dans cet esprit, sélectionnez autant de carrés que nécessaire, ou ajoutez-en d’autres plus tard en revenant à la tâche de sélection des carrés en fonction du temps de microscope disponible. Pour référence, 3000 films ont été collectés pour atteindre une résolution de 1,6 Å d’apo-ferritine. - Sélectionnez des carrés de grille pour la collecte de données dans l’atlas complet ou dans des images de tuiles de haute qualité
- Cliquez avec le bouton droit de la souris sur un carré de grille d’intérêt et choisissez l’option Sélectionner dans le menu contextuel
- Vous pouvez également maintenir la touche Ctrl du clavier enfoncée et cliquer avec le bouton gauche de la souris sur les carrés de grille souhaités.
- Inversement, cliquez avec le bouton droit de la souris et désélectionnez ou maintenez la touche Maj enfoncée et cliquez avec le bouton gauche pour supprimer les carrés.
- Sélectionnez la tâche Sélection de trous dans le panneau de gauche pour sélectionner des trous dans les carrés de grille sélectionnés.
- Cliquez sur le bouton Auto-Eucentrique pour passer automatiquement au premier carré de grille sélectionné, ajuster la hauteur eucentrique et acquérir une image carrée de grille pour trouver des trous de feuille.
- Cliquez sur le bouton Trouver des trous pour trouver des trous de feuille dans l’image.
REMARQUE : Si la recherche de trous n’a pas bien fonctionné (c.-à-d. qu’il y a des trous sautés ou des trous de taille incorrecte dans l’image), vérifiez que les valeurs correctes ont été entrées dans l’entrée Type de quantifoil de la tâche Configuration de session et recherchez à nouveau les trous. Si les trous ne sont toujours pas trouvés correctement, cliquez sur le bouton Mesurer la taille du trou pour régler manuellement le diamètre et la distance du trou et trouver à nouveau les trous. - Cliquez sur le bouton Supprimer les trous près du bouton de la barre de grille pour désélectionner les trous près des barres de grille.
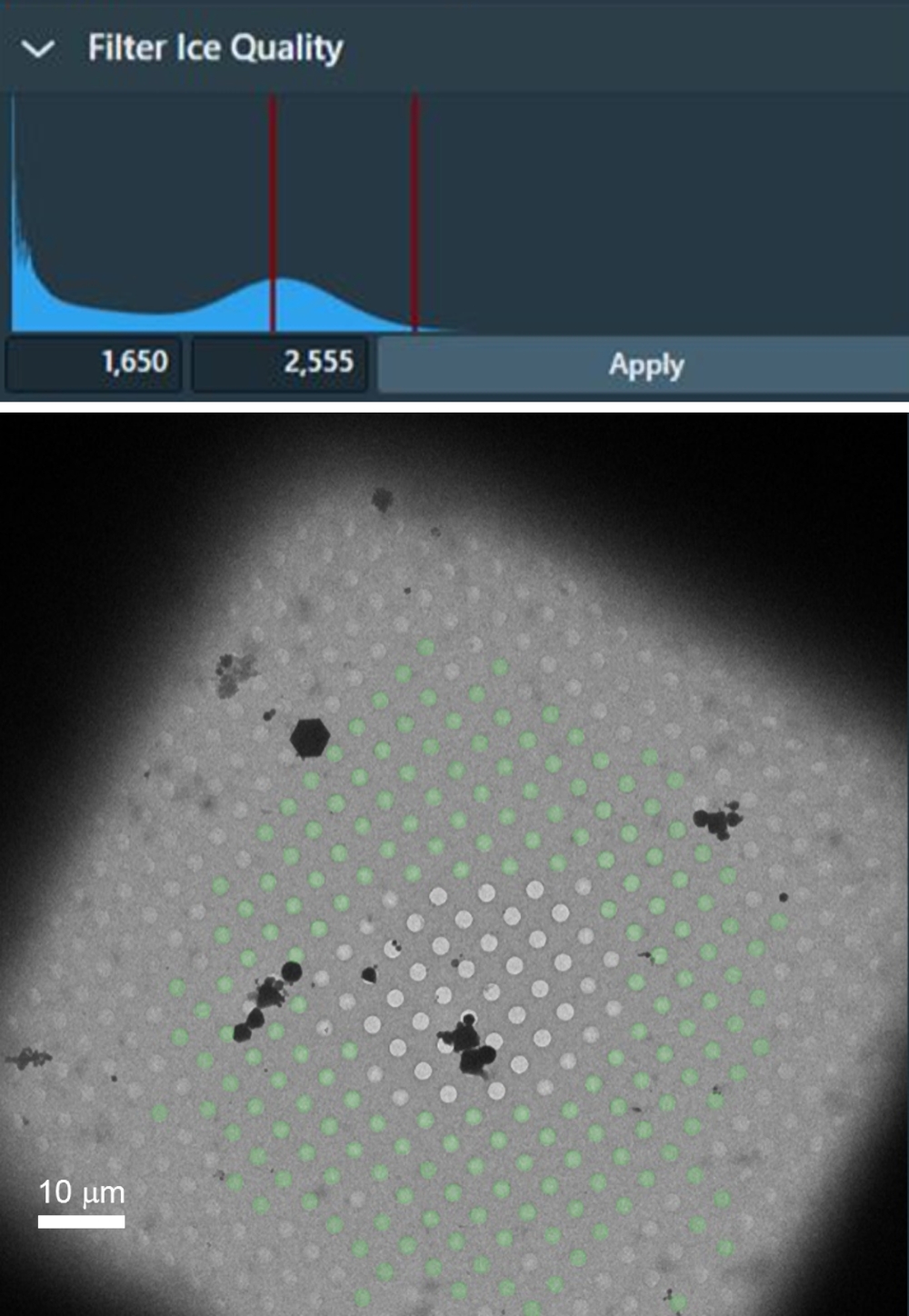
- Ajustez les limites dans l’histogramme de luminosité du filtre à glace (en bas à droite de la fenêtre du logiciel) pour supprimer tous les trous avec de la glace trop épaisse (en utilisant la ligne limite gauche) ainsi que tous les trous vides (en utilisant la ligne limite droite) comme illustré à la figure 4.
REMARQUE: Pour plus d’affinement, des valeurs d’intensité spécifiques peuvent également être saisies dans les zones de texte correspondantes à côté du bouton Appliquer . - Si vous le souhaitez, utilisez la forme de sélection pour affiner manuellement la sélection des trous : cliquez avec le bouton gauche de la souris et faites-la glisser pour désélectionner les trous indésirables. Maintenez la touche Ctrl enfoncée et cliquez avec le bouton gauche de la souris pour pouvoir resélectionner les trous de feuille. Maintenez la touche Maj enfoncée et faites défiler avec la molette de la souris pour ajuster la taille du pinceau.
- Sélectionnez un trou pour définir et tester un modèle d’acquisition de données qui sera utilisé à plusieurs reprises tout au long de la collecte de données : Cliquez avec le bouton droit de la souris sur un trou dans l’image carrée de la grille et sélectionnez l’option Déplacer la scène vers l’emplacement.
- Sélectionnez la tâche Définition de modèle dans le panneau de gauche.
- Sélectionnez le préréglage Trou/Eucentrique et cliquez sur le bouton Acquérir pour acquérir une image.
- Cliquez sur le bouton Rechercher et centrer le trou pour centrer un trou dans l’image.
- Définissez les valeurs de Delay After Image Shift sur 0,5 s (par défaut) et Delay After Stage Shift sur 20 s.
REMARQUE: Comme le diamètre du faisceau parallèle est fixé sur le système cryo-TEM de 200 kV utilisé dans cette étude et correspond généralement à 1,6-1,7 μm avec une ouverture de 50-μm, seulement 1 image peut être acquise par trou en utilisant les grilles R-1.2 / 1.3 et 2 images par trou (bords presque opposés) en utilisant les grilles R-2/1 ou R-2/2. Veuillez noter que la taille des zones d’acquisition définies à l’écran ne correspond pas au diamètre réel du faisceau. La barre d’échelle de l’image peut être utilisée pour estimer une distance de placement appropriée. - Sélectionnez le bouton Ajouter une zone d’acquisition et cliquez sur l’image pour sélectionner l’emplacement dans le trou centré d’où l’acquisition d’image à fort grossissement sera prise.
- Sélectionnez le bouton Ajouter une zone de mise au point automatique et cliquez sur l’image pour sélectionner l’emplacement sur la feuille de support à côté du trou centré où l’autofocus de l’image sera effectué.
REMARQUE: La taille du faisceau pour la mise au point est de 1,6 à 1,7 μm et doit être placée à égale distance des trous voisins sur le carbone. - Cliquez sur la zone d’acquisition verte pour définir une séquence de valeurs de mise au point dans la liste de mise au point dans la section supérieure de la fenêtre du logiciel. Entrez le déconcentrage le plus élevé en premier pour améliorer la visualisation des particules dans une série de tests de la tâche d’exécution de modèle suivante.
- Cliquez sur la zone de mise au point automatique bleue pour définir les paramètres spécifiques à la mise au point automatique dans la même zone :
- Choisissez l’option Après centrage sur la mise au point automatique au début de chaque cluster AFIS.
- Choisissez l’option Objectif objectif pour une mise au point automatique plus rapide et une dérive de scène réduite.
- Sélectionnez la tâche d’exécution du modèle et cliquez sur le bouton Exécuter.
- Observez les différentes étapes de la procédure d’acquisition de données (centrer le trou, autofocus dans la zone sélectionnée et acquisition d’image dans les zones définies) et vérifier la qualité des particules imagées dans l’image finale à fort grossissement.
- Cliquez sur le bouton FFT en bas à droite de la fenêtre d’image à fort grossissement pour vérifier l’image FFT et évaluer visuellement si les anneaux Thon présentent plusieurs oscillations et s’étendent à haute résolution dans l’image FFT.
- Si l’exécution du modèle est terminée avec succès, cliquez sur le bouton Préparer tous les carrés dans la tâche Sélection de trous pour que la collecte de données soit définie automatiquement dans tous les autres carrés de grille sélectionnés en fonction des paramètres utilisés dans ce premier carré de grille.
6. Alignements finaux du microscope avant de commencer la collecte de données
REMARQUE: Pour obtenir les meilleurs résultats haute résolution, les alignements les plus sensibles doivent être effectués exactement aux mêmes paramètres que le mode d’acquisition de données dans le logiciel de collecte de données et juste avant de commencer l’acquisition de données réelle. Ces alignements doivent être effectués à une position avec un carbone de support mince de la grille, suffisamment éloigné de toute barre de grille et aligné à la hauteur eucentrique.
- Sélectionnez la tâche d’exécution du modèle , acquérez une nouvelle image et déplacez-vous avec la scène vers une zone propre sur une feuille de carbone par un clic droit et sélectionnez l’option de menu Déplacer la scène ici.
- Sélectionnez l’onglet Fonctions automatiques et définissez le défocalisation souhaité sur 0 μm et l’itération sur -2 μm. Passez au préréglage Autofocus et cliquez sur le bouton Démarrer pour exécuter la fonction Autofocus.
- Posez l’écran fluorescent et ouvrez le menu Alignements directs dans l’interface utilisateur TEM.
- Pour de meilleurs résultats, les utilisateurs expérimentés peuvent vérifier les alignements des points de pivot : choisissez la tâche nP Beam Tilt pp X dans le menu Alignements directs et chevauchez au maximum les faisceaux rebondissants à l’aide des boutons multifonctions des panneaux de commande. Répétez l’opération pour la tâche np Beam Tilt pp Y .
- Cliquez sur le bouton EF dans le menu de la caméra à écran fluorescent pour afficher un cercle vert qui indique l’ouverture d’entrée du filtre d’énergie et centre le faisceau sur le cercle vert.
REMARQUE: Assurez-vous que la pompe Turbo est arrêtée avant de continuer; les vibrations qui en découlent réduisent le nombre d’anneaux de thon visibles et provoquent fréquemment l’échec des fonctions automatiques. - Revenez à la fenêtre du logiciel et sélectionnez l’onglet Fonctions automatiques dans la barre de menus.
- Sélectionnez la tâche Autostigmate, passez au préréglage Thon Ring et appuyez sur le bouton Démarrer. Observez le processus pour vous assurer que (i) les images prises sont sur du carbone, (ii) les anneaux thon sont clairement visibles et (iii) l’ajustement CTF calculé (lignes pointillées) sont bien placés aux minima thon-ring (figure supplémentaire 2).
- Sélectionnez la tâche Autocoma et appuyez sur le bouton Démarrer . Observez le processus pour vous assurer que les images prises sont sur du carbone et que les anneaux thon sont clairement visibles et que les ajustements calculés (lignes pointillées) sont bien placés aux minima de thon-ring. Zoomez sur chaque image Thon-ring à l’aide de la molette de défilement de la souris (Figure supplémentaire 3).
REMARQUE: Si le microscope est équipé d’un filtre à énergie, le réglage du filtre à énergie doit être effectué dans le cadre de la séquence d’alignement pour centrer la fente du filtre sur le pic de perte zéro et pour corriger toute distorsion dans le prisme du filtre à énergie (étapes 6.9-6.14). Ces alignements doivent être effectués dans une zone vide de la grille, par exemple à l’intérieur d’un carré de grille cassé. - Ouvrez l’interface utilisateur Sherpa et sélectionnez l’application Filtre d’énergie (Figure supplémentaire 4).
- Réglez l’appareil photo sur EF-Falcon, bin 4x, temps d’exposition 0,2 s dans la fenêtre Paramètres .
- Cliquez sur le bouton Centre dans l’option Zéro perte pour centrer la fente du filtre d’énergie zéro perte.
- Cliquez sur le bouton Tune dans l’option Isochromaticité (Figure supplémentaire 4).
- Cliquez sur Le grossissement de réglage et les distorsions de réglage dans l’option Distorsions géométriques et chromatiques (Figure supplémentaire 5, Figure supplémentaire 6)
- Si les résultats ne sont pas conformes aux spécifications indiquées et affichés en rouge dans le rapport de sortie, itérez à nouveau les alignements à partir de l’étape 6.11.
REMARQUE: Bien que la référence de gain du détecteur d’électrons direct puisse être stable pendant des mois, une nouvelle référence de gain doit être prise à l’aide du gestionnaire d’images de référence Falcon 4 si les images de zones vides ne montrent pas une intensité uniforme mais présentent des rayures ou d’autres caractéristiques distinctes. Vous pouvez également calculer une transformée de Fourier rapide (FFT) d’une telle image et vous assurer qu’aucune ligne n’est visible. - Dans la fenêtre du logiciel, sélectionnez l’onglet Préparation et basculez vers le paramètre prédéfini Acquisition de données .
- Réglez le réglage de la dose sur 40 e-/Å2 et cliquez sur le bouton Mesurer le débit de dose .
- Accédez à l’onglet EPU , sélectionnez la tâche Acquisition automatisée et cochez éventuellement la fonction Auto Zero Loss ; cliquez sur le bouton Démarrer l’exécution pour commencer la collecte de données entièrement automatisée.
7. Surveillance et optimisation de la qualité des données lors de la collecte des données
REMARQUE : Pendant que la collecte de données est en cours, les données collectées peuvent être surveillées à l’aide de l’EQM via le portail de gestion des données. EQM effectue la correction de mouvement et la détermination CTF à la volée et affiche les résultats dans le portail. L’utilisateur est alors en mesure de juger de la qualité des acquisitions individuelles, de voir des graphiques sur divers indicateurs de qualité, de filtrer les acquisitions indésirables et d’exporter les données vers leur stockage final, soit à la volée, soit sous forme de traitement par lots.
- Accédez au jeu de données dans lequel le logiciel d’analyse place les données à l’aide d’un navigateur Web.
- Dans la vue d’ensemble de l’ensemble de données, les cartes d’acquisition affichent les informations sur la correction de mouvement et la détermination CTF dans un format graphique. Cliquez sur les cartes individuelles pour obtenir plus d’informations.
- Activez le panneau DataViz pour afficher des graphiques agrégés à partir de l’ensemble de données complet qui montrent la plage de confiance du défocus, de l’astigmatisme et du CTF par acquisition (figure supplémentaire 7).
- Utilisez les filtres situés en haut du panneau pour sélectionner uniquement les données qui se trouvent dans la plage de mise au point demandée, dont l’astigmatisme est proche de zéro et qui peuvent être déterminées jusqu’à la résolution spécifiée (cible). Une fois les filtres configurés, appuyez sur le bouton Appliquer pour afficher uniquement les données choisies dans la fenêtre de vue d’ensemble du jeu de données.
- Lorsque la plupart des images acquises respectent les critères définis, laissez la session de collecte de données se poursuivre. Si seule une très petite fraction des images acquises répond aux critères définis, reconfigurez les paramètres du logiciel d’analyse, déplacez-vous vers une autre zone de l’échantillon (appuyez sur le bouton Next Grid Square dans le logiciel d’analyse) ou arrêtez la collecte de données.
Résultats
Le portail de gestion des données fournit un stockage efficace et structuré des images, données et métadonnées collectées à partir de plusieurs flux de travail expérimentaux dans une seule plate-forme logicielle. Chaque expérience définie dans un projet créé consiste en un flux de travail avec des étapes définies par le client pour capturer des informations d’échantillon, des données collectées et des métadonnées associées sans aucune contrainte afin de fournir une flexibilité et une convivialité maximales pour toutes les expériences possibles et tous les cas d’utilisation (Figure 1, Figure 2). Le portail de gestion des données dispose également d’une fonctionnalité de note de laboratoire pour illustrer les étapes du flux de travail, y compris le traitement d’images avec des résultats intermédiaires, qui peuvent être tous associés à un projet et fournir un enregistrement aussi complet que possible pour l’analyse et la création de rapports et de publications.

Figure 1 : Exemple d’organisation possible des données et métadonnées dans la plateforme de gestion des données. Chaque projet peut consister en plusieurs expériences, telles que la cryo-EM ou la spectroscopie de masse (c’est-à-dire l’étape 2.3 du protocole). Chaque expérience peut inclure plusieurs workflows définis par l’utilisateur (à savoir l’étape de protocole 2.5), chacun composé de plusieurs étapes configurables (à savoir l’étape de protocole 2.7). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 2 : Affichage d’un flux de travail de projet ouvert dans la plate-forme de gestion des données. La figure montre les métadonnées et les notes associées à l’étape ouverte dans le flux de travail. La barre de gauche avec des icônes permet d’accéder rapidement aux différentes options et menus de la plate-forme de gestion des données. Le panneau de gauche comprend une liste des Workflows enregistrés (n’affichant qu’un seul Workflow enregistré « Exp2_ApoF_EFTEM_Grid7 ») et un bouton bleu pour ajouter un nouveau Workflow. Le panneau central affiche les étapes individuelles du flux de travail ouvert, comme illustré pour le flux de travail SPA ici. Le bouton bleu en haut à droite peut ajouter une étape supplémentaire au flux de travail ouvert. Le panneau de droite comprend de l’espace pour les métadonnées enregistrées ou les notes d’entrée utilisateur du flux de travail, qui peuvent inclure du texte, des tableaux et des images. Différentes options de mise en forme du texte sont disponibles. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Les grilles Cryo-EM produites avec des dispositifs traditionnels de congélation par immersion, tels que Vitrobot, affichent généralement un gradient d’épaisseur de glace sur la surface de la grille. Certaines grilles peuvent également être endommagées (pliées) après une manipulation manuelle et / ou un écrêtage dans un support d’anneau de grille automatique. La figure 3 montre des exemples de différentes grilles, comme le montre la vue d’ensemble de l’Atlas. Les grilles avec de la glace épaisse ou des dommages devraient être exclues de toute enquête plus approfondie.

Figure 3: Galerie de différentes grilles comme on le voit dans l’aperçu de l’Atlas. (A) Une mauvaise grille avec de la glace épaisse, (B) une grille pliée avec une mauvaise contamination de la glace et de la glace, (C) une grille acceptable avec un bon gradient de glace, (D) une grille typique avec une bonne glace mince et un petit gradient de glace. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
La sélection de carrés de grille sans dommage et avec une épaisseur de glace optimale est essentielle pour collecter des ensembles de données à haute résolution. L’épaisseur de la glace peut varier même au niveau des carrés de grille individuels, et il est donc important de ne sélectionner que des trous avec une glace d’une épaisseur optimale de chaque carré de grille sélectionné. La figure 4 montre un carré de grille approprié avec une feuille intacte et de la glace mince au centre. Le carré de grille affiché est bon pour définir un filtre pour la sélection automatisée de trous avec de la glace mince dans tous les carrés de grille sélectionnés car il contient une gamme d’épaisseurs de glace différentes ainsi que des trous vides sans glace, ce qui est extrêmement utile pour définir une plage d’intensité appropriée dans le filtre à glace dans la tâche de sélection de trou.

Figure 4 : Exemple de carré de grille avec un gradient d’épaisseur de glace, à partir de carrés de grille vides au centre et de glace épaisse près des barres de grille. Le filtre de qualité de la glace peut être utilisé pour sélectionner la gamme d’intensités à l’intérieur des trous avec l’épaisseur de glace idéale qui sont sélectionnées en conséquence dans le carré de la grille (les trous avec superposition verte). Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Les résultats de référence utilisant le protocole décrit ont été obtenus à l’aide de l’échantillon d’apo-ferritine de souris (apoF) du groupe Kikkawa11. L’ApoF est une protéine hautement α-hélicoïdale qui forme une cage octaédrique très stable. La grande stabilité et la symétrie élevée font de l’apoF un échantillon optimal pour l’imagerie cryo-EM haute résolution et le traitement d’images. L’ApoF est donc devenu un échantillon standard pour évaluer les performances des instruments cryo-EM 11,12,13. Une aliquote congelée contenant un échantillon d’apoF purifié de 15 mg/mL a été décongelée sur de la glace et clarifiée par centrifugation à 10 000 x g pendant 10 min. Le surnageant a été dilué à 5 mg/mL avec 20 mM hePES pH 7,5, 150 mM NaCl. 3 μL de l’échantillon dilué ont été appliqués sur une grille d’or À décharge incandescente R-1.2/1.3, 300 mailles pendant 30 s. Ensuite, les grilles ont été effacées pendant 5 s avant de plonger dans de l’éthane liquide refroidi par de l’azote liquide. La congélation par immersion a été réalisée à l’aide d’un système de vitrification entièrement automatisé à 100% d’humidité et 4 °C. Toutes les grilles ont été clipsées dans des autogrids et chargées dans un Cryo-TEM de 200 kV. Environ 3000 films ont été collectés à un débit de 300 films/h. Les données ont été traitées à l’aide des méthodes décrites11 avec les modifications suivantes: i) la version 4-bêta de Relion a été utilisée à la place de Relion 3.1, ii) la sélection automatisée des particules a été effectuée en utilisant les moyennes de classe 2D des reconstructions apoF précédentes comme références, et iii) le modèle 3D initial a été généré à partir de la reconstruction apoF précédente à une résolution de faible passage à 15-Å. Le regroupement optique n’a pas été effectué pour cet ensemble de données, car il a été prouvé que la procédure AFIS34 utilisée minimise efficacement et de manière fiable les déphasages induits par l’inclinaison du faisceau qui ne limitent pas la qualité des données pour reconstruire des cartes 3D aux résolutions rapportées. Le raffinement 3D après le polissage bayésien et le raffinement CTF ont conduit à une carte de résolution de 1,68 Å. La résolution a été encore améliorée avec la correction de la sphère d’Ewald résultant en une carte de résolution de 1,63 Å. La vue d’ensemble des paramètres de collecte et de traitement des données est présentée dans le tableau 2, et la carte de densité reconstruite finale est illustrée à la figure 5, avec la courbe de corrélation de la coque de Fourier (FSC) illustrée dans la figure supplémentaire 8.
Tableau 2 : Paramètres de collecte de données et de traitement d’images utilisés pour la reconstruction 3D de l’apo-ferritine. Veuillez cliquer ici pour télécharger ce tableau.

Figure 5 : Reconstruction cryo-EM de l’apo-ferritine. (Panneau de gauche) Rendu 3D de la carte cryo-EM apoF reconstruite à une résolution de 1,6 Å. (Panneau de droite) Vue détaillée de la carte reconstruite au niveau des chaînes latérales d’acides aminés individuelles. La densité des chaînes latérales d’acides aminés est bien résolue et le modèle atomique peut être construit sans ambiguïté dans cette carte. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Les effets et les avantages de l’utilisation d’un filtre d’énergie dans les reconstructions SPA ont été évalués à l’aide du protéasome procaryote 20S isolé de T. acidophilum. Le protéasome procaryote 20S a également été utilisé comme échantillon cryo-EM standard car il représente le noyau catalytique stable du complexe protéasome avec la symétrie D7. Les grilles ont été préparées en ajoutant 4,5 μL de l’échantillon de protéasome purifié T. acidophilum 20S sur une grille de cuivre R 2/1 à 200 mailles luminescentes. Les échantillons ont été vitrifiés dans un mélange liquide éthane/propane à l’aide d’un système de vitrification entièrement automatisé réglé à 4 °C et 100 % d’humidité avec une force de transfert de 20 et un temps de transfert de 4,5 s.
Trois ensembles de données différents ont été collectés à partir de la même grille cryo-EM avec des carrés de grille similaires en utilisant une largeur de fente différente du filtre d’énergie dans l’ordre: i) fente entièrement ouverte (pas de fente insérée), ii) fente de 20 eV et iii) fente de 10 eV. Les carrés de la grille ont été sélectionnés à l’aide d’un filtre de qualité de glace dans le logiciel d’analyse. Tous les autres paramètres de collecte et de traitement des données ont été conservés. Les ensembles de données ont été collectés pendant 15 h avec un total de 4000 films et traités à l’aide des méthodes décrites11 à l’aide de Relion 3.1 avec la modification que l’algorithme de prélèvement de particules laplacien de Gaussien a été utilisé pour produire des moyennes de classe 2D initiales pour la sélection de particules basée sur des références à partir des ensembles de données complets. Le même nombre (102 200) de particules sélectionnées au hasard a été choisi et utilisé pour l’itération finale et la reconstruction 3D de chaque ensemble de données. Les variables de traitement des données sont décrites dans le tableau (tableau 3) ci-dessous pour obtenir la carte finale reconstruite de la densité EM illustrée à la figure 6 avec la courbe FSC illustrée à la figure supplémentaire 9. Le regroupement optique et la correction de la sphère d’Ewald n’ont pas non plus été effectués pour ces ensembles de données.
Tableau 3 : Paramètres de collecte de données et de traitement d’images utilisés pour la reconstruction 3D du protéasome T. acidophilum 20S. Veuillez cliquer ici pour télécharger ce tableau.
Tableau 4 : Résumé de la résolution atteinte et du facteur B pour les reconstructions cryo-EM du protéasome T. acidophilum 20S à l’aide d’ensembles de données avec différentes largeurs de fente d’énergie. Veuillez cliquer ici pour télécharger ce tableau.

Figure 6: Effet du filtrage d’énergie sur les images cryo-EM. (A) Images cryo-EM à différentes valeurs de mise au point collectées avec ou sans fente de 10 eV. (B) Vue d’ensemble de la carte cryo-EM du protéasome 20S avec sous-unités segmentées. (C) Vue zoom sur la carte du protéasome 20S avec un modèle atomique ajusté. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
Figure supplémentaire 1 : Tâche d’étalonnage des décalages d’image (ellipse jaune) pour aligner les décalages d’image entre différents préréglages optiques (ellipses rouges) dans le logiciel d’analyse, à l’aide d’un cristal de glace hexagonale visible dans la plage de grossissement complète comprise entre 100x et 165 000x. (Haut) Étalonnage entre les préréglages Acquisition de données et Hauteur trou/Eucentrique, étalonnage (au milieu) entre les préréglages Hole/Eucentric Height et Grid Square, étalonnage (en bas) entre les préréglages Grid Square et Atlas. Veuillez cliquer ici pour télécharger ce fichier.
Figure supplémentaire 2 : Fonction Autostigmate dans un logiciel d’analyse (ellipse jaune). (Image de gauche) Image acquise. (Image de droite) Transfert de Fourier de l’image acquise montrant des anneaux de Thon concentriques et leur ajustement CTF montré dans des faisceaux radiaux. Veuillez cliquer ici pour télécharger ce fichier.
Figure supplémentaire 3 : Interface utilisateur de la fonction Autocoma dans le logiciel d’analyse (ellipse jaune) pour l’alignement du coma. Le panneau d’images montre les images de transfert de Fourier acquises à différentes inclinaisons de faisceau et leurs ajustements CTF qui sont utilisés pour le calcul du coma. Veuillez cliquer ici pour télécharger ce fichier.
Figure supplémentaire 4 : Interface utilisateur du réglage du filtre à énergie. Exemple d’un bon rapport de tunning de l’isochromacité du filtre d’énergie avec tous les paramètres (indiqués en texte vert) dans les spécifications. Veuillez cliquer ici pour télécharger ce fichier.
Figure supplémentaire 5 : Interface utilisateur du réglage du filtre d’énergie. Exemple de bon rapport de réglage des distorsions de grossissement du filtre d’énergie avec tous les paramètres (indiqués en texte vert) dans les spécifications. Veuillez cliquer ici pour télécharger ce fichier.
Figure supplémentaire 6 : Interface utilisateur du réglage du filtre à énergie. Exemple de bon réglage des distorsions chromatiques du filtre d’énergie avec tous les paramètres (indiqués en texte vert) dans les spécifications. Veuillez cliquer ici pour télécharger ce fichier.
Figure supplémentaire 7 : Panneau DataViz du moniteur de qualité de l’EPU avec une vue d’ensemble de la qualité des données dans un ensemble de données cryo-EM collecté. Les graphiques avec des données agrégées de toutes les images/films collectés montrent les valeurs (diagrammes à points) et la distribution (graphiques à barres) de certains indicateurs de qualité critiques, tels que la confiance dans l’ajustement du FCE (bleu), le défocalisation (orange) et l’astigmatisme (vert). Un sous-ensemble des images/films collectés peut être sélectionné en définissant des filtres de paramètres en haut du panneau DataViz. Après avoir appliqué les filtres, les images/films sélectionnés peuvent être exportés pour un traitement ultérieur dans un autre package de traitement d’image, tel que Relion ou CryoSpark. Veuillez cliquer ici pour télécharger ce fichier.
Figure supplémentaire 8 : Courbe FSC de la reconstruction finale de l’apoF à une résolution de 1,6 Å, telle que rapportée par Relion 4-beta. La courbe bleue montre le FSC des cartes 3D masquées à partir de deux reconstructions 3D affinées indépendamment à partir de deux demi-ensembles de données mutuellement exclusifs. Selon l’étalon-or FSC à 0,143, la résolution de la carte 3D finale reconstruite à partir de l’ensemble de données complet correspond à 1,6 Å. La courbe orange montre le FSC des reconstructions 3D masquées avec des phases aléatoires. La chute rapide de la courbe FSC indique que le masque utilisé n’a pas contribué au FSC observé des cartes reconstruites originales (courbe bleue) au-delà de la résolution ~2 Å. Veuillez cliquer ici pour télécharger ce fichier.
Figure supplémentaire 9 : Courbes FSC de la reconstruction finale du protéasome T. acidophilum 20S à l’aide de différentes largeurs de fente du filtre d’énergie, comme indiqué par Relion 3.1. Les courbes bleues montrent le FSC des cartes 3D masquées à partir de deux reconstructions affinées indépendamment à partir de deux demi-ensembles de données de chaque ensemble de données, respectivement. L’étalon-or FSC à 0,143 indique les résolutions obtenues des cartes 3D finales reconstruites à partir des ensembles de données complets respectifs (résolution 2,3 Å, 2,2 Å et 2,1 Å, respectivement). Les courbes rouges montrent le FSC des cartes masquées avec des phases aléatoires. La chute rapide des courbes ROUGES FSC indique que le masque utilisé n’a pas contribué au FSC des cartes reconstruites originales au-delà de la résolution ~3 Å. Les courbes vertes montrent le FSC des cartes 3D non masquées, qui sont influencées par le bruit dans l’ensemble du volume 3D reconstruit et tombent donc plus tôt que le FSC des cartes 3D masquées. Veuillez cliquer ici pour télécharger ce fichier.
Disponibilité des données : Les cartes de densité cryo-EM ont été déposées dans la banque de données EM sous les numéros d’acquisition: apoferritine: EMD 14173, EMPIAR-10973. Protéasome 20S : EMD 14467, EMPIAR-10976.
Discussion
Le protocole décrit suppose que l’optique du microscope TEM utilisé est dans un état bien aligné. Pour la GDT de 200 kV utilisée dans ce protocole, ces alignements de colonnes sont effectués, vérifiés et enregistrés par un technicien de service expérimenté après l’installation du microscope ou toute intervention de service importante. Ces paramètres d’alignement peuvent être rappelés à tout moment dans l’interface utilisateur du microscope. Les utilisateurs peuvent utiliser les procédures d’alignement direct dans l’interface utilisateur du microscope pour modifier à nouveau les paramètres critiques. Certains alignements, tels que l’inclinaison et le changement de canon, sont stables et n’ont pas besoin d’être ajustés quotidiennement par les utilisateurs. Les vérifications et les réalignements (si nécessaire) de l’inclinaison et du décalage du pistolet par le superviseur du microscope sont conseillés deux fois par an. D’autre part, certains alignements sont critiques et doivent être alignés avant chaque collecte de données comme décrit dans le protocole ci-dessus (comme l’astigmatisme objectif et l’alignement sans coma). Si la fonction Autocoma du logiciel d’analyse ne parvient pas à converger, l’alignement des points de pivot d’inclinaison du faisceau et/ou du centre de rotation doit être vérifié et ajusté, et le centrage correct de l’ouverture C2 doit être confirmé. Par la suite, la fonction Autostigmate doit être exécutée car des stigmates objectifs sont également utilisés pour la correction du coma. Ces alignements doivent être itérés jusqu’à ce que les fonctions Autostigmate et Autocomactions réussissent leur première itération. Si nécessaire, une autre zone peut être sélectionnée (par exemple, une feuille de carbone de support sans glace), une mise au point imagée ajustée ou un temps d’acquisition d’image augmenté pour optimiser le rapport signal/bruit dans les images acquises et la visibilité de plusieurs anneaux de Thon dans la transformée de Fourier des images acquises.
Les microscopes cryo-EM modernes génèrent de grandes quantités de données qui dépassent souvent 1 To par ensemble de données pour réaliser des reconstructions 3D haute résolution, en particulier pour les protéines à faible symétrie. Les données et les résultats de cryo-EM sont également généralement complétés par des données et des résultats de méthodes orthogonales permettant de bien comprendre les relations structure-fonction dans chaque projet scientifique. L’organisation des données collectées, leur transfert dans un pipeline de traitement d’images et le partage d’une reconstruction cryo-EM résultante entre collaborateurs imposent des exigences supplémentaires aux nouveaux utilisateurs de la méthodologie cryo-EM pour mettre en place leur infrastructure informatique locale. Les logiciels de gestion de données, tels qu’Athena, facilitent le stockage centralisé des données acquises par tout instrument connecté ou logiciel exploité par un utilisateur enregistré. Les données et métadonnées stockées sont accessibles à l’aide d’une interface de navigateur Web simple par plusieurs utilisateurs, qui peuvent avoir différents rôles dans le projet avec des droits d’accès différents (en tant que propriétaire, collaborateur ou visualiseur) en fonction de leurs informations de connexion et de leur définition de partage de données dans la configuration de l’expérience. Cette numérisation des flux de travail expérimentaux fournit des moyens de partage de données et de métadonnées entre collaborateurs sans duplication inutile et augmente la productivité et la traçabilité des flux de travail utilisés. La mise en œuvre d’une structure générale et personnalisable de projets, d’expériences et de flux de travail dans un logiciel de gestion de données est universelle et permet la personnalisation et l’intégration d’expériences orthogonales à l’aide de méthodes complémentaires dans une base de données de projet unique.
La sélection des zones pour la collecte de données sur une grille cryo-EM est essentielle pour l’acquisition réussie d’ensembles de données à haute résolution. Les grilles cryo-EM produites avec des dispositifs traditionnels de congélation par immersion, tels que le Vitrobot (un système de vitrification entièrement automatisé), affichent généralement un gradient d’épaisseur de glace sur la surface de la grille (Figure 4). Cela peut être bénéfique car la grille contient des zones avec différentes épaisseurs de glace; toutefois, les zones ayant l’épaisseur de glace idéale pour la collecte de données doivent être identifiées comme décrit dans le protocole ci-dessus. Une grille cryo-EM optimale doit contenir le moins de contamination de glace de transfert possible et contenir suffisamment de carrés de grille avec une feuille de support trouée intacte. La collecte de données sur les carrés de grille qui ont des fissures ou des zones cassées n’est pas recommandée car les images collectées seront affectées par une dérive globale beaucoup plus forte lors de l’éclairage par un faisceau d’électrons par rapport aux carrés de grille avec une feuille de support intacte. L’excès de glace cristalline peut obstruer la majorité des trous de feuille et / ou interférer avec la mise au point automatique et de tels carrés de grille doivent également être évités. Les carrés de grille avec de la glace mince présentent généralement de grandes zones vitrées et de nombreux trous de feuille brillants qui sont visibles dans une image prise à l’aide du préréglage Atlas. L’apparition de glace plus épaisse à proximité des barres de grille est à prévoir et non critique car les trous de feuille dans ces zones du carré de la grille sont exclus pendant la procédure de sélection des trous. La présence de plusieurs trous vides dans un carré de grille peut signifier que la glace vitrée dans les trous environnants est extrêmement mince et peut contenir des particules endommagées ou pas de particules du tout. En règle générale, il est sage de choisir des carrés de grille avec une variété d’épaisseurs de glace à différentes régions de la grille pour le dépistage et l’évaluation initiaux afin de comprendre quelles zones ont les meilleures conditions pour la collecte de données à haute résolution et présentent la densité de particules et la distribution d’orientation idéales. Pour les échantillons de protéasomes apoF et 20S utilisés dans cette étude, les zones avec la glace observable la plus mince contiennent les meilleures conditions pour l’imagerie à haute résolution de ces échantillons.
Lors de la sélection automatique de trous dans tous les carrés de grille sélectionnés à l’aide du logiciel de collecte de données, il est conseillé d’effectuer une tâche d’exécution de modèle sur un trou représentatif dans chaque carré de grille pour vérifier et s’assurer que ni les carrés trop épais, trop minces ou inattendus non vitreux n’ont été choisis pour la collecte de données. Lors de l’acquisition des données, les indicateurs de qualité clés des images collectées, tels que la dérive de l’image et l’ajustement CTF, peuvent être surveillés à l’aide de l’EQM. La collecte de données peut ensuite être optimisée en sautant des zones qui produisent des images de mauvaise qualité. Cependant, les images avec des ajustements CTF haute résolution peuvent toujours contenir des images avec des particules dans quelques orientations préférées ou des particules dénaturées dans une couche de glace trop mince. La sélection de particules en temps réel et la classification 2D à partir d’images collectées fourniraient des informations supplémentaires sur la qualité des données structurelles dans les particules imagées et révéleraient à la fois les orientations préférées des particules intactes dans la glace ou la structure incohérente des particules (partiellement) dénaturées. Le calcul des moyennes de classe peut donc aider à affiner davantage les régions appropriées pour la collecte de données, comme cela a déjà été mis en œuvre et montré dans d’autres progiciels23,28.
La sélection des paramètres d’imagerie pour l’acquisition de données, tels que le grossissement, le débit de dose d’électrons et la plage de mise au point, dépend de plusieurs critères, tels que la résolution cible, la taille de la protéine, la concentration de l’échantillon, le débit souhaité du microscope, etc. Pour la caméra à détecteur d’électrons direct utilisée dans ces expériences, le débit de dose d’électrons a été choisi dans la gamme de 4-5 e-/pix/s en sélectionnant une taille et une intensité SPOT appropriées pour maintenir un éclairage parallèle. Comme le montre le tableau 1, une taille SPOT différente peut être utilisée dans le préréglage Trou/Hauteur eucentrique pour assurer un rapport signal/bruit suffisant dans l’image pour le centrage des trous pendant la collecte des données. Le grossissement doit être choisi de telle sorte que la taille des pixels soit au moins 2 à 3 fois inférieure à la résolution cible pour la reconstruction cryo-EM. Cependant, plus le grossissement est élevé (c’est-à-dire une taille de pixel plus petite), plus le champ de vision est capturé dans les images et il y a moins de particules par image, ce qui entraîne finalement un temps de collecte de données plus long pour collecter des images avec suffisamment de particules pour reconstruire des cartes 3D à haute résolution. Pour l’échantillon apoF, nous avons utilisé la taille de pixel de 0,43 Å car nous avions une concentration d’échantillon suffisante pour une densité élevée de particules dans les images et une résolution inférieure à 2 Å de la reconstruction. Pour l’échantillon de protéasome 20S, nous avons utilisé la taille de pixel de 0,68 Å pour couvrir un champ de vision plus large dans les images acquises. Généralement, pour les microscopes TEM de 200 kV, les images cryo-EM sont acquises dans la plage de mise au point de 0,8 à 2,0 μm. Cependant, grâce à l’amélioration du contraste et du rapport signal/bruit à l’aide du filtre d’énergie, les acquisitions de données peuvent être effectuées beaucoup plus près de la mise au point pour mieux préserver les informations haute résolution dans les images acquises en raison de plus petites aberrations et de la réduction correspondante de la fonction d’enveloppe CTF. Nous n’utilisons pas non plus d’ouverture objective car l’ouverture peut introduire des aberrations d’image supplémentaires alors que le contraste de l’image est déjà suffisamment amélioré en utilisant le filtre d’énergie. Pour les échantillons de protéasome apoF et 20S, nous avons utilisé les paramètres de mise au point de 0,5 μm, 0,7 μm et 0,9 μm. Pour les protéines plus petites (<200 kDa), nous avons utilisé des paramètres de mise au point de -0,5 μm, -0,7 μm et -0,9 μm pour améliorer le contraste des particules et faciliter la sélection des particules et l’alignement grossier initial dans l’étape de raffinement 3D de la reconstruction 3D, ce qui a conduit à des cartes 3D de résolution ~ 2,5 Å (résultats non publiés).
Nous avons déjà montré que l’imagerie avec un filtre à énergie améliore le rapport signal/bruit (SNR) dans les images cryo-EM collectées sur les microscopes TEM 300 kV haut de gamme11. En fait, lorsque les électrons traversent un échantillon, deux principaux types d’interactions se produisent: i) les électrons dispersés de manière élastique maintiennent leur énergie et contribuent à la formation d’images en interférant avec le faisceau incident non diffusé via le mécanisme de contraste de phase ii) les électrons diffusés de manière inélastique perdent de l’énergie dans l’échantillon et contribuent principalement au bruit dans les images. Par conséquent, le SNR peut être considérablement amélioré en filtrant les électrons dispersés de manière inélastique, qui ont une énergie inférieure à celle du faisceau incident et des électrons diffusés de manière élastique, à l’aide d’une fente d’énergie étroite. Cependant, il est essentiel d’utiliser un filtre d’énergie suffisamment stable, tel que le Selectris ou le Selectris-X, pour pouvoir utiliser des fentes très étroites (10 eV ou moins) sur de longues (plus de 12 heures) d’acquisition automatisée de données cryoEM haute résolution.
Les images cryo-EM acquises avec des microscopes TEM de 200 kV dans les mêmes conditions qu’avec des microscopes TEM de 300 kV présentent un SNR plus petit à haute résolution (en particulier <4 Å) en raison de la désintégration plus rapide des fonctions de l’enveloppe CTF. Par conséquent, un plus grand nombre de particules (et donc un nombre plus élevé d’images collectées) est nécessaire pour atteindre une certaine résolution lors de l’utilisation de TEM de 200 kV. En outre, la profondeur de champ (10-25 nm dans la plage de résolution 2-3 Å) est également environ 20% plus petite dans les images 200 kV35, ce qui signifie que moins de particules dans la couche de glace (généralement 20-50 nm d’épaisseur) seront entièrement mises au point et contribueront de manière constructive à toutes les caractéristiques haute résolution d’une reconstruction 3D calculée, à moins que les valeurs de défocalisation ne soient affinées pour chaque particule indépendamment dans les étapes ultérieures de la procédure de reconstruction 3D. Pour les particules plus grosses (telles que les virions icosaédriques ou d’autres assemblages macromoléculaires), la taille des particules peut dépasser la profondeur de champ à haute résolution et introduire des erreurs de phase dues à l’approximation planaire de la sphère d’Ewald dans les algorithmes de reconstruction 3D standard36. Ces erreurs peuvent être affinées par des algorithmes avancés qui sont déjà implémentés dans les packages de traitement d’image cryo-EM courants 37,28,39. Comme la sphère d’Ewald a une courbure plus grande dans les données de 200 kV que dans les données de 300 kV, la correction de la sphère d’Ewald est nécessaire à des résolutions relativement plus faibles et/ou pour des assemblages macromoléculaires relativement plus petits lors de l’utilisation de TEM de 200 kV. D’autre part, les images de 200 kV présentent un contraste plus élevé de particules dans la glace mince (20-50 nm) qui est significativement plus mince que le chemin libre moyen de l’électron de 200-300 keV (220-280 nm). Le contraste plus élevé contribue à améliorer l’alignement global correct des particules individuelles, en particulier pour la diffusion faible de protéines plus petites dont la structure n’est pas encore connue, et le modèle de référence 3D n’est pas encore bien établi.
Ici, nous avons démontré sur l’exemple d’un protéasome 20S que le contraste et la qualité de l’image pouvaient être améliorés de la même manière avec un filtre à énergie lors de l’utilisation d’un microscope TEM 200 kV. En utilisant le même nombre de particules, les données collectées à l’aide de la fente de 20 eV ont été reconstruites à une résolution de 2,26 Å par rapport aux données collectées avec la fente d’énergie entièrement ouverte qui n’a été reconstruite qu’à une résolution de 2,34 Å. La meilleure reconstruction a été obtenue à partir des données collectées à l’aide de la fente de 10 eV qui a été reconstruite à une résolution de 2,14 Å. Ces résultats concordent avec la prédiction théorique selon laquelle le filtrage des électrons diffusés de manière inélastique augmente le SNR dans les images collectées et facilite une résolution plus élevée dans les reconstructions cryo-EM à partir du nombre donné de particules, comme résumé dans le tableau 4. Ces résultats ont été confirmés par les facteurs B calculés à partir de ces ensembles de données qui indiquent une meilleure qualité d’images dans les ensembles de données filtrés par l’énergie.
Nous pouvons donc conclure que si les microscopes TEM 300 kV offrent le débit le plus élevé et la résolution la plus élevée possible dans les reconstructions cryo-EM, les microscopes TEM 200 kV fournissent également des ensembles de données de haute qualité pour les reconstructions cryo-EM à haute résolution. Nous avons montré ici que la qualité des images acquises, et donc le temps global de structure, peut être encore améliorée en utilisant le TEM 200 kV équipé d’un filtre d’énergie et d’un détecteur d’électrons direct. Le protocole présenté décrit toutes les étapes nécessaires pour obtenir régulièrement des données cryo-EM à haute résolution à l’aide de cette configuration et révéler de fins détails structurels des structures 3D macromoléculaires, qui sont essentiels pour comprendre les relations structure-fonction clés en biologie structurale et en conception de médicaments basés sur la structure.
Déclarations de divulgation
Sagar Khavnekar ne signale aucun conflit d’intérêts. Les autres auteurs sont des employés de Thermo Fisher Scientific, la division MSD-EM.
Remerciements
Aucun.
matériels
| Name | Company | Catalog Number | Comments |
| AutoGrid rings | Thermo Fisher Scientific | 1036173 | Package of 100x AutoGrid rings for the standard EM grids. |
| C-Clip | Thermo Fisher Scientific | 1036171 | Package of 100 clips that secure the standard EM grids inside the AutoGrid rings. |
| Data Management Platform | Thermo Fisher Scientific | 1160939 | Part of the Glacios base configuraiton; includes Athena Software |
| EPU Quality Monitor | Thermo Fisher Scientific | 1179770 | |
| EPU Software | Thermo Fisher Scientific | 1025080 | Part of the Glacios base configuration |
| Ethane 3.5 | Westfalen | A06010110 | Ethane gas used for making liquid ethane (puritiy at least N35, i.e. 99.95% vol) |
| Falcon 4 200kV | Thermo Fisher Scientific | 1166936 | Direct electron detector |
| Glacios | Thermo Fisher Scientific | 1149551 | 200 kV TEM |
| GloQube Plus Glow Discharge System for TEM Grids and surface modification | Quorum | N/A | also available via Thermo Fisher Scientific (PN 1160602) |
| QuantiFoil grids | Quantifoil | N/A | R-2/1, 300 mesh; carbon foil grid |
| Relion | MRC Laboratory of Molecular Biology | N/A | open source software: https://relion.readthedocs.io/en/release-3.1/ |
| Selectris with Falcon 4 for 200 kV | Thermo Fisher Scientific | 1191753 | Energy filter |
| Selectris X with Falcon 4 for 200 kV | Thermo Fisher Scientific | 1191755 | Energy filter |
| UltrAuFoil grids | Quantifoil | N/A | R-1.2/1.2, 300 mesh; gold foil grids |
| Vitrobot Mk. IV | Thermo Fisher Scientific | 1086439 | Automated vitrification system |
| Whatman 595 filter paper | Thermo Fisher Scientific | AA00420S |
Références
- Wang, J., Hua, T., Liu, Z. -. J. Structural features of activated GPCR signaling complexes. Current Opinions in Structural Biology. 63, 82-89 (2020).
- Chen, S., Gouaux, E. Structure and mechanism of AMPA receptor - auxiliary protein complexes. Current Opinions in Structural Biology. 54, 104-111 (2019).
- Kühlbrandt, W. Structure and mechanisms of F-Type ATP synthases. Annual Review of Biochemistry. 88, 515-549 (2019).
- Laverty, D., et al. Cryo-EM structure of the human α1β3γ2 GABA A receptor in a lipid bilayer. Nature. 565 (7740), 516-520 (2019).
- Liu, F., Zhang, Z., Csanády, L., Gadsby, D. C., Chen, J. Molecular structure of the Human CFTR ion channel. Cell. 169 (1), 85-95 (2017).
- Wrapp, D., et al. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science. 367 (6483), 1260-1263 (2020).
- Ke, Z., et al. Structures and distributions of SARS-CoV-2 spike proteins on intact virions. Nature. 588, 498-502 (2020).
- Yan, R., Zhang, Y., Li, Y., Xia, L., Guo, Y., Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science. 367 (6485), 1444-1448 (2020).
- Walls, A. C., Park, Y. -. J., Tortorici, M. A., Wall, A., McGuire, A. T., Veesler, D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell. 180, 281-292 (2020).
- Chiba, S., et al. Multivalent nanoparticle-based vaccines protect hamsters against SARS-CoV-2 after a single immunization. Communications Biology. 4 (1), 597 (2021).
- Nakane, T., et al. Single-particle cryo-EM at atomic resolution. Nature. 587, 152-156 (2020).
- Bai, X. C. Seeing atoms by single-particle Cryo-EM. Trends in Biochemical Sciences. 46 (4), 253-254 (2021).
- Koh, A., et al. High-resolution cryo-EM at 200kV enabled by Selectris-X and Falcon 4. Microscience Microscopy Congress 2021. , (2021).
- Herzik, M. A., Wu, M., Lander, G. C. Achieving better-than-3-Å resolution by single-particle cryo-EM at 200 keV. Nature Methods. 14 (11), 1075-1078 (2017).
- Herzik, M. A., Wu, M., Lander, G. C. High-resolution structure determination of sub-100 kDa complexes using conventional cryo-EM. Nature Communications. 10 (1), 1032 (2019).
- Wu, M., Lander, G. C., Herzik, M. A. Sub-2 Angstrom resolution structure determination using single-particle cryo-EM at 200 keV. Journal of Structural Biology X. 4, 100020-100029 (2020).
- Mori, T., et al. C-Glycoside metabolism in the gut and in nature: Identification, characterization, structural analyses and distribution of C-C bond-cleaving enzymes. Nature Communications. 12 (1), 6294 (2021).
- Hamdi, F., et al. 2.7 Å cryo-EM structure of vitrified M. musculus H-chain apoferritin from a compact 200 keV cryo-microscope. PLoS One. 15 (5), 0232540 (2020).
- Abbott, S., et al. EMDB web resources. Current Protocols in Bioinformatics. 61 (1), 1-12 (2018).
- Punjani, A., Rubinstein, J. L., Fleet, D. J., Brubaker, M. A. CryoSPARC: algorithms for rapid unsupervised cryo-EM structure determination. Nature Methods. 14 (3), 290-296 (2017).
- Scheres, S. H. W. RELION: Implementation of a Bayesian approach to cryo-EM structure determination. Journal of Structural Biology. 180 (3), 519-530 (2012).
- Gómez-Blanco, J., et al. Using Scipion for stream image processing at Cryo-EM facilities. Journal of Structural Biology. 204 (3), 457-463 (2018).
- Tegunov, D., Cramer, P. Real-time cryo-electron microscopy data preprocessing with Warp. Nature Methods. 16 (11), 1146-1152 (2019).
- Lander, G. C., et al. Appion: an integrated, database-driven pipeline to facilitate EM image processing. Journal of Structural Biology. 166 (1), 95-102 (2009).
- Carragher, B., et al. Current outcomes when optimizing 'standard' sample preparation for single-particle cryo-EM. Journal of Microscopy. 276 (1), 39-45 (2019).
- Drulyte, I., et al. Approaches to altering particle distributions in cryo-electron microscopy sample preparation. Acta Crystallographica. Section D, Structural Biology. 74 (6), 560-571 (2018).
- Bloch, M., Santiveri, M., Taylor, N. M. I. Membrane protein Cryo-EM: Cryo-grid optimization and data collection with protein in detergent. Methods in Molecular Biology. 2127, 227-244 (2020).
- Cheng, A., et al. Leginon: New features and applications. Protein Science. 30 (1), 136-150 (2021).
- Schorb, M., et al. Software tools for automated transmission electron microscopy. Nature Methods. 16 (6), 471-477 (2019).
- Li, X., et al. Electron counting and beam-induced motion correction enable near-atomic-resolution single-particle cryo-EM. Nature Methods. 10 (6), 584-590 (2013).
- Alewijnse, B., et al. Best practices for managing large CryoEM facilities. Journal of Structural Biology. 199, 225-236 (2017).
- Baldwin, P. R., et al. Big data in cryoEM: automated collection, processing and accessibility of EM data. Current Opinion in Microbiology. 43, 1-8 (2018).
- Guo, H., et al. Electron-event representation data enable efficient cryoEM file storage with full preservation of spatial and temporal resolution. International Union of Crystallography Journal. 7, 860-869 (2020).
- Weis, F., Hagen, W. J. H. Combining high throughput and high quality for cryo-electron microscopy data collection. Acta Crystallographica. Section D, Structural Biology. 76 (8), 724-728 (2020).
- Zhou, H., Chiu, W. Determination of icosahedral virus structures by electron cryomicroscopy at subnanometer resolution. Advances in Protein Chemistry. 64, 93-124 (2005).
- DeRosier, D. J. Correction of high-resolution data for curvature of the Ewald sphere. Ultramicroscopy. 81 (2), 83-98 (2000).
- Wolf, M., DeRosier, D. J., Grigorieff, N. Ewald sphere correction for single-particle electron microscopy. Ultramicroscopy. 106 (4-5), 376-382 (2006).
- Leong, P. A., Yu, X., Hong Zhou, Z., Jensen, G. J. Correcting for the ewald sphere in high-resolution single-particle reconstructions. Methods in Enzymology. 482, 369-380 (2010).
- Zivanov, J., et al. New tools for automated high-resolution cryo-EM structure determination in RELION-3. eLife. 7, 42166 (2018).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationThis article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.