
Method Article
Passage rapide, rentable et sans enzyme de cellules souches pluripotentes humaines sur des cellules nourricières par désadhérence médiée par l’acide éthylènediaminetétraacétique
Dans cet article
Résumé
Afin d’éviter les limitations associées au passage enzymatique ou mécanique des cellules souches embryonnaires humaines (CSEh) et des cellules souches pluripotentes induites humaines (CSPhi) cultivées sur des cellules nourricières, nous avons mis au point une méthode rapide, efficace, rentable et à haut rendement pour la récolte de colonies de CSEh ou de CSPhi, maintenues sur une couche de cellules nourricières de fibroblastes de prépuce humain en utilisant la désadhésion médiée par l’EDTA.
Résumé
Les cellules souches pluripotentes humaines (cellules souches embryonnaires humaines, CSEh, et cellules souches pluripotentes induites humaines, CSPhi) ont été cultivées à l’origine sur différents types de cellules nourricières pour être maintenues à l’état indifférencié dans une culture à long terme. Cette approche a été largement supplantée par les protocoles de culture sans alimentateur, mais ceux-ci impliquent des réactifs plus coûteux et peuvent favoriser une transition vers un état d’amorçage, ce qui limite la capacité de différenciation des cellules. Dans les conditions d’alimentation et sans mangeoire, la récolte des colonies hESC ou hiPSC pour le passage est une procédure nécessaire pour étendre les cultures.
Afin de fournir une procédure simple et à haut rendement pour le passage des hESCs/hiPSCs cultivées sur des cellules nourricières, nous avons mis au point une méthode de récolte utilisant la désadhérence provoquée par l’acide chélateur de calcium éthylènediaminetétraacétique (EDTA). Nous avons évalué le rendement et la qualité des cellules traversées résultantes en comparant cette approche à l’approche originale de la récolte mécanique, dans laquelle les colonies sont isolées avec un scalpel sous un microscope (la récolte mécanique a été choisie comme comparateur pour éviter la variabilité des réactifs associée à la récolte enzymatique).
Dans une série d’expériences, deux lignées hESC différentes ont été maintenues sur une couche de cellules nourricières de fibroblastes de prépuce humain. Chaque lignée a été soumise à de multiples passages à l’aide d’un prélèvement à base d’EDTA ou mécanique et évaluée pour la taille et la morphologie de la colonie, la densité cellulaire, l’expression des marqueurs de tige, la différenciation vers les trois couches germinales dans les corps embryoïdes et les aberrations génomiques. Dans une autre série d’expériences, nous avons utilisé la récolte basée sur l’EDTA sur deux lignées hiPSC différentes et avons obtenu des résultats similaires. La désadhérence induite par l’EDTA a permis de gagner du temps et d’obtenir un rendement plus élevé de colonies de taille plus favorable et de morphologie plus uniforme par rapport à la récolte mécanique. Il était également plus rapide que la récolte enzymatique et n’était pas sujet à la variabilité des lots enzymatiques. La méthode de désadhérence induite par l’EDTA facilite également le transfert des lignées hESC/hiPSC de la culture à base de cellules nourricières vers des conditions sans nourrisseur si souhaitée pour une utilisation et une analyse en aval.
Introduction
Le bon maintien des CSEh et des CSPhi in vitro est une méthodologie fondamentale et pratique pour plusieurs pistes de recherche en biologie cellulaire humaine et en biologie du développement. En raison de la tendance inhérente des hESCs et des hiPSCs à se différencier, le maintien de l’état indifférencié in vitro exige un soin et une attention particuliers. Ainsi, la mise au point de protocoles rentables pour la maintenance et le passage des hESC et des hiPSC avec le moins de variabilité méthodologique possible est d’une grande utilité générale.
À l’origine, les hESCs et les hiPSCs ont été cultivés sur différents types de cellules nourricières pour aider à la culture et au maintien à long terme de l’état indifférencié 1,2,3. Plus récemment, la culture dans des conditions sans mangeoire est devenue la norme, car elle évite complètement de traiter avec des cellules nourricières4. Cependant, certains laboratoires et installations centrales cultivent encore des CSEh ou des CSPhi sur des cellules nourricières. La culture sans mangeoire est plus coûteuse car elle nécessite l’utilisation de milieux de culture de compositions spéciales et d’une certaine forme de revêtement de la surface de culture pour assurer l’adhérence de la colonie (composants majeurs de la matrice extracellulaire [MEC] ou un composé commercial de MEC, ou utilisation de plaques revêtues disponibles dans le commerce). La dépense n’est pas négligeable et représente un obstacle financier potentiel pour certains laboratoires intéressés à poursuivre la recherche et le développement basés sur les CSE ou les CSPh. De plus, la culture dans des conditions sans mangeoire a tendance à conduire les hESCs et les hiPSCs à un état moins naïf que celui maintenu sur les cellules nourricières5, ce qui peut compromettre la différenciation ultérieure et conduire à des variations génétiques6.
Historiquement, le passage des hESCs et des hiPSCs cultivés sur des cellules nourricières impliquait une récolte mécanique - à l’aide d’un scalpel pour exciser les colonies au microscope7 - mais cela a ensuite été largement supplanté par la digestion enzymatique avec ou sans grattage doux pour isoler les colonies ou les cellules dissociées. La récolte mécanique est fastidieuse et nécessite une microchirurgie de précision. L’efficacité de la récolte enzymatique peut varier en raison des différences enzymatiques d’un lot à l’autre et tend à favoriser la dissociation complète, ce qui favorise la mort cellulaire à moins d’être contrecarrée par les inhibiteurs de ROCK 8,9 et augmente l’incidence des caryotypes anormaux 9.
Pour tirer parti des coûts réduits et du plus grand potentiel de différenciation de la culture des hESC et des hiPSC sur les cellules nourricières tout en évitant les inconvénients de la récolte mécanique et enzymatique, nous avons mis au point une méthode rapide, efficace, rentable et à haut rendement pour la récolte des colonies de hESC et d’hiPSC maintenues sur une couche nourricière de fibroblastes humains du prépuce en utilisant la désadhérence médiée par EDTA. Nous avons comparé le rendement, la variabilité et la qualité des cellules souches à ceux obtenus avec la récolte mécanique (nous n’avons pas comparé à la digestion enzymatique en raison de la variabilité supplémentaire que cette approche implique). Nous remarquons que la désadhérence médiée par l’EDTA fonctionne également bien pour le transfert de colonies d’une culture basée sur des mangeoires vers des conditions sans nourrisseurs, si cela est souhaité pour une utilisation et des analyses en aval. Cette méthode permet d’assurer une transition avec une méthode de passage cohérente, car la désadhérence induite par l’EDTA est une approche populaire utilisée pour les cultures sans alimentateur.
Protocole
Reportez-vous au tableau des matériaux pour plus de détails sur tous les matériaux, réactifs et instruments utilisés dans ce protocole.
1. Culture de cellules de fibroblastes humains et préparation de la couche de cellules nourricières
- Semez 0,5 × 106 fibroblastes humains du prépuce (ci-après appelés « cellules nourricières ») par fiole de culture T-75 (nombre de flacons selon les besoins) avec 20 mL de milieu Dulbecco modifié (IMDM) d’Iscove avec (p/) 10 % de sérum de veau fœtal (FBS), ci-après appelé « milieu de cellules nourricières ».
- Lorsque les cellules nourricières atteignent 90 % de confluence, retirez le milieu et lavez-les 3 fois avec 10 mL de solution saline tamponnée au phosphate (DPBS) de Dulbecco par flacon pour éviter l’inhibition de la trypsine par les facteurs présents dans le milieu. Ajouter 2 mL de trypsine-EDTA dans chaque fiole et placer la ou les fioles dans un incubateur à 37 °C/5 % de CO2 pendant 5 minutes ou jusqu’à ce que les cellules nourricières se soient détachées de la ou des fioles. Observez le détachement des cellules au microscope sous forme d’agrégats flottants de cellules ou de cellules individuelles.
- Ajouter 5 mL de milieu de cellule nourricière frais préchauffé dans chaque fiole pour inactiver la trypsine-EDTA, et suspendre doucement les cellules nourricières par pipetage.
- Transférez les cellules d’alimentation dans un tube à centrifuger de 15 ml. Bouchez le tube et granulez les cellules d’alimentation par centrifugation à 200 × g pendant 5 min.
- Retirez délicatement le surnageant sans perturber la pastille de la cellule nourricière. Ensuite, remettez soigneusement la pastille en suspension dans 4 mL de milieu de cellule d’alimentation frais. Assurez-vous que les cellules nourricières sont bien remises en suspension avant de les dénombrer à l’aide d’une chambre de comptage de cellules ou d’un autre appareil de comptage de cellules.
- Ajouter 0,5 × 106 cellules nourricières au nombre requis de nouvelles fioles de culture T-75 pour l’expansion, et ajouter 20 mL de milieu de cellule nourricière fraîche dans chaque flacon. Incuber le(s) flacon(s) de culture dans un incubateur à 37 °C/5 % de CO2 jusqu’à ce que les cellules nourricières aient atteint 90 % de confluence.
REMARQUE : Les cellules nourricières peuvent être utilisées au moins jusqu’au passage 25. - Calculer le nombre de cellules nourricières nécessaires pour le nombre de boîtes de culture tissulaire de 35 mm qui seront utilisées pour la culture des CSEh/CSPhi.
REMARQUE : Habituellement, 3,0 × 105 cellules nourricières par boîte de culture tissulaire sont suffisantes pour générer une couche confluente de cellules nourricières. - Pour éviter la prolifération des cellules nourricières, assurez-vous qu’elles sont arrêtées mitotiquement de l’une ou l’autre des deux manières.
REMARQUE : Pour les deux méthodes, un grand lot de cellules nourricières arrêtées mitotiquement peut être généré et congelé dans des aliquotes pour une utilisation ultérieure.- Effectuer l’arrêt mitotique par irradiation gamma en transférant toutes les cellules nourricières nécessaires dans un tube à centrifuger de 50 mL et en complétant avec le milieu de la cellule nourricière jusqu’à un volume total de 5 mL. Transporter immédiatement à température ambiante vers un appareil d’irradiation gamma, et irradier pour arrêter mitotiquement les cellules nourricières (300 kV et 10 mA pendant 20 min).
REMARQUE : Un retard dans le transport peut entraîner une fixation indésirable des cellules d’alimentation à la paroi du tube à centrifuger de 50 mL. Si le transport dure plus de quelques minutes, assurez-vous que les cellules nourricières restent en suspension pendant le transport en agitant continuellement le tube. - Effectuer l’arrêt mitotique à l’aide de la mitomycine C en transférant toutes les cellules nourricières nécessaires dans 5 mL de milieu cellulaire nourricier dans un tube à centrifuger de 50 mL, puis ajouter 15 mL de milieu cellulaire nourricier contenant 20 μg/mL de mitomycine C, et incuber dans un incubateur à 37 °C/5 % de CO2 pendant 3 h. Ajouter 20 mL de PBS à 37 °C, granuler les cellules par centrifugation à 200 × g pendant 5 min, répéter le lavage PBS deux fois de plus et remettre en suspension dans le milieu de la cellule d’alimentation.
- Effectuer l’arrêt mitotique par irradiation gamma en transférant toutes les cellules nourricières nécessaires dans un tube à centrifuger de 50 mL et en complétant avec le milieu de la cellule nourricière jusqu’à un volume total de 5 mL. Transporter immédiatement à température ambiante vers un appareil d’irradiation gamma, et irradier pour arrêter mitotiquement les cellules nourricières (300 kV et 10 mA pendant 20 min).
- Une fois que les cellules nourricières ont été arrêtées mitotiquement, retournez dans la hotte de culture tissulaire et placez les cellules nourricières à 3,0 × 105 cellules par boîte de culture tissulaire de 35 mm, comme suit. Assurez-vous que les cellules nourricières sont complètement remises en suspension, ajoutez le milieu de la cellule nourricière pour atteindre une concentration de cellules nourricières de 1,5 × 105 par mL et ajoutez 2 mL de cette suspension de cellules nourricières dans chaque boîte de culture tissulaire de 35 mm.
- Transvaser les boîtes de culture dans un incubateur à 37 °C/5 % de CO2 . Pour assurer une répartition uniforme des cellules nourricières, déplacez les boîtes de culture lentement mais fermement sur l’étagère de l’incubateur vers l’avant et vers l’arrière 3 fois, suivies d’une pause, puis effectuez la même action de gauche à droite 3 fois. Ne déplacez plus la vaisselle et fermez doucement la porte de l’incubateur.
- Après 24 h, passer du milieu cellulaire nourricier à l’IMDM avec remplacement sérique (SR) à 10 %. Remplacez ce milieu par la suite tous les trois jours. Les cellules nourricières sont prêtes à l’emploi après les 3 premiers jours.
2. Récolte mécanique des colonies de hESC ou hiPSC
- Préchauffer le milieu hESC composé de 80 % de milieu d’aigle modifié de Dulbecco (DMEM), de 20 % de SR, de 1 mM de substitut de glutamine 100x, de 1 mM d’acides aminés non essentiels (NEAA), de 1 mM de pénicilline/streptomycine (P/S), de 0,1 mM de 2-mercaptoéthanol et de 10 ng/mL de facteur de croissance des fibroblastes basiques (bFGF). Le milieu hESC est utilisé pour la culture des hESC ou des hiPSC sur les cellules nourricières.
- Prélever des boîtes de culture tissulaire fraîches de 35 mm contenant des cellules nourricières arrêtées mitotiquement et remplacer le milieu de la cellule nourricière par 1,2 mL de milieu hESC contenant du bFGF au moins 30 minutes avant le transfert des colonies hESC/hiPSC.
- Placer une boîte de culture contenant des colonies hESC/hiPSC sur des cellules nourricières arrêtées mitotiquement sous un microscope avec un grossissement de 10x placé dans une hotte à flux laminaire. Utilisez un scalpel stérile pour couper soigneusement autour de la circonférence de chaque colonie, puis coupez chaque colonie en 5 à 6 morceaux à peu près égaux. Soulevez délicatement les morceaux de colonie avec la pointe de la lame du scalpel afin qu’ils se détachent de la couche de cellules nourricières et flottent librement dans le milieu.
- Essayez d’éviter les régions des colonies qui contiennent des cellules de différenciation, qui apparaissent comme des îlots de cellules plus petites avec des noyaux moins distincts par rapport aux hESCs/hiPSCs au sein d’une colonie.
- Transférez les colonies flottant librement à l’aide d’une pipette de 1 mL dans les nouvelles boîtes de culture contenant les cellules nourricières. Essayez de garder les colonies séparées afin qu’elles ne se développent pas les unes dans les autres plus tard. Déplacez soigneusement les boîtes de culture dans un incubateur cellulaire et évitez de les déranger jusqu’au lendemain.
- Le lendemain, ajouter délicatement 600 μL de milieu hESC contenant du bFGF jusqu’à l’obtention d’un volume final de 1,8 mL. Remplacez le milieu hESC + bFGF chaque jour par la suite jusqu’au prochain passage (généralement après 1 semaine).
3. Récolte des colonies de hESC ou de hiPSC par désadhérence médiée par EDTA
- Prenez des boîtes de culture fraîches avec des cellules nourricières arrêtées mitotiquement et passez de l’IMDM avec 10 % de SR à 1,2 mL de milieu hESC + bFGF préchauffé au moins 30 min avant le transfert des colonies.
- Manipulez une boîte de culture contenant des colonies de hESC ou de hiPSC à la fois. Retirez le milieu hESC + bFGF et lavez les colonies avec 1 mL de DPBS à température ambiante pour éliminer les éventuelles cellules non attachées et les débris cellulaires. Ajouter 1 mL d’EDTA 0,5 mM et incuber pendant 1 min à 37 °C. Si la hotte à flux laminaire est équipée d’une plaque chauffante, effectuez cette étape et les étapes de la section 4 sur la plaque chauffante pour une meilleure désadhérence.
- Après 1 min d’incubation, retirer la solution d’EDTA et ajouter délicatement 1 mL de milieu hESC + bFGF à l’aide d’une pipette de 1 mL. Triturer doucement avec la même pipette pour libérer les colonies de la couche de cellules nourricières. Continuez à triturer soigneusement jusqu’à ce que la couche de cellules nourricières se détache et se replie sur elle-même en une touffe séparée. Retirez la couche de la cellule d’alimentation avec l’embout de la pipette.
- Transférer les colonies hESC/hiPSC en suspension à l’aide d’une nouvelle pipette de 1 mL dans de nouvelles boîtes de culture contenant les cellules nourricières et le milieu hESC + bFGF, en les divisant dans un rapport de 1 :5. Les colonies ont tendance à se répartir uniformément dans chaque nouvelle boîte de culture, mais facilitent cela en déplaçant doucement la boîte d’un côté à l’autre. Remplacez le milieu hESC + bFGF chaque jour par la suite jusqu’au prochain passage (généralement après 1 semaine).
Résultats
Dans les essais et comparaisons documentés ci-dessous, nous avons utilisé deux lignées hESC (H9 et HS429, de WiCell et de l’Institut Karolinska, respectivement) et deux lignées hiPSC (NCS001 et NCS002, toutes deux générées par le Norwegian Core Facility for Human Pluripotent Stem Cells). Les données présentées dans les figures et les tableaux proviennent des lignées hESC, mais des résultats entièrement similaires ont été obtenus à partir des lignées hiPSC.
Entre nos mains, la récolte mécanique a entraîné la division des colonies en environ cinq à six touffes de ~200-250 μm de diamètre, tandis qu’avec la désadhérence induite par l’EDTA suivie d’une trituration, chaque colonie a été divisée en ~10-20 touffes de ~60 μm de diamètre. Nous estimons que le nombre de cellules dans chaque touffe récoltée par EDTA est de ~20. Comme il n’est pas pratique de diviser une colonie en amas de cette taille avec un scalpel, à cet égard, la désadhérence induite par l’EDTA est supérieure, car elle génère des amas d’une taille plus favorable à la survie des cellules de la colonie10,11.
Les colonies hESC/hiPSC récoltées à l’aide de l’EDTA étaient également plus homogènes en taille et en forme que les colonies récoltées mécaniquement (figure 1A-F). En effet, la coupe nécessaire à la récolte mécanique génère des bords inégaux et des tailles de touffes variables. Pour évaluer cela quantitativement, nous avons évalué la circularité de la colonie (en tant que mesure de l’arrondi des bords de la colonie ; une valeur de 1 indique un cercle parfait) 5 jours après le passage en utilisant le protocole ImageJ-win6412. La circularité des colonies était significativement plus faible dans les colonies récoltées mécaniquement (récolte mécanique : 0,61 ± 0,10 ; Récolte basée sur l’EDTA : 0,84 ± 0,01 ; n = 10, p < 0,001, test U de Mann-Whitney, U = 10).
La densité cellulaire dans les colonies récoltées et replaquées, qui est une mesure des interactions cellule-cellule après la récolte pendant la formation de la colonie, était similaire avec la récolte à base d’EDTA et la récolte mécanique (tableau 1 et figure 1G,H). Les colonies récoltées mécaniquement avaient une plus grande tendance à développer une nécrose dans leurs régions centrales (figure 1J). Cela était probablement dû à la variabilité de la forme et, en particulier, à la taille des amas cellulaires isolés mécaniquement, car lorsque ces amas sont trop grands, ils peuvent facilement se replier sur eux-mêmes lorsqu’ils sont transférés dans de nouvelles boîtes de culture. Ce n’était pas le cas des colonies récoltées à l’aide de l’EDTA, qui présentaient uniformément un aspect translucide avec des bords distincts (figure 1I).
Grâce à la récolte basée sur l’EDTA, nous avons pu collecter pratiquement toutes les colonies qui avaient été établies dans un puits en 2 à 3 minutes. En utilisant la récolte mécanique, rassembler toutes les colonies dans un puits serait fastidieux et prendrait beaucoup de temps. Nous n’avons généralement réussi à collecter que ~30%, soit ~20-25 colonies, en utilisant la récolte mécanique, et cela a pris ~20 min. De même, en utilisant la digestion de la collagénase suivie d’un grattage doux, il était généralement difficile de récolter toutes les colonies, bien que la procédure totale n’ait pris que quelques minutes. Ainsi, la récolte à base d’EDTA est aussi rapide ou plus rapide que la récolte enzymatique et plus efficace que la récolte mécanique ou enzymatique.
Afin d’évaluer l’effet des différentes méthodes de récolte sur la tige et la pluripotence, nous avons d’abord soumis les colonies obtenues après 20 passages par récolte à base d’EDTA ou mécanique à une analyse qPCR (Figure 2) et à une coloration immunocytochimique (Figure 3 et Figure 4) pour les marqueurs de tige. Les colonies obtenues à l’aide de l’une ou l’autre méthode présentaient une expression stable des marqueurs de tige à la fois au niveau de l’ARNm et des protéines. Nous avons ensuite évalué la pluripotence par différenciation des trois couches germinales dans les corps embryoïdes (Figure 5 et Figure 6). Les corps embryoïdes générés à partir des hESCs ou des hiPSCs obtenus après 20 passages à l’aide de l’une ou l’autre méthode contenaient un mélange de cellules exprimant des marqueurs couramment évalués pour l’ectoderme, le mésoderme et l’endoderme.
Enfin, nous avons évalué l’incidence des aberrations génomiques dans les hESCs et les hiPSCs passées par chaque méthode à l’aide d’une analyse génétique basée sur la qPCR (voir le tableau des matériaux). Les colonies obtenues après 20 passages en utilisant l’une ou l’autre méthode de récolte présentaient quelques exemples d’écart modeste par rapport à un profil chromosomique diploïde de référence (les anomalies évaluées étaient celles couramment associées à la reprogrammation des CSPhi, mais peuvent également être obtenues dans les CSEh) (Figure 7). Cependant, la tendance de ces écarts était essentiellement la même dans les colonies obtenues après l’une ou l’autre méthode de récolte, ce qui indique qu’ils n’étaient pas liés à la méthode de récolte.

Figure 1 : Morphologie des colonies et densité cellulaire après récolte à base d’EDTA ou mécanique. (A-F) Images représentatives en fond clair de colonies H9 hESC établies en culture sans mangeoire pendant 5 jours après 20 passages à l’aide d’une récolte à base d’EDTA (A-C) ou d’une récolte mécanique (D-F). (G,H) Images de fluorescence représentatives de la densité cellulaire dans les colonies H9 hESC établies après 20 passages à l’aide d’une récolte à base d’EDTA (G) ou d’une récolte mécanique (H). Les noyaux cellulaires sont colorés avec du DAPI. (I,J) Images représentatives en fond clair des colonies H9 hESC établies après 20 passages à l’aide d’une récolte (I) à base d’EDTA ou (J) mécanique. Remarquez la région centrale nécrotique de la colonie récoltée mécaniquement (flèche en J). Toutes les images ont été acquises 5 jours après le 20ème passage. Barres d’échelle = 100 μm. Abréviations : hESC = cellule souche embryonnaire humaine ; EDTA = acide éthylènediaminetétraacétique ; DAPI = 4',6-diamidino-2-phénylindole. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 2 : Expression de l’ARNm marqueur de la tige dans deux lignées hESC (H9 et HS429) générées après récolte à base d’EDTA ou mécanique. Réaction en chaîne par polymérase quantitative en temps réel des marqueurs indiqués dans les hESC H9 (panneau supérieur) et HS429 (panneau inférieur) après un seul passage par récolte mécanique, après 20 passages par récolte mécanique et après 20 passages par récolte à base d’EDTA (dilution 1 :5). Le niveau d’expression est relatif à celui du gène d’entretien ACTB (bêta-actine). Les barres d’erreur indiquent l’écart-type. Abréviations : hESC = cellule souche embryonnaire humaine ; EDTA = acide éthylènediaminetétraacétique. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 3 : Expression des protéines marqueurs de la tige dans la lignée H9 hESC après différentes conditions de récolte. Coloration par immunofluorescence représentative des colonies H9 hESC récoltées mécaniquement (A-E) avant un passage ultérieur, (F-J) après 20 passages en utilisant la récolte mécanique, et (K-O) après 20 passages en utilisant la récolte à base d’EDTA. Barres d’échelle = 100 μm. Abréviations : hESC = cellule souche embryonnaire humaine ; EDTA = acide éthylènediaminetétraacétique ; DAPI = 4',6-diamidino-2-phénylindole. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 4 : Expression des protéines marqueurs de la tige dans la lignée HS429 hESC après différentes conditions de récolte. Coloration par immunofluorescence représentative des colonies HS429 hESC récoltées mécaniquement (A-E) avant un nouveau passage, (F-J) après 20 passages par récolte mécanique et (K-O) après 20 passages par récolte à base d’EDTA. Barres d’échelle = 100 μm. Abréviations : hESC = cellule souche embryonnaire humaine ; EDTA = acide éthylènediaminetétraacétique ; DAPI = 4',6-diamidino-2-phénylindole. Veuillez cliquer ici pour voir une version agrandie de cette figure.
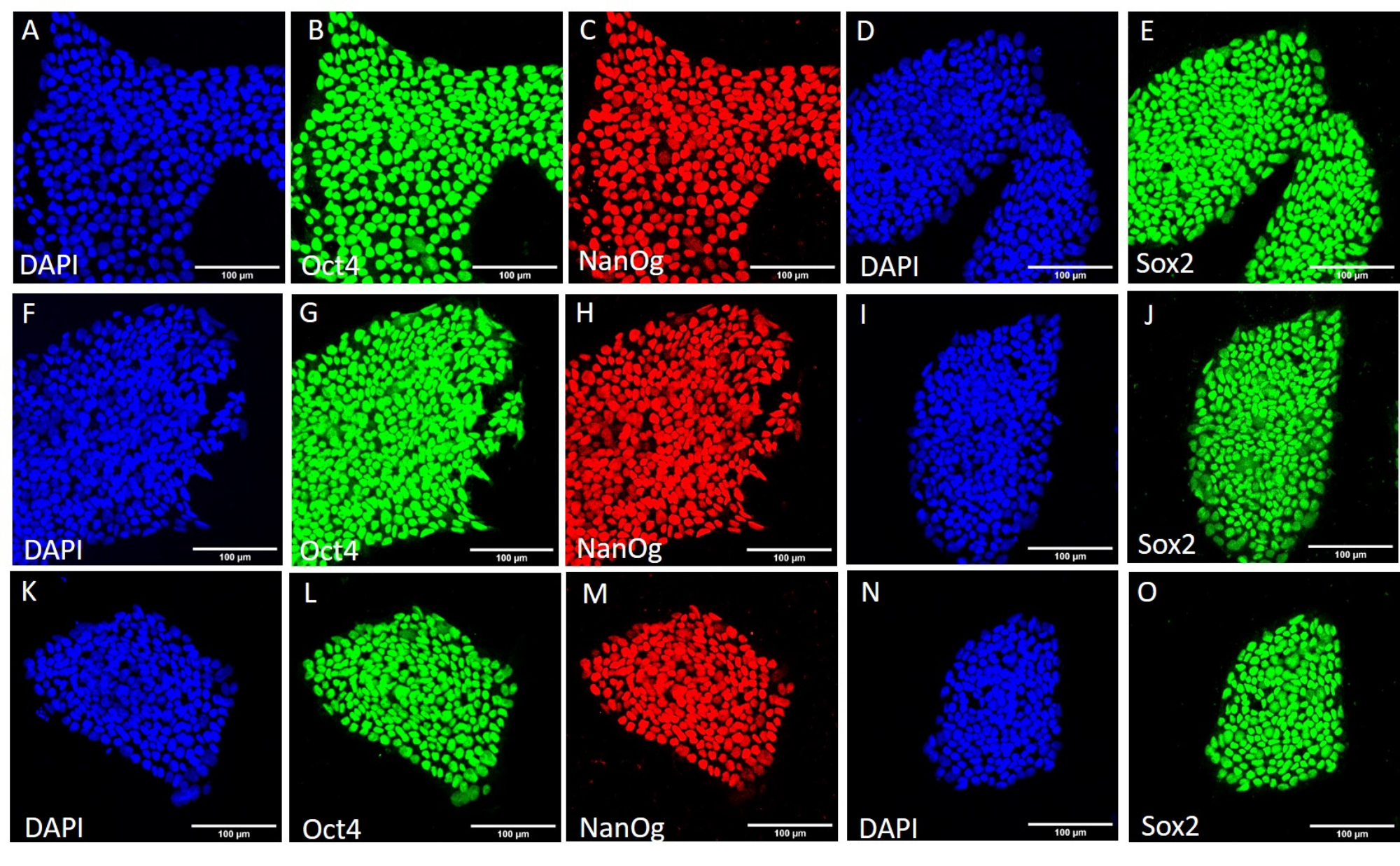{kind=link}

Figure 5 : Expression des marqueurs des trois couches germinales dans les corps embryoïdes générés à partir de la lignée H9 hESC après prélèvement mécanique ou à base d’EDTA. Marquage par immunofluorescence représentatif des marqueurs de l’ectoderme (rangées A et D) (ECTO, TUJI), de l’endoderme (rangées B et E) (ENDO, AFP) et du mésoderme (rangées C et F) (MESO, SMA). EB générés (A-C) après 20 passages de récolte mécanique ou (D-F) après 20 passages de récolte à base d’EDTA. Barres d’échelle = 40 μm. Abréviations : hESC = cellule souche embryonnaire humaine ; EDTA = acide éthylènediaminetétraacétique ; EB = corps embryoïdes. Veuillez cliquer ici pour voir une version agrandie de cette figure.
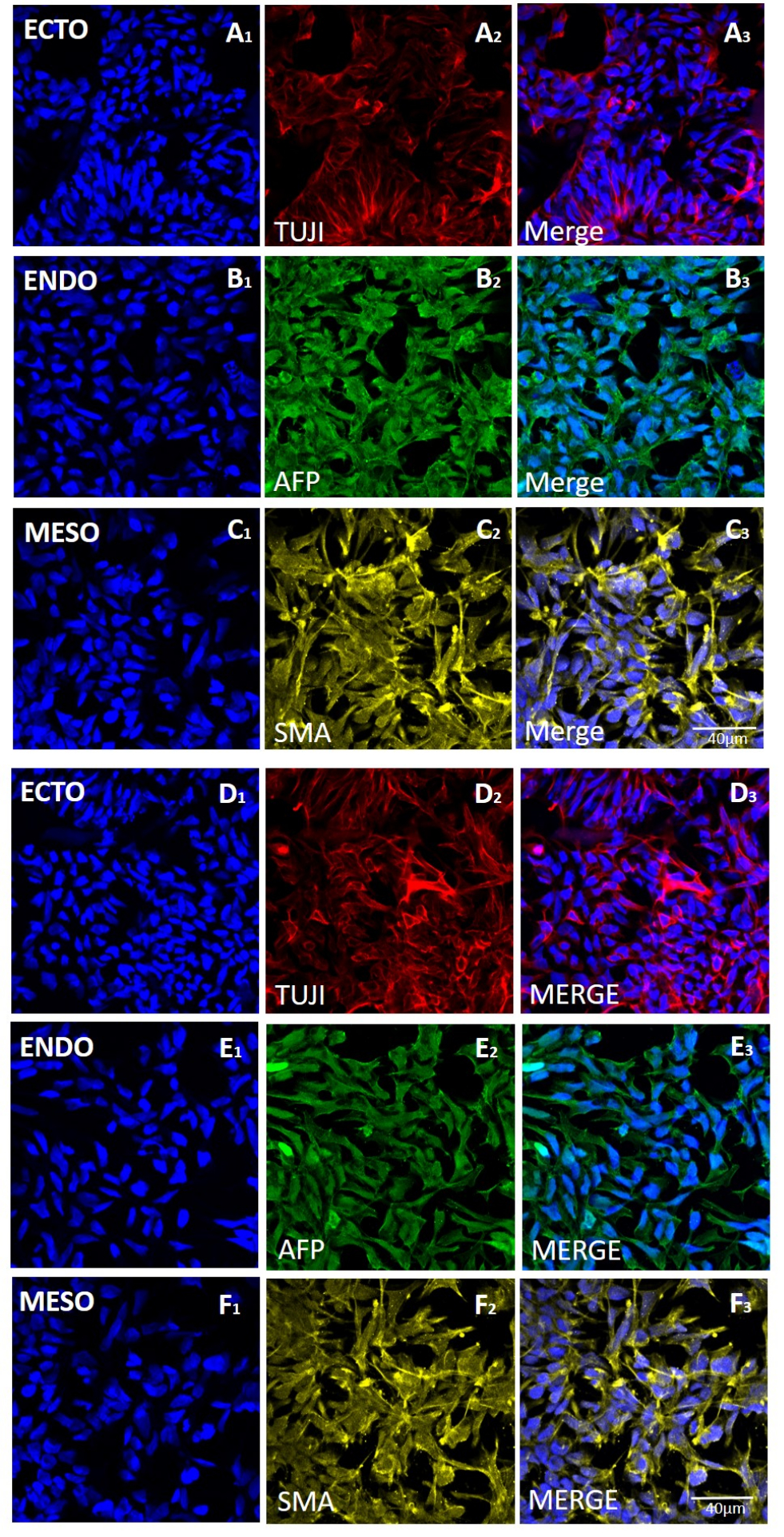{kind=link}

Figure 6 : Expression des marqueurs des trois couches germinales dans les corps embryoïdes générés à partir de la lignée HS429 hESC après prélèvement mécanique ou à base d’EDTA. Marquage par immunofluorescence représentatif des marqueurs de l’ectoderme (rangées A et D) (ECTO, TUJI), de l’endoderme (rangées B et E) (ENDO, AFP) et du mésoderme (rangées C et F) (MESO, SMA). EB générés (A-C) après 20 passages de récolte mécanique (A-C) ou (D-F) après 20 passages de récolte à base d’EDTA. Barres d’échelle = 40 μm. Abréviations : hESC = cellule souche embryonnaire humaine ; EDTA = acide éthylènediaminetétraacétique ; EB = corps embryoïdes. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}

Figure 7 : Analyse génétique basée sur la qPCR des aberrations génomiques courantes dans les lignées HS9 et HS429 ESC et la lignée NCS002 iPSC après 20 passages par récolte mécanique ou à base d’EDTA. La ligne de base à la valeur 2 représente une diploïdie normale à tous les marqueurs chromosomiques. Une valeur de 1 ou 3 représenterait une perte ou un gain, respectivement, du marqueur chromosomique indiqué dans toutes les cellules. Des valeurs intermédiaires comprises entre 1 et 2 ou entre 2 et 3 indiquent la présence d’une perte ou d’un gain du marqueur indiqué dans une fraction des cellules. Il est à noter que le schéma des aberrations est similaire dans les deux conditions de récolte. Abréviations : ESC = cellule souche embryonnaire ; EDTA = acide éthylènediaminetétraacétique ; iPSC = cellule souche pluripotente induite. Veuillez cliquer ici pour voir une version agrandie de cette figure.
{kind=link}
| Densité cellulaire (cellules/mm2) | ||
| H9 | méchant | stdev |
| Vendanges mécaniques avant la poursuite du passage | 3918 | 263.3 |
| Vendanges mécaniques 20 fois | 3868 | 197.7 |
| Récolte EDTA 20 fois | 4080 | 127.8 |
| Réf. HS429 | méchant | stdev |
| Vendanges mécaniques avant la poursuite du passage | 5249 | 565.4 |
| Vendanges mécaniques 20 fois | 5247 | 726.3 |
| Récolte EDTA 20 fois | 4963 | 448.8 |
Tableau 1 : Comparaison des densités cellulaires dans les colonies à partir des deux lignées hESC (H9 et HS429) générées après récolte à base d’EDTA ou mécanique. Les densités cellulaires ont été évaluées soit après un seul passage à l’aide d’une récolte mécanique, soit après 20 passages à l’aide d’une récolte mécanique, soit après 20 passages à l’aide d’une récolte à base d’EDTA (à une dilution de 1 :5). Dans tous les cas, n = 5 colonies.
Discussion
Nous avons décrit une méthode rapide et rentable de récolte des hESC et des hiPSCs cultivées sur des cellules nourricières en utilisant la désadhérence médiée par l’EDTA et nous l’avons comparée principalement à la méthode conventionnelle de récolte mécanique à l’aide d’un scalpel. Nous avons également comparé la récolte basée sur l’EDTA à la récolte enzymatique en ce qui concerne la vitesse de la méthode, mais pas certains aspects de la qualité de la colonie résultante. La raison en est que le prélèvement enzymatique est intrinsèquement plus variable et a été lié à une prévalence plus élevée d’aberrations génomiques5, ce qui pourrait masquer les différences entre les méthodes.
Nous démontrons que la récolte à base d’EDTA est plus rapide et plus efficace que l’une ou l’autre des autres méthodes et génère des colonies plus petites et morphologiquement plus homogènes que la récolte mécanique. Cette dernière caractéristique est bénéfique pour la survie des cellules, car les plus gros amas obtenus par récolte mécanique sont sujets à la nécrose centrale, tandis que la digestion enzymatique a tendance à générer des hESC et des hiPSC isolés, qui sont plus sujets à l’apoptose et nécessitent un traitement supplémentaire, par exemple avec des inhibiteurs de ROCK, pour survivre. La récolte à base d’EDTA peut être utilisée pour au moins 20 passages. Les méthodes de récolte basées sur l’EDTA et mécaniques sont comparables en ce qui concerne la densité cellulaire des colonies, l’expression de l’ARNm et des protéines des gènes souches, la différenciation des trois couches germinales dans les corps embryoïdes et les anomalies génomiques. Si l’objectif est l’efficacité, un rendement plus élevé, moins de variabilité et une manipulation plus douce des hESC et des hiPSC, la récolte à base d’EDTA est préférable.
Nous remarquons également que la récolte à l’aide d’EDTA de CSEh et de CSPhi cultivées sur des cellules nourricières est un moyen peu coûteux de maintenir un état plus naïf et permet une transition en douceur de la culture à base d’alimentation à la culture sans nourrisseur lorsque cela est souhaitable.
Étapes critiques du protocole
Les étapes les plus critiques de la désadhérence médiée par l’EDTA sont la section 3 du protocole (incubation dans la solution d’EDTA) et la section 4 (trituration). Si l’exposition à la solution d’EDTA est supérieure à 1 min, le risque de dissociation complète en cellules individuelles augmente. Cela peut également se produire si la trituration est trop longue ou trop dure. Ce dernier est affecté par la taille de l’embout de la pipette. L’idéal est d’utiliser des pipettes de culture cellulaire de 1 mL comme décrit ici. L’utilisation d’un autre type de pipette avec un diamètre d’embout plus petit est risquée.
Dépannage
Si les cellules nourricières continuent de proliférer, l’arrêt mitotique n’a pas été efficace et un nouveau lot doit être prélevé et la procédure recommencée. Si les colonies ne se détachent pas de la couche de cellules nourricières, il faut s’assurer qu’il n’y a pas de Ca2+ dans l’EDTA et que la boîte de culture contenant les colonies est bien rincée avec du PBS pour éliminer tout milieu de culture cellulaire restant avant d’ajouter l’EDTA. Une trop grande dissociation, qui génère des cellules isolées ou des amas cellulaires trop petits, peut survenir en raison d’une trituration excessive et compromet l’établissement de nouvelles colonies. Le degré de trituration doit être déterminé empiriquement lors d’essais du protocole pour confirmer que les amas cellulaires résultants ont un diamètre de ~60 μm. Si la couche nourricière se détache spontanément de la boîte de culture, en particulier avant que les cellules hESC/hiPSC ne soient prêtes à être récoltées, cela peut être dû au fait que les cellules nourricières n’ont pas été utilisées dans les ~7 jours suivant leur préparation. Par conséquent, le délai d’utilisation des cellules nourricières doit être surveillé attentivement. Si la couche nourricière se dissocie pendant l’exposition à l’EDTA (ce que nous n’avons jamais observé avec les cellules nourricières utilisées ici), le type de cellule nourricière ou leur méthode de culture doivent être modifiés.
Limites de la technique
La principale limite de la technique est qu’elle nécessite une inspection visuelle du processus de désadhérence pour obtenir un résultat positif. Cela signifie que les utilisateurs doivent apprendre à identifier le moment où les colonies se libèrent de la couche de cellules nourricières et où la couche de cellules nourricières se détache du substrat. Cependant, ce n’est pas difficile, et d’après notre expérience, les nouveaux utilisateurs de la technique peuvent la maîtriser en quelques essais.
Il existe également une possibilité inhérente que les CSEh ou les CSPh récoltées soient contaminées par quelques cellules nourricières. Si l’on a l’intention de les transférer dans des conditions autres que celles d’alimentation ou d’isoler les CSEh ou les CSPhi à des fins d’analyse, une telle contamination compromettrait la pureté. Nous remarquons qu’avec les cellules nourricières utilisées ici (fibroblastes du prépuce humain), il est extrêmement difficile de dissocier la couche de cellules nourricières, même avec une digestion enzymatique (non représentée). Étant donné que la couche de cellules nourricières non dissociée est éliminée dans son intégralité, la contamination des CSEh ou des CSPh récoltées est probablement négligeable. De plus, comme les cellules nourricières sont arrêtées mitotiquement, toute contamination finirait par diminuer jusqu’à zéro avec le passage ultérieur des hESCs ou des hiPSCs.
Importance par rapport aux méthodes existantes
La norme actuelle pour la culture des hESC et des hiPSC est de le faire dans des conditions sans feeder, pour lesquelles l’utilisation de l’EDTA pour le passage est répandue. La culture sans mangeoire dépend de l’utilisation de milieux et de substrats de culture spécialement formulés qui garantissent l’adhérence. Ces réactifs entraînent une dépense supplémentaire qui peut dépasser certains budgets de laboratoire. De plus, la culture dans des conditions sans mangeoire a été associée à un potentiel de différenciation perturbé en raison de l’absence de facteurs spécifiques dans les milieux de culture sans mangeoire et d’une transition résultante de l’état naïf à l’état amorcé. La croissance sur des cellules nourricières arrêtées mitotiquement permet d’éviter cette transition et peut ramener les coûts globaux à un niveau gérable, facilitant ainsi l’utilisation plus large des cellules souches pluripotentes dans la recherche en laboratoire.
Déclarations de divulgation
Joel C. Glover en est le directeur et Hege Brincker Fjerdingstad est la directrice quotidienne de la plateforme norvégienne pour les cellules souches pluripotentes humaines. Les auteurs n’ont pas d’intérêts financiers concurrents ou d’autres conflits d’intérêts à divulguer.
Remerciements
Nous remercions Lars Moen pour son aide lors des expériences préliminaires et le Centre norvégien de recherche sur les cellules souches pluripotentes humaines du Centre norvégien de recherche sur les cellules souches de l’hôpital universitaire d’Oslo pour l’utilisation des installations. La ligne hESC H9 a été obtenue auprès de WiCell, et la ligne HS429 hESC a été obtenue auprès d’Outi Hovatta de l’Institut Karolinska. Les deux ont été utilisés conformément aux ententes de transfert de matériel. Les lignées de cellules hiPS NCS001 et NCS002 ont été générées par la plateforme norvégienne de recherche de cellules souches pluripotentes humaines. Cette reprogrammation et tous les travaux rapportés ici ont été effectués avec l’approbation du Comité régional d’éthique du sud-est de la Norvège (approbation REK 2017/110).
matériels
| Name | Company | Catalog Number | Comments |
| 0.5 M EDTA pH 8.0 | Invitrogen | 15575020 | |
| 15 mL centrifuge tubes | Sarstedt | 62.554.502 | |
| 2-mercaptoethanol | Gibco | 31350-010 | |
| 50 mL centrifuge tubes | Sarstedt | 62.547.254 | |
| Basic fibroblast growth factor (bFGF) | PeproTech | AF-100-18B-250UG | |
| Brand Bürker Chamber | Fisher Scientific | 10628431 | |
| Disposable scalpels no.15 | Susann-Morton | 505 | |
| DPBS (1x) without Ca/Mg | Gibco | 14190-094 | |
| Easy Grip tissue culture dish, 35 x 10 mm | Falcon | 353001 | |
| Eppendorf pipette 1 mL | Eppendorf | ||
| Eppendorf pipette 200 μL | Eppendorf | ||
| FBS (Fetal Bovine Serum) | Gibco | 10270-106 | |
| Filter tip 1,000 μL | Sarstedt | 70.1186.210 | |
| Filter tip 200 μL | Sarstedt | 70.760.211 | |
| Gamma Cell 3000 ELAN irradiation machine (alternatively, use Mitomycin C to arrest proliferation) | Best Theratronics | BT/MTS 8007 GC3000E | |
| Glutamax 100x | Gibco | 35050-038 | |
| Growth Factor Reduced Matrixgel | Corning | 734-0269 | |
| H9 hESC line | WiCell | WAe009-A | |
| hPSC Genetic Analysis Kit | Stem Cell Technologies | #07550 | |
| HS429 hESC line | ECACC | KIe024-A | |
| Human Foreskin Fibroblasts -CRL2429 line | ATTC | CRL2429 | |
| IMDM (1x) | Gibco | 21980-032 | |
| iPSC lines | Norwegian Core Facility for Human Pluripotent Stem Cells | NCS001 & NCS002 | |
| Knockout DMEM | Gibco | 10829-018 | |
| Laser Scanning Confocal Microscope or equivalent (we use the LSM 700 from Zeiss) | Zeiss | ||
| Microscope | CETI | ||
| Mitomycin C | Sigma Aldrich | M4287 | |
| Non-essential amino acids (NEAA) | Gibco | 11140.035 | |
| Pipettes, plastic 10 mL | Sarstedt | 86.1254.001 | |
| Pipettes, plastic, 5 mL | Sarstedt | 86.1253.001 | |
| Serum Replacement (SR) | Gibco | 10828-028 | |
| Sterile filters 0.22 um | Sarstedt | 83.1826.102 | |
| T-75 culture flask | ThermoScientific | 156499 | |
| Trypan Blue Stain (0.4 %) | Gibco | 15250-061 | |
| Trypsin-EDTA, 500 mL | Gibco | 25300062 |
Références
- Skottman, H., Hovet, O. Culture conditions for human embryonic stem cells. Reproduction. 132 (5), 691-698 (2006).
- Hovatta, O., et al. A culture system using human foreskin fibroblasts as feeder cells allows production of human embryonic stem cells. Human Reproduction. 18 (7), 1404-1409 (2003).
- Desai, N., Rambhia, P., Gishto, A. Human embryonic stem cell cultivation: historical perspective and evolution of xeno-free culture systems. Reproductive Biology and Endocrinology. 13, 9 (2015).
- Villa-Diaz, L. G., et al. Synthetic polymer coatings for long-term growth of human embryonic stem cells. Nature Biotechnology. 28 (6), 581-583 (2010).
- Watanabe, M., et al. TGFb superfamily signaling regulates the state of human stem cell pluripotency and capacity to create well-structured telencephalic organoids. Stem Cell Reports. 17 (10), 2220-2238 (2022).
- Garitaonandia, I., et al. Increased risk of genetic and epigenetic instability in human embryonic stem cells associated with specific culture conditions. PLoS One. 10 (2), e0118307 (2015).
- Inzunza, J., et al. Derivation of human embryonic stem cell lines in serum replacement medium using postnatal human fibroblasts as feeder cells. Stem Cells. 23, 544-549 (2005).
- Watanabe, K., et al. A ROCK inhibitor permits survival of dissociated human embryonic stem cells. Nature Biotechnology. 25 (6), 681-686 (2007).
- Rivera, T., Zhao, Y., Ni, Y., Wang, J. Human-induced pluripotent stem cell culture methods under cGMP conditions. Current Protocols in Stem Cell Biology. 54 (1), 117 (2020).
- Castro-Viñuelas, R., et al. Tips and tricks for successfully culturing and adapting human induced pluripotent stem cells. Molecular Therapy. Methods & Clinical Development. 23, 569-581 (2021).
- Meng, G., Rancourt, D. E. Derivation and maintenance of undifferentiated human embryonic stem cells. Methods in Molecular Biology. 873, 69-90 (2012).
- Schindelin, J., et al. Fiji: An open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationExplorer plus d’articles
This article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.