
Method Article
Быстрый, экономичный, бесферментный пассаж плюрипотентных стволовых клеток человека на фидерных клетках путем опосредования этилендиаминтетрауксусной кислотой
В этой статье
Резюме
Чтобы избежать ограничений, связанных с ферментативным или механическим пассажем эмбриональных стволовых клеток человека (чЭСК) и индуцированных плюрипотентных стволовых клеток человека (ИПСК), культивируемых на фидерных клетках, мы разработали быстрый, эффективный, экономичный и высокопродуктивный метод сбора колоний чЭСК или ИПСК, поддерживаемых на слое фидерных клеток фибробластов крайней плоти человека, с использованием ЭДТА-опосредованной спайки.
Аннотация
Плюрипотентные стволовые клетки человека (эмбриональные стволовые клетки человека, чЭСК и индуцированные плюрипотентные стволовые клетки человека, ИПСК) первоначально культивировались на различных типах фидерных клеток для поддержания в недифференцированном состоянии при длительном культивировании. Этот подход был в значительной степени вытеснен протоколами культивирования без фидеров, но они включают в себя более дорогостоящие реагенты и могут способствовать переходу в праймированное состояние, которое ограничивает способность клеток к дифференцировке. Как в фидерных, так и в безпитательных условиях сбор геполий hESC или hiPSC для пассажа является необходимой процедурой для выращивания культур.
Чтобы обеспечить простую и высокопроизводительную процедуру пропускания чЭСК/ИПСК, культивируемых на фидерных клетках, мы разработали метод сбора с использованием распайки, вызванной этилендиаминтетрауксусной кислотой (ЭДТА) на основе хелатора кальция. Мы оценили выход и качество полученных пассированных клеток, сравнив этот подход с оригинальным подходом механического забора, при котором колонии выделяют скальпелем под микроскопом (механический забор был выбран в качестве компаратора, чтобы избежать изменчивости реагентов, связанной с ферментативным забором).
В одной серии экспериментов две различные линии чЭСК поддерживались на слое фидерных клеток фибробластов крайней плоти человека. Каждая линия была подвергнута многократным пассажам с использованием ЭДТА или механического сбора и оценена по размеру и морфологии колонии, плотности клеток, экспрессии маркеров стволовости, дифференцировке к трем зародышевым слоям в эмбриоидных телах и геномным аберрациям. В другой серии экспериментов мы использовали сбор на основе ЭДТА на двух разных линиях ИПСК и получили аналогичные результаты. Индуцированная ЭДТА расклеивающаяся адгезия экономила время и давала более высокий выход колоний более благоприятного размера и более однородной морфологии по сравнению с механическим сбором. Он также был быстрее, чем ферментативный сбор, и не был подвержен изменчивости ферментных партий. Метод адгезии, индуцированной ЭДТА, также облегчает перенос линий hESC/hiPSC из культуры на основе фидерных клеток в условия без фидера, если это необходимо, для последующего использования и анализа.
Введение
Надлежащее поддержание чЭСК и ИПСК in vitro является базовой и удобной методологией для нескольких направлений исследований в области клеточной биологии человека и биологии развития. Из-за присущей чЭСК и ИПСК способности к дифференцировке, поддержание недифференцированного состояния in vitro требует особой осторожности и внимания. Таким образом, разработка экономически эффективных протоколов для поддержания и передачи чЭСК и ИПСК с минимально возможной методологической вариативностью имеет большую общую полезность.
Первоначально чЭСК и ИПСК культивировали на различных типах фидерных клеток для содействия длительному культивированию и поддержанию недифференцированного состояния 1,2,3. В последнее время культивирование в условиях отсутствия кормушки стало нормой, так как оно вообще позволяет избежать работы с питательными клетками4. Тем не менее, некоторые лаборатории и основные учреждения до сих пор культивируют чЭСК или ИПСК на фидерных клетках. Безпитательная культура является более дорогой, поскольку требует использования питательных сред специального состава и некоторой формы покрытия поверхности культуры для обеспечения адгезии колонии (основные компоненты внеклеточного матрикса [ВКМ] или коммерческое соединение ВКМ, или использование коммерчески доступных пластин с покрытием). Эти расходы нетривиальны и представляют собой потенциальное финансовое препятствие для некоторых лабораторий, заинтересованных в проведении исследований и разработок на основе чЭСК или ИПСК. Более того, культивирование в условиях без питания имеет тенденцию приводить чЭСК и ИПСК в менее наивное состояние, чем то, которое поддерживается в фидерных клетках5, и это может поставить под угрозу последующую дифференцировку и привести к генетическимвариациям6.
Исторически сложилось так, что передача чЭСК и ИПСК, культивируемых на фидерных клетках, включала механический сбор - использование скальпеля для вырезания колоний под микроскопом7 - но позже это было в значительной степени вытеснено ферментативным расщеплением с мягким соскабливанием или без него для выделения колоний или диссоциированных клеток. Механический сбор урожая утомителен и требует точной микрохирургии. Ферментативный сбор может варьироваться по эффективности из-за различий ферментов от партии к партии и, как правило, способствует полной диссоциации, что способствует гибели клеток, если не противодействовать ингибиторам РОК 8,9, и увеличивает частоту аномальных кариотипов9.
Чтобы воспользоваться преимуществами более низких затрат и большего потенциала дифференцировки культивирования чЭСК и ИПСК на фидерных клетках, избегая при этом недостатков механического и ферментативного сбора, мы разработали быстрый, эффективный, экономичный и высокопродуктивный метод сбора колоний чЭСК и ИПСК на фидерном слое фибробластов крайней плоти человека с использованием ЭДТА-опосредованной спайки. Мы сравнили урожайность, изменчивость и качество стволовых клеток с теми, которые получены при механическом сборе (мы не сравнивали с ферментативным сбраживанием из-за дополнительной изменчивости, которую влечет за собой этот подход). Мы отмечаем, что ЭДТА-опосредованная распайка также хорошо работает для переноса колоний из питательных культур в безпитательные условия, если это необходимо для последующего использования и анализа. Этот метод обеспечивает переход с последовательным методом пассажа, поскольку индуцированная ЭДТА рассеивание является популярным подходом, используемым для культур без корма.
протокол
Подробные сведения обо всех материалах, реагентах и инструментах, используемых в этом протоколе, см. в таблице материалов .
1. Культивирование клеток фибробластов человека и подготовка слоя фидерных клеток
- Засейте 0,5 × 106 фибробластов крайней плоти человека (далее именуемые «фидерными клетками») в каждую колбу для культивирования Т-75 (количество колб по мере необходимости) 20 мл модифицированной среды Дульбекко (IMDM) Iscove с 10% эмбриональной бычьей сывороткой (FBS), далее называемой «питательной клеточной средой».
- Когда питательные ячейки достигнут 90% слияния, удалите среду и промойте 3 раза 10 мл фосфатно-солевого буфера (DPBS) Dulbecco на колбу, чтобы избежать ингибирования трипсина факторами в среде. Добавьте 2 мл трипсина-ЭДТА в каждую колбу и поместите колбу (колбы) в инкубатор с температурой 37 °C/5% CO2 на 5 минут или до тех пор, пока питательные клетки не отделятся от колбы (колб). Наблюдайте отрыв клеток под микроскопом как плавающие агрегаты клеток или одиночные клетки.
- Добавьте в каждую колбу 5 мл свежей предварительно подогретой питательной среды, чтобы инактивировать трипсин-ЭДТА, и осторожно подвешивайте питательные ячейки путем пипетирования.
- Переложите ячейки питателя в центрифужную пробирку объемом 15 мл. Закройте пробирку крышкой и гранулируйте ячейки питателя центрифугированием при 200 × г в течение 5 мин.
- Осторожно удалите надосадочную жидкость, не потревожив гранулу питательной ячейки. Затем осторожно ресуспендируйте гранулу в 4 мл свежей питательной клеточной среды. Перед подсчетом убедитесь, что питательные ячейки полностью ресуспендированы, используя камеру для подсчета клеток или другое устройство для подсчета клеток.
- К необходимому количеству новых колб для культивирования Т-75 добавляют 0,5 × 106 ячеек для расширения и добавляют в каждую колбу 20 мл свежей питательной клеточной среды. Инкубируйте колбу (колбы) для культуры в инкубаторе с температурой 37 °C/5%CO2 до тех пор, пока питательные клетки не достигнут 90% слияния.
ПРИМЕЧАНИЕ: Ячейки питателя можно использовать по крайней мере до прохода 25. - Рассчитайте количество питательных клеток, необходимых для количества 35 мм тканевых культуральных чашек, которые будут использоваться для культивирования чЭСК/ИПСК.
ПРИМЕЧАНИЕ: Обычно 3,0 × 105 фидерных клеток на чашку для культуры тканей достаточно для создания сливающегося слоя фидерных клеток. - Чтобы избежать разрастания питающих клеток, убедитесь, что они митотически остановлены одним из двух способов.
ПРИМЕЧАНИЕ: Для обоих методов может быть сгенерирована большая партия митотически остановленных фидерных клеток, которые могут быть заморожены в аликвотах для последующего использования.- Митотическую остановку проводят гамма-облучением, перенося все необходимые питательные ячейки в центрифужную пробирку объемом 50 мл и доливая питательную клеточную среду до общего объема 5 мл. Немедленно транспортируйте при комнатной температуре в гамма-облучательную установку и облучают для митотической остановки питающих ячеек (300 кВ и 10 мА в течение 20 мин).
ПРИМЕЧАНИЕ: Задержка в транспортировке может привести к нежелательному прикреплению питающих ячеек к стенке центрифужной пробирки объемом 50 мл. Если транспортировка занимает более нескольких минут, убедитесь, что ячейки питателя остаются во взвешенном состоянии во время транспортировки, непрерывно перемешивая трубку. - Митотическую остановку проводят с помощью митомицина С, перенося все питательные клетки, необходимые в 5 мл питательной клеточной среды, в центрифужную пробирку объемом 50 мл, а затем добавляют 15 мл питательной клеточной среды, содержащей 20 мкг/мл митомицина С, и инкубируют в инкубаторе с температурой 37 °C/5% CO2 в течение 3 ч. Добавьте 20 мл PBS с температурой 37 °C, гранулируйте клетки центрифугированием при 200 × г в течение 5 мин, повторите промывку PBS еще два раза и повторно суспендируйте в питательной клеточной среде.
- Митотическую остановку проводят гамма-облучением, перенося все необходимые питательные ячейки в центрифужную пробирку объемом 50 мл и доливая питательную клеточную среду до общего объема 5 мл. Немедленно транспортируйте при комнатной температуре в гамма-облучательную установку и облучают для митотической остановки питающих ячеек (300 кВ и 10 мА в течение 20 мин).
- После того, как питательные клетки были митотически остановлены, вернитесь в колпак для культуры тканей и распределите питательные клетки из расчета 3,0 ×10,5 клеток на 35 мм чашки для культуры тканей, как показано ниже. Убедитесь, что питательные клетки полностью ресуспендированы, добавьте питательную клеточную среду до достижения концентрации питательных клеток 1,5 × 105 на мл и добавьте 2 мл этой суспензии фидерных клеток в каждую 35-миллиметровую чашку для культуры тканей.
- Перенесите чашки с культурами в инкубатор с температурой 37 °C/5% CO2 . Чтобы обеспечить равномерное распределение ячеек кормушки, медленно, но твердо перемещайте чашки для культур на полке инкубатора вперед и назад 3 раза, после чего следует пауза, а затем выполняйте то же действие слева направо 3 раза. Не переставляйте посуду повторно, и аккуратно закройте дверцу инкубатора.
- Через 24 ч перейти с питательной клеточной среды на IMDM с 10% заменой сыворотки (SR). В дальнейшем заменяйте этот носитель каждые три дня. Ячейки кормушки готовы к использованию уже через первые 3 дня.
2. Механический забор гексонов hESC или hiPSC колоний
- Предварительно разогретая среда hESC, состоящая из 80% модифицированной орлиной среды (DMEM) Dulbecco, 20% SR, 1 мМ заменителя глутамина 100x, 1 мМ заменимых аминокислот (NEAA), 1 мМ пенициллина/стрептомицина (P/S), 0,1 мМ 2-меркаптоэтанола и 10 нг/мл основного фактора роста фибробластов (bFGF). Среда hESC используется для культивирования либо hESCs, либо hiPSCs на фидерных клетках.
- По крайней мере, за 30 мин до переноса колоний hESC/hiPSC возьмите свежие 35 мм чашки для культуры тканей, содержащие митотически остановленные фидерные клетки, и замените питательную клеточную среду 1,2 мл среды hESC, содержащей bFGF.
- Поместите культуральную чашку, содержащую колонии hESC/hiPSC, на митотически остановленных фидерных клетках под микроскоп с 10-кратным увеличением, помещенный в колпак ламинарного потока. С помощью стерильного скальпеля аккуратно разрежьте каждую колонию по окружности, а затем разрежьте каждую колонию на 5-6 примерно равных частей. Осторожно приподнимите кусочки колонии кончиком лезвия скальпеля, чтобы они отделились от слоя питательной клетки и свободно плавали в среде.
- Старайтесь избегать областей колоний, содержащих дифференцирующиеся клетки, которые выглядят как островки более мелких клеток с менее отчетливыми ядрами по сравнению с чЭСК/ИПСК внутри колонии.
- Свободно плавающие колонии переносят с помощью пипетки объемом 1 мл в новые чашки для культивирования, содержащие питательные клетки. Старайтесь держать колонии отдельно, чтобы потом они не переросли друг в друга. Осторожно переместите чашки с культурами в клеточный инкубатор и не тревожьте посуду до следующего дня.
- На следующий день осторожно добавьте 600 мкл среды hESC, содержащей bFGF, до конечного объема 1,8 мл. После этого заменяйте среду hESC + bFGF каждый день до следующего прохождения (обычно через 1 неделю).
3. Забор колоний чЭСК или ИПСК с помощью ЭДТА-опосредованной дисадгезии
- Берут свежие чашки для культивирования с митотически остановленными фидерными клетками и переходят с ИМДМ с 10% SR на 1,2 мл предварительно подогретой среды hESC + bFGF не менее чем за 30 мин до переноса колоний.
- Обрабатывайте по одной чашке с культурой, содержащей ЭСК или ИПСК колонии. Удалите среду hESC + bFGF и промойте колонии 1 мл DPBS комнатной температуры, чтобы удалить любые возможные неприкрепленные клетки и клеточный мусор. Добавьте 1 мл 0,5 мМ ЭДТА и инкубируйте в течение 1 мин при 37 °C. Если ламинарный вытяжной шкаф имеет нагревательную пластину, выполните этот шаг и шаги, описанные в разделе 4, на нагревательной пластине для лучшего сцепления.
- После 1 мин инкубации удалите раствор ЭДТА и осторожно добавьте 1 мл среды hESC + bFGF с помощью пипетки объемом 1 мл. Осторожно растирайте той же пипеткой, чтобы освободить колонии из слоя питательной клетки. Продолжайте осторожно растирань, пока слой питательной клетки не разрыхнется и не сложится в отдельный комок. Снимите слой ячейки питателя с помощью наконечника пипетки.
- Перенесите взвешенные колонии hESC/hiPSC с помощью новой пипетки объемом 1 мл в новые культуральные чашки, содержащие фидерные клетки и среду hESC+bFGF, расщепляя в соотношении 1:5. Колонии, как правило, равномерно распределяются в каждой новой чашке с культурой, но облегчают это, осторожно перемещая чашку из стороны в сторону. После этого заменяйте среду hESC + bFGF каждый день до следующего прохождения (обычно через 1 неделю).
Результаты
В анализах и сравнениях, описанных ниже, мы использовали две линии hESC (H9 и HS429, от WiCell и Каролинского института, соответственно) и две линии hiPSC (NCS001 и NCS002, обе получены в Норвежском центре плюрипотентных стволовых клеток человека). Данные, представленные на рисунках и в таблицах, взяты из линий hESC, но совершенно аналогичные результаты были получены из линий hiPSC.
В наших руках механический сбор привел к тому, что колонии были разделены примерно на пять-шесть комков диаметром ~200-250 мкм, в то время как при индуцированной ЭДТА расклеивании с последующей тренировкой каждая колония была разделена на ~10-20 комков диаметром ~60 мкм. По нашим оценкам, количество клеток в каждом сгустке, собранном с помощью ЭДТА, составляет ~20. Поскольку нецелесообразно разбивать колонию на комки такого размера с помощью скальпеля, в этом отношении индуцированная ЭДТА распайка является лучшей, поскольку она генерирует скопления размера, более благоприятного для выживания клеток колонии10,11.
Колонии hESC/hiPSC, собранные с помощью ЭДТА, также были более однородными по размеру и форме по сравнению с колониями, собранными механическим способом (рис. 1A-F). Это связано с тем, что резка, необходимая для механической уборки, приводит к появлению неровных кромок и комков различного размера. Чтобы количественно оценить это, мы оценили круговую структуру колонии (как меру того, насколько округлыми были края колонии; значение 1 указывает на идеальную окружность) через 5 дней после пассажа с помощью протокола ImageJ-win6412. Круговорота колоний были значительно ниже в колониях, собранных механическим способом (механическая уборка: 0,61 ± 0,10; Сбор урожая на основе ЭДТА: 0,84 ± 0,01; n = 10, p < 0,001, U-критерий Манна-Уитни, U = 10).
Плотность клеток в собранных и повторно покрытых колониях, которая является мерой межклеточных взаимодействий после сбора урожая во время формирования колоний, была аналогична при сборе на основе ЭДТА и механическом сборе (Таблица 1 и Рисунок 1G, H). Механически собранные колонии имели большую склонность к развитию некроза в центральных областях (рис. 1J). Это, вероятно, было связано с изменчивостью формы и, в частности, размером механически изолированных клеточных скоплений, поскольку, когда эти скопления слишком велики, они могут легко сворачиваться сами по себе при переносе в новые чашки для культивирования. Это не относится к колониям, собранным с использованием ЭДТА, которые равномерно демонстрировали полупрозрачный вид с четкими краями (рис. 1I).
Используя сбор урожая на основе ЭДТА, мы смогли собрать практически все колонии, которые были созданы в колодце, в течение 2-3 минут. При использовании механической уборки сбор всех колоний в колодец был бы утомительным и трудоемким. Обычно нам удавалось собрать только ~30%, или ~20-25 колоний, используя механическую заготовку, и это занимало ~20 минут. Точно так же, используя расщепление коллагеназы с последующим мягким соскабливанием, обычно было трудно собрать все колонии, хотя вся процедура заняла всего несколько минут. Таким образом, сбор урожая на основе ЭДТА является таким же быстрым или даже более быстрым, чем ферментативный сбор, и более эффективным, чем механический или ферментативный сбор.
Чтобы оценить влияние различных методов сбора на стебельность и плюрипотентность, мы сначала подвергли колонии, полученные после 20 пассажей, с помощью ЭДТА или механического сбора, анализу кПЦР (рис. 2) и иммуноцитохимическому окрашиванию (рис. 3 и рис. 4) на маркеры стволовости. Колонии, полученные с помощью обоих методов, демонстрировали стабильную экспрессию маркеров стволовости как на уровне мРНК, так и на уровне белка. Затем мы оценили плюрипотентность путем дифференцировки на три зародышевых слоя в эмбриоидных телах (рис. 5 и рис. 6). Эмбриоидные тельца, полученные из чЭСК или ИПСК, полученные после 20 пассажей с использованием любого метода, содержали смесь клеток, экспрессирующих обычно оцениваемые маркеры эктодермы, мезодермы и энтодермы.
Наконец, мы оценили частоту геномных аберраций в чЭСК и ИПСК, пассированных каждым методом, с помощью генетического анализа на основе кПЦР (см. таблицу материалов). Колонии, полученные после 20 пассажей с использованием любого метода забора, продемонстрировали некоторые примеры умеренного отклонения от эталонного диплоидного хромосомного паттерна (оцениваемые аномалии были теми, которые обычно связаны с перепрограммированием ИПСК, но могут быть получены и в чЭСК) (рис. 7). Тем не менее, характер этих отклонений был по существу одинаковым в колониях, полученных после любого метода сбора, что указывает на то, что они не были связаны с методом сбора.

Рисунок 1: Морфология колоний и плотность клеток после забора на основе ЭДТА или механического сбора. (A-F) Репрезентативные светлопольные изображения колоний H9 hESC, установленных в культуре без фидера в течение 5 дней после 20 пассажей с использованием (A-C) на основе ЭДТА или (D-F) механического сбора. (Г,Н) Репрезентативные флуоресцентные изображения плотности клеток в колониях H9 hESC, полученные после 20 пассажей с использованием (G) ЭДТА или (H) механического сбора. Ядра клеток окрашивают DAPI. (Я,Дж) Репрезентативные светлопольные изображения колоний H9 hESC, полученные после 20 проходов с использованием (I) ЭДТА или (J) механического сбора. Обратите внимание на некротическую центральную область в колонии, собранной механическим способом (стрелка в J). Все снимки были получены через 5 дней после 20-го прохода. Масштабные линейки = 100 мкм. Сокращения: hESC = эмбриональная стволовая клетка человека; ЭДТА = этилендиаминтетрауксусная кислота; DAPI = 4',6-диамидино-2-фенилиндол. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}

Рисунок 2: Экспрессия мРНК маркера стволовости в двух линиях hESC (H9 и HS429), полученных после механического сбора на основе ЭДТА. Количественная полимеразная цепная реакция в реальном времени указанных маркеров в ЭСК H9 (верхняя панель) и HS429 (нижняя панель) после однократного прохождения с использованием механического сбора, после 20 пассажей с использованием механического сбора и после 20 пассажей с использованием сбора на основе ЭДТА (разведение 1:5). Уровень экспрессии является относительным к уровню экспрессии гена, отвечающего за ведение домашнего хозяйства ACTB (бета-актина). Столбцы погрешности показывают стандартное отклонение. Сокращения: hESC = эмбриональная стволовая клетка человека; ЭДТА = этилендиаминтетрауксусная кислота. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}

Рисунок 3: Экспрессия белков-маркеров стебельчатости в линии H9 hESC после различных условий уборки. Репрезентативное иммунофлуоресцентное окрашивание колоний H9 hESC, собранных механическим способом (A-E) перед дальнейшим пассажем, (F-J) после 20 пассажей с использованием механического сбора и (K-O) после 20 пассажей с использованием сбора на основе ЭДТА. Масштабные линейки = 100 мкм. Сокращения: hESC = эмбриональная стволовая клетка человека; ЭДТА = этилендиаминтетрауксусная кислота; DAPI = 4',6-диамидино-2-фенилиндол. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}

Рисунок 4: Экспрессия белков-маркеров стебельчатости в линии HS429 hESC после различных условий сбора урожая. Репрезентативное иммунофлуоресцентное окрашивание колоний HS429 hESC, собранных механическим способом (A-E) перед дальнейшим пассажем, (F-J) после 20 пассажей с использованием механического сбора и (K-O) после 20 пассажей с использованием сбора на основе ЭДТА. Масштабные линейки = 100 мкм. Сокращения: hESC = эмбриональная стволовая клетка человека; ЭДТА = этилендиаминтетрауксусная кислота; DAPI = 4',6-диамидино-2-фенилиндол. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
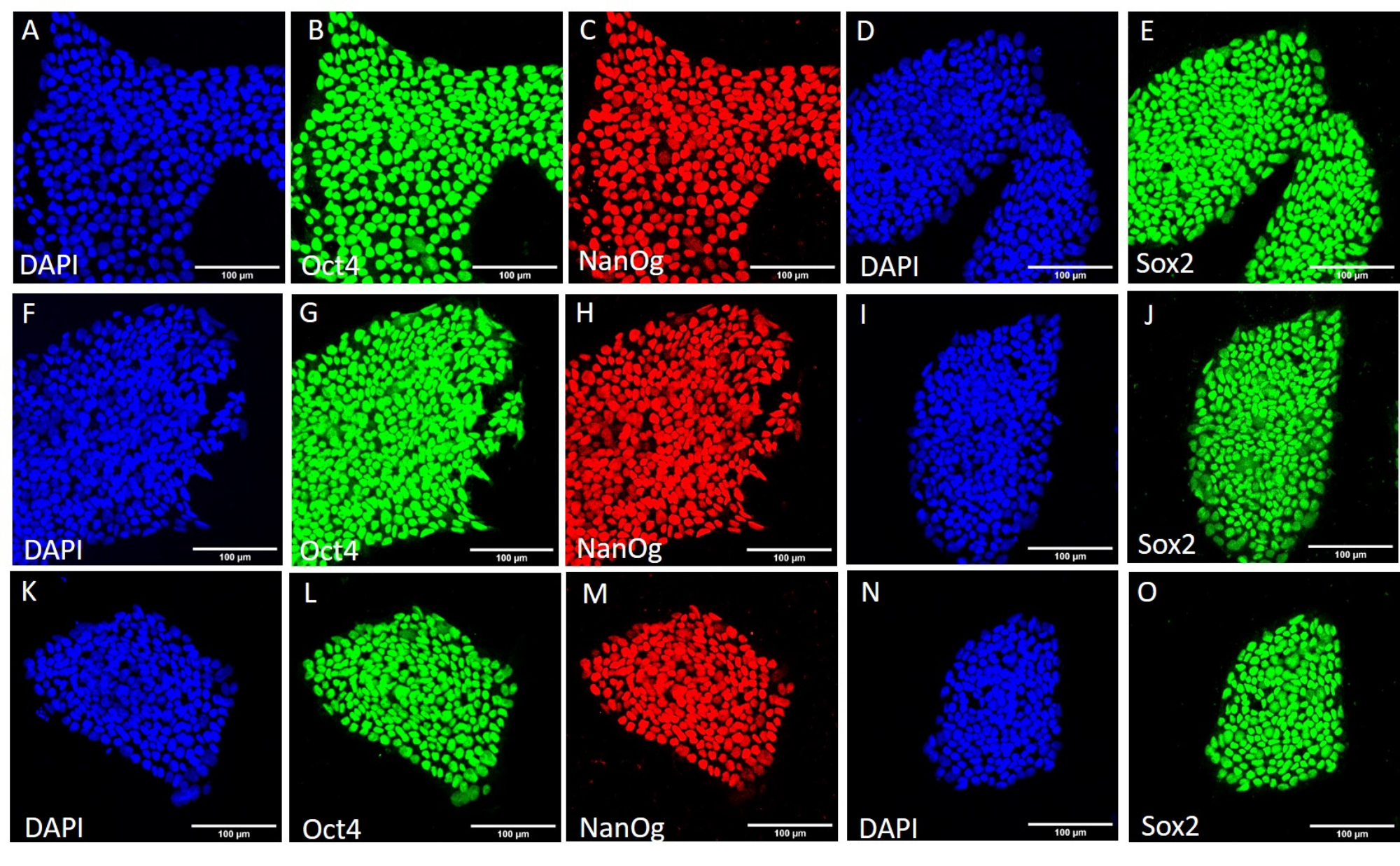{kind=link}

Рисунок 5: Экспрессия маркеров для трех зародышевых слоев в эмбриоидных телах, полученных из линии H9 hESC после механического забора или забора на основе ЭДТА. Репрезентативное иммунофлуоресцентное окрашивание маркеров эктодермы (ряды A и D) (ECTO, TUJI), (ряды B и E) энтодермы (ENDO, AFP) и мезодермы (ряды C и F) (MESO, SMA). ЭБ, образующиеся (A-C) после 20 проходов механической уборки или (D-F) после 20 проходов сбора на основе ЭДТА. Масштабные линейки = 40 мкм. Сокращения: hESC = эмбриональная стволовая клетка человека; ЭДТА = этилендиаминтетрауксусная кислота; ЭБ = эмбриоидные тельца. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
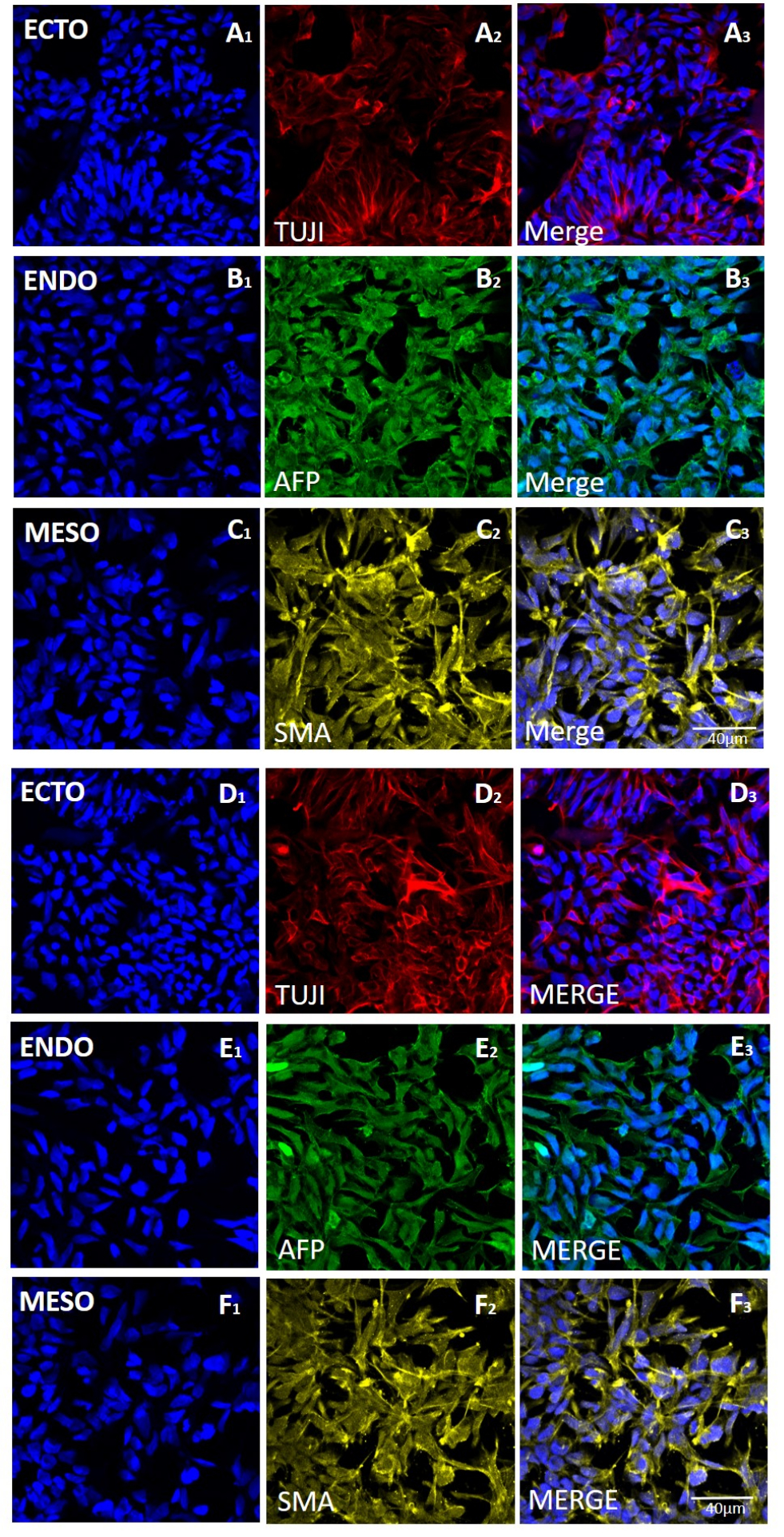{kind=link}

Рисунок 6: Экспрессия маркеров для трех зародышевых слоев в эмбриоидных телах, полученных из линии HS429 hESC после механического забора или забора на основе ЭДТА. Репрезентативное иммунофлуоресцентное окрашивание маркеров эктодермы (ряды A и D) (ECTO, TUJI), (ряды B и E) энтодермы (ENDO, AFP) и мезодермы (ряды C и F) (MESO, SMA). ЭБ, образующиеся (A-C) после 20 проходов механической уборки (A-C) или (D-F) после 20 проходов сборки на основе ЭДТА. Масштабные линейки = 40 мкм. Сокращения: hESC = эмбриональная стволовая клетка человека; ЭДТА = этилендиаминтетрауксусная кислота; ЭБ = эмбриоидные тельца. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}

Рисунок 7: Генетический анализ распространенных геномных аберраций на основе кПЦР в линиях HS9 и HS429 ESC и линии NCS002 iPSC после 20 пассажей с использованием механического забора или забора на основе ЭДТА. Исходный уровень при значении 2 представляет собой нормальную диплоидию по всем хромосомным маркерам. Значение 1 или 3 будет означать потерю или прирост, соответственно, указанного хромосомного маркера во всех клетках. Промежуточные значения между 1 и 2 или между 2 и 3 указывают на наличие потери или усиления указанного маркера во фракции ячеек. Обратите внимание, что характер аберраций одинаков в двух условиях сбора урожая. Сокращения: ESC = эмбриональная стволовая клетка; ЭДТА = этилендиаминтетрауксусная кислота; ИПСК = индуцированная плюрипотентная стволовая клетка. Пожалуйста, нажмите здесь, чтобы увидеть увеличенную версию этого рисунка.
{kind=link}
| Плотность ячеек (ячейки/мм2) | ||
| Н9 | значить | Стандартное уведомление |
| Механизированная уборка урожая перед дальнейшим прохождением | 3918 | 263.3 |
| Механизированный сбор урожая 20 раз | 3868 | 197.7 |
| Урожай ЭДТА 20 раз | 4080 | 127.8 |
| ХС429 | значить | Стандартное уведомление |
| Механизированная уборка урожая перед дальнейшим прохождением | 5249 | 565.4 |
| Механизированный сбор урожая 20 раз | 5247 | 726.3 |
| Урожай ЭДТА 20 раз | 4963 | 448.8 |
Таблица 1: Сравнение плотности клеток в колониях двух линий hESC (H9 и HS429), полученных после механического сбора на основе ЭДТА. Плотность клеток оценивали либо после однократного пассажа с использованием механического сбора, либо после 20 пассажей с использованием механического сбора, либо после 20 пассажей с использованием забора на основе ЭДТА (при разведении 1:5). Во всех случаях n = 5 колоний.
Обсуждение
Мы описали быстрый и экономичный метод сбора чЭСК и ИПСК, культивируемых на фидерных клетках, с использованием ЭДТА-опосредованной распайки, и сравнили его в первую очередь с традиционным методом механического сбора с использованием скальпеля. Мы также сравнили сбор на основе ЭДТА с ферментативным сбором в отношении скорости метода, но не аспектов результирующего качества колоний. Причина этого заключается в том, что ферментативный сбор по своей природе более вариабельен и связан с более высокой распространенностью геномных аберраций5, что может скрыть различия между методами.
Мы демонстрируем, что сбор на основе ЭДТА является более быстрым и эффективным, чем любой из других методов, и создает более мелкие и морфологически более однородные колонии, чем механический сбор. Эта последняя особенность полезна с точки зрения выживания клеток, поскольку более крупные скопления, полученные с помощью механического сбора, склонны к центральному некрозу, в то время как ферментативное расщепление имеет тенденцию генерировать изолированные чЭСК и ИПСК, которые более склонны к апоптозу и требуют дополнительного лечения, например, ингибиторами ROCK, для выживания. Сбор урожая на основе ЭДТА может быть использован не менее чем для 20 проходов. Методы сбора на основе ЭДТА и механического сбора сопоставимы, когда речь идет о плотности клеток колонии, экспрессии мРНК и белков генов стволовости, дифференцировке трех зародышевых слоев в эмбриоидных телах и геномных аномалиях. Если целью является эффективность, более высокая урожайность, меньшая изменчивость и более щадящее обращение с чЭСК и ИПСК, предпочтительнее сбор урожая на основе ЭДТА.
Мы также отмечаем, что сбор ЭСК и ИПСК на основе ЭДТА, культивируемых на фидерных клетках, является недорогим способом поддержания более наивного состояния и обеспечивает плавный переход от культуры на основе фидера к культуре без фидера там, где это желательно.
Критические шаги в рамках протокола
Наиболее важными этапами ЭДТА-опосредованной распайки являются раздел протокола 3 (инкубация в растворе ЭДТА) и раздел 4 (растирание). Если воздействие раствора ЭДТА длится дольше 1 мин, возрастает риск полной диссоциации на единичные клетки. Это также может произойти, если растирание слишком затяжное или слишком резкое. На последнее влияет размер наконечника пипетки. Идеальным вариантом является использование пипеток для клеточных культур объемом 1 мл, как описано здесь. Использование другого типа пипетки с меньшим диаметром наконечника рискованно.
Устранение неполадок
Если питающие клетки продолжают размножаться, митотическая остановка не была эффективной, и необходимо взять новую партию и начать процедуру заново. Если колонии не отделяются от слоя питательных клеток, необходимо убедиться, что в ЭДТА нет Ca2+ и что культуральная чашка, содержащая колонии, хорошо промыта PBS, чтобы удалить оставшуюся клеточную культуральную среду перед добавлением ЭДТА. Из-за чрезмерного растирания может возникнуть слишком сильная диссоциация, которая приводит к образованию изолированных клеток или слишком маленьких скоплений клеток, что ставит под угрозу образование новых колоний. Степень трения должна быть определена эмпирически в пробных запусках протокола, чтобы подтвердить, что результирующие клеточные скопления имеют диаметр ~60 мкм. Если питательный слой самопроизвольно отделяется от чашки для культивирования, особенно до того, как чЭСК/ИПСК будут готовы к сбору, это может быть связано с тем, что питательные клетки не использовались в течение ~7 дней после приготовления. Поэтому следует тщательно следить за временными рамками использования фидерных ячеек. Если питательный слой диссоциирует во время воздействия ЭДТА (чего мы никогда не наблюдали с используемыми здесь фидерными клетками), необходимо изменить либо тип фидерной клетки, либо метод ее культивирования.
Ограничения методики
Основным ограничением методики является то, что она требует визуального осмотра процесса рассеивания для достижения успешного результата. Это означает, что пользователи должны научиться определять, когда колонии высвобождаются из слоя питательных клеток, а слой питательных клеток ослабевает от субстрата. Однако это не сложно, и, по нашему опыту, новые пользователи методики могут освоить ее в течение пары проб.
Существует также неотъемлемая вероятность того, что собранные чЭСК или ИПСК могут быть загрязнены несколькими фидерными клетками. Если намерение состоит в том, чтобы перейти в условия, не связанные с подачей, или изолировать чЭСК или ИПСК для анализов, такое загрязнение может поставить под угрозу чистоту. Отметим, что с используемыми здесь фидерными клетками (фибробластами крайней плоти человека) крайне сложно диссоциировать слой фидерных клеток, даже при ферментативном расщеплении (не показано). Поскольку недиссоциированный слой фидерных клеток удаляется in toto, загрязнение собранных чЭСК или ИПСК, вероятно, будет незначительным. Кроме того, поскольку питающие клетки митотически остановлены, любое загрязнение в конечном итоге уменьшается до нуля при дальнейшем прохождении чЭСК или ИПСК.
Значимость по отношению к существующим методам
В настоящее время нормой культивирования чЭСК и ИПСК является проведение культивирования в условиях без фидера, для чего широко распространено использование ЭДТА для пассажа. Культивирование без фидеров зависит от использования специально разработанных сред и культуральных субстратов, которые обеспечивают прилегание. Эти реагенты влекут за собой дополнительные расходы, которые могут превышать бюджеты некоторых лабораторий. Кроме того, культивирование в условиях без фидера ассоциировалось с нарушенным дифференцирующим потенциалом из-за отсутствия специфических факторов в свободных питательных питательных средах и, как следствие, перехода от наивного состояния к праймированному. Рост на митотически остановленных фидерных клетках позволяет избежать этого перехода и может снизить общие затраты до управляемого уровня, тем самым способствуя более широкому использованию плюрипотентных стволовых клеток в лабораторных исследованиях.
Раскрытие информации
Джоэл С. Гловер является директором, а Хеге Бринкер Фьердингстад – ежедневным менеджером Норвежского центра плюрипотентных стволовых клеток человека. У авторов нет конкурирующих финансовых интересов или других конфликтов интересов, которые они могли бы раскрыть.
Благодарности
Мы благодарим Ларса Моэна за помощь в проведении предварительных экспериментов и Норвежский центр по изучению плюрипотентных стволовых клеток человека при Норвежском центре исследований стволовых клеток при Университетской больнице Осло за использование оборудования. Линия H9 hESC была получена от WiCell, а линия HS429 hESC была получена от Outi Hovatta из Каролинского института. Оба использовались в соответствии с Соглашениями о передаче материалов. Линии hiPSC NCS001 и NCS002 были сгенерированы Норвежским центром плюрипотентных стволовых клеток человека. Это перепрограммирование и вся работа, о которой здесь сообщается, были выполнены с одобрения Регионального комитета по этике Юго-Восточной Норвегии (утверждение REK 2017/110).
Материалы
| Name | Company | Catalog Number | Comments |
| 0.5 M EDTA pH 8.0 | Invitrogen | 15575020 | |
| 15 mL centrifuge tubes | Sarstedt | 62.554.502 | |
| 2-mercaptoethanol | Gibco | 31350-010 | |
| 50 mL centrifuge tubes | Sarstedt | 62.547.254 | |
| Basic fibroblast growth factor (bFGF) | PeproTech | AF-100-18B-250UG | |
| Brand Bürker Chamber | Fisher Scientific | 10628431 | |
| Disposable scalpels no.15 | Susann-Morton | 505 | |
| DPBS (1x) without Ca/Mg | Gibco | 14190-094 | |
| Easy Grip tissue culture dish, 35 x 10 mm | Falcon | 353001 | |
| Eppendorf pipette 1 mL | Eppendorf | ||
| Eppendorf pipette 200 μL | Eppendorf | ||
| FBS (Fetal Bovine Serum) | Gibco | 10270-106 | |
| Filter tip 1,000 μL | Sarstedt | 70.1186.210 | |
| Filter tip 200 μL | Sarstedt | 70.760.211 | |
| Gamma Cell 3000 ELAN irradiation machine (alternatively, use Mitomycin C to arrest proliferation) | Best Theratronics | BT/MTS 8007 GC3000E | |
| Glutamax 100x | Gibco | 35050-038 | |
| Growth Factor Reduced Matrixgel | Corning | 734-0269 | |
| H9 hESC line | WiCell | WAe009-A | |
| hPSC Genetic Analysis Kit | Stem Cell Technologies | #07550 | |
| HS429 hESC line | ECACC | KIe024-A | |
| Human Foreskin Fibroblasts -CRL2429 line | ATTC | CRL2429 | |
| IMDM (1x) | Gibco | 21980-032 | |
| iPSC lines | Norwegian Core Facility for Human Pluripotent Stem Cells | NCS001 & NCS002 | |
| Knockout DMEM | Gibco | 10829-018 | |
| Laser Scanning Confocal Microscope or equivalent (we use the LSM 700 from Zeiss) | Zeiss | ||
| Microscope | CETI | ||
| Mitomycin C | Sigma Aldrich | M4287 | |
| Non-essential amino acids (NEAA) | Gibco | 11140.035 | |
| Pipettes, plastic 10 mL | Sarstedt | 86.1254.001 | |
| Pipettes, plastic, 5 mL | Sarstedt | 86.1253.001 | |
| Serum Replacement (SR) | Gibco | 10828-028 | |
| Sterile filters 0.22 um | Sarstedt | 83.1826.102 | |
| T-75 culture flask | ThermoScientific | 156499 | |
| Trypan Blue Stain (0.4 %) | Gibco | 15250-061 | |
| Trypsin-EDTA, 500 mL | Gibco | 25300062 |
Ссылки
- Skottman, H., Hovet, O. Culture conditions for human embryonic stem cells. Reproduction. 132 (5), 691-698 (2006).
- Hovatta, O., et al. A culture system using human foreskin fibroblasts as feeder cells allows production of human embryonic stem cells. Human Reproduction. 18 (7), 1404-1409 (2003).
- Desai, N., Rambhia, P., Gishto, A. Human embryonic stem cell cultivation: historical perspective and evolution of xeno-free culture systems. Reproductive Biology and Endocrinology. 13, 9 (2015).
- Villa-Diaz, L. G., et al. Synthetic polymer coatings for long-term growth of human embryonic stem cells. Nature Biotechnology. 28 (6), 581-583 (2010).
- Watanabe, M., et al. TGFb superfamily signaling regulates the state of human stem cell pluripotency and capacity to create well-structured telencephalic organoids. Stem Cell Reports. 17 (10), 2220-2238 (2022).
- Garitaonandia, I., et al. Increased risk of genetic and epigenetic instability in human embryonic stem cells associated with specific culture conditions. PLoS One. 10 (2), e0118307 (2015).
- Inzunza, J., et al. Derivation of human embryonic stem cell lines in serum replacement medium using postnatal human fibroblasts as feeder cells. Stem Cells. 23, 544-549 (2005).
- Watanabe, K., et al. A ROCK inhibitor permits survival of dissociated human embryonic stem cells. Nature Biotechnology. 25 (6), 681-686 (2007).
- Rivera, T., Zhao, Y., Ni, Y., Wang, J. Human-induced pluripotent stem cell culture methods under cGMP conditions. Current Protocols in Stem Cell Biology. 54 (1), 117 (2020).
- Castro-Viñuelas, R., et al. Tips and tricks for successfully culturing and adapting human induced pluripotent stem cells. Molecular Therapy. Methods & Clinical Development. 23, 569-581 (2021).
- Meng, G., Rancourt, D. E. Derivation and maintenance of undifferentiated human embryonic stem cells. Methods in Molecular Biology. 873, 69-90 (2012).
- Schindelin, J., et al. Fiji: An open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены