
Method Article
פרוטוקול לזיהוי של אינטראקציות בין חלבונים על הסמך
In This Article
Summary
אנו מציגים וריאציה של (immunoprecipitation כמותיים בשילוב עם מציאה) הגישה המהירה שהוצגה בעבר להבחין בין אמת ושקר אינטראקציות בין חלבונים. הגישה שלנו מבוססת על 15N תיוג מטבולית, האפנון של זיקות של אינטראקציות בין חלבונים על ידי הנוכחות / היעדרות של ה-ATP, immunoprecipitation, וספקטרומטריית מסה כמותית.
Abstract
אינטראקציות בין חלבונים הן בסיסיות לתהליכים ביולוגיים רבים בתא. לכן, האפיון שלהם ממלא תפקיד חשוב במחקר נוכחי ושפע של שיטות לחקירה שלהם זמינה 1. אינטראקציות בין חלבונים לעתים קרובות הן מאוד דינמיות ויכולות להיות תלויות בלוקליזציה subcellular, שינויים שלאחר translational וסביבת החלבון המקומית 2. לכן, הם צריכים להיחקר בסביבה הטבעית שלהם, שלגישות שיתוף immunoprecipitation הן שיטת הבחירה 3. שותפי אינטראקצית שיתוף זרזו מזוהים או על ידי immunoblotting בגישה ממוקדת, או על ידי ספקטרומטריית מסה (LC-MS/MS) באופן לא ממוקד. האסטרטגיה האחרונה לעתים קרובות מושפעת לרעה על ידי מספר גדול של תגליות חיוביות כוזבות, נבע בעיקר מהרגישות הגבוהה של ספקטרומטרים המונים מודרניים שבביטחון לזהות עקבות של חלבונים מזרזים unspecifically. Recenגישה לא להתגבר על בעיה זו מבוססת על הרעיון שסכומים מופחתים של שותפי אינטראקציה ספציפיים יהיו שותף במשקע עם חלבון המטרה נתונה שריכוז סלולרי הוא מופחת על ידי RNAi, ואילו הסכומים של חלבונים מזרזים unspecifically צריכים להיות מושפעים. גישה זו, המכונית מהירה לimmunoprecipitation כמותיים בשילוב עם מציאה 4, מעסיקה תיוג יציב האיזוטופי של חומצות אמינו בתרבית תאים (SILAC) 5 ו-MS לכמת את הכמויות של חלבוני immunoprecipitated מזנים פראיי סוג ועקום למטה. חלבונים הנמצאים בליחס של 1:1 יכולים להיחשב כמזהמים, אלה מועשרים במשקעים מהסוג הפראי כשותפי אינטראקציה ספציפיות של חלבון המטרה. אמנם חדשני, מהיר נושא מספר מגבלות: ראשית, SILAC הוא העלות אינטנסיבית ומוגבל לאורגניזמים שהם אידיאליים לauxotrophic ארגינין ו / או ליזין. תוצאות interconversion יתר על כן, כאשר ארגינין כבד נזון, ארגינין ל-פרולין בadditמסת ional המשמרות עבור כל פרולין בפפטיד ומעט מדללת כבד עם ארגינין אור, מה שהופך את הכימות יותר משעמם ופחות מדויק 5,6. שנית, מהיר דורש כי נוגדני טיטרציה כך שהם אינם הופכים רוויים בחלבון המטרה בתמציות מעקומות למטה מוטציות.
כאן אנו מציגים פרוטוקול מהיר שונה המתגבר על המגבלות האמורות לעיל מהיר על ידי החלפת SILAC תיוג 15 N חילוף חומרים ועל ידי החלפת RNAi בתיווך עקום למטה לאפנון זיקה של אינטראקציות בין חלבונים. אנחנו מדגימים את הישימות של פרוטוקול זה באמצעות Chlamydomonas האצה הירוק unicellular reinhardtii כאורגניזם מודל ומלווה HSP70B הכלורופלסט כחלבון המטרה 7 (איור 1). HSP70s ידוע לאינטראקציה עם עמיתים למלווי מצעים רק במדינת ADP 8 ספציפיים. אנו מנצלים נכס זה כאמצעי לאימות ספציפיתאינטראקציה של HSP70B עם גורם חילופי נוקלאוטיד 9 CGE1.
Protocol
1. נוגדן ספיחה
- תשקול את 120 מ"ג של חלבון Sepharose בצינור חרוטים 15-מ"ל (פלקון). כ15 מ"ג חלבון sepharose נחוץ לכל immunoprecipitation (IP) סכום זה מספיק ל8 כתובות IP. הוסף חוצץ 5 מ"ל 0.1 מ 'פוספט (pH 7.4) ולתת החלבון להתנפח Sepharose למשך 30 דקות ב 4 ° C.
(שים לב כי בכל הצעדים מנקודה זו ואילך צריכות להתבצע עם כפפות, כדי למנוע את הזיהום עם קרטין ועל קרח כדי למנוע פירוק חלבונים / דיסוציאציה מורכבת.)
- צנטריפוגה ל60 שניות ב1000 XG ו4 המעלות צלזיוס לגלולת החלבון הנפוח Sepharose. הסר בזהירות את supernatant ו resuspend את החרוזים במאגר 5 מ"ל 0.1 מ 'פוספט (pH 7.4). חזור על פעולה זו שלוש פעמים לשטוף את החרוזים ביסודיות.
- לאחר השלב האחרון צנטריפוגה, להסיר supernatant ולהשאיר בערך 0.5 מ"ל של חיץ פוספט. הוסף 0.9 מ"ל 0.5 מ 'פוספט חיץ (pH 7.4), 400זיקת μl מטוהרי נוגדנים עיקריים (50 μl לIP) נגד חלבון המטרה (כאן HSP70B), ו16 נוגדנים נגד μl חלבון בקרה (כאן CF1β). מלא עם DDH 2 O להיקף כולל של 5 מ"ל.
(שים לב שנוגדני זיקה-מטוהרים יש להשתמש כדי להפחית את הזיהום על ידי IgGs הנוקב, שמפריע לניתוח ננו LC-MS - לפרוטוקול רואה Willmund et al (2005) 10 CF1β הוא זרז כבקרת טעינה ונבחר.. כי זה הוא שופע, ולאחר תמוגה תא, מציג בשברים מסיסים וקרום. לחלופין, רמות של חלבונים מזהמים ניתן להשתמש כדי לנרמל לטעינה לא שוויונית.)
- אפשר חלבון לחרוזי Sepharose לIgGs לספוג במהלך דגירת 1-HR ב 25 ° C על גלגלת צינור (CAT RM5W, 36 סל"ד).
- צנטריפוגה ל60 שניות ב1000 XG ו4 המעלות צלזיוס לגלולת חלבון לחרוזי Sepharose. הסר בזהירות את supernatant ו Resuspend את החרוזים במאגר 5 מ"ל 0.1 מ 'נתרן borate (pH 9.0). חזור על פעולה זו שלוש פעמים ביסודיות כדי להסיר אמינים שלהרוות crosslinker.
- תשקול את 25.9 מ"ג של טרי, מוצק וdimethylpimelimidate resuspend במאגר זה 5 מ"ל 0.1 מ 'נתרן borate (pH 9.0) כדי להשיג ריכוז סופי של 20 מ"מ. הוסף פתרון זה לחלבון לחרוזי Sepharose.
- אפשר IgGs לקישור לעבור לחלבון למשך 30 דקות ב 25 ° C על גלגלת צינור.
- צנטריפוגה ל60 שניות ב1000 XG ו 4 ° C לגלולה את החרוזים. הסר בזהירות את supernatant ו resuspend את החרוזים ב5 המ"ל 1 M טריס-HCl (pH 7.5) כדי להרוות crosslinker חופשי. חזרו על פעולה זו פעם אחת והדגירה במשך 2 שעות ב 25 ° C או 12-24 שעות ב 4 ° C בגלגלת צינור.
- אופציונלי: אם חלבון לחרוזי Sepharose מצמידים IgGs לא משמשים ישירות עבור ה-IP, אחסון לתקופה של עד שבוע אחד בלבד. לשם כך, צנטריפוגות ל60 שניות ב1000 XG ו4 מעלות צלזיוס לגלולת החרוזים, להסיר בזהירות supernחרוזי atant וresuspend במאגר 5 מ"ל 0.1 מ 'פוספט (pH 7.5) המכילים 0.02% יזידו הנתרן ולאחסן ב 4 מעלות צלזיוס עד לשימוש נוסף.
2. הכנת תמוגה תא, Crosslinking ומדגם
- לגדול שתי תרבויות Chlamydomonas במדיום המכילות 7.5 מ"מ 14 NH 4 Cl או 15 NH 4 Cl כמקור חנקן לצפיפות של ~ 5 X 10 6 תאים / מ"ל. תאים צריכים לעבור לפחות עשרה דורות לתיוג מלא. כאן, תאים גודל במדיום photomixotrophically TAP 11 בסבובי ייקר ב 25 ° C תחת קרינה רציפה עם אור לבן (30 μE מ -2 של -1).
- העברה 2 aliquots כל אחד 14 N-15 ותאי N-כותרת לארבעה צינורות GSA ותאי קציר על ידי צנטריפוגה 4-דקות ב 4000 XG ו4 ° C (מספר התאים נקצר עבור כל aliquot תלוי בריכוז התאי של היעד חלבון וצרכימים שייקבעו דוארmpirically מראש על מנת להבטיח שחלבון המטרה המספק זרז. נקודת התחלה טובה היא 10 9 תאים לכל aliquot, כלומר, 200 מ"ל של תרבות עם 5 10 x 6 תאים / מ"ל.)
לcrosslinking בלבד: במקרה כזה, תהיה crosslinked מערכות חלבון לפני IP, תאים צריכים להיות שטף להסיר אמינים נוכחיים במדיום. לשם כך, תאי resuspend ב 40 מ"ל KH חיץ מראש מקורר (20 מ"מ HEPES-KOH (pH 7.2), 80 המ"מ KCl) ולהעביר אותם לצינורות 50-מיליליטר פלקון. צנטריפוגה ל60 שניות ב1000 XG ו4 ° C. חזור על פעולה זו פעם אחת. - תאי resuspend במאגר תמוגה מ"ל 2 (20 מ"מ HEPES-KOH (pH 7.2), 1 mM MgCl 2, 10 mM KCl, 154 mM NaCl) מראש מקוררים 4 מעלות צלזיוס ולהעביר אותם לצינורות 15-מיליליטר פלקון. איסוף תאים שנותרו בצינורות גז טבעי: עם חיץ נוסף 1 מיליליטר תמוגה כל אחד. הוסף 50 μl 25 x מעכב פרוטאז ו 12.5 μl M MgCl 1 2 (לריכוז סופי של 3.5 מ"מ) לכל aliquot.
- הוסף 150חיץ תמוגה μl, 12.5 μl 1 ז ATP, פוספט 833 μl 270 המ"מ קריאטין וphosphokinase 7 μl 5 מיקרוגרם / μl קריאטין (הריכוז הסופי הוא 2.5 מ"מ ATP, קריאטין פוספט mM 45, ו 7 מיקרוגרם / מ"ל phosphokinase קריאטין) לאחד את aliquots המכיל 14 N-15 ותאים שכותרתו N-(אלה + ATP aliquots).
- הוסף חיץ 930 μl תמוגה ו70 μl 1 U / μl apyrase לaliquots האחר המכיל 14 N-15 ותאים שכותרת N-(אלה aliquots-ATP).
- דגירה עבור 2 דקות ב 25 ° C על גלגלת צינור להקים ATP-לכלות ומדינות ה-ATP-גדושות. אם צעד crosslinking מושמט, להוסיף עוד 1 מ"ל של חיץ תמוגה.
לcrosslinking בלבד: במקרה את האינטראקציות נחקרו חלבונים הן חולפות רצוי ללכוד אותם על ידי צעד crosslinking. לשם כך, להוסיף 500 μl 20 מ"מ dithio-BIS (succinimidyl propionate) (DSP) מומס DMSO (ריכוז סופי הוא 2 מ"מ) לדוארצינור ach ישירות לפני sonication. - Sonicate ארבע פעמים 20 שניות על קרח לשבור תאים עם הפסקות 20-sec בבין לקירור. (אנו משתמשים Bandelin Sonoplus HD2070 עם KE76 קצה בשליטת תפוקה של 75% והמחזור של 60%. ההגדרות הדרושות למכונות אחרות / מכשירים / טיפים צריכות שתיקבענה מראש על מנת להבטיח תמוגה תא מלאה וכדי למנוע שפיכה. )
לcrosslinking בלבד: לאפשר מערכות חלבון לקישור לחצות ידי דוגר לשעה 1 ב 4 ° C בגלגלת צינור. לאחר crosslinking, להשלים כל צינור עם 500 μl M גליצין 1 ולדגור על גלגלת צינור למשך 15 דקות נוספות ב 4 ° C כדי להרוות crosslinker חופשי. - הכן ארבע כריות סוכרוז 6-מ"ל (20 מ"מ HEPES-KOH (pH 7.2), 0.6 סוכרוז M) בSW41 Ti הדק קיר צינורות (פריט Beckman Coulter לא: 344059), להניח בזהירות כל ~ 5.5 מ"ל של lysates הסלולרי על סוכרוז כריות (איזון עם חיץ תמוגה) וצנטריפוגה למשך 30 דקות ב 200.000 XG ו4 מעלות צלזיוס בSW41 טי rotor.
- העברת החלק העליון של השיפוע המכיל מתחמי חלבון מסיס לארבעה צינורות פלקון 15-מ"ל (להימנע מהעברת חלקים מכרית סוכרוז), להוסיף 350 10% μl טריטון X-100 לריכוז סופי של 0.5% לכל אחד מהם, לערבב בזהירות ולהוסיף למאגר תמוגה בהיקף כולל של 7 מ"ל כל אחת.
(העברה 70 μl של כל תמצית מסיסה תא לצינורות 1.5 מיליליטר חרוטים טריים (אפנדורף צינורות) ולהוסיף 70 μl 2 x חיץ SDS-מדגם (4% SDS, 125 המ"מ טריס-HCl (pH 6.8), גליצרול 20%, 10 % 2-mercaptoethanol) לכל אחד ל- PAGE SDS וניתוחי immunoblot.) - השלך את כריות סוכרוז וresuspend את הכדורים בממברנה במאגר תמוגה מ"ל 3 כ"א. הוסף לכל 1 מ"ל 10% טריטון X-100 לריכוז סופי של 2%, sonicate על קרח לפזר כדורים, ולהוסיף למאגר תמוגה בהיקף כולל של 5 מ"ל כל אחת.
- הכן ארבע כריות סוכרוז 6-מ"ל עוד בSW41 צינורות קיר דקים Ti, שכב ~ 5 המ"ל של קרומי solubilized מצעד 2.10 זהירות על סוכרוזכריות, וצנטריפוגה למשך 30 דקות ב 200.000 XG ו4 מעלות צלזיוס ברוטור טי SW41.
- העברת החלק העליון של השיפוע המכיל קומפלקסי חלבונים בממברנה לארבעה צינורות 15-מיליליטר פלקון ולהוסיף חיץ תמוגה (המכיל 2% Triton X-100) לנפח סופי של 7 מ"ל כל אחת. (העברה 70 μl של כל תמצית מסיסה תא לטרי צינורות Eppendorf ולהוסיף 70 μl 2 x-SDS מדגם זה לזה ל- PAGE SDS וניתוחי immunoblot).
3. Immunoprecipitation
- גלולת חלבון לחרוזים המכילים נוגדנים בשילוב Sepharose (מצעדי 1.8 או 1.9) על ידי צנטריפוגה 60-שניות ב1000 XG ו4 ° C, להסיר את supernatant וחרוזי resuspend זהירות ב4 חיץ תמוגה מ"ל. חזור על פעולה זו פעמים כדי לאזן חרוזים במאגר תמוגה.
- מלא עד 8 מ"ל עם חיץ תמוגה ולהעביר 1 מ"ל של השעיה לכל אחד משמונה שפופרות 15-מיליליטר פלקון המכילים מתחמי חלבון מסיסים או קרום מATP-ATP-גדוש ולרוקן 14 N-15 וN-מתויג תאים (מצעד 2.9 ו 2.12).
- דגירה עבור 2 שעות ב 4 ° C בגלגלת צינור לקומפלקסי חלבוני משקע.
- גלול את החרוזים על ידי צנטריפוגה 60-שניות ב1000 XG ו4 מעלות צלזיוס ולהשליך את supernatants. השאר נפח קטן של נוזל על גבי החרוזים כדי להקל על העברה.
- להעביר את החרוזים מכל צינור פלקון 1.5-מיליליטר חרוטי הצינורות (אפנדורף צינורות). לאסוף את כל החרוזים שנשארו בצינורות פלקון, להוסיף עוד 0.8 מיליליטר חיץ תמוגה המכיל טריטון% 0.1 לכל אחד, מערבולת בעדינות, צנטריפוגות ל60 שניות ב1000 XG ו4 מעלות צלזיוס ולהעביר חיץ עם חרוזים השיורי של חצוצרות Eppendorf. חילופי צינור הוא הכרחיים כדי למנוע זיהומים מחלבונים דבקות אל קירות הפלסטיק.
- גלול את החרוזים על ידי צנטריפוגה 15-שניות ב16100 XG ו4 ° C, להסיר את supernatants וחרוזי resuspend במאגר המכיל 1.3 מיליליטר תמוגה טריטון% 0.1 בזהירות. חזרו על פעולה זו פעמים עם buf תמוגהfer המכיל טריטון ופעמים עם טריטון חסר חיץ תמוגה ללשטוף ביסודיות את החרוזים. השאר נפח קטן של נוזל על גבי החרוזים כדי להקל על העברה.
- שוב להעביר את החרוזים לצינורות טריים 1.5-מיליליטר Eppendorf להסיר חלבונים דבוקים לקירות הפלסטיק. שטוף את הצינורות הישנים עם טריטון 1 מיליליטר תמוגה החיץ חסר ולהעביר את כל החרוזים שיורית לחצוצרות הטריות.
- צנטריפוגה במשך 15 שניות ב16100 XG ו4 ° C, להסיר את supernatants הראשון עם פיפטה רגילה, ואז להסיר כל שנותר supernatant לחלוטין עם מזרק המילטון 50-μl.
4. לדוגמא הכנה לnano-LC-MS/MS
- הוסף חיץ 100 μl טרי elution (8 M אוריאה, 25 מ"מ NH 4 3 HCO) לכל צינור, השתמש במאגר elution 100 μl לשטוף את החרוזים נדבקים למזרק ולדגור המילטון למשך 10 דקות בthermomixer ב 800 סל"ד ו 65 מעלות צלזיוס, ובמשך 20 דקות נוספות ב 30 ° C. (הרבה יותר מלאelution של חלבונים כפותים מושג על ידי elution עם 2% משקעי SDS ולאחר מכן עם אצטון 80%).
- צנטריפוגה במשך 15 שניות ב16100 XG ו25 ° C. העברת supernatants לצינורות טריים עם מזרק המילטון 50-μl.
- הוסף חיץ elution 50 μl לחרוזים, השתמש במאגר elution 50 μl לשטוף את החרוזים נדבקים למזרק המילטון וחזור על שלבי דגירה וצנטריפוגה 4.1 ו -4.2, בהתאמה. ברכת eluates בהתאמה.
(העברה 30 μl של eluates לצינורות Eppendorf טריים, להוסיף 30 μl 2 x חיץ SDS-מדגם זה לזה לSDS-PAGE וניתוחי immunoblot.) - שלב eluted משקע מטופל / ATP-+, 14 N-15 וN-מתויג חלבונים מסיסים וקרום כדלקמן:
120 μl 15 N / + μl ATP ו 120 14 N /-ATP
120 μl 14 N / + ATP ו120 μl 15 N /-ATP
120 μl 15 N / + μl ATP ו 120 14 N /-ATP
120 μl 14 N / + ATP ו120 μl 15 N /-ATP - הוסף 1.5 μl המוכן טרי DTT 1 ז לריכוז סופי של 6.5 מ"מ לכל אחד מארבעת הצירופים להפחתת חוב דיסולפיד (כולל אלו בcrosslinker) ודגירה במשך 30 דקות ב 25 ° C.
- הוסף 10.5 μl המוכן iodoacetamide M 0.6 טריים לריכוז סופי של 25 מ"מ לcarboxymethylate את thiols המופחת ודגירה במשך 20 דקות ב 25 ° C בחושך.
- הוסף 256 μl 40 המ"מ NH 4 3 ו 4 HCO μl Lys-C (0.1 מיקרוגרם / μl), צינורות חותמים עם parafilm, ודגירה במשך לפחות 16 שעתי לילה על גלגל סיבוב ב 37 ° C.
- הוסף 470 μl 20 המ"מ NH 4 3 HCO, 10 אצטוניטריל μl 100% (לריכוז סופי של% 1) ו 8 חרוזי טריפסין μl, ולדגור על גלגל סיבוב עבור לפחות 16 שעות ב 37 ° C.
- צנטריפוגה למשך 5 דקות ב16100 XG ו4 ° C, וההעברה לאמבטית supernatants טרי 2-מיליליטר אפנדורףes. שטוף את הצינורות הישנים עם 50 μl 20 המ"מ NH 4 3 HCO, חומצה אצטית 0.5%, וברכה עם supernatants 1.
- לdesalting, להכין תוצרת בית C 18-StageTips על ידי הכנת שני דיסקים מ 18 חומרי Empore C עם מחט מזרק ומעמיד אותם בקצה פיפטה 200-μl. בדרך זו להכין ארבעה טיפים 200-μl. לנקב חורים לתוך המכסים של ארבעה צינורות 2-מיליליטר Eppendorf וטיפים להוסיף.
- תנאי C 18-StageTips עם הפתרון B μl 50 (80 אצטוניטריל%, 0.5% חומצה אצטית). צנטריפוגה ל3 דקות ב 800 גרם ו 25 ° C.
- לאזן C 18-StageTips עם פתרון (0.5% חומצה אצטית, אצטוניטריל 2%) 100 μl. צנטריפוגה ל3 דקות ב 800 גרם ו 25 ° C. חזור על פעולה זו פעם אחת.
- טענת 100 μl של supernatants מdigestions tryptic (4.9) על 18-C StageTips וצנטריפוגה עבור 3 דקות ב 800 גרם ו 25 ° C. חזור על פעולה זו עד שsupernatants השלם יושמוהעמודות.
- שטוף את C 18-StageTips עם צנטריפוגה 100 μl הפתרון א 'ל3 דקות ב 800 גרם ו 25 ° C. חזרו על פעולה זו פעמים.
- Elute פפטידים tryptic לתוך צינור טרי 1.5-מיליליטר אפנדורף עם צנטריפוגה 50 μl פתרון לב 3 דקות ב 800 גרם ו 25 ° C. חזור על פעולה זו פעם אחת. פפטידים יבשים להשלמה במהירות VAC.
- אופציונלי: חותם אפנדורף צינורות עם parafilm ולאחסן ב -80 ° C עד שימוש נוסף.
- Resuspend את הפפטידים המיובשים עם פתרון μl 20 ודגירה במשך שעה לפחות 1 על קרח, שנקטעה על ידי שתי incubations 15-דקות באמבטית sonicator. צנטריפוגה במשך 20 דקות ב16100 XG ו4 מעלות צלזיוס, ולהחיל supernatant לnano-LC-MS/MS.
5. נציג תוצאות
כפי שניתן לראות exemplarily במשך 14 תמציות תאי N-הכותרת באיור 2A, HSP70B וCGE1 המותאמים לשפה מקומית כמעט אך ורק לחלק הקטן המסיס, בלתי תלוי במצב ה-ATP. בתוךלעומת זאת, CF1β הוא מקומי לשברים מסיסים וקרום מועשר, כsonication מזמרת חלק ממנו מהקרום ממוקם CF o, ולכן משמש כטעינת שליטה לשני השברים. כפי שמוצג באיור 2B, כמויות דומות של HSP70B היו זרזו עם הנוגדנים נגד HSP70B מ14 N-15 וN-שכותרת תמציות מסיסות ועצמאיות של מדינת ה-ATP. לעומת זאת, רק מעט היה HSP70B זרז משברים בממברנה עם כמויות גדולות במקצת ממקור שברי קרום ATP-מדולדל בהשוואה לשברים עתירים ה-ATP, ומכאן שמאשר תוצאות קודמות 9. לא CGE1 היה שיתוף זרז עם HSP70B בשברים מסיסים או קרום ATP-עתירים, ואילו כמויות גדולות של CGE1 נלקחו ביחד עם זרזו-HSP70B מATP-מדולדלים שברים מסיסים, וקצת משברים בממברנה ATP-מדולדל.
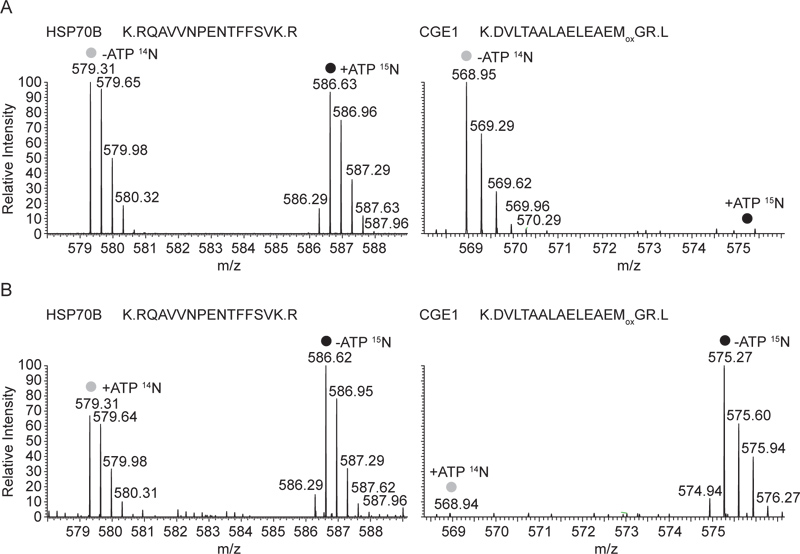
האינטראקציה של CGE1 עם HSP70B רק במדינת ADP הוא ציין גם בניתוח MS: בתרשים 3, MS1 ספקטרום נציג HSP70B וCGE1 פפטידים ממשקעים שנוצרו עם antiserum HSP70B מתמציות תאים מסיסות מוצגים. בניסוי שמוצג באיור 3 א, היו שוקע מתערובות של 14 תמציות N-כותרת חסרת ATP ו15 N-כותרת המכילות תמציות ה-ATP. בעוד הטופס, שהכותרת כבדה וקל של HSP70B הפפטיד התגלה בעוצמות שווות, רק בצורה של האור שכותרת CGE1 פפטיד (תמציות מ- ATP) נמצאה. באיור 3B אותם פפטידים מהמשקע אנטי HSP70B נגזר מתערובות של תמציות תאים מסיסות שבכותרת הדדית מוצגים. בהתאם לכך, זמן זה רק הטופס, שכותרתו הכבד של פפטיד CGE1 (תמציות מ- ATP) זוהה, ואילו זה היה שוב במקרה של שניהם, אור ופפטידים HSP70B כותרתו כבדים.

<חזק> איור 1. זרימת עבודה. תאים ניסיוניים מתויגים מטבולית עם 14 N ו 15 N במשך לפחות 10 דורות, שנקטפו ומסופקים עם או מדולדל מה-ATP. לאחר קומפלקסי חלבוני תמוגה תא אופציונלי ניתן crosslinked (X-קישור) עם DSP. תאי Lysed אז מופרדים במסיס (סול) ושברים בממברנה מועשרת (פל). חלבוני יעד (כאן HSP70B) וחלבון בקרה (כאן CF1β) הם immunoprecipitated עם נוגדנים ספציפיים לחלבון בשילוב חרוזי sepharose A (שחור). לאחר שטיפה, חלבונים זרזו הם eluted ובמישרין נותחו על ידי immunoblotting, או את 14 N-15 ובהתאמה N-כותרת שברים ב+ ATP ו- ATP המדינות נקוו, מתעכלות ונותחו על ידי nano-LC-MS/MS. במקרה הדוגמא שמוצג כאן 15 שבריר N-הכותרת היה מרוקן מה-ATP. פפטידים בהתאם לכך, היחס בין העוצמות (צבעים כהים) שכותרתו כבדים לאור כותרת (צבעים בהירים) מחלבון הבקרה(CF1β), חלבון המטרה (HSP70B) ושאינן מזהמים במיוחד בכריכה צריך להיות סביב 1, בעוד שיחס זה צפוי להיות גבוה מאוד לחלבונים במיוחד באינטראקציה עם חלבון המטרה (CGE1). לחץ כאן לצפייה בדמות גדולה.
{kind=link}

איור 2. ניתוח של הקלט עבור immunoprecipitation HSP70B. חלבון סה"כ היה שחולץ מן מסיס (סול) וקרום מועשר (פל) שברים מדולדלים או מATP (-ATP) או להשלים עם ה-ATP וה-ATP התחדשות מערכת (+ ATP). 0.01% מתמציות החלבונים הופרדו על 10% ג'ל SDS-polyacrylamide, ורמות של HSP70B וCGE1 חלבון יחסית לCF1β שליטת טעינה נותחו על ידי immunoblotting. ניתוח B של immunoprecipitates. HSP70B היה immunoprecipitated מN-14 וN-15 שכותרתו מסיסה בקרום תא ומועשר המכיל תמציות או חסר ה-ATP. חלבונים מתאימים ל3.3% מimmunoprecipitates הופרדו על 10% ג'ל SDS-polyacrylamide ורמות של HSP70B וCGE1 יחסית לטעינת CF1β שליטה נותחו על ידי immunoblotting. לחצו כאן לצפייה בדמות גדולה.
{kind=link}

איור 3. ספקטרום נציג המוני של HSP70B וCGE1 פפטידים מהנוגדים HSP70B immunoprecipitates בוצעו על שברים מסיסים מעורבים (14 N-ATP / 15 + N-ATP). ספקטרום MS מלא של 14 N ו 15 N מתויג פפטידים, המתאימים ל- ATP ו+ מדינות ה-ATP, בהתאמה, מHSP70B וCGE1 שיתוף immunoprecipitated מוצגות. שני פפטידים טעונים פי שלוש, פפטיד HSP70B מכיל 22 אטומי חנקן, CGE1 peptIDE 19, מקביל למעבר המוני של 7.33 מ 'ו6.33 / z, בהתאמה. ספקטרום המוני נציג B מניסוי הגומלין (14 + N-ATP / 15 N-ATP). ספקטרום מלא של MS את אותם 14 N ו 15 N מתויג פפטידים, כאן המתאים ל+ ATP ו- ATP המדינות, בהתאמה, מHSP70B וCGE1 השיתוף immunoprecipitated מוצגים. לחץ כאן לצפייה בדמות גדולה.
{kind=link}
Discussion
יש לנו הצגנו לאחרונה שני שיפורים לגישה המהירה: צעד crosslinking ללכידת אינטראקציות בין חלבונים חולפות (QUICK-X), ומשקעי שליטה לנרמל ליעילות משקעים שווה 6. כאן אנו מציגים פרוטוקול המכיל עוד שני שיפורים של מהיר: ראשון, אנחנו להחליף SILAC 5 עבור 15 N תיוג מטבולית. היתרונות הם כי 15 N תיוג מטבולית הם הרבה יותר זול מאשר SILAC, אם 15 N מסופק כמלח אורגני פשוט. יתר על כן, עם 15 N מטבולית תיוג מהיר יכול להיות מיושם על prototrophic אורגניזמים לכל החומצות האמינו, כמו רוב הצמחים, פטריות וחיידקים. ולבסוף, interconversion הטבוע SILAC 5,6 ארגינין לפרולין אינו מהווה בעיה לכימות של 15 N מתויג פפטידים. דוגמאות לכלים מתאימים להערכה כמותית של 15 נתוני N proteomics הן 12 MSQUANT אוOMIQS 13.
שנית, אנחנו מציגים את זיקת אפנון כאמצעי להפחתת כמות החלבונים באינטראקציה עם חלבון המטרה הניתן בדגימה אחת לעומת אחרת במפורש. היתרונות של גישה זו הם שהוא עוקף את הבנייה של מוטציות, שמציאה עבור כמה מערכות מודל קשה לייצר או לא יכול להיות שנוצרו בכל במקרה של חלבוני היעד חיוניים. יתר על כן, הוא נמנע מפרשנויות מוטעות הנגרמות על ידי ביטוי חלבון הפרש פוטנציאלי המתרחש כתגובה של התא לדפיקות למטה חלבון מטרה: אם חלבונים אחרים הם מוסדר כלפי מטה, כמו גם ולחצות-מגיבים עם antiserum משמש לimmunoprecipitation, הם יתפרשו כשותפים אמיתיים של אינטראקציה של חלבון המטרה. לבסוף, זיקת אפנון מבטל את הצורך של מציאת יחס נכון נוגדן לאנטיגן.
למרות שאנחנו מיישמים הפרוטוקול שלנו לreinhardtii Chlamydomonas כאורגניזם מודל, זה יכול בקלות להיות מותאם לכל אורגניזם אחר שניתן לגדל בתרבית תאים, והוא מסוגל להשתמש באמוניום חנק או כמקור חנקן. אפנון זיקה של קומפלקסי חלבונים על ידי ATP / ADP ישירות עשוי להיות מיושם למלווים אחרים שאינטראקציה עם מצעים וחלבוני עוקבה תלויה במצב ATP, כמו GroEL/HSP60/Cpn60 או Hsp90 מערכות מלווה 14,15, או לכל מערכת אחרת שבי זיקות מחייבות הם מווסתים על ידי ATP. זיקת אפנון צריך גם לעבוד למקרים בם זיקות בין אינטראקציות חלבון שונו על ידי תרופות מסוימות, כגון radicicol או geldanamycin במקרה של Hsp90 מערכות 15.
מגבלה ברורה של הפרוטוקול שלנו היא שזה דורש זיקה-מטוהרי נוגדנים נגד חלבון המטרה הידועה להיות רגיש לטיפול / תרופה שמודולציה זיקתו לחלבוני שותפים ספציפיים. לכן, אין זה שיטת תפוקה גבוהה.
Disclosures
אין ניגודי האינטרסים הכריזו.
Acknowledgements
אנו מודים לאוליבייה Vallon antiserum נגד CF1β. עבודה זו נתמכה על ידי אגודת מקס פלנק ומענקים מForschungsgemeinschaft דויטשה (Schr 617/5-1) וBundesministerium für Bildung und Forschung (FORSYS הביולוגיה המערכתית יוזמה, GoFORSYS פרויקט).
Materials
| Name | Company | Catalog Number | Comments |
| שם המגיב | חברה | מספר קטלוגים | תגובות (אופציונלי) |
| ProteinA Sepharose | סיגמה אולדריץ | P3391 | |
| DMP (דימתיל pimelimidate) | סיגמה אולדריץ | D8388 | אחסן מיובש ב -20 ° C, לפזר ישירות לפני השימוש |
| פרוטאז מעכב (מלא, ללא EDTA) | רוש יישומי מדע | 11873580001 | |
| ATP (Adenosin-5'-triphosphat) | קרל רוט | K054 | |
| קריאטין פוספאט | סיגמה אולדריץ | 27920 | |
| phosphokinase קריאטין | סיגמה אולדריץ | C7886 | ; |
| DSP Dithiobis [succinimidyl propionate] | Thermo Sientific | 22585 | אחסן מיובש ב 4 ° C |
| 15 NH 4 Cl | קיימברידג' איזוטופ מעבדות, אנדובר, מסצ'וסטס | NLM-467 | |
| Lys-C (Endoproteinase Lys-C) | רוש יישומי מדע | 11047825001 | |
| חרוזי טריפסין (Poroszyme ללא יכולת לזוז, טריפסין) | אפלייד Biosystems | 2-3127-00 | מערבב נמרצות ישירות לפני השימוש |
| דיסק Empore 18 C 47 מ"מ | ריאן | 12145004 |
References
- Perrakis, A., Musacchio, A., Cusack, S., Petosa, C. Investigating a macromolecular complex: the toolkit of methods. J. Struct. Biol. 175, 106-112 (2011).
- Gavin, A. C., Maeda, K., Kuhner, S. Recent advances in charting protein-protein interaction: mass spectrometry-based approaches. Curr. Opin. Biotechnol. 22, 42-49 (2011).
- Markham, K., Bai, Y., Schmitt-Ulms, G. Co-immunoprecipitations revisited: an update on experimental concepts and their implementation for sensitive interactome investigations of endogenous proteins. Anal. Bioanal. Chem. 389, 461-473 (2007).
- Selbach, M., Mann, M. Protein interaction screening by quantitative immunoprecipitation combined with knockdown (QUICK. Nat Methods. 3, 981-983 (2006).
- Ong, S. E., Mann, M. A practical recipe for stable isotope labeling by amino acids in cell culture (SILAC). Nat Protoc. 1, 2650-2660 (2006).
- Heide, H. Application of quantitative immunoprecipitation combined with knockdown and cross-linking to Chlamydomonas reveals the presence of vesicle-inducing protein in plastids 1 in a common complex with chloroplast HSP90C. Proteomics. 9, 3079-3089 (2009).
- Nordhues, A., Miller, S. M., Muhlhaus, T., Schroda, M. New insights into the roles of molecular chaperones in Chlamydomonas and Volvox. International review of cell and molecular biology. 285, 75-113 (2010).
- Mayer, M. P., Bukau, B. Hsp70 chaperones: cellular functions and molecular mechanism. Cell Mol. Life Sci. 62, 670-684 (2005).
- Schroda, M., Vallon, O., Whitelegge, J. P., Beck, C. F., Wollman, F. A. The chloroplastic GrpE homolog of Chlamydomonas: two isoforms generated by differential splicing. The Plant Cell. 13, 2823-2839 (2001).
- Willmund, F., Schroda, M. HEAT SHOCK PROTEIN 90C is a bona fide Hsp90 that interacts with plastidic HSP70B in Chlamydomonas reinhardtii. Plant Physiol. 138, 2310-2322 (2005).
- Harris, E. H. The Chlamydomonas Sourcebook: Introduction to Chlamydomonas and Its Laboratory Use. , Elsevier, Academic Press. San Diego, CA. (2008).
- Mortensen, P. MSQuant, an open source platform for mass spectrometry-based quantitative proteomics. J. Proteome Res. 9, 393-403 (2010).
- M?hlhaus, T., Weiss, J., Hemme, D., Sommer, F., Schroda, M. Quantitative Shotgun Proteomics Using a Uniform 15N-Labeled Standard to Monitor Proteome Dynamics in Time Course Experiments Reveals New Insights into the Heat Stress Response of Chlamydomonas reinhardtii. Mol. Cell Proteomics. 10, (2011).
- Bukau, B., Horwich, A. L. The Hsp70 and Hsp60 chaperone machines. Cell. 92, 351-366 (1998).
- Wandinger, S. K., Richter, K., Buchner, J. The Hsp90 chaperone machinery. J. Biol. Chem. 283, 18473-18477 (2008).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved