
Method Article
לימוד TGF-β איתות ו TGF-β-induced אפיתל-ל-mesenchymal מעבר בסרטן השד ותאים נורמליים
In This Article
Erratum Notice
Summary
אנו מתארים זרימת עבודה שיטתית כדי לחקור איתות TGF-β ו- EMT הנגרמת על ידי β TGF על ידי לימוד ביטוי החלבון והגנים המעורבים במסלול איתות זה. השיטות כוללות סופג מערבי, בדיקת כתב לוציפראז, qPCR, וכתמי כשל חיסוני.
Abstract
שינוי פקטורי גדילה-β (TGF-β) הוא גורם רב תכליתי מופרש הממלא תפקיד מפתח בתקשורת בין-תאית. הפרעות של איתות TGF-β יכול להוביל לסרטן השד. TGF-β מעורר את השפעותיו על התפשטות ובידול באמצעות משטח תא ספציפי TGF-β סוג I קולטנים מסוג II (כלומר, TβRI ו TβRII) המכילים תחום פנימי סרין / threonine קינאז. על היווצרות מורכבת הטרומרית הנגרמת על ידי TGF-β, מופעל TβRI מעורר איתות תאי על ידי phosphorylating SMAD2 ו SMAD3. אלה SMADs מופעל טופס קומפלקסים הטרומריים עם SMAD4 כדי לווסת גנים היעד ספציפי, כולל מעכב הפעלת פלסמינוגן 1 (PAI-1, מקודד על ידי הגן SERPINE1). האינדוקציה של מעבר אפיתל-mesenchymal (EMT) מאפשר תאים סרטניים אפיתל באתר הראשי או במהלך קולוניזציה באתרים מרוחקים כדי להשיג פנוטיפ פולשני ולהניע התקדמות הגידול. TGF-β פועל כגרום חזק של פלישה לסרטן השד על ידי נהיגה EMT. כאן, אנו מתארים שיטות שיטתיות לחקור TGF-β איתות ותגובות EMT באמצעות תאים MCF10A-RAS אנושיים קדם-מיניים (M2) ותאי אפיתל NMuMG העכבר כדוגמאות. אנו מתארים שיטות כדי לקבוע TGF-β-induced SMAD2 זרחון על ידי סופג המערבי, SMAD3 / SMAD4 תלוי פעילות שעתוק באמצעות פעילות כתב לוציפראז ו SERPINE1 ביטוי גנים היעד על ידי תגובת שרשרת כמותית בזמן אמת פולימראז (qRT-PCR). בנוסף, מתוארות שיטות לבחון את ה- EMT הנגרמת על ידי TGF-β על ידי מדידת שינויים במורפולוגיה, ביטוי סמן אפיתל ומזנכימלי, כתמי אקטין סיביים וכתם כשל חיסוני של E-cadherin. שני מעכבי קינאז קולטן TGF-β מולקולה קטנה סלקטיבית, GW788388 ו- SB431542, שימשו לחסימת זרחון SMAD2 המושרה β על ידי TGF, גנים ממוקדים ושינויים בביטוי סמן EMT. יתר על כן, אנו מתארים את ההשתנות של השד mesenchymal Py2T תאי גידול אפיתל מורין לתוך adipocytes. שיטות לבחון TGF-β-induced איתות EMT בסרטן השד עשוי לתרום גישות טיפוליות חדשות לסרטן השד.
Introduction
ציטוקינים שינוי פקטורי גדילה-β (TGF-β) הוא אב טיפוס של קבוצה גדולה של פוליפפטידים רגולטוריים הקשורים מבנית ותפקודית כולל TGF-βs (כלומר, TGF-β1, -β2 ו -β3), חלבונים מורפוגניים עצם (BMPs) ו activins1,2. ציטוקינים אלה ממלאים תפקידים חשובים בהתפתחות העוברית ובתחזוקת הומאוסטזיס רקמות ואיברים3. misregulation של TGF-β יכול להוביל למגוון רחב של מחלות, כולל סרטן4,5. TGF-β ממלא תפקיד מורכב, כפול בהתקדמות סרטן: בתאי אפיתל נורמליים טרום מלמיניים, TGF-β מתנהג כמו מדכא גידול על ידי עיכוב התפשטות וגרימת אפופטוזיס6,7; עם זאת, בשלב המאוחר של התקדמות הגידול, כאשר תגובות ציטוסטטיות נחסמות על ידי הפעלה של אונקוגנים או אובדן של גנים מדכאי גידולים, TGF-β פועלת כמשפרת גידולים על ידי קידום מעבר אפיתל-mesenchymal (EMT) בתאי סרטן, ובכך לאפשר פלישת תאים סרטניים גרורות, הפועל על תאים microenvironment הגידול, וגירוי אנגיוגנזה והתחמקות החיסון8,9,10.
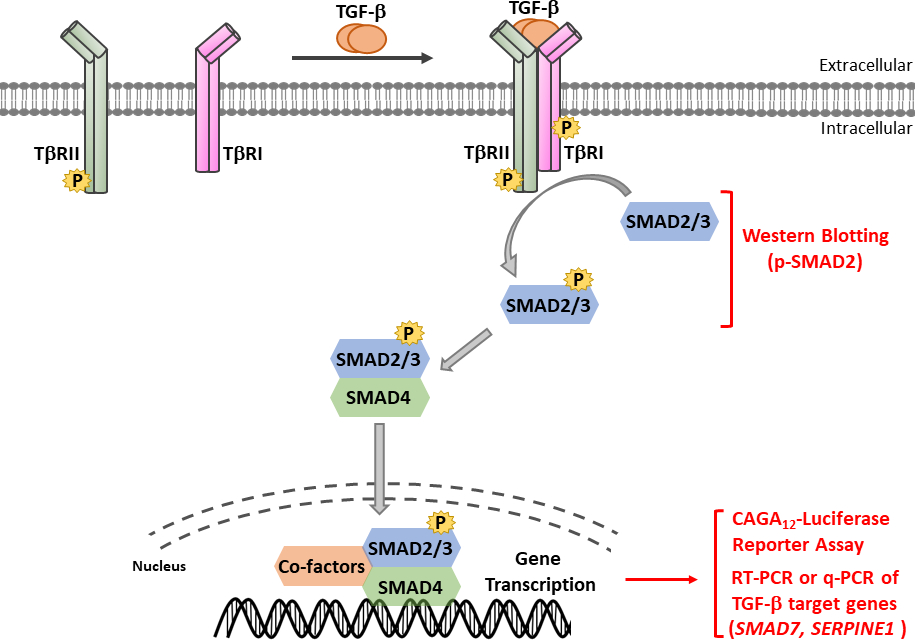
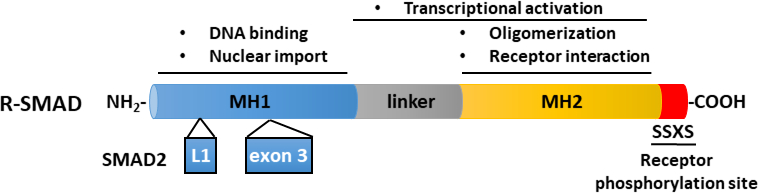
TGF-β מופרש כמו מולקולת קודמן לא פעיל המכיל את β TGF-מסוף קרבוקסיה בוגרת ופפטיד הקשורים להשהיה (LAP)11. קומפלקס קטן זה יכול להיות כבול בקובאליות על ידי חלבון סמוי TGF-β מחייב (LTBP)12. שחרורו של בוגר TGF-β יכול להיות מתווך על ידי פעולה של פרוטאזות ספציפיות כי cleave LAP או על ידי משיכה מכנית של LAP בתהליך תלוי integrin13,14. בנוסף ל- LTBP, גליקופרוטאין A חוזר דומיננטי (GARP) מתבטא מאוד על פני השטח של תאי T רגולטוריים (Tregs) וממלא תפקיד דומה לזה של LTBP בוויסות ההפעלה של TGF-β15,16. GARP נקשר ישירות β TGF-β סמויים באמצעות הצמדה דיסולפידית ושיוך לא-קוולנטי. ההפעלה של TGF-β ממתחם GARP / TGF-β דורשת אונטגרינים17. בוגרת TGF-β נקשר TGF-β קולטני סרין / threonine קינאז, כלומר, TGF-β סוג I (TβRI) ו TGF-β סוג II (TβRII) קולטנים18 ליזום איתות. הכריכה של TGF-β ל- TβRII מקדמת את גיוס TβRI והקמת מתחם הטרומרי. לאחר מכן, TβRI הוא phosphorylated על ידי קינאז TβRII על שאריות סרין ותרונין במוטיב קצר גליצין- וסרין עשיר (GS), וכתוצאה מכך הפעלתו19,20. עם ההפעלה, TβRI מופעל מגויסים phosphorylates המצעים שלה: שני SMADs ספציפי לקולטן (R-SMADs) הכוללים SMAD2 ו SMAD3 (איור 1). R-SMADs חולקים מבנה כולל דומה עם שני תחומים שנקראים Mad homology, MH1 ו- MH2, המופרדים באמצעות אזור קישור עשיר בפרולין (איור 2). מוטיב כריכת ה- DNA בתחום MH1 של SMAD3 אינו נשמר בין SMAD2 ל- SMAD3, ו- SMAD2 אינו יכול לאגד ישירות DNA עקב שתי הוספות בתחום MH1 שלו (exon 3 ו- L1). SMAD2 ו-SMAD3 יכולים להיות מופעלים על ידי הזרחון של מוטיב ה-SSXS בטרמיני C שלהם (איור 2). Phosphorylated SMAD2/3 יוצר מתחמים הטרומריים עם מתווך SMAD משותף, SMAD4, אשר טרנסלוקציה לתוך הגרעין כדי לווסת את שעתוק של גנים היעד (איור 1)7,21. מסלול איתות SMAD קנוני זה מוסדר במדויק ומייצר תגובות ספציפיות לתאים ולרקמות כגון ויסות גורל התא וגרורות תאים סרטניים ופלישה22. בנוסף לאיתות TGF-β-SMAD, מסלולי איתות שאינם SMAD יכולים להיות מופעלים ישירות גם על ידי קולטנים כדי לווסת תגובות סלולריות במורד הזרם23.
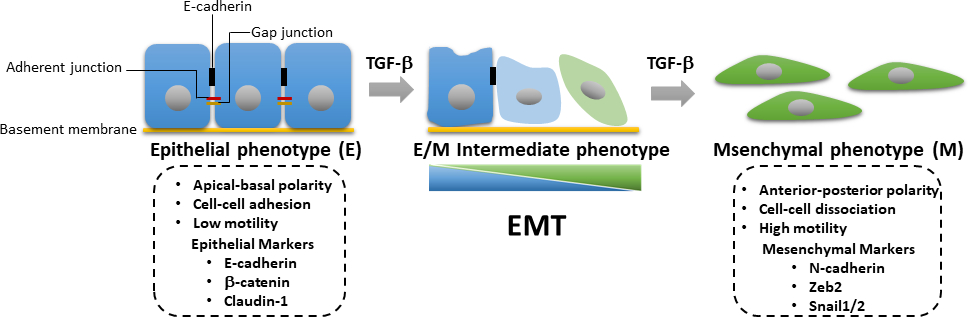
במהלך התקדמות הגידול, ההפעלה של TGF-β-induced SMAD-induced מסלולים עצמאיים SMAD נדרשים אינדוקציה של EMT. EMT הוא תהליך הפיך שבו תאים סרטניים dedifferentiate מפנוטיפ אפיתל, אשר קשורה לאובדן של מגעים תא-תא וירידה קוטביות אפית-בסיסית, כדי פנוטיפ mesenchymal עם תנועתיות משופרת ויכולת הפלישה24. EMT מאופיין בביטוי מוגבר של חלבוני סמן mesenchymal, כולל N-cadherin, vimentin, Zeb2 ו חילזון1/2, ואת הסרת הפיקוח בו זמנית של סמני אפיתל, כגון E-cadherin ו β-catenin (איור 3)25. עם זאת, המעבר מאפיתל למצב mesenchymal הוא לעתים קרובות לא שלם, ותאים לקבל מאפיינים אפיתל מעורב mesenchymal (E/M). מאמר שפורסם לאחרונה על ידי האגודה הבינלאומית EMT הציע לתאר את התהליך של תאים שעברו מצבי ביניים E / M phenotypic כמו פלסטיות אפיתל-mesenchymal (EMP)26. פלסטיות זו מתייחסת EMT חלקית, מצב E / M היברידית, מצב EMT metastable, רצף EMT ספקטרום EMT26. במהלך EMT, תאים סרטניים להשיג תכונות תא גזע סרטני (CSC) ולהיות עמידים יותר אפופטוזיס הנגרמת על ידי ניתוק27. בעוד EMT אחראי על רכישת פנוטיפ פולשני בתאים סרטניים ראשוניים ומניע התקדמות סרטן, לעומת זאת, מעבר mesenchymal-אפיתל (MET) הוכח לשחק תפקיד חשוב בצמחייה של תאים סרטניים שהופצו באתרים גרורתיים רחוקים28,29. מחקר שנערך לאחרונה הראה כי תאים סרטניים השד נגזר EMT ניתן transdifferentiated לתוך adipocytes, אשר עשוי להציע הזדמנות לעכב גרורות להתגבר על עמידות לטיפול בתאי הגידול וסרטן התדרדר30. בשל התפקיד החשוב של TGF-β איתות בהפעלה של EMT ב carcinogenesis השד, אנו מציגים פרוטוקולים מפורטים עבור סופג המערבי, בדיקת שעתוק לוציפראז, תגובת שרשרת כמותית בזמן אמת פולימראז (qRT-PCR), ו immunofluorescence לחקירה של TGF-β איתות, TGF-β-induced EMT, ואת transdifferentiation של תאי גידול אפיתל השד מורין נגזר EMT לתוך אדיפוציטים. טכניקות אלה הן הכלים האנליטיים הנפוצים ביותר בתחום הביולוגיה של התא. qRT-PCR משמש לזיהוי, אפיון וכימות רמות ביטוי mRNA באופן כמותי. בהשוואה ל- PCR כמותי (qPCR), טכניקה חלופית, ניתן להשתמש בתעתיק ההפוך (RT)-PCR כדי לקבוע ביטוי mRNA באופן חצי כמותי31,32. כתמים מערביים משמשים לבחינת רמות חלבון ספציפיות במדגם ליסנט תא נתון עם יתרונות של רגישות וספציפיות, באופן חצי כמותי. לכן, אנו מציגים זרימת עבודה שיטתית כדי לנתח שינויים מביטוי גנים לביטוי חלבון כדי לעזור לחקור איתות TGF-β שניתן להחיל גם על מסלולי איתות אחרים.
Protocol
1. ניתוח של זרחון SMAD2 הנגרמת על ידי TGF-β באמצעות סופג מערבי
הערה: תאי MCF10A-Ras של השד האנושי הפרה-מיניים שימשו כדוגמה לחקירת תגובות איתות TGF-β33. באופן עקרוני, השיטות המתוארות להלן חלות גם על קווי תאים אחרים בעלי תגובה β TGF.
- תרבות קו תא אפיתל השד MCF10A-ראס ב 37 °C (77 °F) במדיום הנשר שונה של Dulbecco (DMEM)/F12 המכיל L-גלוטמין עם 5% סרום סוס, 20 ng/mL גורם גדילה אפידרמיס (EGF), 10 מ"ג / מ"ל אינסולין, 100 ng / mL כולרה enterotoxin, 0.5 מ"ג / מ"ל הידרוקורטיזון, ו 1:100 פניצילין-סטרפטומיצין (עט-סטרפטוצין).
- טריפסינייז MCF10A-ראס תאים עם 1 מ"ל של 0.25% טריפסין-EDTA במשך 1 דקה ולספור תאים קיימא באמצעות מונה התא.
- תאי זרע לתוך צלחות 6-באר בצפיפות של 5×105 תאים / גם.
- לאחר צמיחה של לילה, לטפל בתאים עם או TGF-β (5 ng/mL) או חיץ ליגנד (4 mM HCl, 0.1% שומן ללא חומצת שומן סרום אלבומין (BSA)) במשך 1 שעה, ולאחר מכן להסיר מדיום תרבות בעדינות לשטוף את התאים פעמיים עם 1 מ"ל של תמיסת מלח פוספט אגירה (PBS).
- תאים מגניבים בצלחות 6 בארות על קרח ומוסיפים 150 μL של מבחני משקעים חיסוניים רדיו (RIPA) מאגר תמוגה (150 mM NaCl, 0.1% טריטון X-100, 0.5% נתרן deoxycholate, 0.1% SDS, 50 מ"מ Tris-HCl pH 8.0, וקוקטייל מעכב פרוטאז מיני הוסיף טרי). אפשר לתמוגה להמשיך על הקרח במשך 10 דקות.
- לגרד תאים חסידים את המנה באמצעות מגרד תא פלסטיק, ואז בעדינות להעביר את המתלה התא לתוך צינור microcentrifuge precooled.
- צנטריפוגה התא lysate במשך 10 דקות ב 150 כוח צנטריפוגלי (x g) ב 4 מעלות צלזיוס ולהעביר את supernatant לצינור טרי 1.5 מ"ל microcentrifuge.
- למדוד את ריכוז החלבון באמצעות ערכת מבחני חלבון תואמת חומר ניקוי (DC).
- טען 30 מיקרוגרם חלבון מכל דגימה על 10% נתרן דודקיל סולפט פוליאקרילמיד ג'ל אלקטרופורזה (SDS-PAGE) ג'ל ולהפעיל את הג'ל במתח של 100 V במשך 1.5-2 שעות.
- מעבירים את החלבונים מהג'ל לקרום פוליווינילידן דיפלוריד (PVDF) של 45 מיקרומטר במתח של 110 וולט למשך 1-1.5 שעות.
- מעבירים את קרום PVDF למיכל מתאים עם צד החלבון (הצד שפונה לג'ל) למעלה, ושוטף בקצרה את הקרום במים מזוקקים.
- להשליך את המים, להוסיף פתרון פונסאו S, ולשים את הממברנה על פלטפורמת נדנדה במשך 1-2 דקות.
- דה כתם הממברנה עם מים מזוקקים על ידי שטיפה מהירה ולאחר מכן לשטוף אותו במשך 1 דקה.
- לאחר מכן, שים את קרום PVDF על קופסת אור, וצלם תמונה כדי לבדוק אם יש עומסי חלבון שווים.
- לשטוף את הממברנה עם מלוחים טריס-אגירה עם Tween 20 (TBST, 20 mM Tris-HCl, pH 7.4, 150 mM NaCl, ו 0.1% Tween 20) עד שאין כתמים גלויים.
- שים את הממברנה בחסימת חיץ (5% חלב רזה בתמיסת TBST) ודגירה אותו במשך שעה בטמפרטורת החדר.
- לשטוף את הממברנה פעמיים עם TBST.
- דגירה הממברנה עם נוגדנים ראשוניים נגד זרחן-SMAD2 (p-SMAD2; 1:1000, תוצרת בית 34),סה"כ SMAD2/3 1:1000 ו glyceraldehyde 3-פוספט דהידרוגנאז (GAPDH; 1:1000), לילה ב 4 מעלות צלזיוס.
- לשטוף את הממברנה פעמיים עם TBST ולהדגיר את הממברנה עם נוגדן משני נגד ארנב או עכבר (1:5000) במשך 2 שעות.
- דגירה קרום PVDF עם מצע ECL המערבי במשך 30 שניות, ולזהות את האות באמצעות מערכת הדמיה.
- חזור על הניסויים לפחות שלוש פעמים כדי להשיג משולשים ביולוגיים.
2. ניתוח של תגובות תמלול תלויות SMAD3 הנגרמות על ידי TGF-β
- בצע את ה-CAGA12תלוי SMAD3/SMAD4 -לוציפראז שעתוק כתב assay.
- תרבות ותאי MCF10A-Ras עם ניסיון כמתואר בשלב 1. תאי זרע לתוך צלחות 24-באר ב 5×104 תאים / באר ולאפשר לתאים לדבוק בן לילה.
- יום לאחר הזריעה, cotransfect התאים בכל באר עם 100 ng של TGF-β / SMAD3-בלתי ניתן להקרבה (CAGA)12 כתב שעתוק לוציפראז לבנות35 ו 80 ng של מבנה ביטוי β-גלקטוזידאז באמצעות פוליאתילנימין (PEI). ההעתקה של β-גלקטוזידאז משמשת לנרמול הבדלים ביעילות התת-הדבקה בין בארות שונות. להקים כל קבוצת ניסוי משולשת.
- לאחר 24 שעות של דגירה, תאים גוועים ברעב עם גלוקוז גבוה DMEM ללא סרום, 6 שעות מאוחר יותר, להוסיף TGF-β (5 ng/mL) או חיץ ליגנד (4 mM HCl, 0.1% BSA) כבקרת רכב לתאים.
- לאחר עוד 24 שעות של דגירה, לשטוף תאים פעמיים עם PBS מראש.
- מוסיפים 120 μL / גם 1× מאגר תמוגה בעדינות לנער את הצלחת ב 4 מעלות צלזיוס במשך 20 דקות.
- העבר 30 μL של lysate לכל באר של microplate 96-well לבן assay כדי למדוד פעילות לוציפראז באמצעות לומינומטר.
- העבר 50 μL של lysate לכל באר של צלחת שקופה 96-היטב כדי למדוד פעילות β-גלקטוזידאז.
- לנרמל את פעילות לוציפראז לפעילות β-גלקטוזידאז ולחזור על הניסויים לפחות שלוש פעמים כדי להשיג משולשים ביולוגיים.
- לנתח את הביטוי של TGF-β גנים היעד באמצעות תגובת שרשרת כמותית בזמן אמת פולימראז (qRT-PCR).
- תרבות ותאי MCF10A-Ras עם ניסיון כמתואר בשלב 1. תאי זרע לתוך צלחות 6-באר ב 5×105 תאים / באר ולאפשר לתאים לדבוק בן לילה.
- לטפל בתאים עם TGF-β (5 ng/mL) או מאגר ליגנד (4 mM HCl, 0.1% BSA) במשך 6 שעות, ולאחר מכן לשטוף את התאים פעמיים עם 1 מ"ל של PBS.
- לבודד את ה-RNA הכולל באמצעות ערכת בידוד RNA.
- לקבוע את ריכוז RNA עם NanoDrop ולבצע סינתזת cDNA עם 1 מיקרוגרם של RNA באמצעות ערכת סינתזה cDNA גדיל הראשון.
- השתמש במערכת זיהוי PCR בזמן אמת כדי לבצע כמותי בזמן אמת PCR (qRT-PCR) עם cDNA מדולל פי עשרה בתערובת תגובת μL 10 הכוללת פריימרים אנושיים ספציפיים קדימה והפוך עבור GAPDH (לנורמליזציה), SERPINE1 (קידוד חלבון PAI-1, גן יעד TGF-β/SMAD), SMAD7 (גן יעד TGF-β/SMAD) ו- qPCR מאסטר מיקס. להקים כל קבוצת ניסוי משולשת.
הערה: רצפי פריימר המשמשים לזיהוי גנים אנושיים ממוקדים ב qRT-PCR מפורטים בטבלה 1. - השתמש בתנאי התגובה הבאים qPCR: אתחול, 95 °C (69 °F) במשך 3 דקות; דנטורציה, 95 מעלות צלזיוס למשך 10 שניות; חישול, 60 מעלות צלזיוס למשך 30 שניות; והרחבה, 80 מעלות צלזיוס למשך 10 שניות; דנטורציה, חישול והרחבה חוזרים על עצמם 40 פעמים.
- חזור על הניסויים לפחות שלוש פעמים כדי להשיג משולשים ביולוגיים.
3. ניתוח של EMT הנגרמת על ידי TGF-β
- לנתח את הביטוי של סמני EMT ברמת החלבון באמצעות סופג מערבי.
הערה: הניתוח של EMT הנגרמת על ידי TGF-β מוצג באמצעות תאי אפיתל NMuMG העכבר כדוגמה36,37.- תרבות NMuMG תאים ב 37 °C (70 °F) במדיום גלוקוז DMEM גבוה בתוספת 10% סרום שור עוברי ו 1:100 עט סטרפטוקוקוס. השתמש בשיטות המתוארות בשלב 1 לבידוד וזיהוי חלבונים.
הערה: הנוגדנים הבאים משמשים בניסוי זה: E-cadherin, 1:1000; נ-קדרין, 1:1000; חילזון, 1:1000; שבלול, 1:1000; טובולין, 1:1000(איור 3).
- תרבות NMuMG תאים ב 37 °C (70 °F) במדיום גלוקוז DMEM גבוה בתוספת 10% סרום שור עוברי ו 1:100 עט סטרפטוקוקוס. השתמש בשיטות המתוארות בשלב 1 לבידוד וזיהוי חלבונים.
- נתח את הביטוי של סמני EMT ברמת mRNA באמצעות PCR כמותי בזמן אמת כמתואר בשלב 2.2.
הערה: כל פריימרים לעכברים המשמשים עבור qRT-PCR, כולל CDH1 (קידוד חלבון E-cadherin), SNAILו- ZEB2 (איור 3) מפורטים בטבלה 1. - לנתח את תהליך EMT באמצעות שפעת חיסונית עקיפה וכתם פלואורסצנטי ישיר.
- כתמי כשל חיסוני עקיף של E-cadherin
- מניחים כיסויי זכוכית מרובעים סטריליים בגודל 18 מ"מ בצלחות של 6 בארות (כיסוי אחד לבאר).
- זרע 1×105 תאי NMuMG עם 2 מ"ל של DMEM מלא לכל צלחת 6 באר ולאפשר לתאים לדבוק בן לילה.
- מעבירים בעדינות את הכיסויים עם תאים דבקים לצלחת חדשה של 6 בארות ומוסיפים 2 מ"ל של מדיום תרבות לבארות.
- לטפל בתאים עם TGF-β (5 ng/mL) או מאגר ליגנד (4 mM HCl, 0.1% BSA) במשך 2 ימים.
- הסר את מדיום התרבות בעדינות לשטוף את התאים פעמיים עם 1 מ"ל של PBS מראש.
- לתקן את התאים על ידי הוספת 1 מ"ל של 4% paraformaldehyde ודגרת במשך 30 דקות בטמפרטורת החדר. לאחר מכן, לשטוף בעדינות את התאים פעמיים עם 1 מ"ל של PBS.
- לחלחל לתאים הקבועים עם 0.1% טריטון X-100 במשך 10 דקות בטמפרטורת החדר ולשטוף את התאים פעמיים עם PBS.
- חסום את התאים עם 5% BSA ב PBS במשך שעה אחת בטמפרטורת החדר ולשטוף את התאים פעמיים עם PBS.
- הוסף את הנוגדן העיקרי נגד E-cadherin (מדולל 1:1000 ב PBS) לחלק העליון של כל כריכה דגירה במשך 1 שעה בטמפרטורת החדר.
- הסר את הנוגדן העיקרי ולשטוף את הכיסוי עם PBS שלוש פעמים.
- הוסף את הנוגדן המשני Alexa Fluor 555 (מדולל 1:500 ב- PBS) לחלק העליון של כל כיסוי ודגר במשך שעה בטמפרטורת החדר תוך כיסוי בנייר אלומיניום כדי להגן מפני אור.
- הסר את הנוגדן המשני ולשטוף את הכיסוי עם PBS שלוש פעמים.
- הר את הכיסוי (תאים הפונים כלפי מטה) על שקופיות זכוכית באמצעות מדיום הרכבה עם 4′,6-diamidino-2-פנילינדול (DAPI) ולאחסן את השקופיות רכוב בתיבה ב 4 °C (6 °F), מוגן מפני אור.
- שימו לב לכתמים באמצעות מיקרוסקופיה קונפוקלית SP8.
- כתמי פלואורסצנטיות ישירים של סיבים (F)-actin.
- הכן דגימות לפי שלבים 3.3.1.1. ל-3.3.1.9.
- הכתים את התאים על ידי הוספת Alexa Fluor 488 Phalloidin (1:1000) במשך שעה אחת בטמפרטורת החדר בחושך כדי לדמיין אקטין סיבי (F-actin).
- לשטוף תאים שלוש פעמים עם PBS.
- טען את החלקה על שקופיות זכוכית באמצעות מדיום הרכבה עם DAPI וצלם תמונות באמצעות מיקרוסקופיה קונפוקלית SP8.
- כתמי כשל חיסוני עקיף של E-cadherin
תוצאות
ניתוח איתות TGF-β
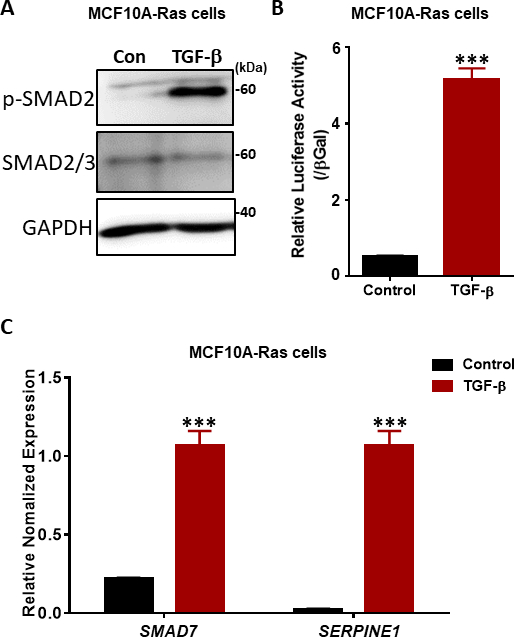
השלב המרכזי באיתות TGF-β הוא הזרחון של שתי שאריות הסרין של מסוף הקרבוקסי ביותר במוטיב SSXS(איור 2)מאת TβRI kinase38,39. כדי לחקור תגובות איתות TGF-β, ביצענו סופג המערבי של SMAD2 זרחן. בתאי ה-MCF10A-Ras של השד האנושי, הזרחן של SMAD2 גדל באופן משמעותי בתגובה לגירוי β TGF במשך שעה, בעוד שהביטוי של SMAD2/3 הכולל לא הושפע מטיפול TGF-β (איור 4A). על ידי שימוש ב-SMAD3/4-induced TGF-β-induced CAGA-luc שעתוק הכתב assay, מצאנו כי TGF-β המושרה במידה ניכרת כתב לוציפראז בקו תאים MCF10A-ראס בהשוואה לתאים שאינם מטופלים (איור 4B). יתר על כן, ראינו כי מטרות גנים שעתוק ישיר מאופיין היטב של TGF-β כולל SMAD7 ו SERPINE1 (קידוד חלבון PAI-1), באו לידי ביטוי מאוד בתאי MCF10A-Ras שטופלו β TGF(איור 4C).
ניתוח של EMT הנגרמת על ידי TGF-β
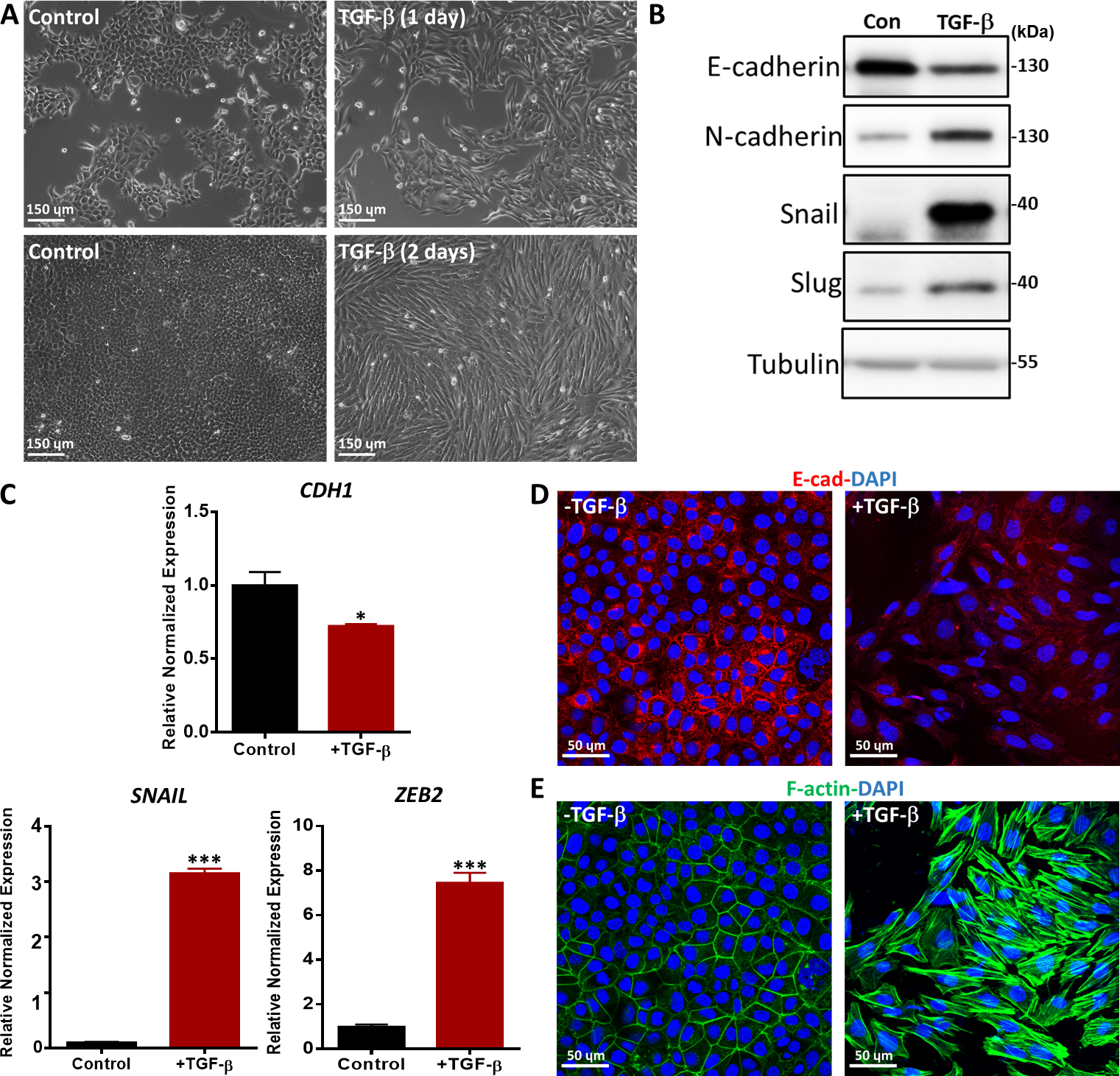
הערכנו את ה- EMT המושרה על ידי TGF-β בשיטות שונות, כגון שינויים מורפולוגיים, ביטוי של סמני EMT ברמות mRNA וחלבון וכתמים אימונופלואורסצנטיים של סמני EMT36. תאי אפיתל NMuMG שטופלו β TGF במשך יום ויומיים השתנו ממורפולוגיה אפיתל קלאסית למורפולוגיה דמוית מסנצ'יאל בצורת ציר, כפי שמוצג על ידי מיקרוסקופיית ניגוד פאזה (איור 5A). בהתאם לשינויים המורפולוגיים, ראינו כי טיפול TGF-β הוביל לעלייה בביטוי החלבון של סמנים mesenchymal, כולל N-cadherin, חילזון, ו שבלול37 (איור 5B). לעומת זאת, E-cadherin, סמן אפיתל, היה downregulated בתאי NMuMG לאחר 2 ימים של טיפול TGF-β (איור 5B). בנוסף, ביצענו תגובת שרשרת כמותית בזמן אמת פולימראז (qRT-PCR) כדי לחקור את ביטוי הגנים של סמני EMT. CDH1 (קידוד חלבון E-cadherin) ירד באופן משמעותי, בעוד סמנים mesenchymal כגון SNAIL ואבץ אצבע E-box מחייב homeobox 2 (ZEB2) גדלו במידה ניכרת לאחר גירוי TGF-β בתאי NMuMG לעומת תאים לא מטופלים(איור 5C). TGF-β-induced EMT בתאי NMuMG אושרה עוד יותר על ידי כתמי כשל חיסוני של E-cadherin. לאחר גירוי TGF-β במשך יומיים, תאי NMuMG הביעו פחות E-cadherin מאשר תאים בקבוצת הביקורת uninduced, כפי שנותח על ידי מיקרוסקופיה confocal (איור 5D). יתר על כן, תאי NMuMG בנוכחות TGF-β יצרו יותר סיבי מתח אקטין, כפי שמוצג על ידי מיקרוסקופיה קונפוקלית (איור 5E).
SB431542 ו- GW788388 מעכבים איתות TGF-β ו- EMT הנגרמת על ידי TGF-β
SB431542 הוא מעכב תחרותי ATP של תחום קינאז של TβRI, המכונה גם קולטן activin כמו קינאז 5 (ALK5), בעוד GW788388 מעכב פעילות TβRI ו TβRII קינאז. שני המעכבים יכולים לעכב קולטן TGF-β איתות40. לכן, טיפלנו בתאי NMuMG עם ריכוזים שונים של GW788388 בנוכחות TGF-β במשך שעה אחת. כצפוי, GW788388 עיכב זרחון SMAD2 הנגרמת על ידי TGF-β באופן תלוי מינון (איור 6A). בנוסף, הזרחן בתיווך β TGF של SMAD2 נחסם על ידי טיפול SB431542 (איור 6A). Phosphorylated SMAD2/3 יוצר קומפלקס הטרומרי עם SMAD4 ו translocates לתוך הגרעין כדי לווסת את שעתוק של גנים היעד. לכן, חקרנו את הטרנסלוקציה של SMAD2/3 בתאי NMuMG על ידי כתמי כשל חיסוני של SMAD2/3. הנתונים הראו כי הן SB431542 והן GW788388 עיכבו באופן משמעותי את הטרנסלוקציה הגרעינית הנגרמת על ידי TGF β והצטברות של SMAD2/3 בתאי NMuMG (איור 6B). יתר על כן, ההשפעות המעכבות של SB431542 ו- GW788388 נצפו גם ברמות הביטוי mRNA של גנים חשובים של יעד TGF-β המעורבים ב- EMT, כולל PAI-1, SNAIL, E-cadherin ו- Fibronectin (איור 6C). נתונים אלה הראו כי SB431542 ו- GW788388 חסמו איתות TGF-β ו- EMT הנגרמת על ידי TGF-β.
ניתוח של transdifferentiation של תאים מסרטן השד mesenchymal לתוך adipocytes
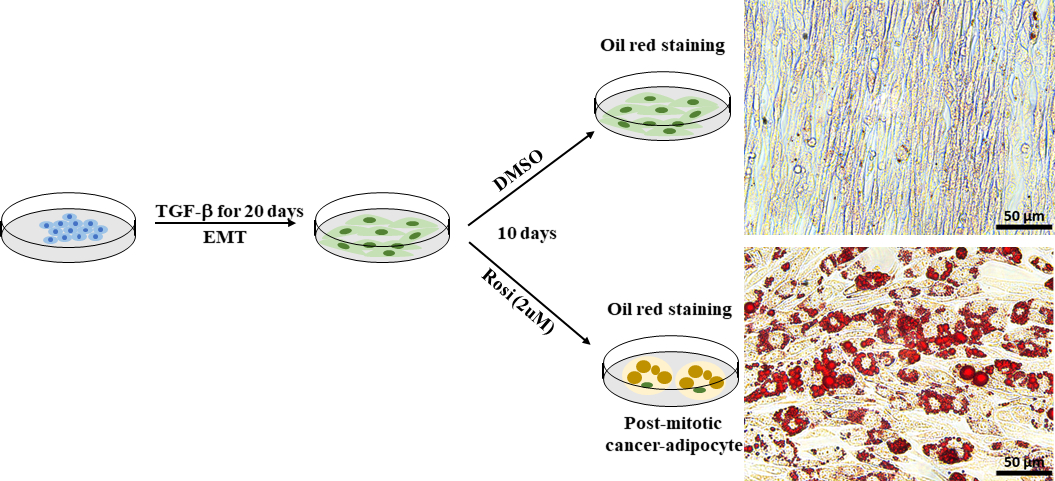
EMT ממלא תפקיד חיוני בשיפור הפלסטיות התאית בסרטן ותוצאות התפתחות עמידות לטיפול. פלסטיות תא סרטן יכול להיות ממוקד ישירות מעוכב עם גישה טרנס בידול, כגון אדיפוגנזה כפויה30. השתמשנו בתאי סרטן השד Py2T מורין, אשר נגזרו בלוטת החלב של גידול החלב של העכבר וירוס פוליומה התיכון גידול אנטיגן (MMTV-PyMT) עכבר מהונדס, כמודל הסלולר של פלסטיות תאים סרטניים הנגרמת על ידי EMT. בהתבסס על פרוטוקול הוקמה41, טיפלנו EMT נגזר Py2T מורין סרטן השד תאים עם התרופה נגד סוכרת rosiglitazone במשך 10 ימים כדי לגרום אדיפוגנזה. Adipogenesis הוערך על ידי הדמיה טיפות השומנים באמצעות שמן אדום O כתמים. תאי שומן זוהו בקלות בתאי סרטן השד Py2T מורין שטופלו ברוזיגליטזון (איור 7), אשר הראו כי טיפול עם rosiglitazone לבד מספיק כדי לקדם את התעתיק של תאים סרטניים בשד נגזר EMT לתוך אדיפוציטים.

איור 1: איתות TGF-β/SMAD. איתות TGF-β יוזם עם מחייב של TGF-β לקולטן מסוג TGF-β II (TβRII), קינאז פעיל מכונן, כי phosphorylates TGF-β סוג אני קולטן (TβRI). לאחר מכן, מופעל TβRI קינאז זרחן SMAD2/3. פפטיד המכיל את מוטיב SSXS של SMAD2 עם שתי שאריות סרין מסוף קרבוקסיה שימש להשגת אנטיסרה פוליקלוני זיהוי זרחן-SMAD2 (p-SMAD2). לכן, ניתוח של ביטוי p-SMAD2 על ידי סופג מערבי יכול לשמש כדי לקבוע את ההפעלה של מסלול איתות TGF-β. Phosphorylated SMAD2/3 יכול ליצור קומפלקסים הטרומריים עם SMAD4, אשר לאחר מכן טרנסלוקציה לתוך הגרעין כדי לווסת תגובות שעתוק. CAGA12-luciferase כתב assay ו PCR בזמן אמת כמותי (qRT-PCR) עבור ביטוי mRNA של TGF-β גנים היעד כגון SMAD7 ו SERPINE1 (קידוד חלבון PAI-1), ניתן להשתמש כדי לנתח TGF-β-induced SMAD3-induced תגובות שעתוק תלויות. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 2: מבנה סכמטי של מזהי R-SMAD (SMAD2 ו-SMAD3). התחומים MH1 (כחול) ו- MH2 (צהוב) נשמרים בין מזהי R-SMAD, אך אזור המקשר (אפור) אינו נשמר. תחום MH1 של SMAD3 מכיל מוטיב מחייב DNA, בעוד ש- SMAD2 אינו יכול לאגד דנ"א ישירות, עקב הכנסה (exon 3) בתחום MH1 שלו. תחום MH2 מתווך אוליגומריזציה SMAD, אינטראקציה עם קולטני TGF-β, מחייב חלבון והוא מעורב ברגולציה שעתוק. SMAD2 ו SMAD3 יכול להיות מופעל על ידי זרחון של מוטיב SSXS (באדום) בטרמיני C שלהם. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 3: EMT הנגרמת על-ידי β TGF. במהלך מעבר אפיתל-mesenchymal הנגרמת על ידי TGF-β (EMT), התאים עוברים אובדן אפיתל ורכישת מאפיינים mesenchymal עם תנועתיות תאים משופרת ויכולת פלישה. האינדוקציה של EMT מובילה לביטוי של סמנים mesenchymal כגון N-cadherin, Zeb2, ו חילזון1/2, כמו גם downregulation של סמני אפיתל כולל E-cadherin, β קטנין, קלודין-1. ההפסד המצטבר או הרווח של מאפייני אפיתל/mesenchymal (E/M) גורם לתא להיכנס למצבי ביניים באופן הפיך. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 4: תגובות איתות β TGF בתאי MCF10A-Ras. (A)תאי MCF10A-Ras טופלו עם או בלי TGF-β (2.5 ng/mL) במשך שעה אחת, ו lysates התא היו immunoblotted עבור SMAD2 זרחן (p-SMAD2), סה"כ SMAD2/3 ו GAPDH (כבקרת טעינה). סמן הגודל מוצג מימין. קון: קבוצת בקרה ללא טיפול TGF-β. (ב)ניתוח של פעילות TGF-β (5 ng/mL) באמצעות פעילות CAGA12-luciferase (LUC) התלויה ב- SMAD3-SMAD4 בתאי MCF10A-Ras. הערכים מנורמלים לפעילות β-גלקטוזידאז (βGal). הנתונים מבוטאים כ ממוצע ± s.d, n = 3. מבחן t של התלמיד, ***P ≤ 0.001 (C) qRT-PCR ניתוח של TGF-β גנים היעד SMAD7 ו SERPINE1 (קידוד חלבון PAI-1) בתאי MCF10A-ראס שטופלו TGF-β (2.5 ng/mL) במשך 6 שעות. GAPDH שימש כפקד פנימי. הנתונים מבוטאים כ ממוצע ± s.d, n = 3. מבחן התלמידים, ***P ≤ 0.001. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 5: EMT הנגרמת על-ידי β TGF בתאי NMuMG. (A) מורפולוגיה של תאי NMuMG שטופלו β TGF (2.5 ng/mL) במשך יום או יומיים. בנוכחות TGF-β, תאי NMuMG transdifferentiated לתוך פנוטיפ mesenchymal. סרגל קנה מידה = 150 מיקרומטר (B) תאי NMuMG טופלו עם או בלי TGF-β (5 ng/ mL) במשך 2 ימים, וסמני EMT נותחו על ידי סופג מערבי. סמן הגודל הוא כפי שצוין מימין. קון: קבוצת בקרה ללא טיפול TGF-β. (C) ניתוח ביטוי גנים של סמני EMT (CDH1 (קידוד חלבון E-cadherin), חילזון ו ZEB2) בתאי NMuMG שטופלו במשך יומיים עם TGF-β (5 ng/mL). GAPDH שימש כפקד פנימי. התוצאות מבוטאות כ ממוצע ± s.d., n = 3. *P ≤ 0.05, **P ≤ 0.01, ***P ≤ 0.001. (D)תאי NMuMG הוכתמו על ידי אימונופלואורסצנטיות כדי לזהות את הביטוי של סמן אפיתל E-cadherin (אדום) לאחר טיפול TGF-β (2.5 ng/mL) במשך 2 ימים. גרעינים היו מוכתמים ב- DAPI (כחול). תמונות נלכדו במיקרוסקופיה קונפוקלית. סרגל קנה מידה = 50 מיקרומטר (E) תאי NMuMG היו מוכתמים בפלואורסצנטין-פאלודין (ירוק) כדי לדמיין F-actin. גרעינים היו מוכתמים ב- DAPI (כחול). סרגל קנה מידה = 50 מיקרומטר.
{kind=link}

איור 6: TGF-βsignaling ו- EMT הנגרמת על ידי TGF-β עכבות על ידי SB431542 ו- GW788388. (A)תאי NMuMG טופלו עם 10 מיקרומטר של SB431542 (SB) או הריכוזים המצוינים של GW788388 (GW) בנוכחות או היעדר TGF-β (5 ng/mL) במשך שעה אחת. lysates התא היו immunoblotted עבור p-SMAD2, SMAD2/3 ו GAPDH. (B)תאי NMuMG טופלו עם 5 מיקרומטר של SB431542 (SB) או 10 מיקרומטר של GW788388 (GW) בנוכחות או היעדר 5 ng/mL של TGF-β במשך שעה אחת מוכתם על ידי immunofluorescence כדי לזהות את ההעתקה הגרעינית של SMAD2/3 (ירוק). תמונות נלכדו במיקרוסקופיה קונפוקלית. (C) ביטוי של גנים היעד TGF-β, כולל PAI-1 וגנים קידוד סמני EMT, כולל SNAIL, E-Cadherin ו Fibronectin, נותחו על ידי תגובת שרשרת פולימראז תעתיק הפוך (RT-PCR) בתאי NMuMG לאחר טיפול SB או GW וגירוי TGF-β במשך 48 שעות. GAPDH שימש כבקרת טעינה. הפקד מציין תאים שאינם מטופלים. נתון זה שונה מ פיטרסן M. ואח' . 34 באישור המפרסם. לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
{kind=link}

איור 7: ניתן לגרום לתאי סרטן השד מורין Py2T שמקורם ב-EMT להבדיל ביניהם לאדיפוציטים. Py2T מורין סרטן השד תאים היו מגורה עם 2 ng/mL TGF-β במשך 20 ימים כדי לגרום EMT להשלים. לאחר מכן, התאים טופלו גם עם DMSO כבקרת רכב או rosiglitazone (2 מיקרומטר) במשך 10 ימים כדי לאפשר את הבידול של תאים סרטניים mesenchymal ולעורר אדיפוגנזה. המדיום הוחלף כל יומיים. לאחר 10 ימים של טיפול, התאים היו מוכתמים בשמן אדום O. Scale bar = 50 מיקרומטר.
{kind=link}
| מינים | שם גנים | קדימה (5' עד 3') | הפוך (5' עד 3') | ||
| אנושי | גאפ-די | TGCACCACCAקטגקטגC | GGקטגקטגקטגאג | ||
| SMAD7 | TCCAגאטגTCC | GTCCגאטגקטג'קג'יק | |||
| סרפין1 | קקאוטגאגג'קקט | קטרג'יג'גטגטגאקאק | |||
| העכבר | גאפ-די | TGGקאגטגאטגאטגג'ק | אאגאטגטגאטגגקטקטקג'ג | ||
| CDH1 | ACCAAAGTCGCTGAAGTC | גאגאטגטקאטגאטג | |||
| חילזון | קגקטגקאקטגט | GCGAGGGCCCCGGAGCA | |||
| ZEB2 | TTCTGCAAGCTCTGTAGCC | TTCTGGCCCCTGCAT | |||
טבלה 1: פריימרים המשמשים עבור qRT-PCR.
Discussion
TGF-β/SMAD איתות ממלא תפקיד מרכזי בהתקדמות סרטן השד, כפי שהוא יכול לקדם את פולשניות התא סרטן השד גרורות על ידי גרימת EMT7. כאן, תיארנו זרימת עבודה הגיונית כדי לחקור איתות ביוזמת TGF-β מהפעלת SMAD הנגרמת על ידי קולטן לתגובות שעתוק וביולוגיות בתיווך SMAD. התחלנו בתיאור הניתוח של זרחון SMAD2, המשכנו עם תגובות שעתוק תלויות SMAD3 הנגרמות על ידי TGF β וביטוי סמן EMT הן ברמת הגן והן ברמת החלבון כדי לנתח את תגובת האיתות TGF-β / SMAD, ולבסוף בדקתי את ה- EMT המושרה על ידי TGF-β. השתמשנו כתב שעתוק CAGA12-luciferase המכיל תיבות CAGA נגזר מקדם PAI-1, כדי לפקח על הפעילות של TGF-β / SMAD איתות מסלול35. מבנה כתב זה דורש SMAD3 ו- SMAD4 להפעלה. מחקרים קודמים הראו כי ההפלה של SMAD4 מוחלש TGF-β-induced CAGA12-luciferase פעילות37. בנוסף לבדיקת הכתב, קביעת מצב הזרחן של SMADs אנדוגני, כולל SMAD2 ו SMAD3, היא דרך נוספת לחקור את תגובת איתות TGF-β. ואכן, חברים אחרים במשפחת TGF-β, כגון גורם צמיחה ובידול (GDF)-8/מיוסטטין ו- GDF-9, גם מתמר אותות באמצעות חלבוני SMAD2/3 על ידי עיסוק TβRI42,43,44. בנוסף לכתב CAGA12-luciferase, מספר כתבים דומים שימשו כדי לזהות את ההפעלה של איתות TGF-β. לדוגמה, כתב שעתוק (SBE) 4 -Lux עם רכיביתגובההנגזרים ממקדם JunB יכול להיגרם ביעילות על ידי TGF-β, activins ו BMPs45.
כתמים מערביים ו qPCR שימשו לניתוח TGF-β-induced EMT, שהן שיטות קלאסיות לחקור את הביטוי של סמני אפיתל (כלומר, E-cadherin) וסמנים mesenchymal (כלומר, N-cadherin, חילזון, שבלול ו Zeb2). ביצענו גם כתמי כשל חיסוני עקיף של E-cadherin וכתם פלואורסצנטי ישיר של F-actin. מבחנים אלה עוד יותר אימת את פנוטיפ mesenchymal של תאים לאחר טיפול TGF-β. המגבלה של כתמי כשל חיסוני היא שיש לתקן תאים לפני הדגירה עם נוגדנים והדמיה, וקשה לחקור שינויים בביטוי סמן EMT בתאים חיים. לאחרונה, העיצוב של קווי תא כתב EMT, כגון A549 ריאות אדנוקרצינומה-vimentin-RFP, אפשר לפקח על הטרנספורמציה של תאי אפיתל לתאים mesenchymal בזמן אמת באמצעות הביטוי של חלבון פלואורסצנטי אדום (RFP)מתויג vimentin. פלטפורמה זו יכולה להיות מנוצלת עבור הקרנת סמים ופיתוח תרופות חדשות46. צבע LifeAct, פפטיד 17-חומצת אמינו, שיכול להכתים מבני F-actin בתאים חיים, הופך לכלי רב ערך כדי לדמיין את ציטוסקלטון actin בזמן אמת מבלי להפריע לתהליכים הסלולר47. במחקר זה, השתמשנו בשני מעכבי מולקולה קטנה, SB431542 ו- GW788388, כדי לאמת את ההשפעה המעכבת שלהם על איתות TGF-β ו- EMT הנגרמת על ידי TGF-β. יש לציין כי GW788388 מעכב בעוצמה את פעילות TβRI ו- TβRII, בעוד של- SB431542 יש השפעה מעכבת רק על TβRI (ו- ALK4 ו- ALK7). מחקרים קודמים גילו כי GW788388 הוא חזק יותר vivo מאשר SB43154240. בנוסף לעיכוב של EMT, GW788388 הפחית את הביטוי של סמני פיברוזיס בכליה, ואת הממשל אוראלי של GW788388 בעכברים סוכרתיים ירד במידה ניכרת גלומרולופתיה25,48.
EMT ממלא תפקיד חיוני בקידום פלסטיות תא סרטן ותוצאות עמידות לתרופות גרורות 49. לכן, מיקוד תאים נגזרי EMT עם תרופות ציטוטוקסיות ספציפיות50 או גרימת חלוקה מחדש באמצעות מעבר mesenchymal-to-אפיתל (MET)51 הוצע כגישה להתגבר על גרורות תאים סרטניים והתנגדות לטיפול. עם זאת, MET תורם לשגשוג של תאים סרטניים מופצים באיברים רחוקים52, אשר עשוי להיות מועיל בעת שימוש בהיפוך הטיפולי של EMT. לאחרונה, מחקר חדש דיווח על גישה טיפולית transdifferentiation על ידי מיקוד ישיר EMT נגזר תאים סרטניים השד עבור בידול לתוך adipocytes30. מחקרו של ישי-רונן ואח' Al.30 השתמשו בתאי סרטן אפיתל מורין Py2T שעברו מעבר לתאים mesenchymal בתגובה לטיפול ארוך טווח עם TGF-β. הם הראו כי rosiglitazone בשילוב עם מעכבי MEK משופרת בידול אפיתל ו adipogenesis. עם זאת, מצאנו כי rosiglitazone לבד היה מספיק כדי לגרום transdifferentiation של תאים מורין Py2T mesenchymal לתוך adipocytes.
לסיכום, השיטות המשמשות במחקר זה סיפקו זרימת עבודה הגיונית כדי לחקור איתות TGF-β ו- EMT הנגרמת על ידי TGF-β. שני המעכבים, SB431542 ו- GW788388, יכולים לחסום תגובות הנגרמות על ידי TGF-β ו- EMT. בנוסף, הדגמנו גם rosiglitazone לבד גורם adipogenesis מסוימים TGF-β-induced תאים סרטן השד mesenchymal. למרות שהשתמשנו רק בכמה קווי תאים סרטניים בשד כדי לחקור תגובות TGF-β, השיטות המתוארות כאן יכול להיות extrapolated לתאים אחרים (סרטן). כאן, השתמשנו בריכוזים שונים של β TGF כדי לגרום לתגובות סלולריות. ברוב סוגי התאים, TGF-β מפעילה את פעילותה הביולוגית בטווח הריכוז של 0.01-10 ng/mL53 וגורם לאיתות בתבנית תגובת מינון. בתאי אנדותל ראשוניים, כולל תאי אנדותל אבי העורקים שור, TGF-β המושרה הביטוי המשמעותי של SMAD2 phosphorylated ב 0.025 ng/mL, הגיע למקסימום ב 0.25 ng/mL, ונשאר ברמה זו בתגובה ריכוזים גבוהים יותר53. במחקר שלנו, השתמשנו בריכוז גבוה של TGF-β (5 ng/mL) בתאי MCF10A-ראס לבדיקת הכתב שעתוק כדי לקבל תגובות חזקות. SMAD2 זרחון וביטוי גנים היעד יכול להיגרם על ידי TGF-β במינון נמוך; לכן, השתמשנו 2.5 ng/mL TGF-β לטיפול בתאים. עם זאת, ריכוז העבודה המתאים ביותר תלוי בסוג התא ובאפקטים המשוערים. כדי לקבוע את הריכוז הטוב ביותר של TGF-β, מומלץ לטפל בתאים במינונים שונים (מנמוך לגבוה).
Disclosures
למחברים אין מה לחשוף.
Acknowledgements
אנו מכירים בתמיכת מועצת המלגות הסינית (CSC) ל- J.Z. ולמרכז הגנומיקה לסרטן בהולנד (CGC). NL) לפ.ט.ד.
Materials
| Name | Company | Catalog Number | Comments |
| Reagent | |||
| 18 mm-side square glass coverslips | Menzel Gläser | 631-1331 | |
| 4′,6-diamidino-2-phenylindole (DAPI) | Vector Laboratories | H-1200 | |
| Alexa Fluor 488 Phalloidin | Thermo Fisher Scientific | A12379 | |
| Alexa Fluor 555 secondary antibody | Thermo Fisher Scientific | A-21422 | |
| Anti-E-cadherin antibody | BD Biosciences | 610181 | |
| anti-glyceraldehyde 3-phosphate dehydrogenase(GAPDH) antibody | Merck Millipore | MAB374 | |
| Anti-N-cadherin antibody | BD Biosciences | 610920 | |
| Anti-Slug antibody | Cell Signaling Technology | 9585 | |
| anti-SMAD2/3 antibody | Becton Dickinson | 610842 | |
| Anti-Snail antibody | Cell Signaling Technology | 3879 | |
| Anti-Tubulin antibody | Cell Signaling Technology | 2148 | |
| Bovine Serum Albumin | Sigma-Aldrich | A2058 | |
| Cholera enterotoxin | Sigma-Aldrich | C8052 | |
| Clarity Western ECL Substrate | Bio-Rad | 1705060 | |
| DC protein assay kit | Bio-Rad | 5000111 | |
| DMEM-high glucose | Thermo Fisher Scientific | 11965092 | |
| DMEM-high glucose medium | Thermo Fisher Scientific | 11965092 | |
| Dulbecco’s modified Eagle’s medium (DMEM)/F12 | Thermo Fisher Scientific | 11039047 | |
| epidermal growth factor (EGF) | Merck Millipore | 01-107 | |
| Fetal bovine serum (FBS) | BioWest | S1860-500 | |
| GoTaq qPCR Master Mix | PROMEGA | A600X | |
| Horse serum | Thermo Fisher Scientific | 26050088 | |
| Hydrocortisone | Sigma-Aldrich | H0135 | |
| Insulin | Sigma-Aldrich | 91077C | |
| Mini Protease Inhibitor Cocktail | Roche | 11836153001 | |
| NucleoSpin RNA II kit | BIOKE´ | 740955 | |
| Penicillin-streptomycin (Pen-Strep) | Thermo Fisher Scientific | 15140148 | |
| Polyethylenimine (PEI) | Polyscience | 23966 | |
| Polyvinylidene difluoride (PVDF) membrane | Merck Millipore | IPVH00010 | |
| Ponceau S solution | Sigma-Aldrich | P7170 | |
| RevertAid First Strand cDNA Synthesis Kit | Thermo Fisher Scientific | K1621 | |
| Rosiglitazone | Sigma-Aldrich | 557366-M | |
| Skimmed milk | Campina: Elk | ||
| Equipment | |||
| ChemiDoc Imaging System | Bio-Rad | 17001402 | |
| CFX Connect Detection System | Bio-Rad | 1855201 | |
| Luminometer | Perkin Elmer | 2030-0050 | |
| NanoDrop 2000/2000c Spectrophotometers | Thermo Fisher Scientific | ND-2000 |
References
- Tzavlaki, K., Moustakas, A. TGF-β Signaling. Biomolecules. 10 (3), (2020).
- Hao, Y., Baker, D., Ten Dijke, P. TGF-β-Mediated Epithelial-Mesenchymal Transition and Cancer Metastasis. International Journal of Molecular Sciences. 20 (11), (2019).
- Morikawa, M., Derynck, R., Miyazono, K. TGF-β and the TGF-β Family: Context-Dependent Roles in Cell and Tissue Physiology. Cold Spring Harbor Perspectives in Biology. 8 (5), (2016).
- Derynck, R., Akhurst, R. J. Differentiation plasticity regulated by TGF-β family proteins in development and disease. Nature Cell Biology. 9 (9), 1000-1004 (2007).
- Massague, J. How cells read TGF-β signals. Nature Reviews Molecular Cell Biology. 1 (3), 169-178 (2000).
- Seoane, J., Gomis, R. R. TGF-β Family Signaling in Tumor Suppression and Cancer Progression. Cold Spring Harbor Perspectives in Biology. 9 (12), (2017).
- Colak, S., Ten Dijke, P. Targeting TGF-β Signaling in Cancer. Trends in Cancer. 3 (1), 56-71 (2017).
- Suriyamurthy, S., Baker, D., ten Dijke, P., Iyengar, P. V. Epigenetic Reprogramming of TGF-β Signaling in Breast Cancer. Cancers (Basel). 11 (5), (2019).
- Drabsch, Y., Ten Dijke, P. TGF-β signalling and its role in cancer progression and metastasis). Cancer and Metastasis Reviews. 31 (3-4), 553-568 (2012).
- Goumans, M. J., Ten Dijke, P. TGF-β Signaling in Control of Cardiovascular Function. Cold Spring Harbor Perspectives in Biology. 10 (2), (2018).
- Robertson, I. B., Rifkin, D. B. Regulation of the Bioavailability of TGF-β and TGF-β-Related Proteins. Cold Spring Harbor Perspectives in Biology. 8 (6), (2016).
- Robertson, I. B., et al. Latent TGF-β-binding proteins. Matrix Biology. 47, 44-53 (2015).
- Khan, Z., Marshall, J. F. The role of integrins in TGF-β activation in the tumour stroma. Cell and Tissue Research. 365 (3), 657-673 (2016).
- Jenkins, G. The role of proteases in transforming growth factor-β activation. The International Journal of Biochemistry & Cell Biology. 40 (6-7), 1068-1078 (2008).
- Tran, D. Q., et al. GARP (LRRC32) is essential for the surface expression of latent TGF-β on platelets and activated FOXP3+ regulatory T cells. Proceedings of the National Academy of Sciences of the United States of America. 106 (32), 13445-13450 (2009).
- Stockis, J., Colau, D., Coulie, P. G., Lucas, S. Membrane protein GARP is a receptor for latent TGF-β on the surface of activated human Treg. European Journal of Immunology. 39 (12), 3315-3322 (2009).
- Wang, R., et al. GARP regulates the bioavailability and activation of TGF-β. Molecular Biology of the Cell. 23 (6), 1129-1139 (2012).
- Vander Ark, A., Cao, J., Li, X. TGF-β receptors: In and beyond TGF-β signaling. Cellular Signalling. 52, 112-120 (2018).
- Heldin, C. H., Miyazono, K., ten Dijke, P. TGF-β signalling from cell membrane to nucleus through SMAD proteins. Nature. 390 (6659), 465-471 (1997).
- ten Dijke, P., Hill, C. S. New insights into TGF-β-Smad signalling. Trends in Biochemical Sciences. 29 (5), 265-273 (2004).
- Miyazono, K. TGF-β signaling by Smad proteins. Cytokine & Growth Factor Reviews. 11 (1-2), 15-22 (2000).
- Yu, Y., Feng, X. H. TGF-β signaling in cell fate control and cancer. Current Opinion in Cell Biology. 61, 56-63 (2019).
- Zhang, Y. E. Non-Smad pathways in TGF-β signaling. Cell Research. 19 (1), 128-139 (2009).
- Katsuno, Y., Lamouille, S., Derynck, R. TGF-β signaling and epithelial-mesenchymal transition in cancer progression. Current Opinion in Oncology. 25 (1), 76-84 (2013).
- Lamouille, S., Xu, J., Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nature Reviews Molecular Cell Biology. 15 (3), 178-196 (2014).
- Yang, J., et al. Guidelines and definitions for research on epithelial-mesenchymal transition. Nature Reviews Molecular Cell Biology. 21 (6), 341-352 (2020).
- Zhang, Y., Weinberg, R. A. Epithelial-to-mesenchymal transition in cancer: complexity and opportunities. Frontiers in Medicine. 12 (4), 361-373 (2018).
- Brabletz, T., Kalluri, R., Nieto, M. A., Weinberg, R. A. EMT in cancer. Nature Reviews Cancer. 18 (2), 128-134 (2018).
- Ocana, O. H., et al. Metastatic colonization requires the repression of the epithelial-mesenchymal transition inducer Prrx1. Cancer Cell. 22 (6), 709-724 (2012).
- Ishay-Ronen, D., et al. Gain Fat-Lose Metastasis: Converting Invasive Breast Cancer Cells into Adipocytes Inhibits Cancer Metastasis. Cancer Cell. 35 (1), 17-32 (2019).
- Smith, C. J., Osborn, A. M. Advantages and limitations of quantitative PCR (Q-PCR)-based approaches in microbial ecology. FEMS Microbiology Ecology. 67 (1), 6-20 (2009).
- Bustin, S. A., Mueller, R. Real-time reverse transcription PCR (qRT-PCR) and its potential use in clinical diagnosis. Clinical Science. 109 (4), 365-379 (2005).
- Sundqvist, A., et al. JUNB governs a feed-forward network of TGF-β signaling that aggravates breast cancer invasion. Nucleic Acids Research. 46 (3), 1180-1195 (2018).
- Persson, U., et al. The L45 loop in type I receptors for TGF-β family members is a critical determinant in specifying Smad isoform activation. FEBS Letters. 434 (1-2), 83-87 (1998).
- Dennler, S., et al. Direct binding of Smad3 and Smad4 to critical TGF-β-inducible elements in the promoter of human plasminogen activator inhibitor-type 1 gene. The EMBO Journal. 17 (11), 3091-3100 (1998).
- Piek, E., Moustakas, A., Kurisaki, A., Heldin, C. H., ten Dijke, P. TGF-β type I receptor/ALK-5 and Smad proteins mediate epithelial to mesenchymal transdifferentiation in NMuMG breast epithelial cells. Journal of Cell Science. 112, Pt 24 4557-4568 (1999).
- Deckers, M., et al. The tumor suppressor Smad4 is required for transforming growth factor-β-induced epithelial to mesenchymal transition and bone metastasis of breast cancer cells. Cancer Research. 66 (4), 2202-2209 (2006).
- Souchelnytskyi, S., et al. Phosphorylation of Ser465 and Ser467 in the C terminus of Smad2 mediates interaction with Smad4 and is required for transforming growth factor-β signaling. Journal of Biological Chemistry. 272 (44), 28107-28115 (1997).
- Abdollah, S., et al. TβRI phosphorylation of Smad2 on Ser465 and Ser467 is required for Smad2-Smad4 complex formation and signaling. Journal of Biological Chemistry. 272 (44), 27678-27685 (1997).
- Petersen, M., et al. Oral administration of GW788388, an inhibitor of TGF-β type I and II receptor kinases, decreases renal fibrosis. Kidney International. 73 (6), 705-715 (2008).
- Gubelmann, C., et al. Identification of the transcription factor ZEB1 as a central component of the adipogenic gene regulatory network. Elife. 3, 03346(2014).
- Nickel, J., Ten Dijke, P., Mueller, T. D. TGF-β family co-receptor function and signaling. Acta Biochimica et Biophysica Sinica. 50 (1), 12-36 (2018).
- Mazerbourg, S., et al. Growth differentiation factor-9 signaling is mediated by the type I receptor, activin receptor-like kinase 5. Molecular Endocrinology. 18 (3), 653-665 (2004).
- Rebbapragada, A., Benchabane, H., Wrana, J. L., Celeste, A. J., Attisano, L. Myostatin signals through a transforming growth factor-β-like signaling pathway to block adipogenesis. Molecular and Cellular Biology. 23 (20), 7230-7242 (2003).
- Jonk, L. J., Itoh, S., Heldin, C. H., ten Dijke, P., Kruijer, W. Identification and functional characterization of a Smad binding element (SBE) in the JunB promoter that acts as a transforming growth factor-β, activin, and bone morphogenetic protein-inducible enhancer. Journal of Biological Chemistry. 273 (33), 21145-21152 (1998).
- Thiery, J. P., Sleeman, J. P. Complex networks orchestrate epithelial-mesenchymal transitions. Nature Reviews Molecular Cell Biology. 7 (2), 131-142 (2006).
- Riedl, J., et al. Lifeact: a versatile marker to visualize F-actin. Nature Methods. 5 (7), 605-607 (2008).
- Gellibert, F., et al. Discovery of 4-{4-[3-(pyridin-2-yl)-1H-pyrazol-4-yl]pyridin-2-yl}-N-(tetrahydro-2H- pyran-4-yl)benzamide (GW788388): a potent, selective, and orally active transforming growth factor-β type I receptor inhibitor. Journal of Medicinal Chemistry. 49 (7), 2210-2221 (2006).
- Ishay-Ronen, D., Christofori, G. Targeting Cancer Cell Metastasis by Converting Cancer Cells into Fat. Cancer Research. 79 (21), 5471-5475 (2019).
- Gupta, P. B., et al. Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell. 138 (4), 645-659 (2009).
- Pattabiraman, D. R., et al. Activation of PKA leads to mesenchymal-to-epithelial transition and loss of tumor-initiating ability. Science. 351 (6277), (2016).
- Gao, D., et al. Myeloid progenitor cells in the premetastatic lung promote metastases by inducing mesenchymal to epithelial transition. Cancer Research. 72 (6), 1384-1394 (2012).
- Goumans, M. J., et al. Balancing the activation state of the endothelium via two distinct TGF-β type I receptors. The EMBO Journal. 21 (7), 1743-1753 (2002).
Erratum
Formal Correction: Erratum: Studying TGF-β Signaling and TGF-β-induced Epithelial-to-mesenchymal Transition in Breast Cancer and Normal Cells
Posted by JoVE Editors on 12/15/2020. Citeable Link.
An erratum was issued for: Studying TGF-β Signaling and TGF-β-induced Epithelial-to-mesenchymal Transition in Breast Cancer and Normal Cells. NMuMG was mislabeled as cancer cells.
NMuMG cells are non-transformed epithelial breast cells established from a mouse mammary gland. NMuMG cells are frequently used as a model system to investigate TGF-β-induced epithelial to mesenchymal transition (EMT). EMT is induced by TGF-β in premalignant human MCF 10A-RAS (M2) cells and mouse MMTV-PyMT breast cancer cell line. The article has been updated to reflect this.
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved