
Method Article
Untersuchung von TGF-β Signalisierung und TGF-β-induzierten epitheliaal-mesenchymalen Übergang bei Brustkrebs und normalen Zellen
In diesem Artikel
Erratum Notice
Zusammenfassung
Wir beschreiben einen systematischen Workflow zur Untersuchung der TGF-β Signalisierung und tGF-β-induzierten EMT, indem wir die Protein- und Genexpression untersuchen, die an diesem Signalweg beteiligt sind. Die Methoden umfassen Western Blotting, ein Luziferase Reporter Assay, qPCR, und Immunfluoreszenz Färbung.
Zusammenfassung
Die Transformation von Wachstumsfaktor-β (TGF-β) ist ein sezernierter multifunktionaler Faktor, der eine Schlüsselrolle in der interzellulären Kommunikation spielt. Störungen der TGF-β-Signalisierung kann zu Brustkrebs führen. TGF-β seine Auswirkungen auf die Proliferation und Differenzierung über spezifische Zelloberflächen-TGF-β Typ-I- und Typ-II-Rezeptoren (d. h. T-RI und T-RII) hervor, die eine intrinsische Serin-/Threoninkinase-Domäne enthalten. Bei TGF-β-induzierter heteromerer komplexer Bildung entlockt die aktivierte T-RI die intrazelluläre Signalisierung durch Phosphorylieren von SMAD2 und SMAD3. Diese aktivierten SMADs bilden heteromere Komplexe mit SMAD4, um bestimmte Zielgene zu regulieren, einschließlich plasminogener Aktivierungshemmer 1 (PAI-1, kodiert durch das SERPINE1-Gen). Die Induktion des epitheliaal-mesenchymalen Übergangs (EMT) ermöglicht Epithelkrebszellen an der primären Stelle oder während der Besiedlung an entfernten Stellen, um einen invasiven Phänotyp zu erhalten und die Tumorprogression voranzutreiben. TGF-β wirkt als potenter Induktor einer Brustkrebsinvasion, indem sie EMT antreibt. Hier beschreiben wir systematische Methoden zur Untersuchung von TGF-β Signalisierung und EMT-Antworten anhand prämaligner menschlicher MCF10A-RAS (M2)-Zellen und Maus-NMuMG-Epithelzellen als Beispiele. Wir beschreiben Methoden zur Bestimmung der TGF-β-induzierten SMAD2-Phosphorylierung durch Western Blotting, SMAD3/SMAD4-abhängige Transkriptionsaktivität mit Luziferase-Reporteraktivität und SERPINE1-Zielgenexpression durch quantitative Echtzeit-Polymerase-Kettenreaktion (qRT-PCR). Darüber hinaus werden Methoden zur Untersuchung von TGF-β-induzierten EMT beschrieben, indem Veränderungen in der Morphologie, epitheliale und mesenchymale Markerexpression, fadenförmige Aktinfärbung und Immunfluoreszenzfärbung von E-Cadherin gemessen werden. Zwei selektive kleine Molekül-TGF-β-Rezeptorkinase-Inhibitoren, GW788388 und SB431542, wurden verwendet, um TGF-β-induzierte SMAD2-Phosphorylierung, Zielgene und Veränderungen der EMT-Markerexpression zu blockieren. Darüber hinaus beschreiben wir die Transdifferenzierung von mesenchymalen Py2T murine epitheliale Tumorzellen in Adipozyten. Methoden zur Untersuchung von TGF-β-induzierte Signalisierung und EMT bei Brustkrebs können zu neuen therapeutischen Ansätzen für Brustkrebs beitragen.
Einleitung
Der Zytokin transformierende Wachstumsfaktor-β (TGF-β) ist der Prototyp einer großen Gruppe strukturell und funktional verwandter regulatorischer Polypeptide, einschließlich TGF-s (d.h. TGF-1, -2 und -3), Knochenmorphogene Proteine (BMPf) und Aktivine1,2. Diese Zytokine spielen alle eine wichtige Rolle bei der embryonalen Entwicklung und bei der Aufrechterhaltung der Gewebe- und Organhomöostase3. Die Fehlregulation von TGF-β kann zu einer Vielzahl von Krankheiten führen, einschließlich Krebs4,5. TGF-β spielt eine komplexe, doppelte Rolle bei der Krebsprogression: In normalen und prämalignen Epithelzellen verhält sich TGF-β als Tumorsuppressor, indem es die Proliferation hemmt und Apoptose induktisiert6,7; Im späten Stadium der Tumorprogression, in dem zytostatische Reaktionen durch die Aktivierung von Onkogenen oder den Verlust von Tumorsuppressorgenen blockiert werden, fungiert TGF-β jedoch als Tumorverstärker, indem es den epitheliaal-mesenchymalen Übergang (EMT) in Krebszellen fördert und dadurch eine Invasion von Krebszellen und Metastasen ermöglicht, die auf Zellen in der Tumormikroumgebung wirken und anitogene undImmunevasionnstimulieren.
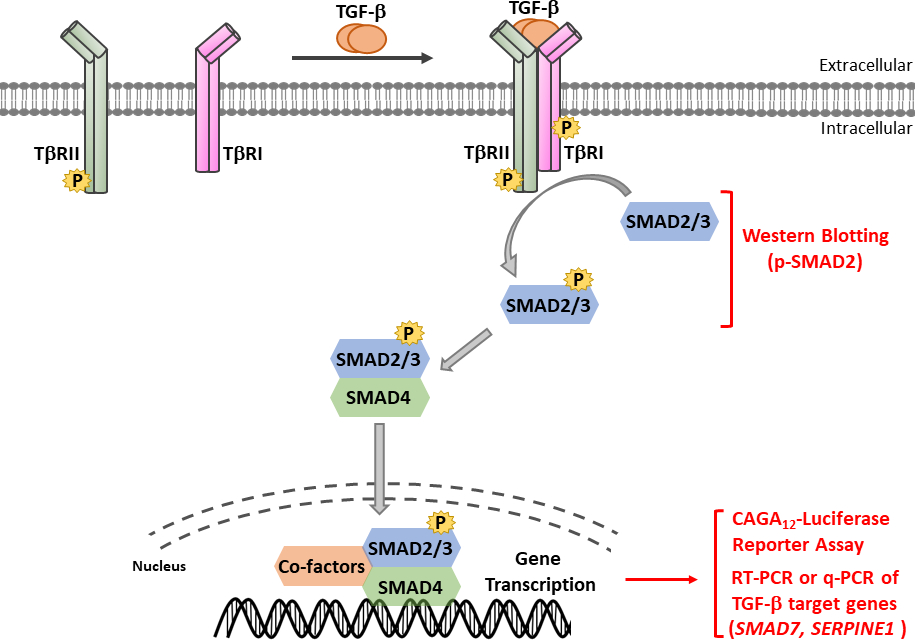
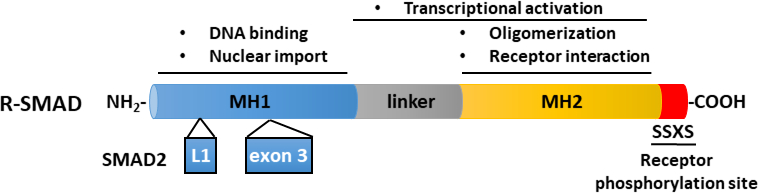
TGF-β wird als inaktives Vorläufermolekül abgesondert, das das ausgereifte Carboxy-Terminal TGF-β und latenzassoziiertes Peptid (LAP)11enthält. Dieser kleine Komplex kann kovalent durch latentes TGF-β-bindendes Protein (LTBP)12gebunden werden. Die Freisetzung von ausgereiften TGF-β kann durch die Wirkung spezifischer Proteasen, die LAP spalten, oder durch das mechanische Ziehen von LAP in einem integrinabhängigen Prozess vermittelt werden13,14. Neben LTBP ist Glycoprotein A Wiederholungen vorherrschend (GARP) stark auf der Oberfläche von regulatorischen T-Zellen (Tregs) exprimiert und spielt eine ähnliche Rolle wie LTBP bei der Regulierung der Aktivierung von TGF-β15,16. GARP bindet direkt an latente TGF-β durch Disulfidverknüpfung und nichtkovalente Assoziation. Die Aktivierung von TGF-β aus dem GARP/TGF-β-Komplex erfordert Integrine17. Reifen TGF-β bindet an TGF-β Serin-/Threoninkinase-Rezeptoren, d.h. TGF-β Typ I (T-RI) und TGF-β Typ-II-Rezeptoren18, um die Signalisierung zu initiieren. Die Bindung von TGF-β an T-RII fördert die Rekrutierung von T-RI und die Bildung eines heteromeren Komplexes. In der Folge wird die T-RI durch die Kinase T-RII auf Serin- und Threoninrückstände in einem kurzen Glycin- und Serin-reichen (GS) Motiv phosphoryliert, was zu seiner Aktivierung19,20führt. Bei der Aktivierung rekrutiert und phosphoryliert die aktivierte T-RI ihre Substrate: die beiden rezeptorspezifischen SMADs (R-SMADs), die SMAD2 und SMAD3 enthalten (Abbildung 1). R-SMADs haben eine ähnliche Gesamtstruktur mit zwei sogenannten Mad Homology-Domänen, MH1 und MH2, die durch eine prolinereiche Linker-Region getrennt sind (Abbildung 2). Das DNA-Bindungsmotiv innerhalb der MH1-Domäne von SMAD3 ist zwischen SMAD2 und SMAD3 nicht konserviert, und SMAD2 kann DNA aufgrund von zwei Einfügungen in seine MH1-Domäne (Exon 3 und L1) nicht direkt binden. SMAD2 und SMAD3 können durch die Phosphorylierung des SSXS-Motivs in ihrem C-termini aktiviert werden (Abbildung 2). Phosphoryliertes SMAD2/3 bildet heteromische Komplexe mit einem gemeinsamen SMAD-Mediator, SMAD4, der sich in den Kern translokalisiert, um die Transkription von Zielgenen zu modulieren (Abbildung 1)7,21. Dieser kanonische SMAD-Signalweg ist präzise reguliert und erzeugt spezifische Zell- und Gewebereaktionen wie die Regulierung des Zellschicksals und tumorzellmetastasierung und Invasion22. Neben der TGF-β-SMAD-Signalisierung können nicht-SMAD-Signalwege auch direkt von Rezeptoren aktiviert werden, um nachgeschaltete zelluläre Reaktionen zu regulieren23.
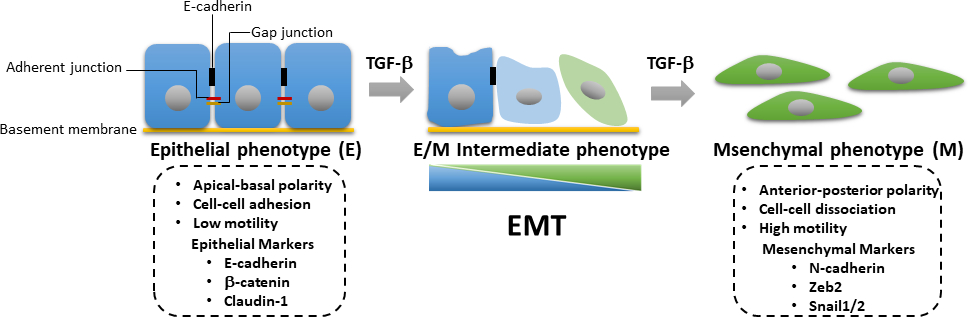
Während der Tumorprogression wird die Aktivierung von TGF-β-induzierten SMAD-abhängigen und SMAD-unabhängigen Bahnen für die Induktion von EMT benötigt. EMT ist ein reversibler Prozess, bei dem Tumorzellen von einem epitheliaalen Phänotyp, der mit dem Verlust von Zell-Zell-Kontakten und verminderter apikal-basaler Polarität verbunden ist, zu einem mesenchymalen Phänotyp mit verbesserter Motilität und Invasionsfähigkeit24dedifferenzieren. EMT ist gekennzeichnet durch eine erhöhte Expression von mesenchymalen Markerproteinen, einschließlich N-Cadherin, Vimentin, Zeb2 und Snail1,2, und die gleichzeitige Downregulation von Epithelmarkern wie E-Cadherin und β-Catenin (Abbildung 3)25. Der Übergang von einem Epithel in einen mesenchymalen Zustand ist jedoch oft unvollständig, und Zellen erhalten gemischte epitheliale und mesenchymale (E/M) Eigenschaften. Ein kürzlich erschienenes Papier der International EMT Association schlug vor, den Prozess von Zellen, die sich zwischen den phänotypischen E/M-Zuständen befinden, als epitheliale-mesenchymale Plastizität (EMP)26zu beschreiben. Diese Plastizität bezieht sich auf partielle EMT, einen hybriden E/M-Status, einen metastabilen EMT-Zustand, ein EMT-Kontinuum und ein EMT-Spektrum26. Während der EMT erhalten Tumorzellen Krebsstammzelleigenschaften (CSC) und werden resistenter gegen ablösende Apoptose27. Während EMT für den Erwerb eines invasiven Phänotyps in primären Tumorzellen verantwortlich ist und die Krebsprogression antreibt, spielt der mesenchymale-epitheliale Übergang (MET) nachweislich eine wichtige Rolle beim Wachstum von disseminierten Tumorzellen an entfernten metastasierenden Stellen28,29. Eine aktuelle Studie hat gezeigt, dass EMT-abgeleitete Brustkrebszellen in Adipozyten transdifferenziert werden können, was eine Möglichkeit bieten könnte, Metastasen zu hemmen und Therapieresistenzen in Tumorzellen und rückfälligem Krebs zu überwinden30. Aufgrund der wichtigen Rolle der TGF-β Signalisierung bei der Aktivierung von EMT bei der Brustkarzinogenese Wir präsentieren detaillierte Protokolle für Western Blotting, einen luziferase-Transkriptionsreporter-Assay, quantitative Echtzeit-Polymerase-Kettenreaktion (qRT-PCR) und Immunfluoreszenz für die Untersuchung der TGF-β-Signalisierung, TGF-β-induzierte EMT und die Transdifferenzierung von EMT-abgeleiteten murinen Brustepitheltumorzellen in Adipozyten. Diese Techniken sind die am häufigsten verwendeten Analysewerkzeuge im Bereich der Zellbiologie. qRT-PCR wird verwendet, um mRNA-Expressionsniveaus quantitativ zu erkennen, zu charakterisieren und zu quantifizieren. Im Vergleich zur quantitativen PCR (qPCR), einer alternativen Technik, kann die Reverse Transkription (RT)-PCR verwendet werden, um die mRNA-Expression semiquantitativ zu bestimmen31,32. Western Blotting wird verwendet, um bestimmte Proteinspiegel in einer bestimmten Zelllysatprobe mit Vorteilen der Empfindlichkeit und Spezifität semiquantitative nativ zu untersuchen. So präsentieren wir einen systematischen Workflow zur Analyse von Veränderungen von der Genexpression zur Proteinexpression, um die TGF-β-Signalisierung zu untersuchen, die auch auf andere Signalwege angewendet werden kann.
Protokoll
1. Analyse der TGF-β-induzierten SMAD2-Phosphorylierung mit Western Blotting
HINWEIS: Prämaligne menschliche Brust MCF10A-Ras-Zellen wurden als Beispiel verwendet, um TGF-β Signalreaktionen33zu untersuchen. Grundsätzlich gelten die nachstehend beschriebenen Methoden auch für andere TGF-β-responsive Zelllinien.
- Kultur der Brustepithelzelllinie MCF10A-Ras bei 37 °C in Dulbeccos modifiziertem Eagle-Medium (DMEM)/F12 mit L-Glutamin mit 5% Pferdeserum, 20 ng/ml epidermaler Wachstumsfaktor (EGF), 10 mg/ml Insulin, 100 ng/ml Cholera-Enterotoxin, 0,5 mg/ml Hydrocortison und 1:100 Penicillin-Streptomycin (Pen-Strep).
- Trypsinize MCF10A-Ras Zellen mit 1 ml 0,25% Trypsin-EDTA für 1 Minute und zählen lebensfähige Zellen mit einem Zellzähler.
- Samenzellen in 6-Well-Platten mit einer Dichte von 5×105 Zellen/gut.
- Nach dem über Nacht Wachstum, behandeln Sie Zellen mit entweder TGF-β (5 ng/ml) oder Ligandenpuffer (4 mM HCl, 0,1% fettsäurefreies Rinderserumalbumin (BSA)) für 1 Stunde, und entfernen Sie dann Kulturmedium und waschen Sie die Zellen vorsichtig zweimal mit 1 ml phosphatgepufferter Kochsaline (PBS).
- Kühlen Sie Zellen in 6-Well-Platten auf Eis und fügen Sie 150 l vorgekühlten Radioimmun-Fällungs-Assay (RIPA) Lysepuffer (150 mM NaCl, 0,1% Triton X-100, 0,5% Natriumdesoxycholat, 0,1% SDS, 50 mM Tris-HCl pH 8.0 und frisch zugesetztem Mini-Protease-Hemmer-Cocktail) hinzu. Lassen Sie die Lyse 10 Minuten auf Demeis weitergehen.
- Mit einem Kunststoffzellenschaber anhaftende Zellen von der Schale abkratzen und die Zellsuspension vorsichtig in ein vorgekühltes Mikrozentrifugenrohr übertragen.
- Zentrifugieren Sie die Zelle 10 Minuten lang bei 150 Zentrifugalkraft (x g) bei 4 °C und übertragen Sie den Überstand in ein frisches 1,5-ml-Mikrozentrifugenrohr.
- Messen Sie die Proteinkonzentration mit einem Waschmittel-kompatiblen (DC) Protein-Assay-Kit.
- 30 g Protein aus jeder Probe auf ein 10% Natriumdodecylsulfat-Polyacrylamid-Gel-Elektrophorese (SDS-PAGE)-Gel geben und das Gel 1,5-2 Stunden lang mit einer Spannung von 100 V laufen lassen.
- Übertragen Sie die Proteine aus dem Gel auf eine 45-m Polyvinylidendifluorid -Membran (PVDF) mit einer Spannung von 110 V für 1-1,5 Stunden.
- Übertragen Sie die PVDF-Membran in einen geeigneten Behälter mit der Proteinseite (der Seite, die dem Gel zugewandt war) nach oben, und spülen Sie die Membran kurz in destilliertem Wasser ab.
- Entsorgen Sie das Wasser, fügen Sie ponceau S Lösung, und legen Sie die Membran auf eine Schaukelplattform für 1-2 Minuten.
- Entflecken Sie die Membran mit destilliertem Wasser, indem Sie sie schnell abspülen und dann 1 Minute waschen.
- Legen Sie dann die PVDF-Membran auf einen Leuchtkasten, und machen Sie ein Bild, um die gleiche Gesamt-Proteinbelastung zu überprüfen.
- Waschen Sie die Membran mit Tris-gepufferter Salin mit Tween 20 (TBST, 20 mM Tris-HCl, pH 7,4, 150 mM NaCl und 0,1% Tween 20), bis keine sichtbare Färbung vorhanden ist.
- Setzen Sie die Membran in einen Sperrpuffer (5% Magermilch in TBST-Lösung) und inkubieren Sie sie 1 Stunde bei Raumtemperatur.
- Waschen Sie die Membran zweimal mit TBST.
- Inkubieren Sie die Membran mit Primärantikörpern gegen Phospho-SMAD2 (p-SMAD2; 1:1000, hausgemachte 34), insgesamt SMAD2/3 1:1000 und Glyceraldehyd 3-Phosphat-Dehydrogenase (GAPDH; 1:1000), über Nacht bei 4 °C.
- Waschen Sie die Membran zweimal mit TBST und inkubieren Sie die Membran mit einem sekundären Antikörper gegen Kaninchen oder Maus (1:5000) für 2 Stunden.
- Inkubieren Sie die PVDF-Membran mit westlichem ECL-Substrat für 30 Sekunden und erkennen Sie das Signal mit einem Bildgebungssystem.
- Wiederholen Sie die Experimente mindestens dreimal, um biologische Triplicate zu erhalten.
2. Analyse von TGF-β-induzierten SMAD3-abhängigen Transkriptionsreaktionen
- Führen Sie den SMAD3/SMAD4-abhängigen CAGA12-luciferase Transkriptionsreporter-Assay durch.
- Kultur und Trypsinize MCF10A-Ras-Zellen, wie in Schritt 1 beschrieben. Samenzellen in 24-Well-Platten bei 5×104 Zellen/Well und lassen die Zellen über Nacht haften.
- Am Tag nach der Aussaat kotranstranssektieren Sie die Zellen in jedem Brunnen mit 100 ng des TGF-β/SMAD3-induziden (CAGA)12 luziferase Transkriptionsreporter konstrukt35 und 80 ng des β-Galactosidase-Expressionskonstrukts mit Polyethylenimin (PEI). Die Transfektion von β-Galactosidase wird verwendet, um Unterschiede in der Transfektionseffizienz zwischen verschiedenen Brunnen zu normalisieren. Richten Sie jede Experimentelle Gruppe in dreifacher Ausfertigung ein.
- Nach 24 Stunden Inkubation, verhungernde Zellen mit serumfreiem DMEM-hoher Glukose und 6 Stunden später TGF-β (5 ng/ml) oder Ligandenpuffer (4 mM HCl, 0,1% BSA) als Fahrzeugsteuerung zu den Zellen hinzufügen.
- Nach weiteren 24 Stunden Inkubation, waschen Zellen zweimal mit vorgewärmten PBS.
- 120 L/well 1× Lysepuffer hinzufügen und die Platte bei 4 °C 20 Minuten lang sanft schütteln.
- Übertragen Sie 30 l Lysat auf jeden Brunnen einer 96-Well-weißen Assay-Mikroplatte, um die Luziferase-Aktivität mit einem Luminometer zu messen.
- Übertragen Sie 50 l Lysat auf jeden Brunnen einer 96-gut transparenten Platte, um β-Galactosidase-Aktivität zu messen.
- Normalisieren Sie die Luziferase-Aktivität auf die β-Galactosidase-Aktivität und wiederholen Sie die Experimente mindestens dreimal, um biologische Triplizen zu erhalten.
- Analysieren Sie die Expression von TGF-β Zielgene mit quantitativer Echtzeit-Polymerase-Kettenreaktion (qRT-PCR).
- Kultur und Trypsinize MCF10A-Ras-Zellen, wie in Schritt 1 beschrieben. Samenzellen in 6-Well-Platten bei 5×105 Zellen/Well und lassen die Zellen über Nacht haften.
- Behandeln Sie Zellen mit TGF-β (5 ng/ml) oder Ligandenpuffer (4 mM HCl, 0,1% BSA) für 6 Stunden, und waschen Sie die Zellen dann zweimal mit 1 ml PBS.
- Isolieren Sie die gesamte RNA mit einem RNA-Isolationskit.
- Bestimmen Sie die RNA-Konzentration mit einem NanoDrop und führen Sie die cDNA-Synthese mit 1 g RNA mit einem ersten Strang-cDNA-Synthese-Kit durch.
- Verwenden Sie ein Echtzeit-PCR-Erkennungssystem, um quantitative Echtzeit-PCR (qRT-PCR) mit zehnfach verdünnter cDNA in einem 10-L-Reaktionsgemisch durchzuführen, das spezifische menschliche Vorwärts- und Reverse-Primer für GAPDH (für die Normalisierung), SERPINE1 (Kodierung des PAI-1-Proteins, ein TGF-β/SMAD-Zielgen), SMAD7 (TGF-β/SMAD-Zielgen) und qPCR-Zielgen enthält. Richten Sie jede Experimentelle Gruppe in dreifacher Ausfertigung ein.
HINWEIS: Die Primersequenzen, die zum Nachweis menschlicher Zielgene in qRT-PCR verwendet werden, sind in Tabelle 1aufgeführt. - Verwenden Sie die folgenden qPCR-Reaktionsbedingungen: Initialisierung, 95 °C für 3 Minuten; Denaturierung, 95 °C für 10 Sekunden; Glühende, 60 °C für 30 Sekunden; und Verlängerung, 80 °C für 10 Sekunden; Denaturierung, Glühen und Verlängerung werden 40 Mal wiederholt.
- Wiederholen Sie die Experimente mindestens dreimal, um biologische Triplicate zu erhalten.
3. Analyse der TGF-β-induzierten EMT
- Analysieren Sie die Expression von EMT-Markern auf Proteinebene mit Western Blotting.
HINWEIS: Die Analyse von TGF-β-induzierten EMT wird anhand von Mund-NMuMG-Epithelzellen als Beispiel36,37gezeigt.- Kultur NMuMG-Zellen bei 37 °C in DMEM-hohem Glukosemedium, ergänzt mit 10% fetalem Rinderserum und 1:100 Pen-Strep. Verwenden Sie die in Schritt 1 beschriebenen Methoden zur Proteinisolierung und -detektion.
HINWEIS: In diesem Experiment werden folgende Antikörper verwendet: E-Cadherin, 1:1000; N-Cadherin, 1:1000; Schnecke, 1:1000; Schnecke, 1:1000; Tubulin, 1:1000 (Bild 3).
- Kultur NMuMG-Zellen bei 37 °C in DMEM-hohem Glukosemedium, ergänzt mit 10% fetalem Rinderserum und 1:100 Pen-Strep. Verwenden Sie die in Schritt 1 beschriebenen Methoden zur Proteinisolierung und -detektion.
- Analysieren Sie die Expression von EMT-Markern auf mRNA-Ebene mit quantitativer Echtzeit-PCR, wie in Schritt 2.2 beschrieben.
HINWEIS: Alle für qRT-PCR verwendeten Mausprimer, einschließlich CDH1 (Kodierung des E-Cadherin-Proteins), SNAILund ZEB2 (Abbildung 3), sind in Tabelle 1aufgeführt. - Analysieren Sie den EMT-Prozess mit indirekter Immunfluoreszenz und direkter Fluoreszenzfärbung.
- Indirekte Immunfluoreszenzfärbung von E-Cadherin
- Legen Sie sterile 18 mm-seitige quadratische Glasabdeckungen in 6-Well-Platten (ein Abdeckungsslip pro Brunnen).
- Samen 1×105 NMuMG-Zellen mit 2 ml komplettem DMEM pro 6-Well-Platte und lassen die Zellen über Nacht haften.
- Bewegen Sie die Abdeckungen mit anhaftenden Zellen vorsichtig auf eine neue 6-Well-Platte und fügen Sie 2 ml Kulturmedium zu den Brunnen hinzu.
- Behandeln Sie die Zellen mit TGF-β (5 ng/ml) oder Ligandenpuffer (4 mM HCl, 0,1% BSA) für 2 Tage.
- Entfernen Sie das Kulturmedium und waschen Sie die Zellen vorsichtig zweimal mit 1 ml vorgewärmten PBS.
- Fixieren Sie die Zellen, indem Sie 1 ml 4% Paraformaldehyd hinzufügen und 30 Minuten bei Raumtemperatur inkubieren. Dann die Zellen vorsichtig zweimal mit 1 ml PBS waschen.
- Permeabilisieren Sie die festen Zellen mit 0,1% Triton X-100 für 10 Minuten bei Raumtemperatur und waschen Sie die Zellen zweimal mit PBS.
- Blockieren Sie die Zellen mit 5% BSA in PBS für 1 Stunde bei Raumtemperatur und waschen Sie die Zellen zweimal mit PBS.
- Fügen Sie den primären Antikörper gegen E-Cadherin (verdünnt 1:1000 in PBS) an die Oberseite jedes Deckelschlupfes und inkubieren für 1 Stunde bei Raumtemperatur.
- Entfernen Sie den primären Antikörper und waschen Sie den Deckelrutsch dreimal mit PBS.
- Fügen Sie den Sekundärantikörper Alexa Fluor 555 (verdünnt 1:500 in PBS) an die Oberseite jedes Coverslips und inkubieren Sie für 1 Stunde bei Raumtemperatur, während Sie mit Aluminiumfolie abdecken, um vor Licht zu schützen.
- Entfernen Sie den sekundären Antikörper und waschen Sie den Deckelrutsch dreimal mit PBS.
- Montieren Sie den Deckelschlupf (Zellen nach unten) auf Glasschlitten mit Montagemedium mit 4,6-Diamidino-2-Phenylindole (DAPI) und lagern Sie die montierten Dias in einer Box bei 4 °C, geschützt vor Licht.
- Beobachten Sie die Färbung mit der konfokalen SP8-Mikroskopie.
- Direkte Fluoreszenzfärbung von fadenförmigem (F)-Aktin.
- Bereiten Sie die folgenden Schritte 3.3.1.1 vor. bis 3.3.1.9.
- Färben Sie die Zellen, indem Sie Alexa Fluor 488 Phalloidin (1:1000) für 1 Stunde bei Raumtemperatur im Dunkeln hinzufügen, um fadenförmiges Actin (F-Actin) zu visualisieren.
- Waschen Sie Zellen dreimal mit PBS.
- Montieren Sie den Deckelrutsch mit DAPI auf Glasschlitten und nehmen Sie Bilder mit der konfokalen SP8-Mikroskopie auf.
- Indirekte Immunfluoreszenzfärbung von E-Cadherin
Ergebnisse
Analyse der TGF-β Signalisierung
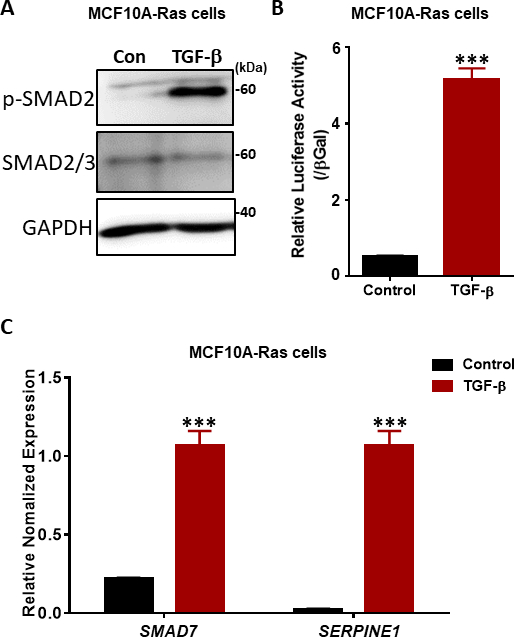
Der schlüsselfertige Schritt bei der TGF-β-Signalisierung ist die Phosphorylierung der beiden carboxy Terminal Serinrückstände im SSXS-Motiv (Abbildung 2) von T-RI Kinase38,39. Um TGF-β Signalreaktionen zu untersuchen, führten wir western blotting von phosphoryliertem SMAD2 durch. In den prämalignen MCF10A-Ras-Zellen der menschlichen Brust erhöhte sich die Phosphorylierung von SMAD2 als Reaktion auf die TGF-β Stimulation für 1 Stunde signifikant, während die Expression von SMAD2/3 insgesamt nicht durch TGF-β Behandlung beeinflusst wurde (Abbildung 4A). Durch die Verwendung des TGF-β-induzierten SMAD3/4-gesteuerten CAGA-luc-Transkriptions-Reporter-Assays fanden wir heraus, dass TGF-β den Luziferase-Reporter in der MCF10A-Ras-Zellenlinie im Vergleich zu nicht behandelten Zellen deutlich induzierte (Abbildung 4B). Darüber hinaus beobachteten wir, dass gut charakterisierte direkte transkriptionsbezogene Genziele von TGF-β einschließlich SMAD7 und SERPINE1 (Kodierung des PAI-1-Proteins) in TGF-β-behandelten MCF10A-Ras-Zellen hochgradig exprimiert wurden (Abbildung 4C).
Analyse von TGF-β-induzierter EMT
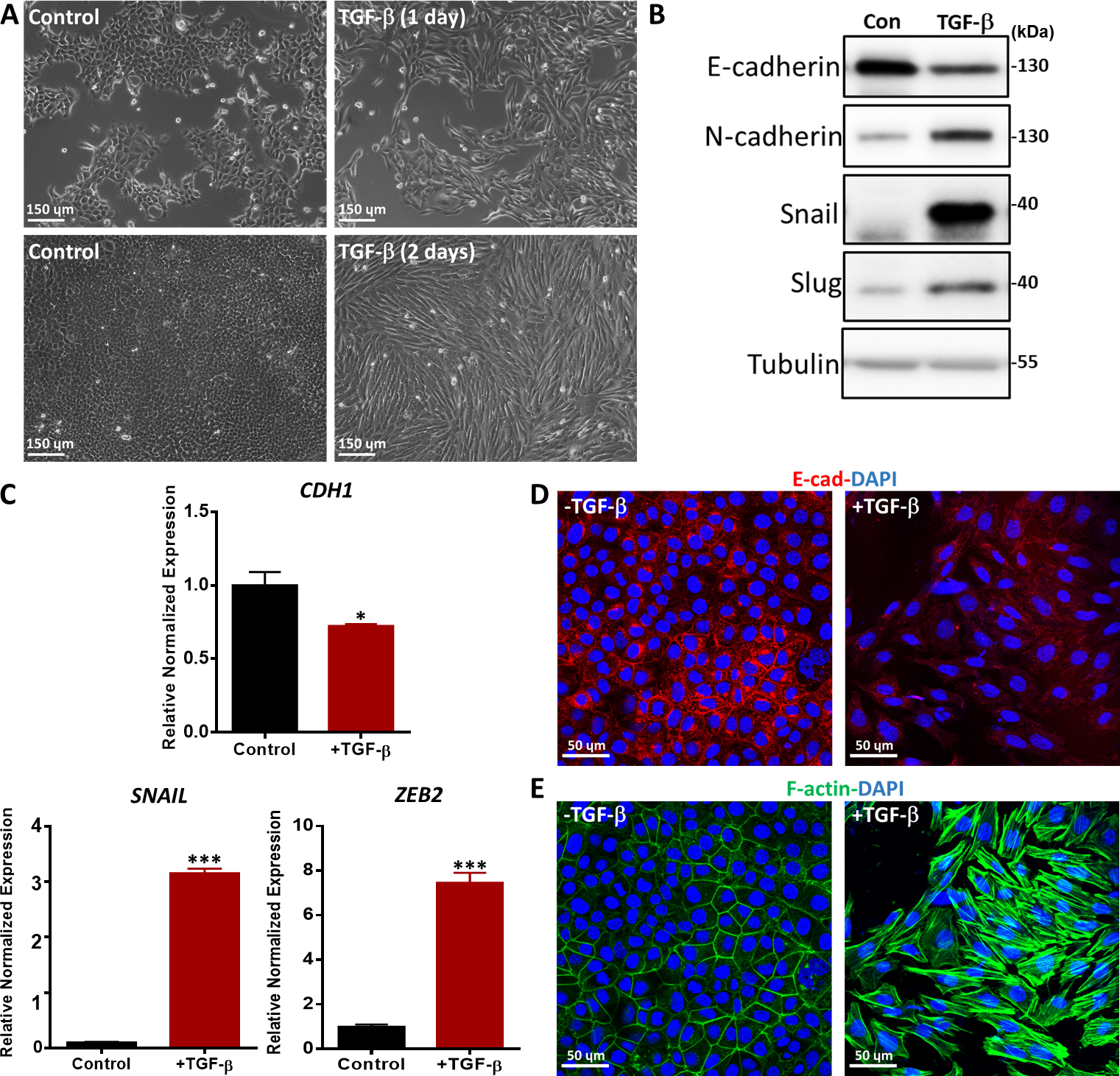
Wir bewerteten TGF-β-induzierte EMT mit verschiedenen Methoden, wie morphologischen Veränderungen, der Expression von EMT-Markern auf mRNA- und Proteinspiegel und der Immunfluoreszenzfärbung von EMT-Markern36. NMuMG-Epithelzellen, die 1 und 2 Tage lang mit TGF-β behandelt wurden, wechselten von einer klassischen epitheliaalen Morphologie zu einer spindelförmigen mesenchymalen Morphologie, wie die Phasenkontrastmikroskopie zeigt (Abbildung 5A). In Übereinstimmung mit den morphologischen Veränderungen beobachteten wir, dass die TGF-β Behandlung zu einer Erhöhung der Proteinexpression von mesenchymalen Markern führte, einschließlich N-Cadherin, Schnecke und Schnecke37 (Abbildung 5B). Im Gegensatz dazu wurde E-Cadherin, ein Epithelmarker, in NMuMG-Zellen nach 2 Tagen TGF-β Behandlung nach unten reguliert (Abbildung 5B). Darüber hinaus führten wir eine quantitative Echtzeit-Polymerase-Kettenreaktion (qRT-PCR) durch, um die Genexpression von EMT-Markern zu untersuchen. CDH1 (Kodierung des E-Cadherin-Proteins) wurde signifikant verringert, während mesenchymale Marker wie SNAIL und Zinkfinger E-Box-bindung Stoltoso-Lösung 2 (ZEB2) nach TGF-β Stimulation in NMuMG-Zellen im Vergleich zu unbehandelten Zellen deutlich erhöht wurden(Abbildung 5C). TGF-β-induzierte EMT in NMuMG-Zellen wurde durch Immunfluoreszenzfärbung von E-Cadherin weiter bestätigt. Bei TGF-β Stimulation für 2 Tage exprimierten NMuMG-Zellen weniger E-Cadherin als Zellen in der nicht induzierten Kontrollgruppe, wie durch konfokale Mikroskopie analysiert (Abbildung 5D). Darüber hinaus bildeten NMuMG-Zellen in Gegenwart von TGF-β mehr Actin-Stressfasern, wie die konfokale Mikroskopie zeigt (Abbildung 5E).
SB431542 und GW788388 hemmen TGF-β Signalisierung und TGF-β-induzierte EMT
SB431542 ist ein ATP-Wettbewerbshemmer der Kinase-Domäne von T-RI, auch als Aktivinrezeptor-ähnliche Kinase 5 (ALK5) bezeichnet, während GW788388 die Kinase-Aktivität von T-RI und T-RII hemmt. Beide Inhibitoren können die TGF-β-Rezeptor-Signalisierung40hemmen. So behandelten wir NMuMG-Zellen mit unterschiedlichen Konzentrationen von GW788388 in Gegenwart von TGF-β 1 Stunde lang. Wie erwartet hemmte GW788388 die TGF-β-induzierte SMAD2-Phosphorylierung dosisabhängig (Abbildung 6A). Zusätzlich wurde die TGF-β-vermittelte Phosphorylierung von SMAD2 durch SB431542-Behandlung blockiert (Abbildung 6A). Phosphoryliertes SMAD2/3 bildet mit SMAD4 einen heteromeren Komplex und translokalisiert sich in den Kern, um die Transkription von Zielgenen zu modulieren. Daher untersuchten wir die Translokation von SMAD2/3 in NMuMG-Zellen durch Immunfluoreszenzfärbung von SMAD2/3. Die Daten zeigten, dass sowohl SB431542 als auch GW788388 die TGF-β-induzierte Kerntranslokation und Akkumulation von SMAD2/3 in NMuMG-Zellen signifikant hemmten (Abbildung 6B). Darüber hinaus wurden die hemmenden Wirkungen von SB431542 und GW788388 auch in den mRNA-Expressionsniveaus wichtiger TGF-β Ansinnen-Gene beobachtet, die an EMT beteiligt sind, einschließlich PAI-1, SNAIL, E-Cadherin und Fibronectin (Abbildung 6C). Diese Daten legten nahe, dass SB431542 und GW788388 tGF-β-Signalisierung und TGF-β-induzierte EMT blockierten.
Analyse der Transdifferenzierung von mesenchymalen Brustkrebszellen in Adipozyten
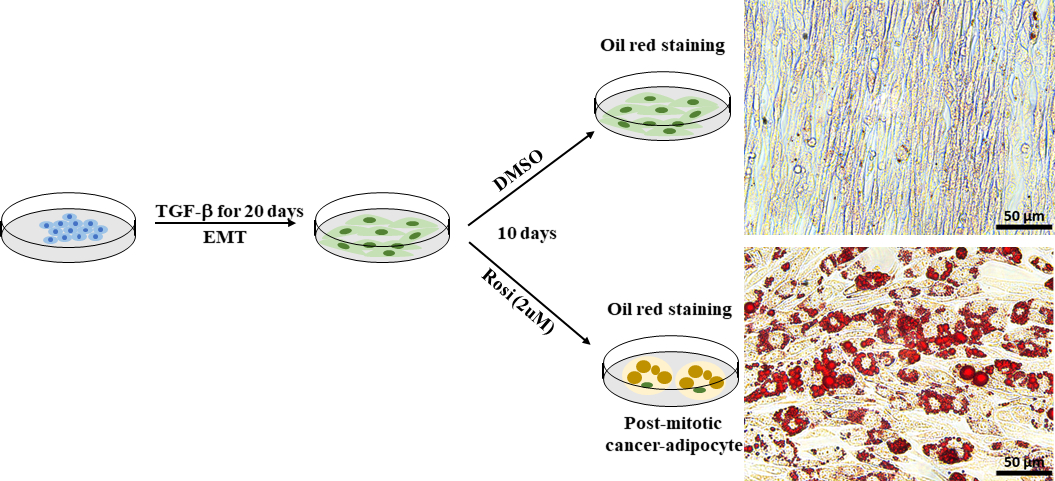
EMT spielt eine wichtige Rolle bei der Verbesserung der zellulären Plastizität bei Krebserkrankungen und führt zur Entwicklung von Therapieresistenzen. Krebszellplastizität kann direkt gezielt und gehemmt werden mit einem Transdifferenzierungsansatz, wie z.B. erzwungene Adipogenese30. Wir verwendeten Py2T-Murine-Brustkrebszellen, die aus der Brustdrüse eines Maus-Mammary-Tumorvirus-Polyom-Mitteltumor-Antigens (MMTV-PyMT) transgene Maus abgeleitet wurden, als zelluläres Modell der EMT-induzierten Krebszellplastizität. Basierend auf einem etablierten Protokoll41behandelten wir EMT-abgeleitete Py2T-murine Brustkrebszellen 10 Tage lang mit dem Antidiabetika um das Antidiabetika Rosiglitazon, um eine Adipogenese zu induzieren. Die Adipogenese wurde durch Visualisierung von Lipidtröpfchen mit ölroter O-Färbung bewertet. Fettzellen wurden leicht in rosiglitazonbehandelten Py2T-Murine-Brustkrebszellen nachgewiesen (Abbildung 7), die zeigten, dass die Behandlung mit Rosiglitazon allein ausreicht, um die Transdifferenzierung von EMT-abgeleiteten Brustkrebszellen in Adipozyten zu fördern.

Abbildung 1: TGF-β/SMAD-Signalisierung. TGF-β Signalisierung initiiert mit der Bindung von TGF-β an TGF-β Typ-II-Rezeptor (T-RII), eine konstitutiv aktive Kinase, die TGF-β Typ-I-Rezeptor (T-RI) phosphoryliert. Dann wird die aktivierte T-RI-Kinase sMAD2/3 aktiviert. Ein Peptid, das das SSXS-Motiv von SMAD2 mit zwei carboxy terminal engetylierten Serinrückständen enthält, wurde verwendet, um polyklonale Antisera zu erhalten, die Phosphor-SMAD2 (p-SMAD2) erkennen. Daher kann die Analyse der p-SMAD2-Expression durch Western Blotting verwendet werden, um die Aktivierung des TGF-β Signalwegs zu bestimmen. Phosphorylierte SMAD2/3 können mit SMAD4 heteromere Komplexe bilden, die sich dann in den Kern transortieren, um Transkriptionsreaktionen zu modulieren. Der CAGA12-luciferase Reporter-Assay und quantitative Echtzeit-PCR (qRT-PCR) für die mRNA-Expression von TGF-β Zielgene wie SMAD7 und SERPINE1 (Codierung PAI-1-Protein) können verwendet werden, um TGF-β-induzierte SMAD3-abhängige Transkriptionsreaktionen zu analysieren. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 2: Schematische Struktur von R-SMADs (SMAD2 und SMAD3). Die Domänen MH1 (blau) und MH2 (gelb) werden zwischen R-SMADs konserviert, der Linkerbereich (grau) jedoch nicht konserviert. Die MH1-Domäne von SMAD3 beherbergt ein DNA-bindendes Motiv, während SMAD2 DNA aufgrund einer Einfügung (exon 3) in seine MH1-Domäne nicht direkt binden kann. Die MH2-Domäne vermittelt Die SMAD-Oligomerisierung, Interaktion mit TGF-β-Rezeptoren und Proteinbindung und ist an der Transkriptionsregulation beteiligt. SMAD2 und SMAD3 können durch die Phosphorylierung des SSXS-Motivs (in rot) in ihrem C-Termini aktiviert werden. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 3: TGF-β-induzierte EMT. Während des TGF-β-induzierten epitheliaal-mesenchymalen Übergangs (EMT) erleiden die Zellen den Verlust von Epithel und den Erwerb von mesenchymalen Eigenschaften mit verbesserter Zellmotilität und Invasionsfähigkeit. Die Induktion von EMT führt zur Expression von mesenchymalen Markern wie N-Cadherin, Zeb2 und Snail1/2 sowie zur Downregulation von Epithelmarkern wie E-Cadherin, β-Catenin und Claudin-1. Der akkumulierte Verlust oder Gewinn epitheliale/mesenchymaler (E/M) Eigenschaften bewirkt, dass eine Zelle in reversible Weise in Zwischenzustände eindringt. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 4: TGF-β Signalantworten in MCF10A-Ras-Zellen. (A) MCF10A-Ras-Zellen wurden entweder mit oder ohne TGF-β (2,5 ng/ml) für 1 Stunde behandelt, und Zelllysate wurden für phosphorylierte SMAD2 (p-SMAD2), sMAD2/3 insgesamt und GAPDH (als Ladekontrolle) immunoblotted. Die Größenmarkierung wird auf der rechten Seite angezeigt. Con: Kontrollgruppe ohne TGF-β Behandlung. (B) Analyse der TGF-β (5 ng/ml) Aktivität mit dem SMAD3–SMAD4-abhängigen CAGA12-luciferase (LUC) Transkriptionsreporter in MCF10A-Ras-Zellen. Die Werte werden auf β-Galactosidase-Aktivität normalisiert. Die Daten werden als Mittelwert ± s.d, n = 3 ausgedrückt. Schüler-t-Test, ***P ≤ 0.001 (C) qRT-PCR-Analyse der TGF-β Zielgene SMAD7 und SERPINE1 (Kodierung des PAI-1-Proteins) in MCF10A-Ras-Zellen, die 6 Stunden lang mit TGF-β (2,5 ng/ml) behandelt wurden. GAPDH wurde als interne Kontrolle verwendet. Die Daten werden als Mittelwert ± s.d, n = 3 ausgedrückt. Schüler-t-Test, ***P ≤ 0,001. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 5: TGF-β-induzierte EMT in NMuMG-Zellen. (A) Morphologie von NMuMG-Zellen, die 1 oder 2 Tage lang mit TGF-β (2,5 ng/ml) behandelt wurden. In Gegenwart von TGF-β, NMuMG-Zellen in einen mesenchymalen Phänotyp transdifferenziert. Scale bar = 150 m (B) NMuMG-Zellen wurden 2 Tage lang mit oder ohne TGF-β (5 ng/ml) behandelt, und EMT-Marker wurden durch Western Blotting analysiert. Die Größenmarkierung ist wie auf der rechten Seite angegeben. Con: Kontrollgruppe ohne TGF-β Behandlung. (C) Genexpressionsanalyse von EMT-Markern (CDH1 (Kodierung des E-Cadherin-Proteins),SNAIL und ZEB2) in NMuMG-Zellen, die 2 Tage lang mit TGF-β (5 ng/ml) behandelt wurden. GAPDH wurde als interne Kontrolle verwendet. Die Ergebnisse werden als Mittelwert ± s.d., n = 3 ausgedrückt. Schüler-T-Test, *P ≤ 0,05, **P ≤ 0,01, ***P ≤ 0,001. (D) NMuMG-Zellen wurden durch Immunfluoreszenz gefärbt, um die Expression des Epithelmarkers E-Cadherin (rot) nach Der TGF-β (2,5 ng/ml) Behandlung für 2 Tage zu erkennen. Nuclei wurden mit DAPI (blau) konterkariert. Die Bilder wurden mit konfokaler Mikroskopie aufgenommen. Scale bar = 50 'm (E) NMuMG-Zellen wurden mit Fluorescein-Phalloidin (grün) gefärbt, um F-Actin zu visualisieren. Nuclei wurden mit DAPI (blau) konterkariert. Maßstabsleiste = 50 m. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 6: Die TGF-Signalisierung und die TGF-β-induzierte EMT wurden durch SB431542 und GW788388 gehemmt. (A) NMuMG-Zellen wurden 1 Stunde lang mit 10 M SB431542 (SB) oder den angegebenen Konzentrationen von GW788388 (GW) in Gegenwart oder Abwesenheit von TGF-β (5 ng/ml) behandelt. Die Zelllysate wurden für p-SMAD2, SMAD2/3 und GAPDH immunoblotted. (B) NMuMG-Zellen wurden mit 5 nM SB431542 (SB) oder 10 M GW788388 (GW) in Gegenwart oder Abwesenheit von 5 ng/ml TGF-β für 1 Stunde behandelt und durch Immunfluoreszenz gefärbt, um die nukleare Translokation von SMAD2/3 (grün) zu erkennen. Die Bilder wurden mit konfokaler Mikroskopie aufgenommen. (C) Die Expression von TGF-β Zielgene, einschließlich PAI-1 und Gene, die EMT-Marker kodieren, einschließlich SNAIL, E-Cadherin und Fibronectin, wurden 48 Stunden lang durch Reverse-Transkriptase-Polymerase-Kettenreaktion (RT-PCR) in NMuMG-Zellen nach SB- oder GW-Behandlung und TGF-β Stimulation analysiert. GAPDH diente als Ladekontrolle. Control bezeichnet nicht behandelte Zellen. Diese Zahl wurde von Petersen M. et al.geändert. 34 mit Genehmigung des Herausgebers. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}

Abbildung 7: EMT-abgeleitete Py2T-murine Brustkrebszellen können induziert werden, um sich in Adipozyten zu differenzieren. Py2T murine Brustkrebszellen wurden mit 2 ng/mL TGF-β für 20 Tage stimuliert, um vollständige EMT zu induzieren. Anschließend wurden die Zellen entweder mit DMSO als Fahrzeugkontrolle oder Rosiglitazon (2 M) für 10 Tage behandelt, um die Differenzierung von mesenchymalen Krebszellen zu ermöglichen und eine Adipogenese zu induzieren. Das Medium wurde alle 2 Tage gewechselt. Nach 10 Tagen Behandlung wurden die Zellen mit ölrot O. Scale bar = 50 m gefärbt. Bitte klicken Sie hier, um eine größere Version dieser Abbildung anzuzeigen.
{kind=link}
| Spezies | Genname | Vorwärts (5' bis 3') | Rückwärts (5' bis 3') | ||
| Menschlichen | GAPDH | TGCACCACCAACTGCTTAGC | GGCATGGACTGTGGTCATGAG | ||
| SMAD7 | TCCAGATGCTGTGCCTTCC | GTCCGAATTGAGCTGTCCG | |||
| SERPINE1 | CACAAATCAGACGGCAGCACT | CATCGGGCGTGGTGAACTC | |||
| Maus | GAPDH | TGGCAAAGTGGAGATTGTTGCC | AAGATGGTGATGGGCTTCCCG | ||
| CDH1 | ACCAAAGTGACGCTGAAGTC | GAGGATGTACTTGGCAATGG | |||
| Schnecke | CAGCTGGCCAGGCTCTCGGT | GCGAGGGCCTCCGGAGCA | |||
| ZEB2 | TTCTGCAAGCCTCTGTAGCC | TTCTGGCCCCATTGCATCAT | |||
Tabelle 1: Primer für qRT-PCR.
Diskussion
TGF-β/SMAD-Signalisierung spielt eine zentrale Rolle bei der Brustkrebsprogression, da sie die Invasivität und Metastasierung von Brustkrebszellen fördern kann, indem emT7induzieren. Hier beschrieben wir einen logischen Workflow zur Untersuchung der TGF-β-initiierten Signalisierung von der rezeptorinduzierten SMAD-Aktivierung bis hin zu SMAD-vermittelten Transkriptions- und biologischen Reaktionen. Wir begannen mit der Beschreibung der Analyse der SMAD2-Phosphorylierung, setzten uns mit TGF-β-induzierten SMAD3-induzierten Transkriptionsreaktionen und EMT-Marker-Expression sowohl auf Gen- als auch auf Proteinebene fort, um die TGF-β/SMAD-Signalreaktion zu analysieren, und untersuchten schließlich TGF-β-induzierte EMT. Wir verwendeten den CAGA12-luciferase Transkriptionsreporter mit CAGA-Boxen, die vom PAI-1-Promoter abgeleitet wurden, um die Aktivität des TGF-β/SMAD-Signalwegs35zu überwachen. Dieses Reporterkonstrukt erfordert SMAD3 und SMAD4 für die Aktivierung. Frühere Studien haben gezeigt, dass der Knockdown von SMAD4 abgeschwächt TGF-β-induzierte CAGA12-luciferase Aktivität37. Neben dem Reporter-Assay ist die Bestimmung des Phosphorylierungsstatus von endogenen SMADs, einschließlich SMAD2 und SMAD3, eine weitere Möglichkeit, die TGF-β Signalreaktion zu untersuchen. Tatsächlich erfassen auch andere Mitglieder der TGF-β Familie, wie z. B. Wachstums- und Differenzierungsfaktor (GDF)-8/Myostatin und GDF-9, Signale über SMAD2/3-Proteine, indem sie T-RI42,43,44. Neben dem CAGA12-luciferase Reporter wurden mehrere ähnliche Reporter verwendet, um die Aktivierung von TGF-β Signalisierung zu erkennen. Beispielsweise kann ein Transkriptionsreporter (SBE)4-Lux mit Antwortelementen, die vom JunB-Promotor abgeleitet wurden, effizient durch TGF-β, Aktivine und BMPs45induziert werden.
Western Blotting und qPCR wurden verwendet, um TGF-β-induzierte EMT zu analysieren, die klassische Methoden zur Untersuchung der Expression von Epithelmarkern (d. h. E-Cadherin) und mesenchymalen Markern (d. h. N-Cadherin, Schnecke, Slug und Zeb2). Wir führten auch indirekte Immunfluoreszenzfärbung von E-Cadherin und direkte Fluoreszenzfärbung von F-Actin durch. Diese Assays bestätigten den mesenchymalen Phänotyp der Zellen nach TGF-β Behandlung. Die Einschränkung der Immunfluoreszenzfärbung besteht darin, dass Zellen vor der Inkubation mit Antikörpern und Bildgebung fixiert werden müssen, und es ist schwierig, Veränderungen der EMT-Markerexpression in lebenden Zellen zu untersuchen. Kürzlich hat das Design von EMT-Reporterzelllinien, wie A549 Lungenadenokarzinom-Vimentin-RFP, es ermöglicht, die Transformation von Epithelzellen in mesenchymale Zellen in Echtzeit über die Expression von rotem fluoreszierendem Protein (RFP)-markiertem Vimentin zu überwachen. Diese Plattform könnte für Drogenscreening und neue Arzneimittelentwicklung genutzt werden46. LifeAct Farbstoff, ein 17-Aminosäure-Peptid, das F-Actin-Strukturen in lebenden Zellen färben kann, wird zu einem wertvollen Werkzeug, um das Aktin-Zytoskelett in Echtzeit zu visualisieren, ohne die zellulären Prozesse zu stören47. In dieser Studie verwendeten wir zwei kleinmolekulare Inhibitoren, SB431542 und GW788388, um ihre hemmende Wirkung auf TGF-β Signalisierung und TGF-β-induzierte EMT zu validieren. Insbesondere hemmt GW788388 die T-RI- und T-RII-Aktivität stark, während SB431542 nur eine hemmende Wirkung auf T-RI (und ALK4 und ALK7) hat. Frühere Studien zeigten, dass GW788388 in vivo stärker ist als SB43154240. Zusätzlich zur Hemmung von EMT reduzierte GW788388 die Expression von Fibrosemarkern in der Niere, und die orale Verabreichung von GW788388 bei diabetischen Mäusen verringerte die Glomerulopathie25,48.
EMT spielt eine wesentliche Rolle bei der Förderung der Plastizität von Krebszellen und führt zu Medikamentenresistenz und Metastasierung 49. Daher wurde die Ausrichtung auf EMT-abgeleitete Zellen mit spezifischen zytotoxischen Medikamenten50 oder die induzierende Redifferenzierung über den mesenchymal-epitheliaalen Übergang (MET)51 als Ansatz zur Überwindung der Metastasierung von Krebszellen und der Therapieresistenz vorgeschlagen. ME trägt jedoch zur Proliferation von disseminierten Krebszellen in entfernten Organenbei 52, was kontraproduktiv sein könnte, wenn die therapeutische Reversion von EMT verwendet wird. Kürzlich berichtete eine neue Studie über einen therapeutischen Transdifferenzierungsansatz, indem EMT-abgeleitete Brustkrebszellen direkt zur Differenzierung in Adipozyten30untersucht wurden. Die Studie von Ishay-Ronen et. 30 verwendete Py2T-Murine-Epithelkrebszellen, die als Reaktion auf eine Langzeitbehandlung mit TGF-β einen Übergang zu mesenchymalen Zellen durchlaufen hatten. Sie zeigten, dass Rosiglitazon in Kombination mit MEK-Inhibitoren die epitheliale Differenzierung und Adipogenese verbesserte. Wir fanden jedoch heraus, dass Rosiglitazon allein ausreichte, um die Transdifferenzierung von mesenchymalen Py2T-Mininzellen in Adipozyten zu induzieren.
Zusammenfassend lässt sich sagen, dass die in dieser Studie verwendeten Methoden einen logischen Workflow zur Untersuchung der TGF-β Signalisierung und der TGF-β-induzierten EMT lieferten. Die beiden Inhibitoren SB431542 und GW788388 können TGF-β-induzierte Reaktionen und EMT blockieren. Darüber hinaus haben wir gezeigt, dass Rosiglitazon allein in bestimmten TGF-β-induzierten mesenchymalen Brustkrebszellen Adipogenese induziert. Obwohl wir nur mehrere Brustkrebs-Zelllinien verwendet haben, um TGF-β-Antworten zu untersuchen, könnten die hier beschriebenen Methoden auf andere (Krebs-)Zellen extrapoliert werden. Hier haben wir verschiedene TGF-β Konzentrationen verwendet, um zelluläre Reaktionen zu induzieren. Bei den meisten Zelltypen übt Die TGF-β ihre biologische Aktivität im Konzentrationsbereich von 0,01-10 ng/mL53 aus und induziert Signalisierung in einem Dosis-Wirkungs-Muster. In primären Endothelzellen, einschließlich der aortenförmigen Endothelzellen von Rindern, induzierte TGF-β die substantielle Expression von phosphoryliertem SMAD2 bei 0,025 ng/ml, erreichte ein Maximum von 0,25 ng/ml und blieb auf diesem Niveau als Reaktion auf höhere Konzentrationen53. In unserer Studie verwendeten wir eine hohe Konzentration von TGF-β (5 ng/ml) in MCF10A-Ras-Zellen für den Transkriptionsreporter-Assay, um starke Antworten zu erhalten. SMAD2-Phosphorylierung und Zielgenexpression können durch TGF-β in niedriger Dosis induziert werden; Daher verwendeten wir 2,5 ng/ml TGF-β zur Behandlung von Zellen. Die am besten geeignete Arbeitskonzentration hängt jedoch vom Zelltyp und den geschätzten Wirkungen ab. Zur Bestimmung der besten Konzentration von TGF-β wird die Behandlung der Zellen mit unterschiedlichen Dosen (von niedrig bis hoch) empfohlen.
Offenlegungen
Die Autoren haben nichts zu verraten.
Danksagungen
Wir würdigen die Unterstützung des Chinese Scholarship Council (CSC) für J.Z. und das Cancer Genomics Centre Netherlands (CGC. NL) zu P.t.D.
Materialien
| Name | Company | Catalog Number | Comments |
| Reagent | |||
| 18 mm-side square glass coverslips | Menzel Gläser | 631-1331 | |
| 4′,6-diamidino-2-phenylindole (DAPI) | Vector Laboratories | H-1200 | |
| Alexa Fluor 488 Phalloidin | Thermo Fisher Scientific | A12379 | |
| Alexa Fluor 555 secondary antibody | Thermo Fisher Scientific | A-21422 | |
| Anti-E-cadherin antibody | BD Biosciences | 610181 | |
| anti-glyceraldehyde 3-phosphate dehydrogenase(GAPDH) antibody | Merck Millipore | MAB374 | |
| Anti-N-cadherin antibody | BD Biosciences | 610920 | |
| Anti-Slug antibody | Cell Signaling Technology | 9585 | |
| anti-SMAD2/3 antibody | Becton Dickinson | 610842 | |
| Anti-Snail antibody | Cell Signaling Technology | 3879 | |
| Anti-Tubulin antibody | Cell Signaling Technology | 2148 | |
| Bovine Serum Albumin | Sigma-Aldrich | A2058 | |
| Cholera enterotoxin | Sigma-Aldrich | C8052 | |
| Clarity Western ECL Substrate | Bio-Rad | 1705060 | |
| DC protein assay kit | Bio-Rad | 5000111 | |
| DMEM-high glucose | Thermo Fisher Scientific | 11965092 | |
| DMEM-high glucose medium | Thermo Fisher Scientific | 11965092 | |
| Dulbecco’s modified Eagle’s medium (DMEM)/F12 | Thermo Fisher Scientific | 11039047 | |
| epidermal growth factor (EGF) | Merck Millipore | 01-107 | |
| Fetal bovine serum (FBS) | BioWest | S1860-500 | |
| GoTaq qPCR Master Mix | PROMEGA | A600X | |
| Horse serum | Thermo Fisher Scientific | 26050088 | |
| Hydrocortisone | Sigma-Aldrich | H0135 | |
| Insulin | Sigma-Aldrich | 91077C | |
| Mini Protease Inhibitor Cocktail | Roche | 11836153001 | |
| NucleoSpin RNA II kit | BIOKE´ | 740955 | |
| Penicillin-streptomycin (Pen-Strep) | Thermo Fisher Scientific | 15140148 | |
| Polyethylenimine (PEI) | Polyscience | 23966 | |
| Polyvinylidene difluoride (PVDF) membrane | Merck Millipore | IPVH00010 | |
| Ponceau S solution | Sigma-Aldrich | P7170 | |
| RevertAid First Strand cDNA Synthesis Kit | Thermo Fisher Scientific | K1621 | |
| Rosiglitazone | Sigma-Aldrich | 557366-M | |
| Skimmed milk | Campina: Elk | ||
| Equipment | |||
| ChemiDoc Imaging System | Bio-Rad | 17001402 | |
| CFX Connect Detection System | Bio-Rad | 1855201 | |
| Luminometer | Perkin Elmer | 2030-0050 | |
| NanoDrop 2000/2000c Spectrophotometers | Thermo Fisher Scientific | ND-2000 |
Referenzen
- Tzavlaki, K., Moustakas, A. TGF-β Signaling. Biomolecules. 10 (3), (2020).
- Hao, Y., Baker, D., Ten Dijke, P. TGF-β-Mediated Epithelial-Mesenchymal Transition and Cancer Metastasis. International Journal of Molecular Sciences. 20 (11), (2019).
- Morikawa, M., Derynck, R., Miyazono, K. TGF-β and the TGF-β Family: Context-Dependent Roles in Cell and Tissue Physiology. Cold Spring Harbor Perspectives in Biology. 8 (5), (2016).
- Derynck, R., Akhurst, R. J. Differentiation plasticity regulated by TGF-β family proteins in development and disease. Nature Cell Biology. 9 (9), 1000-1004 (2007).
- Massague, J. How cells read TGF-β signals. Nature Reviews Molecular Cell Biology. 1 (3), 169-178 (2000).
- Seoane, J., Gomis, R. R. TGF-β Family Signaling in Tumor Suppression and Cancer Progression. Cold Spring Harbor Perspectives in Biology. 9 (12), (2017).
- Colak, S., Ten Dijke, P. Targeting TGF-β Signaling in Cancer. Trends in Cancer. 3 (1), 56-71 (2017).
- Suriyamurthy, S., Baker, D., ten Dijke, P., Iyengar, P. V. Epigenetic Reprogramming of TGF-β Signaling in Breast Cancer. Cancers (Basel). 11 (5), (2019).
- Drabsch, Y., Ten Dijke, P. TGF-β signalling and its role in cancer progression and metastasis). Cancer and Metastasis Reviews. 31 (3-4), 553-568 (2012).
- Goumans, M. J., Ten Dijke, P. TGF-β Signaling in Control of Cardiovascular Function. Cold Spring Harbor Perspectives in Biology. 10 (2), (2018).
- Robertson, I. B., Rifkin, D. B. Regulation of the Bioavailability of TGF-β and TGF-β-Related Proteins. Cold Spring Harbor Perspectives in Biology. 8 (6), (2016).
- Robertson, I. B., et al. Latent TGF-β-binding proteins. Matrix Biology. 47, 44-53 (2015).
- Khan, Z., Marshall, J. F. The role of integrins in TGF-β activation in the tumour stroma. Cell and Tissue Research. 365 (3), 657-673 (2016).
- Jenkins, G. The role of proteases in transforming growth factor-β activation. The International Journal of Biochemistry & Cell Biology. 40 (6-7), 1068-1078 (2008).
- Tran, D. Q., et al. GARP (LRRC32) is essential for the surface expression of latent TGF-β on platelets and activated FOXP3+ regulatory T cells. Proceedings of the National Academy of Sciences of the United States of America. 106 (32), 13445-13450 (2009).
- Stockis, J., Colau, D., Coulie, P. G., Lucas, S. Membrane protein GARP is a receptor for latent TGF-β on the surface of activated human Treg. European Journal of Immunology. 39 (12), 3315-3322 (2009).
- Wang, R., et al. GARP regulates the bioavailability and activation of TGF-β. Molecular Biology of the Cell. 23 (6), 1129-1139 (2012).
- Vander Ark, A., Cao, J., Li, X. TGF-β receptors: In and beyond TGF-β signaling. Cellular Signalling. 52, 112-120 (2018).
- Heldin, C. H., Miyazono, K., ten Dijke, P. TGF-β signalling from cell membrane to nucleus through SMAD proteins. Nature. 390 (6659), 465-471 (1997).
- ten Dijke, P., Hill, C. S. New insights into TGF-β-Smad signalling. Trends in Biochemical Sciences. 29 (5), 265-273 (2004).
- Miyazono, K. TGF-β signaling by Smad proteins. Cytokine & Growth Factor Reviews. 11 (1-2), 15-22 (2000).
- Yu, Y., Feng, X. H. TGF-β signaling in cell fate control and cancer. Current Opinion in Cell Biology. 61, 56-63 (2019).
- Zhang, Y. E. Non-Smad pathways in TGF-β signaling. Cell Research. 19 (1), 128-139 (2009).
- Katsuno, Y., Lamouille, S., Derynck, R. TGF-β signaling and epithelial-mesenchymal transition in cancer progression. Current Opinion in Oncology. 25 (1), 76-84 (2013).
- Lamouille, S., Xu, J., Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nature Reviews Molecular Cell Biology. 15 (3), 178-196 (2014).
- Yang, J., et al. Guidelines and definitions for research on epithelial-mesenchymal transition. Nature Reviews Molecular Cell Biology. 21 (6), 341-352 (2020).
- Zhang, Y., Weinberg, R. A. Epithelial-to-mesenchymal transition in cancer: complexity and opportunities. Frontiers in Medicine. 12 (4), 361-373 (2018).
- Brabletz, T., Kalluri, R., Nieto, M. A., Weinberg, R. A. EMT in cancer. Nature Reviews Cancer. 18 (2), 128-134 (2018).
- Ocana, O. H., et al. Metastatic colonization requires the repression of the epithelial-mesenchymal transition inducer Prrx1. Cancer Cell. 22 (6), 709-724 (2012).
- Ishay-Ronen, D., et al. Gain Fat-Lose Metastasis: Converting Invasive Breast Cancer Cells into Adipocytes Inhibits Cancer Metastasis. Cancer Cell. 35 (1), 17-32 (2019).
- Smith, C. J., Osborn, A. M. Advantages and limitations of quantitative PCR (Q-PCR)-based approaches in microbial ecology. FEMS Microbiology Ecology. 67 (1), 6-20 (2009).
- Bustin, S. A., Mueller, R. Real-time reverse transcription PCR (qRT-PCR) and its potential use in clinical diagnosis. Clinical Science. 109 (4), 365-379 (2005).
- Sundqvist, A., et al. JUNB governs a feed-forward network of TGF-β signaling that aggravates breast cancer invasion. Nucleic Acids Research. 46 (3), 1180-1195 (2018).
- Persson, U., et al. The L45 loop in type I receptors for TGF-β family members is a critical determinant in specifying Smad isoform activation. FEBS Letters. 434 (1-2), 83-87 (1998).
- Dennler, S., et al. Direct binding of Smad3 and Smad4 to critical TGF-β-inducible elements in the promoter of human plasminogen activator inhibitor-type 1 gene. The EMBO Journal. 17 (11), 3091-3100 (1998).
- Piek, E., Moustakas, A., Kurisaki, A., Heldin, C. H., ten Dijke, P. TGF-β type I receptor/ALK-5 and Smad proteins mediate epithelial to mesenchymal transdifferentiation in NMuMG breast epithelial cells. Journal of Cell Science. 112, Pt 24 4557-4568 (1999).
- Deckers, M., et al. The tumor suppressor Smad4 is required for transforming growth factor-β-induced epithelial to mesenchymal transition and bone metastasis of breast cancer cells. Cancer Research. 66 (4), 2202-2209 (2006).
- Souchelnytskyi, S., et al. Phosphorylation of Ser465 and Ser467 in the C terminus of Smad2 mediates interaction with Smad4 and is required for transforming growth factor-β signaling. Journal of Biological Chemistry. 272 (44), 28107-28115 (1997).
- Abdollah, S., et al. TβRI phosphorylation of Smad2 on Ser465 and Ser467 is required for Smad2-Smad4 complex formation and signaling. Journal of Biological Chemistry. 272 (44), 27678-27685 (1997).
- Petersen, M., et al. Oral administration of GW788388, an inhibitor of TGF-β type I and II receptor kinases, decreases renal fibrosis. Kidney International. 73 (6), 705-715 (2008).
- Gubelmann, C., et al. Identification of the transcription factor ZEB1 as a central component of the adipogenic gene regulatory network. Elife. 3, 03346(2014).
- Nickel, J., Ten Dijke, P., Mueller, T. D. TGF-β family co-receptor function and signaling. Acta Biochimica et Biophysica Sinica. 50 (1), 12-36 (2018).
- Mazerbourg, S., et al. Growth differentiation factor-9 signaling is mediated by the type I receptor, activin receptor-like kinase 5. Molecular Endocrinology. 18 (3), 653-665 (2004).
- Rebbapragada, A., Benchabane, H., Wrana, J. L., Celeste, A. J., Attisano, L. Myostatin signals through a transforming growth factor-β-like signaling pathway to block adipogenesis. Molecular and Cellular Biology. 23 (20), 7230-7242 (2003).
- Jonk, L. J., Itoh, S., Heldin, C. H., ten Dijke, P., Kruijer, W. Identification and functional characterization of a Smad binding element (SBE) in the JunB promoter that acts as a transforming growth factor-β, activin, and bone morphogenetic protein-inducible enhancer. Journal of Biological Chemistry. 273 (33), 21145-21152 (1998).
- Thiery, J. P., Sleeman, J. P. Complex networks orchestrate epithelial-mesenchymal transitions. Nature Reviews Molecular Cell Biology. 7 (2), 131-142 (2006).
- Riedl, J., et al. Lifeact: a versatile marker to visualize F-actin. Nature Methods. 5 (7), 605-607 (2008).
- Gellibert, F., et al. Discovery of 4-{4-[3-(pyridin-2-yl)-1H-pyrazol-4-yl]pyridin-2-yl}-N-(tetrahydro-2H- pyran-4-yl)benzamide (GW788388): a potent, selective, and orally active transforming growth factor-β type I receptor inhibitor. Journal of Medicinal Chemistry. 49 (7), 2210-2221 (2006).
- Ishay-Ronen, D., Christofori, G. Targeting Cancer Cell Metastasis by Converting Cancer Cells into Fat. Cancer Research. 79 (21), 5471-5475 (2019).
- Gupta, P. B., et al. Identification of selective inhibitors of cancer stem cells by high-throughput screening. Cell. 138 (4), 645-659 (2009).
- Pattabiraman, D. R., et al. Activation of PKA leads to mesenchymal-to-epithelial transition and loss of tumor-initiating ability. Science. 351 (6277), (2016).
- Gao, D., et al. Myeloid progenitor cells in the premetastatic lung promote metastases by inducing mesenchymal to epithelial transition. Cancer Research. 72 (6), 1384-1394 (2012).
- Goumans, M. J., et al. Balancing the activation state of the endothelium via two distinct TGF-β type I receptors. The EMBO Journal. 21 (7), 1743-1753 (2002).
Erratum
Formal Correction: Erratum: Studying TGF-β Signaling and TGF-β-induced Epithelial-to-mesenchymal Transition in Breast Cancer and Normal Cells
Posted by JoVE Editors on 12/15/2020. Citeable Link.
An erratum was issued for: Studying TGF-β Signaling and TGF-β-induced Epithelial-to-mesenchymal Transition in Breast Cancer and Normal Cells. NMuMG was mislabeled as cancer cells.
NMuMG cells are non-transformed epithelial breast cells established from a mouse mammary gland. NMuMG cells are frequently used as a model system to investigate TGF-β-induced epithelial to mesenchymal transition (EMT). EMT is induced by TGF-β in premalignant human MCF 10A-RAS (M2) cells and mouse MMTV-PyMT breast cancer cell line. The article has been updated to reflect this.
Nachdrucke und Genehmigungen
Genehmigung beantragen, um den Text oder die Abbildungen dieses JoVE-Artikels zu verwenden
Genehmigung beantragenThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Alle Rechte vorbehalten