
Method Article
発現クローニングによりGrb2のシグナル伝達複合体を研究するために高コンテンツイメージングワークフロー
要約
有能な膜貫通型受容体のシグナリング小説を同定するためのハイコンテンツスクリーニング方法が記載されている。この方法は、大規模な自動化に適しているとの予測を可能にするタンパク質の結合および哺乳動物細胞におけるタンパク質複合体の細胞内局在。
要約
成長因子受容体によるシグナル伝達は、細胞が増殖と分化を維持するために不可欠であり、厳格な管理を必要とします。シグナル伝達は、膜貫通型受容体と下流シグナル伝達カスケードの活性化に外部リガンドの結合によって開始される。分裂促進シグナル伝達の重要な調節因子はGrb2は、酵素活性を欠いている2 SH3ドメインが隣接内部SH2(Src相同2)ドメインから成るモジュラー蛋白質である。 Grb2のは構成的にそのN末端SH3ドメインを介して、GTPアーゼソン·オブ·Sevenless(SOS)に関連付けられています。 Grb2のSH2ドメインは、このように、SOS-RAS-MAPキナーゼシグナル伝達カスケードに受容体の活性化を結合するリン酸化チロシン残基における成長因子受容体に結合する。また、シグナリングおよび受容体のエンドサイトーシスの正または負の調節因子としてGrb2のための他の役割が記載されている。 Grb2のモジュラー組成物は、受容体とtransducの様にドッキングできることを示唆している1月3日の異なる経路に沿って多数の電子信号。
ここで説明するには、形質膜へのGrb2の募集を監視する簡単な顕微鏡検査アッセイである。それが刺激に応答して、4月6日 、緑色蛍光タンパク質(GFP)タグ付きGrb2の細胞内局在の変化を測定するアッセイから適合させられます。 cDNA発現により細胞膜に、例えば活性化上皮成長因子受容体(EGFR)リクルート、GFP-Grb2のGrb2のように結合し、続いて細胞内のエンドソーム区画に再配置することを原形質膜受容体。 Grb2のin vivoでのタンパク質複合体で識別するために、この技術はGrb2の細胞内局在の変化に基づいてゲノムワイドな高コンテンツ画面を実行するために使用することができます。 cDNA発現クローンを、トランスフェクションおよび画像取得の調製は、以下に詳細に説明されています。このような酵母2-HYなどのタンパク質相互作用パートナーを識別するために使用される他のゲノム手法と比較して、BRIDは、この手法は単純な顕微鏡ベースアッセイによる相互作用のサブ携帯サイトで哺乳動物細胞におけるタンパク質複合体の可視化を可能にします。したがって、そのような局在化のパターンとして質的特徴は、同様に相互作用の定量的な強さとして、評価することができる。
プロトコル
1。 cDNA発現クローンをピッキング
- cDNAライブラリーは、96ウェルフォーマットでグリセロールストックの単一の細菌クローンとして提供されています。 Norgen CP7200のピッカーメニューでRearrayコマンドを使用して、96ディープウェル(井戸1ミリリットル)をプレートに1.5 mlのLB /アンプにソースプレートや接種から細菌クローンを選択します。
- ガス透過性のシールでディープウェルプレートを密封し、振とう機で37℃で一晩インキュベートする。
2。 cDNA発現ライブラリーの作製
- テーブルトップ遠心機でプレートアダプタを使用し、20分間2,000 rpmで細菌を含むディープウェルプレートを回転させます。
- 細菌ペレットをそのまま残して、ウェルから培地を吸引。
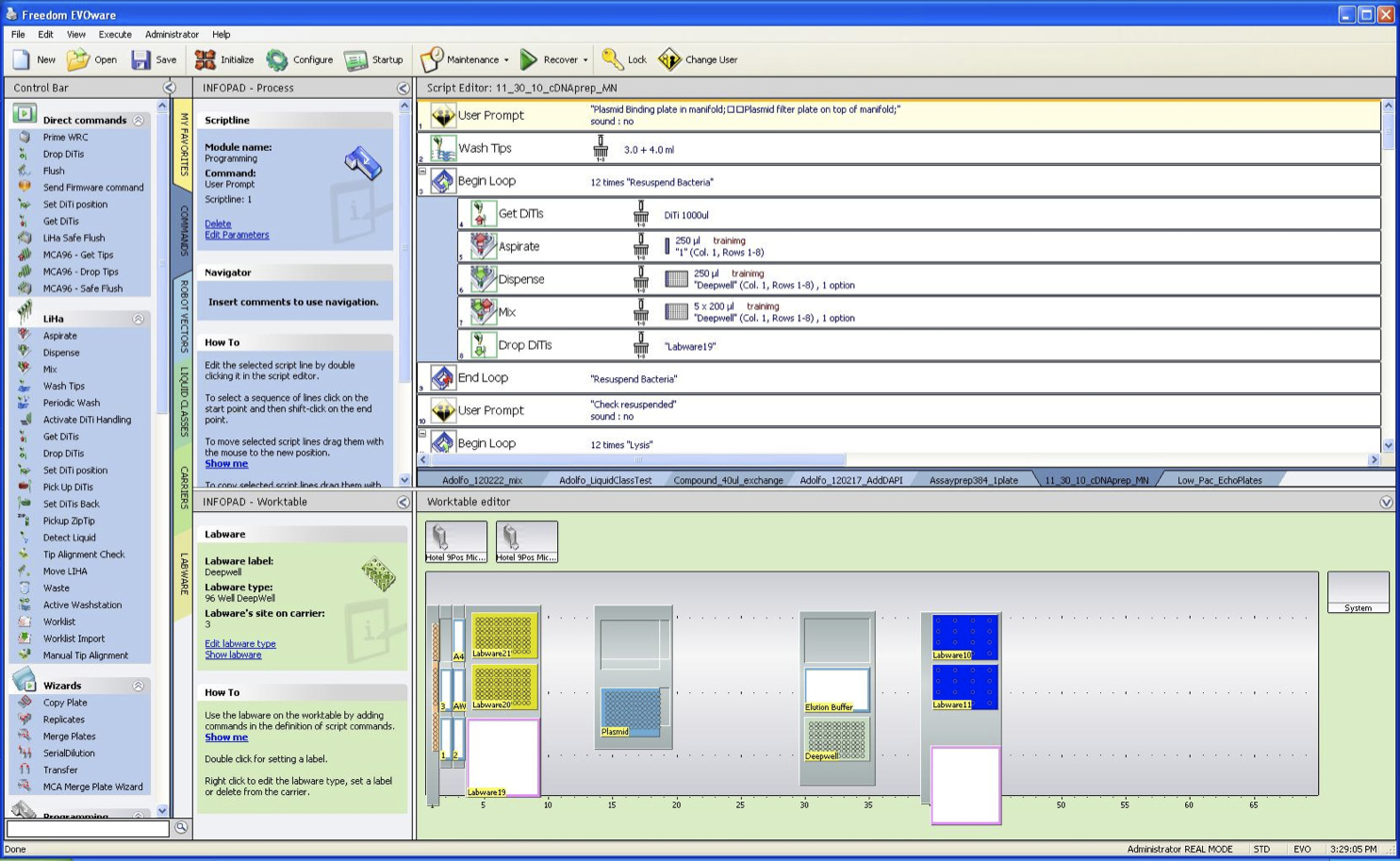
- 図2に示すように、ラボ·オートメーション·ワークステーションのデッキを構成します。プラスミド調製用の溶液はトラフ1月5日に配布されており、ディープウェルプレートはデッキ上に配置されます。溶出溶液によるマルチウェルトラフディープウェルプレートに隣接して配置される。ヒントはチップラックにロードされます。
- 真空マニホールドを組み立てます。プラスミド結合プレートはマニホールドの内部に配置され、プラスミドフィルタープレート、マニホールドの上に配置されます。
- ピペッティングにより250μlの溶液1(再懸濁緩衝液)でペレットを再懸濁します。
- 250μlの溶液2(溶解バッファー)を添加することにより細菌を溶解します。プレートを密封し、完全な混合を可能にするために反転されます。
- 2分のタイマーを起動します。
- 350μlの溶液3(中和バッファー)を追加します。プレートをシールし、3回転倒。
- プラスミドフィルタープレートに細菌溶解物を転送します。
- 5分間吸引してください。
- フィルタープレートを取り外し、マニホールドの上に結合プレートを配置します。 1分間吸引してください。
- 追加の洗浄(AW)のバッファー500μlを加える。
- 2分間吸引してください。
- 洗浄緩衝液4の900μlを添加する。 2分間吸引してください。
- 繰り返しの2.14 TEP。さらに15分間吸引してください。
- マニホールド内のDNAコレクションプレートを置きます。結合プレートに溶出バッファー100μLを加え、2分間インキュベートする。
- 10分間真空。
- 8000ではを使用して、コレクションプレートとメジャーDNA濃度を取り外します。一般的に〜の収量は100 ng /μlに、この方法で期待することができる。
3。細胞播種
- HEK293細胞をトリプシン処理し、計数されています。
- 2×10 4個の細胞をThermoFisher 96ウェルマルチドロップを使用して、パーキンエルマーViewplateの各ウェルに分注されています。
- 細胞を37℃で一晩インキュベート℃および5%CO 2で 。
4。トランスフェクション
- 丸底96ウェルプレートのウェルあたり25μlの無血清培地でのcDNAを100ngと100ngのGFP-Grb2のプラスミドDNAのミックスを調製する。
- ウェルあたり0.5μlのTransfectinを含む25μlの無血清培地を追加します。
- ミックスとタイマーfoを開始R 30分。
- 穏やかなディスペンス法を用いて細胞にTransfectinのcDNA /ミックス50μlを移します。代わりに、トランスフェクション混合物をプレートに分注し、その後第一細胞が上に追加することができます(リバーストランスフェクション)。
- ℃、5%CO 2、37℃で一晩細胞をインキュベートします。
5。画像収集
- オペラソフトウェアを起動します。実験のセットアップの画面のスナップショットを図3に示します。 "設定"タブを選択し、20倍、客観的かつ正確なプレートタイプを選択します。 "襟"は異なるプレートタイプでピントを可能にするために客観的に正しい値に設定されていることを確認します。
- "顕微鏡"タブを選択します。 FITC(488レーザ)やUVなどの露出2(365レーザー)などの露出の定義1。両方のエクスポージャーのUVフィルタをアクティブにして、カメラ1とカメラ2への露出2〜露出1を割り当てます。露出1露出2の40ミリに対して800ミリ秒までの露光時間を設定します。
- "顕微鏡"タブを選択します。 eを定義FITC(488レーザ)やUVなどの露出2(365レーザー)などxposure 1。両方のエクスポージャーのUVフィルタをアクティブにして、カメラ1とカメラ2への露出2〜露出1を割り当てます。露出1露出2の40ミリに対して800ミリ秒までの露光時間を設定します。
- 暴露1を選択します。高さが0μmにフォーカスを設定します。 "フォーカス"を選択します。一度当て、カメラ1を公開します。露出面を最適化し、 "高さを取る"をクリックして、フォーカスの高さを調整します。 〜3000の最大ピクセル強度を与えるために露光時間とレーザーパワーを変更します。露出パラメータを保存します。露出2についても同じ手順を繰り返します。
- "実験の定義"タブを選択します。レイアウトとsublayoutを作成します。関連するレイアウト、露出、参照画像、skewcropファイルとsublayoutをドラッグアンドドロップします。実験を保存します。
- "自動実験"タブを選択し、画像を取得。
6。画像解析(1.46h ImageJのバージョンに基づいています。)
- 使用して独自のパーキンに変換します。フレックスファイルがタグ付きのイメージ形式に変換します。tifファイルFlex2Volocityアカペラスクリプト。
- スタックで、その結果、イメージシーケンスとしてはImageJで画像をインポートします。
- Hyperstackメニュー項目にイメージ/ Hyperstack /スタックを選択することにより、2チャンネルhyperstackにスタックに変換します。スライスの数は、インポートされたスタックの半分に設定する必要があります。
- イメージ/ DUPLICATEコマンドを使用してGFPのチャネルの重複:重複hyperstackのチェックボックスをマークしてチャンネル指定:1-1。来世重複スタックを使用します。
- プロセスを選択/スタックヒストグラムを用いて、0.01%の飽和画素のコントラストを高めるとImage/Type/8-bitと8ビットに変換します。
- コントラスト閾値(パラメータ1)Q = 30とバーンセンアルゴリズムを使用して、15ピクセル半径ローカル適応分割を定義します。画像/調整/自動ローカルしきい値メニューに黒の背景に白のチェックボックスオブジェクトにチェックを入れます。
- セグメンテーションの品質管理:元の画像に分割されたオブジェクトの輪郭オーバーレイを生成します。
- さまざまな機能測定領域は、それぞれの細胞小器官の平均と合計強度:粒子を分析することによって抽出し直す。結果ファイルを保存します。
- Rの機能の結果ファイルを開き、ヒストグラムを生成します。
7。代表的な結果
通常の条件下ではGrb2のは、全体の細胞全体にローカライズされています。増殖因子受容体の刺激によって、それは細胞膜に転位し、その後、 図4に示すように、エンドソームに取り込まれます。バインディングGrb2のできる細胞表面受容体の発現は、この移行を誘導するのに十分である。典型的なスクリーニング実験では、GFP-Grb2の局在化には変化は観察されない。しかし、タンパク質を発現させたときに、サブ携帯サイトへの新兵のGrb2は、蛍光顕微鏡により容易に視覚化することができる局在の変化があること。エンドソーム様構造へのGFP-Grb2の再局在におけるEGFR結果のインスタンス、発現のために( 図4)。このように、ゲノムワイドなライブラリを適用するとき、それは小説Grb2結合する細胞表面タンパク質を同定することができることが期待できる。
この方法は、細胞表面タンパク質の検出に限定されない。例えば、Dynamin2は、GFP-Grb2のは、エンドソームのような募集します ( 図5)を表示するの細胞内局在の変化を誘導する。 Dynamin2はGrb2のC末端のSH3ドメインに結合し、この複合体9,10のエンドサイトーシスに関与している。したがって、このアプローチは、一般的なタンパク質の結合複合体の同定を可能にし、SH2ドメインの相互作用パートナーの同定に限定されるものではない。
転座のいくつかの異なるタイプは、原形質膜、エンドソームに、他の細胞質小胞の構造や核への局在化が期待できる。したがって、すべての画像が、目の検査を受けておくことをお勧めします。しかし、cDNAの大規模なセットをスクリーニングする際に自動画像解析アルゴリズムは、より実用的である。このケースでは、アルゴリズムの組み合わせは、このような細胞の形状の変化を識別するために往復核および一般形態学的なアルゴリズムを識別するために、エンドソーム、細胞質/核検出を識別するために、スポットの検出として、望ましい。これらの異なる表現型の検出方法の使用は、他の場所で11に詳細に議論されてきた。

図1実験の全体的なスキーム。実験は細部cDNAライブラリーの増幅と準備、無料のオープンソフトウェアImageJを用いて、cDNAベクターをプラスレポーターコンストラクト、画像取得と画像解析のトランスフェクションで始まる完全な高コンテンツスクリーニングワークフローを。

Fテカンデッキのigure 2章構成。 (1)左側で、5百ミリリットル谷はバッファ1(40ミリリットル)、2(40ミリリットル)、3(50ミリリットル)、AW(60ミリリットル)とA4(100ミリリットル)で満たされる必要があります。トラフA4は、手順の間に一度再充填する必要があります。 (2)真空マニホールドは、この例では14番目に位置しています。 (3)細菌ペレットとディープウェルプレートは25mlの溶出緩衝液で溶出バッファートラフの隣に、30位に位置しています。 (4)8チャネル·ヘッド用使い捨てヒントは左側にチップラックにロードする必要があり、マルチチャンネルヘッドのためのヒントは、位置40に記入する必要があります。この実験で使用されているテカンFreedomEvo液ハンドラが500μlの注射器が装備されています。 拡大図を表示するには、ここをクリックしてください 。
{kind=link}

図オペラLX上の画像取得の3。オートメーション。 A)の[設定]タブ(1)を選択します。実験(2)に必要なレーザーラインをアクティブにします。画像取得(3)カメラを選択します。実験(4)のためのプレート型と対物レンズを選択します。 B)は顕微鏡タブ(1)を選択します。露出1(2,3)用の光源と露光時間を定義します。よく1に焦点を当てると高さ(4)を取る。 C)実験の定義]タブ(1)を選択します。ドラッグや露出、skewcrop、リファレンス、レイアウトやsublayoutファイル(2)をドロップします。実験ファイル(3)をドラッグ·アンド·ドロップします。 D)の自動実験(1)を選択します。実験ファイル(2)をドラッグ·アンド·ドロップします。適切なバーコード(3)を割り当てます。スタートボタン(4)をクリックすることで画像収集を開始します。 拡大図を表示するには、ここをクリックしてください 。
{kind=link}

図4、GFP-Grb2の転。 GFP-Grb2は貫通成長因子受容体の過剰発現時に優勢な細胞質に局在からエンドソーム様構造へ移転し、その後エンドソームに内部化する。左側には、COS M6の細胞の顕微鏡画像を一過性のGFP-Grb2の(上部)またはGFP-Grb2のプラスEGFR(下)を用いてトランスフェクトしたことを示しています。エンドソーム様構造へのGFP-Grb2の移転におけるEGFR結果の共発現。

図5、GFP-Grb2のcDNAを誘起転座の例。 Grb2の体を介したエンドサイトーシスに関与するGTPアーゼDynamin2は、エンドソーム様構造(円内)に、GFP-Grb2は再配置されます。この実験では、GFP-Grb2のはHEK293T細胞にDNM2で同時トランスフェクトされた。 24時間と画像がOPEで取得した後、細胞をHoechst 33342で染色したRA LX。ワングリーン(GFP)のチャンネルの視野とUV(青;ヘキスト33342)が表示されます。
ディスカッション
発現クローニングは、このようなウイルス受容体および血液細胞抗原12のような斬新な細胞成分を同定するために、過去に使用されている強力なツールです。ここでは、Grb2のに結合する新規な推定シグナル伝達受容体の同定を容易にする方法を説明します。
プロトコルのいくつかの重要なステップがあります。
- 自動化されたプラスミドの調製のためには、細菌ペレットを完全に再懸濁を持つことが必要不可欠です。時には、それは厳密な揺れステップまたはピペットでさらに混合を含める必要があります。
- ソリューション2と3を添加すると、完全な混合が不可欠であると我々は手動で反転封印されたプレートが自動化されたシェーカーを使用するよりも効率的であることを見出した。それはDNAの切断につながるとしてピはこのステップで避けるべきである。
- すべての真空手順では、1は、液体が完全に排出されていることを確認する必要がありますプレートから。そうでなければ、低収率は予想されています。
- 溶出の際には、溶出液は、プラスミド結合プレートの下部にあるフォームをドロップすることは非常に一般的です。これらのドロップは、溶出プレートに収集する必要があるので、プラスミドの結合プレートを慎重に持ち上げられ、下部に残留滴を慎重に溶出液をプレートに移しておくことをお勧めします。
- トランスフェクション時には、複合体形成の時間が重要です。我々は、通常20〜30分に準拠しています。短い時間が推奨されていないながら、最高1時間まで長い時間が、歩留まりを低下させることなく問題ありません。
- 顕微鏡は、倍率の選択が重要になります。ここで使用されるオペラLXのシステムでは、20倍で解像度は、高品質の画像を得るために十分である。高解像度の画像が必要な場合は、目的を変更することができますが、セルの統計的に有意な数値を得るためには、フィールドと高い取得時間数の増加が必要とされる。一般的に、我々は目指してウェルごとに結像される少なくとも100個の細胞を得ることができます。
Grb2の転座検定は、EGFRキナーゼ活性化6の小分子阻害剤を識別するために、他のグループによって使用されています。その場合には、EGFRは、具体的にリガンドが細胞膜にGrb2の動員を引き起こすで有効になります。この相互作用の崩壊は、その後、小分子化合物を用いて検討することができます。同様に、それは、siRNAスクリーンは、c-KITまたはエリスロポエチン受容体などのEGFR-Grb2のシグナリングまたは他のGrb2結合成長因子受容体に関与する内因性遺伝子を同定するために用いることができることを想定することができる。従って、この技術のために複数の潜在的用途がある。類似したアプローチは、Shcを、GABまたはIRSなどの他のアダプター分子のためのGFPタグレポーターシステムに適用することができる。
哺乳動物細胞で、この細胞ベースのアッセイを使用する大きな利点の1つは、それが生理的にrelの識別を可能にするということですevant相互作用。アッセイは、タンパク質複合体形成のために読み出していますが、より重要なのは、関連する相互作用が正しいサブ携帯サイトで監視されます。この点では、この技術は、酵母ツーハイブリッドまたはin vitroアッセイのような他のタンパク質-タンパク質相互作用の方法から成果物を克服しています。それは間接的な相互作用はまたGrb2の募集をもたらす可能性があること、しかし注意すべきである。同様に、結合パートナーの転写アップレギュレーションは、cDNA発現によって誘導することができる。これらの可能性を区別するためには、直接的および間接的な結合の効果を区別するために、適切な二次アッセイを行うことが必要である。
結論としては、GFP-アダプター分子転アッセイは、ゲノムワイドなスクリーニングや創薬アプリケーションのための高いポテンシャルをお約束します。
開示事項
特別な利害関係は宣言されません。
謝辞
この作品は、医学研究評議会とマリー·キュリー国際社会復帰グラント方式(JKVへ)によってサポートされていました。
資料
| 試薬の名称 | 会社 | カタログ番号 | コメント(オプション) |
| cDNAライブラリー | Origene | ||
| LB +アンペア | |||
| ガス透過性のシール | ThermoScientific | AB-0718 | |
| Nucleospin 96プラスミドキット | マシュレ-ナーゲル | 740625.4 | |
| Transfectin | BioRad社 | 170-3351 | |
| Hoechst33342 | 分子プローブ | H3570 | |
| Viewplate 96 F TC | パーキンエルマー | 6005182 | |
| RapidPick | ハドソン | 当社までCP7200 | |
| 自由エボ | テカン | 真空マニホールド付き | |
| 384マルチドロップ | サーモ | ||
| オペラLX | パーキンエルマー |
参考文献
- Sastry, L., Cao, T., King, C. R. Multiple Grb2-protein complexes in human cancer cells. Int. J. Cancer. 70, 208-213 (1997).
- Li, S., Couvillon, A. D., Brasher, B. B., Van Etten, R. A. Tyrosine phosphorylation of Grb2 by Bcr/Abl and epidermal growth factor receptor: a novel regulatory mechanism for tyrosine kinase signaling. Embo. J. 20, 6793-6804 (2001).
- Bisson, N. Selected reaction monitoring mass spectrometry reveals the dynamics of signaling through the GRB2 adaptor. Nat. Biotechnol. 29, 653-658 (2011).
- Ketteler, R. The Feynman trajectories: determining the path of a protein using fixed-endpoint assays. J. Biomol. Screen. 15, 321-326 (2010).
- Yamazaki, T. Role of Grb2 in EGF-stimulated EGFR internalization. J. Cell. Sci. 115, 1791-1802 (2002).
- Antczak, A. Domain-Based Biosensor Assay to Screen for Epidermal Growth Factor Receptor Modulators. in Live Cells. ASSAY and Drug Development Technologies. 10, 24-36 (2012).
- Su, J., Batzer, A., Sap, J. Receptor tyrosine phosphatase R-PTP-alpha is tyrosine-phosphorylated and associated with the adaptor protein Grb2. J. Biol. Chem. 269, 18731-18734 (1994).
- Hertog, J. den, Tracy, S., Hunter, T. Phosphorylation of receptor protein-tyrosine phosphatase alpha on Tyr789, a binding site for the SH3-SH2-SH3 adaptor protein GRB-2 in vivo. Embo. J. 13, 3020-3032 (1994).
- Miki, H. Association of Ash/Grb-2 with dynamin through the Src homology 3 domain. J. Biol. Chem. 269, 5489-5492 (1994).
- Seedorf, K. Dynamin binds to SH3 domains of phospholipase C gamma and GRB-2. J. Biol. Chem. 269, 16009-16014 (1994).
- Carpenter, A. E. CellProfiler: image analysis software for identifying and quantifying cell phenotypes. Genome Biol. 7, R100(2006).
- Seed, B. Developments in expression cloning. Curr. Opin. Biotechnol. 6, 567-573 (1995).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved