
Method Article
癌細胞スフェロイド培養における潜在的に新規薬剤標的を同定するためのスクリーニング機能ゲノミクスを利用
要約
Identifying novel drug targets that transition from pre-clinical testing to human trials is a scientific priority. To that end, here we describe a functional genomics approach for examining the impact of gene depletion on cancer cell line spheroids, which more appropriately model human cancers in vivo.
要約
癌における機能ドライバイベントの同定は、新規な薬物標的の次の世代の発見のための癌生物学の我々の理解を促進する中心的かつ不可欠です。癌のより複雑なモデルは、in vivoで腫瘍形成を駆動し、臨床試験への前臨床モデルからの移行を行う新しい治療法の有効性を増加させる要因を完全に理解することが必要であることが明らかになってきています。
ここでは、siRNA機能的スクリーニングに供することができる均一で再現性の腫瘍スフェロイドを生成するための方法論を提示します。これらのスフェロイドは、従来の2次元培養物中に存在していない固形腫瘍で発見された多くの特徴を表示します。我々は、いくつかの一般的に使用される乳癌細胞株は、このプロトコルに適していることを示しています。さらに、我々はそれらを確認し、乳癌細胞系BT474を用いたプルーフの原理データを提供します上皮成長因子受容体HER2およびホスファチジルイノシトール-4,5-ビホスフェート3-キナーゼ(PIK3CA)の変異の増幅に依存する腫瘍スフェロイドとして増殖させました。最後に、我々はさらに、免疫組織化学を使用して、これらの依存関係の空間的影響を調査し、確認することができます。
概要
固形腫瘍は、正常患者を治療することができることで重要な課題で臨床医を提示する重要な組織学的、遺伝的およびマイクロ環境内腫瘍の不均一性を、表示します。小説を識別するために使用されるモデルの大部分は、治療は、これらの特徴の多くを組み込んでいないターゲット。確かに、診療所で利用電流標的療法は、二次元(2D)培養条件下で増殖させた癌細胞株に依存スクリーニングアプローチを採用し10年間で開発されてきました。このような受容体チロシンキナーゼ阻害剤として、様々な成功、もたらしたが、それは癌のより複雑なモデルが完全にin vivoで腫瘍形成を駆動してから移行する新しい治療法の数を増加させる要因を理解するために必要とされることが明らかになってきています臨床試験への前臨床モデル。また、現在ではよく2D培養システムがで反映しないことが理解されます行動1,2を生体内。例えば、不十分な血管新生化腫瘍では、微小環境内の酸素や栄養素の需要が供給を上回るように、ハイとローの配達の領域が開発しています。腫瘍における低酸素(低酸素症)の存在は、このような炭酸脱水酵素IX(CAIX)などの確立された低酸素マーカーのための腫瘍切片の免疫組織化学染色によって検出されるように、このようになどの機能を組み込んだ乳癌3,4で貧しい臨床転帰と相関しますスクリーニングモデルに低酸素症は、in vivoでより有効になる新規な薬物標的を発見するために私たちの能力を高めることができます。確かに、成功した低酸素症が含まれている攻撃的な腫瘍を標的とすることは、臨床的優先度5です。
従来の2DのsiRNAスクリーニングの改変は、より正確な腫瘍微小環境中の癌細胞が遭遇する条件の要素を再現する試みにおいて、いくつかの遺伝子番目の同定につながっていますインビボでの腫瘍増殖のために重要であることが見出されています。これらは、低血清条件6、低酸素状態7の下と組み合わせ8で行われ、機能ゲノムスクリーンを含みます。低血清中で増殖させた場合、例えば、6-Phosphofructo -2-キナーゼ/フルクトース-2,6-ビホスファターゼ4(PFKFB4)、炭素入る解糖の調節を担うタンパク質のサイレンシングは、唯一の転移に由来する前立腺癌細胞株においてアポトーシスを誘導しました。 PFKFB4の枯渇が完全に前立腺癌細胞株の異種移植片6の成長を切除、一方同じ条件下で、正常な前立腺細胞株におけるPFKFB4のサイレンシングは、効果がなかったです。
乳癌細胞株の延長パネルにおいて、モノカルボキシラーゼトランスポーター4(MCT4)のサイレンシングは、優先的に、低酸素条件下で細胞株の増殖の減少につながりました。この脆弱性は、乳癌細胞株直交異方性xenogra in vivoで検証しました。FTS。おそらく最も驚くべきことに、アセチル-CoAシンセターゼ2(ACSS2)、アセチル-CoAへの酢酸の変換に関与する酵素、栄養ストレス条件下で還元癌細胞数(低酸素および血清)のサイレンシングが、通常の培養条件の下でほとんどまたは全く効果がなかったです8。 ACSS2のアブレーションは、栄養勾配が腫瘍微小環境内で孤立して存在し、これらの地域に存在する細胞が腫瘍の進行8のために必須であることをしないことを示唆している乳癌および前立腺癌異種移植片の増殖に影響を与えました。また、ACSS2も腫瘍における増大ACSS2活性は不利な条件下での増殖をサポートする基本的なメカニズムであり得ることを示唆し、神経膠芽腫および肝細胞癌9,10において重要であることが見出されました。
集合的に、これらの研究は、 インビボで遭遇する条件を反復するとsiRNAのスクリーニングを行うことGの同定を可能にすることを証明します癌の生存に必須エン。同様に、栄養ストレス条件下で2D癌細胞の増殖に影響を与えるように、これらの研究における標的遺伝子の枯渇は、腫瘍異種移植片6,8で観察されたものをミラーリング、癌細胞株のスフェロイドの成長を阻害しました。したがって、癌細胞株のスフェロイドはACSS2サイレンシングに感受性を付与する腫瘍微小環境に遭遇する条件のいくつかを含有します。実際、スフェロイドは、栄養勾配(血清および酸素)、pHの変化、3次元(3D)細胞 - 細胞接触だけでなく、細胞周期停止およびアポトーシスを受けている細胞で増殖セレリティの変化を表示します。これは、壊死領域のがんスフェロイドにおける存在、従来の2D培養では見られない特徴が例示されます。
癌細胞のスフェロイドは既に小分子阻害剤をスクリーニングするために、より生物学的に関連するモデルとして使用されてきたが、これは唯一の化合物の有効性の検証やコンポの再利用を可能にしていますundsもともと他の疾患11のために設計します。現在の回転楕円体のスクリーニング方法は、高含量の方法のハイスループットで特定の遺伝子の枯渇の分析を可能にするものではありません。ここで、我々は、初めて癌細胞株のスフェロイドにおける低分子干渉RNA(siRNA)技術を利用し、特定の遺伝子の依存関係を解明するための機能ゲノミクスパイプラインを説明します。私たちは、人間の乳癌における200最も頻繁に変異遺伝子を標的とするsiRNAでオーダーメイドのライブラリを設計し、回転楕円体のサイズとBT474乳癌スフェロイドにおける代謝活性上の遺伝子の枯渇の影響を評価しました。我々は、堅牢かつ再現3D培養物ではサイレンシングERBB2およびPIK3CAの影響を検出することができました。さらに、我々は、免疫組織化学を使用して、BT474スフェロイドの空間的なアーキテクチャ上の遺伝子の枯渇の影響を評価することができます。
プロトコル
96ウェルのsiRNAプレートの調製
注:96ウェルプレートの外縁部は、このように96ウェルプレートあたり60のsiRNAの量を制限し、他のウェルと比較して蒸発しやすいです。これを制限するために、プレーン培地またはPBSで外側のウェルに記入します。また、ヒットの検証画面は、プレートバイアスを緩和することをお勧めします。
- 超低付着プレートの適切なウェルに500 ngの/(製造者の推奨に従って)低血清培地中でμL、アリコート10μLを - 250にsiRNAを希釈します。三連ですべての画面を実行します。
注:トランスフェクション効率を評価することができることを確認するために、プレートのデザインに適切な非標的(陰性)と(そのようなUBBとPLK1として、正)殺害のsiRNAコントロールを組み込みます。細胞は、トランスフェクション混合物に添加されるように、siRNAの作業のための低付着プレートを使用することは、重要です。私たちは、いくつかのメーカーは、超低付着することを除外することはできませんプレートは、スフェロイド成長に微妙な影響を与えることがあります。このように、我々はこれらがエンドユーザーによってテストされることをお勧めします。 - -20℃で接着シールとストアとカバープレート。
細胞株の2リバーストランスフェクション
- 画面の日に、室温でプレートを解凍し、卓上遠心機を用いて千×gで5分間スピン。各ウェルマルチチャンネルピペットを使用することに最適化されたトランスフェクション試薬を含む低血清培地の10μLを追加します。形成するためのsiRNAを含むトランスフェクション複合体のために15分間プレートを残します。
注:この研究は、機能的ゲノムスクリーニング(10%ウシ胎児血清(FBS)および抗生物質を補充した高グルコースDMEM(ダルベッコ改変イーグル培地)中で培養した)のための乳癌細胞株BT474を利用しました。 - スクリーニングされる細胞(0.05%トリプシン及び0.53mMのEDTA)は、フラスコから剥離されるまで、細胞株specifの適切な量を使用して中和をトリプシン処理ベンチトップ型遠心分離機上のIC媒体1,000×gで5分間スピンはトリプシンを削除します。媒体の適切な量で細胞を再懸濁し、細胞カウンターを使用して、正確な細胞数を決定します。
注:一般的にウェル当たり5000個の細胞は、試験したほとんどの細胞株全体のスフェロイド形成のために十分です。正確に正確なサイズの比較のための単一細胞懸濁液として細胞を計数することが重要です。 - 冷たい培地(4℃)で180μLあたり5,000個の細胞に細胞を希釈します。
注:再構成基底膜マトリックス成分の添加は、負にトランスフェクション効率に影響し、使用されていないことをお勧めします。細胞株の最適化は、スフェロイド形成能力を確認し、ノックダウン効果は、画面の前に行われるべきです。 - リザーバーに細胞懸濁液を移し、ピペットで混和し、siRNAが(2.1)以前用意含む96ウェル超低接着プレートの各ウェルに180μLを追加します。
- Centrifug電子その後10分間予冷4℃の遠心機で千×gでプレートし、37℃の組織培養インキュベーターに戻ります。
- 注:細胞がウェルの底にあるモザイクとして表示されます。次の12の上に - 24時間後、細胞は、単一の球を形成するために一緒に凝集します。
- 24時間後、細胞を観察するには、ウェルの中心における単一のスフェロイドを形成します。成長を促進するために、各ウェルに、完全培地100μLを追加します。
- 3日後に培地を補充します。静かに、各ウェルから培地100μLを除去し、新鮮な培地100μLを追加します。
- 7日目に、定量的に時間をかけてスフェロイドの成長を監視することができ、プレートリーダー上で自動化されたスフェロイドの大きさを定量化する(第3節を参照してください)。
- その後、(4章参照)発光細胞生存率色素を用いて細胞生存率を決定します。
3.自動画像取得
- ベンチトップ上でプレートをスキャンし、7日目のマイクロウェルプレートのイメージングサイトメーター。
- <、'96型ウェルプレート「適切なプレートタイプを選択して、実験名を入力します:LI>次の項目を選択し、ソフトウェアを開きます。注意:追加の情報は、ソフトウェアに追加することができます。
- 「Tumorsphere」アプリケーションを選択します。回転楕円体にフォーカスがあると、最適なコントラストを持つようにフォーカスを変更します。
注:私たちは、回転楕円体のサイズの大幅な変動が予想される」画像基づいてフォーカス」をお勧めします。 - スキャンを必要とする井戸を選択し、「スキャン開始」。
- プレートスキャナソフトウェアを使用して、オブジェクトマスクが正確に回転楕円体の大きさを表していることを確認してください。コロニー直径、国境の拡張、最小厚さと精度の設定を調整することでこれを行い、各細胞株に特異的には11,12をテストしました。
注記:回転楕円体の領域は、ソフトウェアアルゴリズムを用いて計算されます。例えば、正確に「高」に精度を調整する7日間増殖させたBT474スフェロイドの面積を計算しますdは200μMに最小コロニーの直径を設定します。これは、回転楕円体領域の適切なスフェロイドマスク代表を生成する必要があります。
注:これらのデータは、注釈付きデータファイルの形でで、エクスポート機能を使用して、マシンからエクスポートすることができます。日付は、プレートの中央値正規化され、組み合わされ、回転楕円体領域に統計的に有意な効果を持っていたsiRNAを同定するために分析のいずれかのZスコアまたは厳密に標準化平均差(SSMD)によってです。
- 「Tumorsphere」アプリケーションを選択します。回転楕円体にフォーカスがあると、最適なコントラストを持つようにフォーカスを変更します。
4.細胞生存率の決定
- 回転楕円体領域が計算された後、発光生存細胞の色素を用いて生存率を決定します。製造元の指示に従って試薬を準備します。
- 慎重に各ウェルから培地100μLを除去し、生存率染料の100μLを追加します。 15分間プレートをその後、発光プレートリーダーを使用してスキャンインキュベート。
注:これらのデータは、その後でで、エクスポート機能を使用して、マシンからエクスポートすることができます注釈付きデータファイルの形式。日付は、プレートの中央値正規化され、組み合わされ、回転楕円体生存率に統計的に有意な効果を持っていたsiRNAを同定するために分析のいずれかのZスコアまたは厳密に標準化平均差(SSMD)によってです。
結果
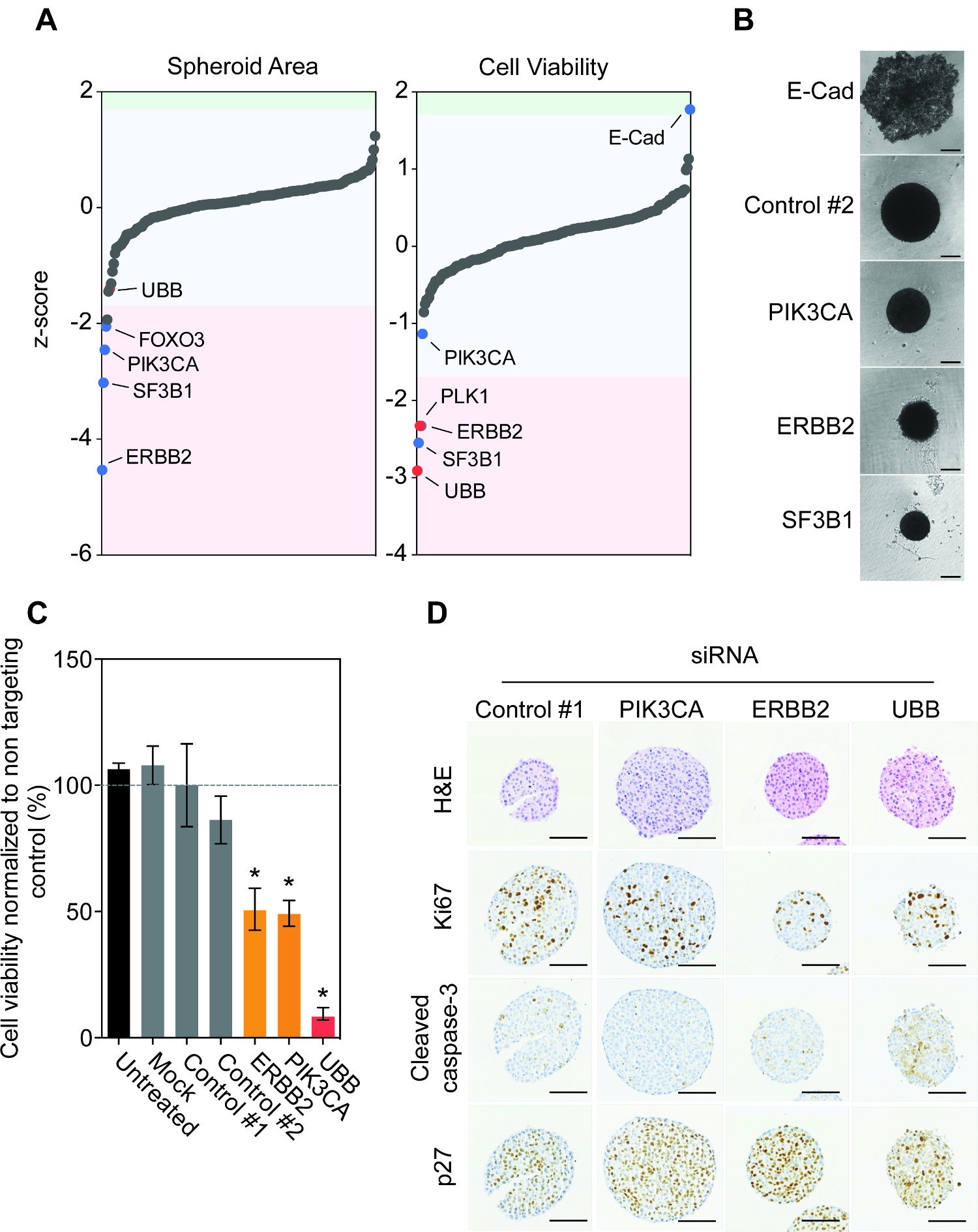
超低付着の96ウェルプレート中の回転楕円体アッセイはより容易にインビボで腫瘍に見られる生理学的条件を再現するコンテキストの潜在的な発癌性のためのハイスループット表現型の評価を提供します。実際、癌細胞株MCF10DCIS.comとBT474フォームタイトスフェロイド構造( 図1A)と回転楕円体のセクションの免疫組織学的調査は、細胞および核形態の明確な空間的な変化を示しました。時間が経つにつれて、このようなBT474スフェロイドのようないくつかのスフェロイドは、積極的な固形腫瘍( 図1B)の共通の特徴を壊死領域を開発しています。一部のスフェロイドは、MDA-MB-231細胞株などの壊死性コアを開発したが、逆に切断されたcaspsase-3の発現、アポトーシスのマーカー( 図1C)と相関増殖マーカーKi67を、で表示マークされ変動いたしません。多数の細胞株が実際レジェップあることを確立するためにsiRNA媒介遺伝子サイレンシングBT474の的は、MCF10DCIS.com、MDA-MB-231およびJIMT1細胞を逆7日間のsiRNAでトランスフェクトしました。試験した全ての細胞株( 図1D)に必須の遺伝子ユビキチンB(UBB)有意に減少スフェロイドの生存率のサイレンシングしながら、トランスフェクション(モック)試薬または対照siRNAのトランスフェクションの存在は、回転楕円体の生存能力に影響を及ぼしませんでした。
私たちは、選択されていない、ER +、HER2 +とトリプルネガティブ乳癌において最も頻繁に変異遺伝子を包含し、人間siRNAライブラリーを設計しました。このライブラリは、MYC、PIK3CAとTP53とその貢献発癌に確立されていないもののような既知の機能の遺伝子で構成されています。画面には、いくつかの非標的化対照siRNA(対照#1、コントロール#2)を含有し、siRNAは、殺害のコントロール( 表1)として機能し、このようなPLK1とUBBなどの必須遺伝子を、標的とします。我々は、乳房CANCを使用することにしましたそれらは容易に再構成基底膜を添加せずにスフェロイドを形成するような細胞株BT474、ER、馬車馬細胞株を確立し、公知のゲノムのアーキテクチャを有しています。例えば、BT474細胞はエストロゲン受容体(ER +)のために陽性である、ヒト上皮増殖因子受容体2(HER2 +)を過剰発現し、TP53(E285K)とPIK3CA(K111N)13に変異を保有します。
上記で概説したプロトコルを用いて、我々はインパクト遺伝子の枯渇は、siRNAをリバーストランスフェクション( 図1E)の7日後に回転楕円体の大きさ及び生存率に与えた監視しました。興味深いことに、遺伝子の大部分は、回転楕円体領域または生存能力( 図2A)に大きな影響を持っていませんでした。 FOXO3、PIK3CA、ERBB2およびSF3B1のサイレンシングは、回転楕円体のサイズの中で最も重要な再現性の減少をもたらしました。この減少はまた、ERBB2とSF3B1サイレンシング後に回転楕円体の生存率で観察されました。心強いことに、我々はCONF明視野顕微鏡( 図2B)を使用して、回転楕円体のサイズにサイレンシングPIK3CA、ERBB2およびSF3B1の影響をirmed。我々は以前、多数の細胞株モデルに必須の遺伝子としてSF3B1を同定し、したがってsiSF3B1はUBB 14に加えて、良好な殺害コントロールを表します。興味深いことに、E-カドヘリンのすべての200の遺伝子の唯一のサイレンシングは、回転楕円体の生存率( 図2A)の有意な増加をもたらしました。スフェロイド形態の調査は、E-カドヘリンのサイレンシングは、生存細胞が低接着ウェル( 図2B)の下で休んで、スフェロイドアーキテクチャの完全な崩壊をもたらしたことを示しました。スフェロイド容積画面データを手動で再調査は、これが原因で設定されたサイズ制限より上であるオブジェクトにも観察されたが、領域の定量化から排除されたことを示しました。以前に強調したように、BT474細胞は、受容体チロシンキナーゼHER2を過剰発現し、中の発癌性変異を抱きます PIK3CA(K111N)。我々は、非標的化コントロールのトランスフェクションは効果( 図2C)を有していなかったERBB2およびPIK3CAのサイレンシングは、減少した回転楕円体の生存率が得られたことを確認しました。
次に、回転楕円体組織学に対するsiRNA枯渇の影響を調べました。 BT474スフェロイドは、逆非標的コントロールsiRNAおよびsiRNAはPIK3CA、ERBB2およびUBBを標的にトランスフェクトしました。 ERBB2およびUBBのサイレンシングは、Ki67には、siRNAの( 図2D)を制御するために比較してプロ増殖マーカーの減少をもたらしました。プロアポトーシスマーカーの活性化は、カスパーゼ3のみHER2およびPIK3CAの枯渇は、アポトーシスにつながるのではなく、細胞毒性より細胞増殖抑制性であったなかったことを示唆している、UBBサイレンシング後に観察された切断されました。実際、HER2およびPIK3CAのサイレンシングがトランスフェクトされたスフェロイドを制御するために比較して、細胞周期停止タンパク質p27のタンパク質の発現の増加をもたらしました。
ENT」FO:キープtogether.withinページ= "1">まとめると、これらの結果は、BT474細胞を発癌性HER2およびPIK3CA 3Dスフェロイドとして増殖させた場合のシグナリングによって駆動されることを示すより重要なことに、これらの結果は、それが可能であることを示します。デザインと堅牢かつ再現癌細胞株スフェロイドにおける遺伝子の数百のオーダーメイドのsiRNAスクリーニングライブラリを実装しています。
図1:3 次元成長のための細胞株の最適化。 A.乳癌細胞株は、MCF10DCIS.com及びBT474は、7日間の低接着プレートで培養しました。明代表画像は、倒立顕微鏡を用いて撮影しました。スケールバー=100μmです。 B. MCF10DCIS.comとBT474スフェロイドは、28日間培養しました。 4日間 - 新鮮な培地を100μLごとに3補充しました。スフェロイドは、3.8%ホルムアルデヒド、(e)に固定し、mbedded、切断し、ヘマトキシリンおよびエオシン(H&E)で染色しました。代表的な画像は、低域と高倍率で示されています。スケールバーはそれぞれ、100ミクロンと33ミクロンを表します。 C. MDA-MB-231スフェロイドは、21日間培養しました。 4日間 - 新鮮な培地を100μLごとに3補充しました。スフェロイドを、3.8%ホルムアルデヒドで固定包埋、切片およびKi67および開裂カスパーゼ-3で染色しました。代表的な画像を示します。スケールバー=100μmです。 D. BT474、MCF10DCIS.com、MDA-MB-231、およびJIMT1細胞株が逆モック(トランスフェクション試薬のみ)、および対照siRNAとUBBでトランスフェクトした、超低付着プレートはその後スフェロイドを形成するように回転させました。新鮮な培地(100μl)を、細胞生存率を定量化した、7日後の日1と4に追加されました。データは、#1を制御するために正規化三重で行わ二つの独立した生物学的複製の平均±SDを表します。統計的有意性はUNPAを用いて計算しましたIREDスチューデントt検定(p <0.05)。 E.機能的に癌細胞株スフェロイドにおける遺伝子依存性を調べるために使用される逆トランスフェクションプロトコールを要約する流れ図。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}

図2:BT474 スフェロイド暴く発癌の依存関係のゲノム機能研究。 A. BT474細胞は、逆三連で200個の遺伝子のヒトsiGENOME siRNAライブラリーを用いてトランスフェクトしました。回転楕円体のサイズと生存率を観察しました。生データ値は、プレートの中央値正規化z-スコアが1.7倍を超える有意なアウトライアー15メジアンプレートの標準偏差を特定するために計算されたました。辺境の遺伝子ERBB2、SF3B1、PLK1とUBB aは注意してくださいndはE-CAD。大幅に回転楕円体の領域及び生存率を増加または減少したsiRNAは、赤は対照siRNAのを示している場合には、青と赤で網掛けされています。 B.細胞は、逆非標的制御、E-カドヘリン(E-CAD)、PIK3CA、ERBB2またはUBB siRNAをトランスフェクトした後、スフェロイドを形成するために、超低取付板にスピンしました。 7日後、代表的な回転楕円体の画像は、倒立顕微鏡を用いて撮影した明視野。スケールバー=100μmです。 C.細胞は、逆非標的制御、PIK3CA、ERBB2またはUBB siRNAをトランスフェクトした後、スフェロイドを形成するために、超低取付板にスピンしました。 7日後、細胞生存率を定量しました。データは、#1を制御するために正規化三重で行わ二つの独立した生物学的複製の平均±SDを表します。統計的有意性は、不対スチューデントt検定(p <0.05)を用いて計算しました。 D. 7日後に、スフェロイドは、固定された埋め込み、切片にし、染色しましたH&E、Ki67に、切断されたカスパーゼ3、およびp27のため。代表的な画像を示します。スケールバー=100μmです。 siControlスフェロイドのH&Eが原因で処理し、染色のために選択された個々の無傷のスフェロイドのアーチファクトに小さく表示されます注意してください。しかし、これはスキャンした画像と細胞生存率の結果を用いて観察される変化を損ないません。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| 1の非標的 | TP53 | DST | KMT2C | 未処理の1 | FCGBP | ARID1B | FBXM7 | TTC40 | 2を非標的 | ||
| GATA3 | TTN | MUC12 | MUC4 | AHNAK | HUWEI1 | DNAH11 | ITPR2 | ABCA13 | CREBBP | ||
| MAP2K4 | PIK3CA | F5 | APOB | ANKRD30A | MUC17 | DNAH17 | LAMA2 | エース | CSMD2 | ||
| STARD9 | USH2A | FAT3 | LPR2 | CSMD3 | MYO18B | DNAH5 | MDN1 | ARHGAP5 | DNAH9 | ||
| CXCR3 | MUC16 | RB1 | PKHD1L1 | DNAH2 | SYNE2 | DYNC1H1 | PCLO | CACNA1B | ERBB2 | ||
| PLK1 | SYNE1 | LYST | PTEN | SPTA1 | 未処理の2 | VHL | RYR1 | COL6A3 | UBB | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| 1の非標的 | ENAM | RYR2 | BRCA2 | 未処理の1 | FMN2 | HECW1 | LAMB4 | SI | 2を非標的 | ||
| FHOD3 | MACF1 | RYR3 | C2ORF16 | DMD | FRG1 | HERC2 | MYH11 | STAB1 | ZDBF2 | ||
| GOLGA6L2 | NEB | SMG1 | CACNA1E | DNA14 | GCC2 | HIVEP2 | NIPBL | TANC1 | ZNF536 | ||
| HMCN1 | NF1 | UBR5 | CACNA1F | DYNC2H1 | GON4L | HYDIN | PKD1L1 | TF | ANK3 | ||
| HRNR | OBSCN | USP34 | CYMA5 | FAM208B | GPR112 | ITSN2 | RNF213 | TPR | ASPM | ||
| PLK1 | PCDH15 | XIRP2 | COL7A1 | FLG2 | 未処理の2 | VHL | SAGE1 | UNC80 | UBB | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| 1の非標的 | DCHS2 | MUC5B | ZFHX4 | 未処理の1 | MYO9A | SPHKAP | CXORF22 | NCOR1 | VPS13D | ||
| ATR | DMXL2 | MXRA5 | ANK2 | KIAA1210 | PRUNE2 | TCHH | DANH6 | NOTCH2 | ANKRD12 | ||
| BIRC6 | DNAH10 | TENM1 | ATM | LRP1 | SCN10A | VPS13C | DNAH7 | SPEN | C5ORF42 | ||
| CDH1 | DNAH3 | PEG3 | DIDO1 | MAP1A | SCN2A | IPTR3 | ERBB3 | SRRM2 | CCDC88A | ||
| CUBN | NOCK11 | RELN | DNAH8 | MED12 | SHROOM2 | CEP350 | FAT4 | SZT2 | CHD4 | ||
| PLK1 | EYS | SACS | KIAA1109 | MED13 | 未処理の2 | VHL | KMT2A | VPS13A | UBB | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
| 非標的 | QSER1 | ARID1A | WDFY3 | 未処理の1 | SDK1 | TEX15 | LAMA1 | 2を非標的 | |||
| COL14A1 | SHROOM3 | ATRX | EFCAB5 | SF3B1 | CBFB | AHNAK2 | |||||
| CSMD1 | TBX3 | KIAA0947 | FOXA1 | ITPR1 | DDX3X | KIF4A | |||||
| MEFV | UBR4 | MYCBP2 | INPPL1 | FLG | HECTD4 | FAT2 | |||||
| MGAM | VCAN | NBEAL1 | MAP3K1 | AKAP9 | GPR98 | FOXO3 | |||||
| PLK1 | ZNF462 | SETX | NRP1 | HERC1 | 未処理の2 | VHL | UBB | ||||
表1:プレートレイアウトと人間のsiRNAライブラリデコンボリューション。表には、画面で使用される低接着プレート用のsiRNAプールとレイアウトの各コンテンツが含まれています。
ディスカッション
癌の三次元モデルは、ますます選択的に癌細胞を殺すために設計されている既知の及び新規な化合物の有効性を評価するために使用されています。癌細胞スフェロイドは、このように、3Dでの有効性を増加している化合物は、インビボでの効果を有する可能性がより高いインビボでの腫瘍において遭遇するものと類似の条件を表示する構造です。しかし、これらのモダリティは、癌を治療する際にかなりの効果を持つことができる薬物設計の対象とされていない潜在的な新規標的の同定を可能にするものではありません。
私たちは、癌細胞株スフェロイドで最大7日間耐久性の遺伝子サイレンシングのために許可されたsiRNA機能ゲノミクスのアプローチを開発しました。 siRNAスクリーニングを行うことができる前に、最適化を必要とするプロトコルには、いくつかの重要なステップがあります。大規模で再現可能な実行可能な球状体を形成する能力が不可欠です。また、ppropriateトランスフェクション条件を厳密に最適化されるべきです。我々は、適切な非標的といくつかの異なるトランスフェクション試薬を試用し、画面をしようとする前にコントロールを殺すことをお勧めします。我々は、いくつかの一般的に使用される乳癌細胞株、すなわちBT474、MCF10DCIS.com、MDA-MB-231及びJIMT1がsiRNAトランスフェクションに適したことを示すことができました。さらに、我々はBT474のスフェロイドにおける乳癌における200最も頻繁に変異遺伝子をスクリーニングする実証の原則データを提供する、HER2の増幅およびPIK3CAの発癌性変異に自分の依存関係を確認しました。興味深いことに、転写因子FOXO3のサイレンシングは、回転楕円体のサイズの縮小が、生存率に有意な影響をもたらしました。 FOXO3は、彼らの環境16をより容易に適応できるように、癌細胞の代謝能力を変化させる、低酸素状態に対する応答を調節することが知られています。それはATPの存在量を検出すると、この役割は、潜在的に細胞生存率の読み取りを妨げる可能性、細胞代謝の主な製品の一つ。
スフェロイド小型化の観察を支持し、HeLa細胞におけるFOXO3のサイレンシングは、腫瘍増殖を障害およびアポトーシス17を誘導した異種移植片が示されています。いくつかの遺伝子は、偽陽性をもたらす可能性がそれらの3D構造を、保持する癌細胞の能力に影響を与える可能性があることに留意することが重要です。例えば、E-カドヘリンのノックダウンは、BT474スフェロイド構造の溶解をもたらしました。これは、以前E-カドヘリンは、抗体18をターゲットに用いて報告されていました。任意のスクリーニングプラットフォームと同様に、潜在的なターゲットは、観察される効果の再現性を評価するために再スクリーニングされるべきです。技術、siRNA媒介遺伝子ノックダウンの、すなわち一時的な性質には制限があります。サイレンシング持続7日間より長くは、siRNAを用いて達成可能ではありませんでした。
このアプローチの利点は、様々な他の生物群系と結合することができるということですTRIC染料だけでなく、回転楕円体の低酸素症の空間的な情報を与えるか、アポトーシスを起こしている細胞を監視する、例えば、回転楕円体の生存率を評価するもの。プレートリーダーのスキャンは、比較的迅速かつ非侵襲性であるため、さらに、回転楕円体のサイズにsiRNAの影響は、時間の経過だけでなく、実験終了時点で評価することができます。確かに、我々は現在、我々のスクリーニングパイプライン内でこれらの道のいくつかを模索しています。新規の依存関係を識別するために、3D培養物を利用する別のアプローチは、標的またはタンパク質の特定のファミリーの広い範囲のいずれかを阻害する化学ライブラリーの使用です。実際、Bitler ら 。卵巣明細胞癌19でARID1A状態とEZH2阻害剤との間の合成致死相互作用を識別するために、このターゲットにアプローチを利用しました。 CRISPR-Cas9遺伝子編集技術の発見もオルガノイド培養およびin vivoでの遺伝子スクリーニングの開発を可能にしました。しかし、トン彼のアプローチは、適切な動物施設を有するに依存し、20法外な費用がかかることもできます。
結論として、我々は新たな癌標的または確立されたターゲットの強固な検証の識別を可能にすることが、より正確にモデル化酸素およびin vivoでの腫瘍の微小環境の特徴である栄養勾配、プロトコルを概説していると信じています。さらに、我々のプロトコルは、スフェロイドを形成し、従って日常ハイスループットsiRNAのスクリーニングのための癌研究コミュニティで使用することができる細胞系の任意のタイプに適用することができます。
開示事項
Open Access fees were supported by Nexcelom Bioscience, LLC.
謝辞
The authors would like to thank Fredrik Walberg (FACS and imaging facility, ICR) for assistance with microscopy. This work was funded by Breast Cancer Now. RN is the recipient of a Breast Cancer Now Career Development Fellowship (2011MaySF01).
資料
| Name | Company | Catalog Number | Comments |
| Lullaby | Oz Biosciences | LL70500 | lipid-based transfection reagent |
| Viromer | Lipocalyx | VB-01LB-01 | virus-like polymer transfection reagent |
| Ultra-low attachment plate | Corning | CLS7007 | 96 well plate |
| Foil plate seals | ThermoFisher | AB-0626 | |
| Luminescent cell viability dye | Promega | G7570 | CellTitre-Glo |
| Pipette tips (200 μL) | Starlab | S1111-0806 | |
| Pipette tips (10 μL) | Starlab | S1111-3800 | |
| Pipette tips (1, 000 μL) | Starlab | S1122-1830 | |
| Serological pipettes (5 mL) | Sarstedt | 86.1253.025 | |
| Serological pipettes (10 mL) | Sarstedt | 86.1254.025 | |
| Serological pipettes (25 mL) | Sarstedt | 86.1685.020 | |
| RPMI Media | GIBCO | 11875-093 | |
| DMEM Media | GIBCO | 11965-084 | |
| Opti-MEM | GIBCO | 31985070 | |
| Feta bovine serum | GIBCO | 16140063 | |
| siRNA | Dharmacon | Cherry picked library | |
| Countess Cell Counter | ThermoFisher Scientific | AMQAX1000 | |
| Cell counting chamber slides | ThermoFisher Scientific | C10312 | |
| Celigo S | Nexcelom | contact company | |
| Victor X5 | Perkin Elmer | contact company | |
| Benchtop centrifuge | Various | ||
| Axiovert Inverted brightfield microscope | Zeiss | contact company | |
| Tissue culture CO2; Incubator | Various | ||
| Mulitichannel pipette | Various |
参考文献
- Seton-Rogers, S. E., et al. Cooperation of the ErbB2 receptor and transforming growth factor beta in induction of migration and invasion in mammary epithelial cells. Proc Natl Acad Sci U S A. 101 (5), 1257-1262 (2004).
- Debnath, J., Muthuswamy, S. K., Brugge, J. S. Morphogenesis and oncogenesis of MCF-10A mammary epithelial acini grown in three-dimensional basement membrane cultures. Methods. 30 (3), 256-268 (2003).
- Chia, S. K., et al. Prognostic significance of a novel hypoxia-regulated marker, carbonic anhydrase IX, in invasive breast carcinoma. J Clin Oncol. 19 (16), 3660-3668 (2001).
- Trastour, C., et al. HIF-1alpha and CA IX staining in invasive breast carcinomas: prognosis and treatment outcome. Int J Cancer. 120 (7), 1451-1458 (2007).
- Wilson, W. R., Hay, M. P. Targeting hypoxia in cancer therapy. Nat Rev Cancer. 11 (6), 393-410 (2011).
- Ros, S., et al. Functional metabolic screen identifies 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 4 as an important regulator of prostate cancer cell survival. Cancer Discov. 2 (4), 328-343 (2012).
- Baenke, F., et al. Functional screening identifies MCT4 as a key regulator of breast cancer cell metabolism and survival. J Pathol. 237 (2), 152-165 (2015).
- Schug, Z. T., et al. Acetyl-CoA synthetase 2 promotes acetate utilization and maintains cancer cell growth under metabolic stress. Cancer Cell. 27 (1), 57-71 (2015).
- Mashimo, T., et al. Acetate is a bioenergetic substrate for human glioblastoma and brain metastases. Cell. 159 (7), 1603-1614 (2014).
- Comerford, S. A., et al. Acetate dependence of tumors. Cell. 159 (7), 1591-1602 (2014).
- Vinci, M., Box, C., Eccles, S. A. Three-dimensional (3D) tumor spheroid invasion assay. J Vis Exp. (99), e52686 (2015).
- Vinci, M., Box, C., Zimmermann, M., Eccles, S. A. Tumor spheroid-based migration assays for evaluation of therapeutic agents. Methods Mol Biol. 986, 253-266 (2013).
- Barretina, J., et al. The Cancer Cell Line Encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature. 483 (7391), 603-607 (2012).
- Maguire, S. L., et al. SF3B1 mutations constitute a novel therapeutic target in breast cancer. J Pathol. 235 (4), 571-580 (2015).
- Brough, R., et al. Functional viability profiles of breast cancer. Cancer Discov. 1 (3), 260-273 (2011).
- Ferber, E. C., et al. FOXO3a regulates reactive oxygen metabolism by inhibiting mitochondrial gene expression. Cell Death Differ. 19 (6), 968-979 (2012).
- Jensen, K. S., et al. FoxO3A promotes metabolic adaptation to hypoxia by antagonizing Myc function. EMBO J. 30 (22), 4554-4570 (2011).
- Ivascu, A., Kubbies, M. Diversity of cell-mediated adhesions in breast cancer spheroids. Int J Oncol. 31 (6), 1403-1413 (2007).
- Bitler, B. G., et al. Synthetic lethality by targeting EZH2 methyltransferase activity in ARID1A-mutated cancers. Nat Med. 21 (3), 231-238 (2015).
- Dow, L. E., et al. Inducible in vivo genome editing with CRISPR-Cas9. Nat Biotechnol. 33 (4), 390-394 (2015).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved