
Method Article
Utilisant la génomique fonctionnelle de dépistage pour identifier les cibles de médicaments potentiellement nouveaux dans Cancer Cell Cultures Spheroid
Dans cet article
Résumé
Identifying novel drug targets that transition from pre-clinical testing to human trials is a scientific priority. To that end, here we describe a functional genomics approach for examining the impact of gene depletion on cancer cell line spheroids, which more appropriately model human cancers in vivo.
Résumé
L'identification des événements de pilotes fonctionnels dans le cancer est au cœur de favoriser notre compréhension de la biologie du cancer et indispensable pour la découverte de la prochaine génération de nouvelles cibles thérapeutiques. Il devient évident que les modèles plus complexes de cancer sont nécessaires pour apprécier pleinement les facteurs qui stimulent la tumorigenèse in vivo et d' augmenter l'efficacité de nouvelles thérapies qui font la transition à partir de modèles pré-cliniques pour les essais cliniques.
Nous présentons ici une méthode pour générer des sphéroïdes tumoraux uniformes et reproductibles qui peuvent être soumis à un criblage fonctionnel siRNA. Ces sphéroïdes présentent de nombreuses caractéristiques que l'on trouve dans les tumeurs solides qui ne sont pas présentes dans la culture à deux dimensions traditionnelles. Nous montrons que plusieurs lignées de cellules de cancer du sein couramment utilisées se prêtent à ce protocole. En outre, nous fournissons la preuve de principe des données utilisant le cancer du sein lignée cellulaire BT474, confirmant leurdépendant de l'amplification du récepteur du facteur de croissance épidermique HER2 et la mutation de la phosphatidylinositol-4,5-biphosphate 3-kinase (PIK3CA) lorsqu'elles sont cultivées sous forme de sphéroïdes de tumeur. Enfin, nous sommes en mesure d'étudier plus avant et de confirmer l'impact spatial de ces dépendances en utilisant immunohistochimie.
Introduction
Les tumeurs solides affichent histologique significative, l'hétérogénéité intra-tumorale génétique et micro-environnement, qui présente aux cliniciens un défi important d'être en mesure de traiter les patients avec succès. La majorité des modèles utilisés pour identifier de nouvelles thérapies ciblées ne comportent pas beaucoup de ces caractéristiques. En effet, les thérapies ciblées actuelles utilisées en clinique ont été développés dans la dernière décennie, en utilisant des approches de criblage qui reposent sur des lignées de cellules cancéreuses cultivées dans des conditions bidimensionnelles (2D) de la culture. Bien que cela ait provoqué des succès divers, tels que les inhibiteurs du récepteur tyrosine kinase, il devient évident que les modèles plus complexes de cancer sont nécessaires pour apprécier pleinement les facteurs qui stimulent la tumorigenèse in vivo et d' augmenter le nombre de nouvelles thérapies qui font la transition de modèles précliniques aux essais cliniques. De plus, il est maintenant bien apprécié que les systèmes de culture 2D ne reflètent envivo comportement 1,2. Par exemple, dans les tumeurs vascularisées mal, car la demande d'oxygène et de nutriments dans le microenvironnement supérieure à l'offre, les régions de livraisons hautes et basses se développent. La présence de faible teneur en oxygène (hypoxie) dans les tumeurs, telles que détectées par la coloration immunohistochimique de coupes de tumeur pour les marqueurs de hypoxiques établis tels que l' anhydrase carbonique IX (CAIX), est en corrélation avec un résultat clinique plus faible dans le cancer du sein 3,4 Ainsi , les caractéristiques incorporant tel que hypoxie dans les modèles de dépistage peut améliorer notre capacité à découvrir de nouvelles cibles thérapeutiques qui seront plus efficaces in vivo. En effet, ciblant avec succès des tumeurs agressives qui contiennent l' hypoxie est une priorité clinique 5.
La modification de criblage traditionnelle 2D ARNsi, dans une tentative pour récapituler avec plus de précision les éléments des conditions rencontrées par les cellules cancéreuses dans le microenvironnement tumoral, a conduit à l'identification de plusieurs gènes eà se sont révélés être importants pour la croissance tumorale in vivo. Ceux - ci comprennent des écrans de génomique fonctionnelle effectuées dans des conditions de sérum bas 6, des conditions hypoxiques 7 et 8 en combinaison. Par exemple, le silençage de 6-phosphofructo-2-kinase / fructose-2,6-biphosphatase 4 (PFKFB4), une protéine responsable de la régulation du carbone glycolyse entrant, seul induit l'apoptose dans les lignées de cancer de la prostate dérivées de métastases lorsqu'il est cultivé dans un faible taux sérique. Silencing de PFKFB4 dans des lignées cellulaires de la prostate normales dans les mêmes conditions n'a eu aucun effet, alors que, l' épuisement des PFKFB4 complètement ablatée la croissance de la prostate lignée cellulaire de cancer xénogreffes 6.
Dans un panneau d'extension de lignées cellulaires de cancer du sein, le silençage de mono-carboxylase transporteur 4 (MCT4), de préférence conduit à la réduction de la croissance de la lignée cellulaire dans des conditions de faible teneur en oxygène. Cette vulnérabilité a été validée in vivo dans le cancer du sein lignée cellulaire orthotrope xenografts. Peut-être le plus frappant, réduisant au silence de l'acétyl-CoA synthétase 2 (ACSS2), une enzyme responsable de la conversion de l'acétate en acétyl-CoA, réduit le cancer nombre de cellules dans des conditions de stress en nutriments (faible teneur en oxygène et le sérum), mais a eu peu ou pas d'effet dans des conditions normales de culture 8. Ablation de ACSS2 impacté la croissance du cancer du sein et de la prostate xénogreffes suggérant que les gradients de nutriments ne sont pas isolés dans le microenvironnement de la tumeur et que les cellules qui se trouvent dans ces régions sont essentielles à la progression tumorale 8. En outre, ACSS2 a également été jugée importante dans le glioblastome et le carcinome hépatocellulaire 9,10, ce qui suggère que l' augmentation de l' activité de ACSS2 dans des tumeurs pourrait être un mécanisme essentiel qui soutient la croissance , dans des conditions défavorables.
Collectivement, ces études démontrent que récapitulant les conditions rencontrées in vivo et la réalisation d' écrans d'ARNsi permet l'identification de gènes essentiels pour la survie du cancer. Ainsi que d'un impact sur la croissance des cellules cancéreuses en 2D dans des conditions de stress en nutriments, l' épuisement des gènes cibles dans ces études ont inhibé la croissance de la sphéroïde lignée cellulaire de cancer, ce qui reflète ce qui a été observé dans des xénogreffes de tumeur à 6,8. Ainsi, les sphéroïdes de lignées cellulaires cancéreuses contiennent plusieurs des conditions rencontrées dans le microenvironnement tumoral qui confèrent une sensibilité à ACSS2 silençage. En effet, les sphéroïdes présentent des gradients de nutriments et d'oxygène (sérum), des variations de pH, en trois dimensions (3D) de contact cellule-cellule, mais aussi, des altérations dans la célérité proliférative des cellules subissant une apoptose et un arrêt du cycle cellulaire. Ceci est illustré par la présence d'un cancer dans sphéroïdes des régions nécrotiques, une caractéristique non trouvée dans la culture traditionnelle 2D.
sphéroïdes de cellules cancéreuses ont déjà été utilisés comme modèles biologiquement plus pertinents pour cribler des inhibiteurs de petites molécules, mais cela ne permet la validation de l'efficacité du composé ou du reciblage des compounds conception à l' origine pour d' autres maladies 11. méthodes de dépistage sphéroïde actuelles ne permettent pas pour l'analyse de l'épuisement du gène spécifique dans un haut débit de manière à haute teneur. Ici, nous décrivons pour la première fois, un pipeline de génomique fonctionnelle pour découvrir les dépendances de gènes spécifiques en utilisant des petits ARN interférents (siRNA) la technologie dans le cancer sphéroïdes de lignées cellulaires. Nous avons conçu une bibliothèque sur mesure avec siRNA ciblant les 200 gènes les plus fréquemment mutés dans les cancers du sein humains et évalué l'impact de l'épuisement du gène de la taille sphéroïde et l'activité métabolique dans BT474 sphéroïdes de cancer du sein. Nous avons pu détecter robuste et reproductible l'impact de ERBB2 et PIK3CA taire dans les cultures 3D. De plus, nous avons pu évaluer l'impact de l'épuisement des gènes sur l'architecture spatiale de sphéroïdes BT474 utilisant immunohistochimie.
Protocole
1. Préparation de 96 puits siARN plaques
NOTE: Les bords extérieurs d'une plaque de 96 puits sont plus sujettes à l'évaporation par rapport à d'autres puits, limiter ainsi la quantité de siRNA à 60 plaque à 96 puits. Remplissez dans les puits extérieurs avec un milieu ordinaire ou PBS pour limiter cela. En outre, un écran de validation des résultats est recommandée pour atténuer les biais des plaques.
- Diluer les ARNsi à 250 - 500 ng / ul dans un milieu de sérum réduit (selon les recommandations du fabricant) et aliquote de 10 ul dans les puits appropriés de la plaque de fixation ultra-faible. Effectuer tous les écrans en triple.
NOTE: Incorporer non-ciblage (négatif) et de tuer (positif, comme UBB et PLK1) contrôles de siRNA appropriées dans la conception de la plaque pour assurer l'efficacité de transfection peut être évaluée. L'utilisation de plaques à faible fixation pour le travail de l'ARNsi est critique, car les cellules seront ajoutés au mélange de transfection. Nous ne pouvons pas exclure que certains fabricants ultra-faible attachementplaques peuvent avoir des effets subtils sur la croissance sphéroïde. En tant que tel, nous recommandons que ceux-ci sont testés par l'utilisateur final. - Des plaques de recouvrement avec des joints adhésifs et conserver à -20 ° C.
2. inverse Transfection de la lignée cellulaire
- Le jour de l'écran, dégivrer les plaques à la température ambiante et tourner pendant 5 min à 1000 xg en utilisant une centrifugeuse de paillasse. Ajouter 10 ul de milieu sérique réduit contenant le réactif de transfection optimale dans chaque puits en utilisant une pipette à canaux multiples. Laisser les plaques pendant 15 min pour les complexes de transfection contenant siRNA pour former.
NOTE: Cette étude a utilisé la ligne du cancer du sein de cellules BT474 pour le criblage génomique fonctionnelle (cultivées en haute DMEM de glucose (milieu de Eagle modifié par Dulbecco) supplémenté avec 10% de sérum de veau fœtal (FBS) et des antibiotiques). - Trypsiniser les cellules à cribler (0,05% de trypsine et d'EDTA 0,53 mM) jusqu'à ce qu'ils se détachent de la fiole, à neutraliser en utilisant un volume approprié de la lignée cellulaire spécifmoyen ic et de spin pendant 5 min à 1000 xg dans une centrifugeuse de paillasse pour éliminer la trypsine. Resuspendre les cellules avec un volume approprié de milieu et de déterminer le nombre de cellules précis en utilisant un compteur de cellules.
NOTE: Typiquement 5.000 cellules par puits sont suffisantes pour la formation sphéroïde dans la plupart des lignées cellulaires testées. Il est important de compter avec précision les cellules en suspension de cellules individuelles pour des comparaisons précises de taille. - Diluer les cellules à 5.000 cellules par 180 ul dans un milieu froid (4 ° C).
NOTE: L'ajout de reconstitués membrane basale composants de la matrice aura un impact négatif efficacité de la transfection et il est recommandé qu'il ne soit pas utilisé. l'optimisation de la ligne cellulaire pour confirmer la capacité de sphéroïde formant et l'efficacité knockdown doit être effectué avant l'écran. - Transférer la suspension cellulaire à un réservoir, une pipette pour mélanger et ajouter 180 uL à chaque puits de la plaque de fixation 96 puits ultra-faible contenant l'ARNsi préparé précédemment (2,1).
- Centrifuge la plaque à 1000 x g dans une pré-refroidi à 4 ° C centrifuger pendant 10 min, puis retour à une culture incubateur à 37 ° C tissulaire.
- REMARQUE: Les cellules apparaissent sous la forme d'une mosaïque dans le fond des puits. Au cours des prochaines 12-24 h les cellules vont regrouper ensemble pour former une seule sphère.
- Après 24 h, observer les cellules forment une seule sphéroïde dans le centre du puits. Ajouter 100 ul de milieu complet à chaque puits pour favoriser la croissance.
- Reconstituer milieu après trois jours. Retirez délicatement 100 pi de milieu de chaque puits et ajouter 100 pi de milieu frais.
- Au jour 7, de quantifier la taille sphéroïde automatisée sur un lecteur de plaques qui peuvent surveiller la croissance quantitative sphéroïde au fil du temps (voir la section 3).
- Ensuite, déterminer la viabilité des cellules en utilisant un colorant de viabilité cellulaire luminescent (voir la section 4).
3. Automated Image Acquisition
- Balayez les plaques sur un banc-top, micro-puits plaque imagerie cytomètre le jour 7.
- Ouvrez le logiciel, sélectionnez les éléments suivants: la plaque '96 ', sélectionnez le type de plaque approprié et entrez un nom d'expérience. REMARQUE: Toute information supplémentaire peut également être ajouté dans le logiciel.
- Sélectionnez l'application 'Tumorsphere'. Modifier la mise au point de telle sorte que le sphéroïde est mise au point et a un contraste optimal.
NOTE: Nous vous recommandons «foyer image sur la base" comme des variations importantes dans les tailles sphéroïde sont attendus. - Sélectionnez les puits qui nécessitent la numérisation et «scan Start '.
- Utilisation du logiciel du scanner de plaque, assurez-vous que le masque d'objet représente avec précision la taille de sphéroïde. Faites ceci en ajustant les paramètres diamètre de la colonie, la dilatation des frontières, l' épaisseur minimum et de précision, spécifiques à chaque lignée cellulaire testée 11,12.
NOTE: La zone sphéroïde sera alors calculée en utilisant l'algorithme du logiciel. Par exemple, pour calculer avec précision la zone de sphéroïdes BT474 cultivées pendant sept jours d'ajuster la précision de 'high' und régler le diamètre des colonies minimum à 200 uM. Cela devrait produire un représentant du masque sphéroïde appropriée de la zone sphéroïde.
NOTE: Ces données peuvent ensuite être exportées à partir de la machine, en utilisant la fonction d'exportation, sous la forme d'un fichier de données annotée. La date est la plaque médiane normalisée, combinées et analysées soit par z-score ou différence moyenne strictement standardisée (DMES) pour identifier les ARNsi qui ont eu un effet statistiquement significatif sur la zone sphéroïde.
4. Détermination de la viabilité cellulaire
- Après la zone sphéroïde a été calculée, déterminer la viabilité en utilisant un colorant cellulaire de viabilité luminescent. Préparer le réactif selon les instructions du fabricant.
- Retirez délicatement 100 pi de milieu de chaque puits et ajouter 100 pi de viabilité colorant. Incuber les plaques pendant 15 min numériser puis en utilisant un lecteur de plaque luminescente.
NOTE: Ces données peuvent ensuite être exportées à partir de la machine, en utilisant la fonction d'exportation, dansla forme d'un fichier de données annotée. La date est la plaque médiane normalisée, combinées et analysées soit par z-score ou différence moyenne strictement standardisée (DMES) pour identifier les ARNsi qui ont eu un effet statistiquement significatif sur la viabilité sphéroïde.
Résultats
Essais sphéroïde fixation des plaques ultra-basses 96 puits fournissent une évaluation phénotypique à haut débit pour oncogenicity potentiel dans un contexte qui récapitule plus facilement les conditions physiologiques trouvés dans les tumeurs in vivo. En effet, les lignées cellulaires du cancer MCF10DCIS.com et forment BT474 serrés les structures sphéroïdales (figure 1A) et d' investigation immunohistochimique de sections sphéroïdes ont montré des changements spatiaux distincts dans la morphologie cellulaire et nucléaire. Au fil du temps, certains sphéroïdes tels que sphéroïdes BT474 développent des régions nécrotiques, une caractéristique commune des tumeurs solides agressives (figure 1B). Certains sphéroïdes ne développent pas de noyaux nécrotiques, tels que la lignée de cellules MDA-MB-231, mais font affichage variation marquée dans le marqueur de prolifération Ki67, qui est en corrélation inverse avec clivée caspsase-3 expression, un marqueur de l' apoptose (figure 1C). Pour établir que les lignées cellulaires multiples sont en effet receptive à l'ARNsi à médiation par gene silencing BT474, MCF10DCIS.com, MDA-MB-231 et les cellules ont été JIMT1 inverse transfectées avec l'ARNsi pendant sept jours. La présence de réactif de transfection (mock) ou la transfection de siARN de contrôle n'a eu aucun effet sur la viabilité sphéroïde, tout en réduisant au silence le gène essentiel Ubiquitin B (UBB) réduit de manière significative la viabilité des sphéroïde dans toutes les lignées cellulaires testées (Figure 1D).
Nous avons conçu une bibliothèque humaine siRNA qui englobait les gènes les plus fréquemment mutés dans unselected, ER +, HER2 + et les cancers du sein triple négatif. Cette bibliothèque se compose de gènes de fonction connue, tels que MYC, PIK3CA et TP53 et ceux dont la contribution à la cancérogenèse est pas établie. L'écran contient également plusieurs ciblant non siRNA de contrôle (contrôle n ° 1, de commande n ° 2) et des siRNA ciblant des gènes essentiels, tels que PLK1 et UBB, qui agissent comme tuer des contrôles (tableau 1). Nous avons choisi d'utiliser le canc du seiner les lignées cellulaires BT474 car ils forment facilement des sphéroïdes sans l'addition d'une membrane basale reconstituée, sont des lignées cellulaires établies cheval de bataille et ont une architecture génomique connue. Par exemple, les cellules BT474 sont positives pour les récepteurs d'œstrogènes (ER +), surexpriment le récepteur humain de croissance épidermique du facteur 2 (HER2 +) et des mutations dans le port TP53 (E285K) et PIK3CA (K111N) 13.
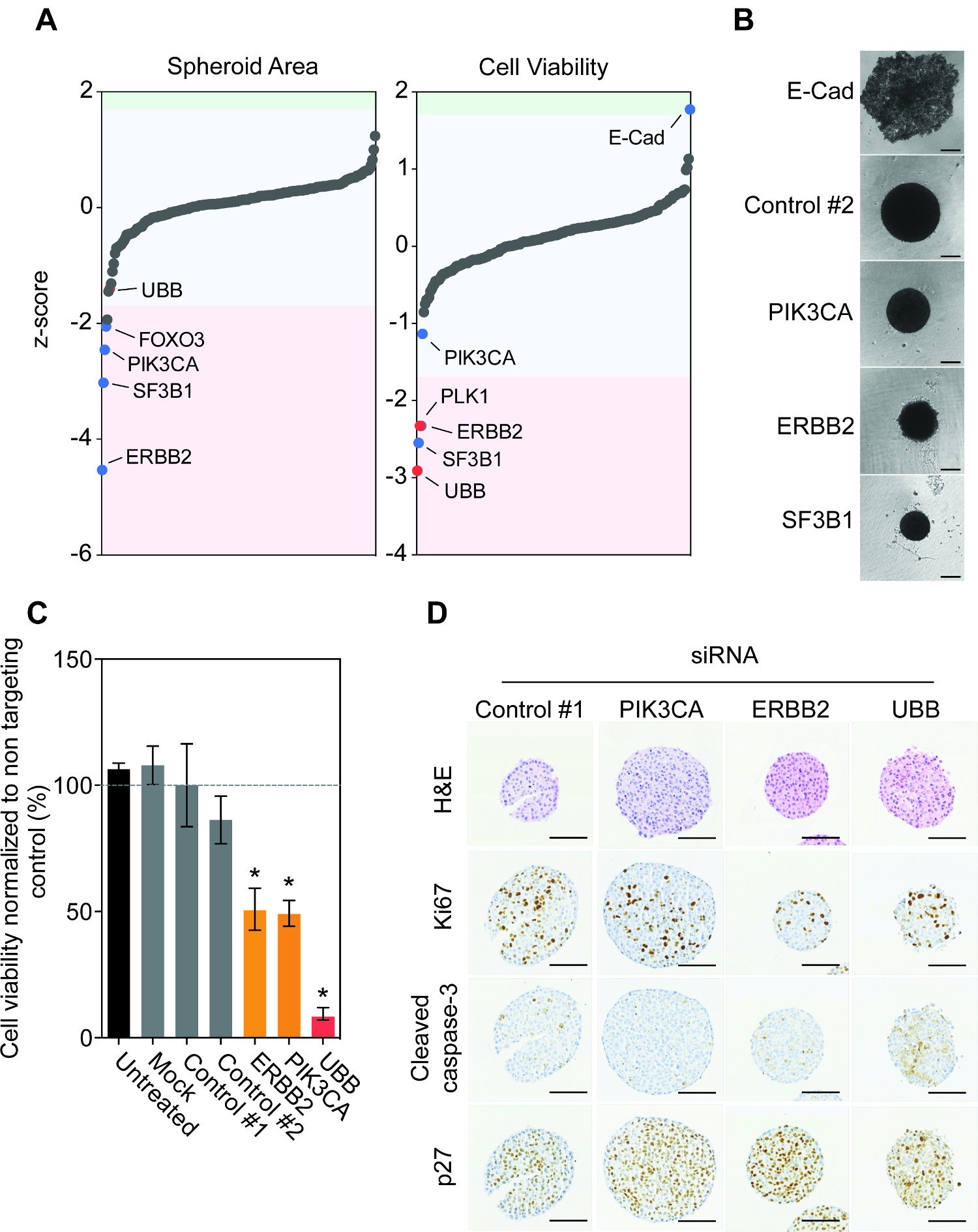
En utilisant le protocole décrit ci - dessus, nous avons surveillé la diminution du gène d'impact a eu sur la taille et la viabilité sphéroïde après sept jours de la transfection inverse de siRNA (figure 1E). Fait intéressant, la majorité des gènes n'a pas eu d'effet significatif sur la surface sphéroïdale ou la viabilité (figure 2A). Silencing de FOXO3, PIK3CA, ERBB2 et SF3B1 a abouti à la réduction reproductible la plus significative de la taille sphéroïde. Cette réduction a également été observée dans la viabilité sphéroïde après ERBB2 et SF3B1 silencieux. Encourageant, nous CONFirmé l'impact de PIK3CA, ERBB2 et SF3B1 taire sur la taille de sphéroïde en utilisant la microscopie à champ lumineux (figure 2B). Nous avons déjà identifié SF3B1 comme un gène essentiel dans de nombreux modèles de lignées cellulaires et donc siSF3B1 représente un bon contrôle de la mise à mort en plus de 14 UBB. Fait intéressant, les 200 gènes de seulement la réduction au silence de la E-cadhérine a entraîné une augmentation significative de la viabilité sphéroïde (figure 2A). Etude de la morphologie sphéroïde a montré que E-cadhérine silencing a entraîné un effondrement complet de l' architecture sphéroïde, avec des cellules viables reposant sur le fond de l'attache basse bien (figure 2B). Manuel de réexamen des données d'écran du volume sphéroïde a montré que cela avait également été observée, mais avait été éliminé de la zone de quantification en raison de l'objet étant au-dessus des restrictions jeu de taille. Comme souligné précédemment, les cellules BT474 surexpriment le récepteur tyrosine kinase HER2 et abritent une mutation oncogénique dans PIK3CA (K111N). Nous avons confirmé que le silençage de l' ERBB2 et PIK3CA conduit à une réduction sphéroïde viabilité, tandis que la transfection avec des témoins non-cible n'a eu aucun effet (figure 2C).
Suivant nous avons étudié l'impact de siRNA déplétion sur l'histologie sphéroïde. sphéroïdes BT474 ont été transfectées avec inverse non-ciblage siRNA contrôle et siRNA ciblant PIK3CA, ERBB2 et UBB. Silençage ERBB2 et UBB a entraîné une réduction du marqueur de prolifération Ki67 pro-comparé à siRNA contrôle (figure 2D). L'activation du marqueur pro-apoptotique caspase-3 a été observée après seulement UBB silençage, ce qui suggère que l'épuisement de HER2 et PIK3CA n'a pas donné lieu à une apoptose, mais étaient cytostatique plutôt que cytotoxique. En effet, le silençage de HER2 et PIK3CA a pour résultat une augmentation de l'expression de la protéine du cycle cellulaire, la protéine p27 d'arrêt par rapport au témoin sphéroïdes transfectées.
ent "fo: keep-together.within-page =" 1 "> Pris ensemble, ces résultats montrent que les cellules BT474 sont entraînées par oncogène HER2 et PIK3CA signalisation lorsqu'il est cultivé comme sphéroïdes 3D Plus important encore, ces résultats montrent qu'il est possible de. la conception et la mise en œuvre d'une bibliothèque sur mesure de dépistage de siRNA de centaines de gènes dans sphéroïdes de lignées cellulaires de cancer avec vigueur aux et reproductible.
Figure 1: Optimisation de la ligne cellulaire pour la croissance en 3 dimensions. A. Les lignées cellulaires de cancer du sein, et BT474 MCF10DCIS.com ont été cultivées dans des plaques de fixation basse pendant 7 jours. des images représentatives en fond clair ont été prises avec un microscope inversé. Les barres d'échelle = 100 um. B. MCF10DCIS.com et BT474 sphéroïdes ont été cultivées pendant 28 jours. 100 ul de milieu frais a été réapprovisionné tous les 3 - 4 jours. Spheroids ont été fixées dans 3,8% de formaldéhyde, embedded, sectionnées et colorées à l'hématoxyline et à l'éosine (H & E). images représentatifs sont présentés à un grossissement faible et élevé. Les barres d'échelle représentent 100 um et 33 um, respectivement. C. MDA-MB-231 sphéroïdes ont été cultivées pendant 21 jours. 100 ul de milieu frais a été réapprovisionné tous les 3 - 4 jours. Spheroids ont été fixées dans 3,8% de formaldéhyde, noyées, sectionnés et colorés avec Ki67 et clivés caspase-3. images représentatives sont indiquées. Les barres d'échelle = 100 um. D. BT474, MCF10DCIS.com, MDA-MB-231 et JIMT1 lignées cellulaires ont été transfectées avec inversion (réactif de transfection uniquement) simulée et les ARNsi de contrôle et de facturation à l' utilisation, les plaques de fixation ultra-basses ont ensuite été centrifugés pour former des sphéroïdes. Du milieu frais (100 pi) a été ajoutée aux jours 1 et 4. Au bout de 7 jours, la viabilité cellulaire a été quantifiée. Les données représentent la moyenne ± écart type de deux répliques biologiques indépendantes réalisées en triple exemplaire normalisée pour contrôler # 1. La signification statistique a été calculée à l'aide d'une APNUÉtudiants IRED test t (p <0,05). E. Un organigramme résumant le protocole de transfection inverse utilisé pour interroger la dépendance fonctionnelle du gène dans le cancer sphéroïdes de lignée cellulaire. S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
{kind=link}

Figure 2: Investigation génomique fonctionnelle des BT474 Spheroids Uncovers oncogènes dépendances. Cellules A. BT474 ont été inverse transfectées avec le gène de 200 siGENOME humaine bibliothèque de siRNA en triple. taille Spheroid et la viabilité a été observée. Les valeurs de données brutes étaient plaques médianes normalisées et z-scores ont été calculés pour identifier les valeurs aberrantes importantes supérieures à 1,7x l'écart type de la plaque médiane 15. Remarque gènes périphériques ERBB2, SF3B1, PLK1 et un UBBnd E-cad. ARNsi qui ont augmenté ou réduit de manière significative la zone sphéroïde et la viabilité sont ombrées en bleu et rouge, où le rouge représente les siARN de contrôle de. B. Les cellules ont été transfectées avec inverse non-ciblage de contrôle, E-cadhérine (E-Cad), PIK3CA, ERBB2 ou siRNA UBB puis centrifugés dans une plaque de fixation ultra-bas pour former des sphéroïdes. Après 7 jours, Brightfield images sphéroïdes représentatives ont été prises à l'aide d'un microscope inversé. Les barres d'échelle = 100 um. C. Les cellules ont été transfectées avec inverse non-ciblage de contrôle, PIK3CA, ERBB2 ou siRNA UBB puis centrifugés dans une plaque de fixation ultra-bas pour former des sphéroïdes. Au bout de 7 jours, la viabilité cellulaire a été quantifiée. Les données représentent la moyenne ± écart type de deux répliques biologiques indépendantes réalisées en triple exemplaire normalisée pour contrôler # 1. La signification statistique a été calculée à l' aide d' un des étudiants non appariés t-test (p <0,05). D. Au bout de 7 jours, les sphéroïdes ont été fixées, incorporées, sectionnés et colorésH & E, Ki67, caspase-3, et p27. images représentatives sont indiquées. Les barres d'échelle = 100 um. Notez que le H & E des sphéroïdes siControl apparaissent plus petits en raison d'un artefact de traitement et individuels intacts sphéroïdes choisis pour la coloration. Toutefois, cela ne porte pas atteinte à des changements observés avec les images numérisées et les résultats de la viabilité cellulaire. S'il vous plaît cliquer ici pour voir une version plus grande de cette figure.
{kind=link}
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | dix | 11 | 12 |
| La non-cible 1 | TP53 | DST | KMT2C | 1 non traitée | FCGBP | ARID1B | FBXM7 | TTC40 | Non ciblage 2 | ||
| GATA3 | TTN | MUC12 | MUC4 | AHNAK | HUWEI1 | DNAH11 | itpr2 | ABCA13 | CREBBP | ||
| MAP2K4 | PIK3CA | F5 | APOB | ANKRD30A | MUC17 | DNAH17 | LAMA2 | AS | CSMD2 | ||
| STARD9 | USH2A | FAT3 | LPR2 | CSMD3 | MYO18B | DNAH5 | MDN1 | ARHGAP5 | DNAH9 | ||
| CXCR3 | MUC16 | RB1 | PKHD1L1 | DNAH2 | SYNE2 | DYNC1H1 | PCLO | CACNA1B | ERBB2 | ||
| PLK1 | SYNE1 | LYST | PTEN | SPTA1 | 2 non traitée | VHL | RYR1 | COL6A3 | UBB | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | dix | 11 | 12 |
| La non-cible 1 | ENAM | RYR2 | BRCA2 | 1 non traitée | FMN2 | HECW1 | LAMB4 | SI | Non ciblage 2 | ||
| FHOD3 | macf1 | RYR3 | C2ORF16 | DMD | FRG1 | HERC2 | MYH11 | STAB1 | ZDBF2 | ||
| GOLGA6L2 | BEC | SMG1 | CACNA1E | DNA14 | GCC2 | HIVEP2 | NIPBL | TANC1 | ZNF536 | ||
| HMCN1 | NF1 | UBR5 | CACNA1F | DYNC2H1 | GON4L | HYDIN | PKD1L1 | TF | ANK3 | ||
| HRNR | OBSCN | USP34 | CYMA5 | FAM208B | GPR112 | ITSN2 | RNF213 | TPR | ASPM | ||
| PLK1 | PCDH15 | XIRP2 | COL7A1 | FLG2 | 2 non traitée | VHL | SAGE1 | UNC80 | UBB | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | dix | 11 | 12 |
| La non-cible 1 | DCHS2 | MUC5B | ZFHX4 | 1 non traitée | MYO9A | SPHKAP | CXORF22 | NCoR1 | VPS13D | ||
| ATR | DMXL2 | MXRA5 | ANK2 | KIAA1210 | PRUNE2 | TCHH | DANH6 | NOTCH2 | ANKRD12 | ||
| BIRC6 | DNAH10 | TENM1 | AU M | LRP1 | SCN10A | VPS13C | DNAH7 | SPEN | C5ORF42 | ||
| CDH1 | DNAH3 | PEG3 | DIDO1 | MAP1A | SCN2A | IPTR3 | ERBB3 | SRRM2 | CCDC88A | ||
| CUBN | NOCK11 | RELN | DNAH8 | MED12 | SHROOM2 | CEP350 | FAT4 | SZT2 | CHD4 | ||
| PLK1 | EYS | SACS | KIAA1109 | MED13 | 2 non traitée | VHL | KMT2A | VPS13A | UBB | ||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | dix | 11 | 12 |
| Non-ciblage | QSER1 | ARID1A | WDFY3 | 1 non traitée | sdk1 | TEX15 | LAMA1 | Non ciblage 2 | |||
| COL14A1 | SHROOM3 | ATRX | EFCAB5 | SF3B1 | CBFB | AHNAK2 | |||||
| CSMD1 | TBX3 | KIAA0947 | FOXA1 | ITPR1 | DDX3X | KIF4A | |||||
| MEFV | UBR4 | MYCBP2 | INPPL1 | FLG | HECTD4 | FAT2 | |||||
| MGAM | VCAN | NBEAL1 | MAP3K1 | AKAP9 | GPR98 | FOXO3 | |||||
| PLK1 | ZNF462 | SETX | NRP1 | HERC1 | 2 non traitée | VHL | UBB | ||||
Tableau 1: Disposition de plaque et siRNA Human Library De-convolution. Le tableau contient le contenu de chacune des piscines siARN et mise en page pour les plaques à faible fixation utilisés dans l'écran.
Discussion
des modèles tridimensionnels de cancer sont de plus en plus utilisées pour évaluer l'efficacité des composés connus et nouveaux qui ont été conçus pour tuer sélectivement des cellules cancéreuses. Sphéroïdes de cellules cancéreuses sont des structures qui présentent des conditions similaires à celles rencontrées dans les tumeurs in vivo, donc des composés ayant une efficacité accrue en 3D sont plus susceptibles d'avoir un effet in vivo. Cependant, ces modalités ne permettent pas l'identification de nouvelles cibles potentiellement qui n'a pas fait l'objet de la conception de médicaments qui pourraient avoir une efficacité considérable dans le traitement du cancer.
Nous avons développé un siRNA génomique fonctionnelle approche qui a permis l'inactivation de gène durable pour un maximum de sept jours dans le cancer sphéroïdes lignée cellulaire. Il y a plusieurs étapes critiques du protocole qui nécessitent une optimisation devant un écran de siRNA peut être effectuée. L'aptitude à former des sphéroïdes viables et reproductibles à grande échelle est essentielle. Par ailleurs, unconditions de transfection ppropriate doivent être rigoureusement optimisés. Nous suggérons trialing plusieurs réactifs de transfection différentes avec des non-ciblage approprié et des contrôles tuer avant d'essayer l'écran. Nous avons pu montrer que plusieurs lignées de cellules de cancer du sein couramment utilisés, à savoir BT474, MCF10DCIS.com, MDA-MB-231 et JIMT1 étaient prête à siRNA transfection. En outre, nous fournissons la preuve de principe des données de dépistage des 200 gènes les plus fréquemment mutés dans le cancer du sein chez les sphéroïdes BT474, ont confirmé leur dépendance sur l'amplification de HER2 et la mutation oncogénique de PIK3CA. Fait intéressant, le silençage du FOXO3 facteur de transcription a entraîné une réduction de la taille sphéroïde, mais aucun effet significatif sur la viabilité. FOXO3 est connu pour réguler la réponse à l' hypoxie, ce qui modifie la capacité métabolique des cellules cancéreuses qui leur permet d'adapter plus facilement à leur environnement 16. Ce rôle pourrait interférer avec la lecture de la viabilité des cellules qu'il détecte ATP abondance, L'un des principaux produits du métabolisme cellulaire.
À l' appui de l'observation d'une réduction de la taille sphéroïde, il a été démontré que le silençage de FOXO3 HeLa dans des xénogreffes de tumeur altération de la croissance et de l' apoptose 17 induite. Il est important de noter que certains gènes peuvent influer sur la capacité des cellules cancéreuses à conserver leur architecture 3D, ce qui pourrait entraîner des résultats faussement positifs. Par exemple, knockdown de E-cadhérine a abouti à la dissolution de la structure sphéroïde BT474. Cela avait déjà été signalé en utilisant E-cadhérine anticorps 18 ciblé. Comme avec toute plateforme de criblage, des cibles potentielles devraient être de nouveau criblées pour évaluer la reproductibilité de l'effet observé. Il y a des limites à la technique, à savoir la nature transitoire du gène knockdown de siRNA à médiation. Soutenus au silence plus de sept jours n'a pas été possible avec le siRNA.
L'avantage de cette approche est qu'elle peut être couplée avec divers autres biomecolorants triques non pas seulement ceux qui évaluent la viabilité sphéroïde, par exemple, en donnant l'information spatiale de l'hypoxie sphéroïde ou des cellules de surveillance subissant une apoptose. En outre, parce que les scans de lecteur de plaque sont relativement rapide et non invasive, l'impact de siRNA sur la taille sphéroïde peut être évaluée au fil du temps plutôt que juste au point final expérimental. En effet, nous étudions actuellement plusieurs de ces avenues au sein de notre pipeline de dépistage. Une approche alternative qui utilise des cultures 3D pour identifier de nouvelles dépendances est l'utilisation des bibliothèques chimiques qui inhibent large, soit une gamme de cibles ou de familles particulières de protéines. En effet, Bitler et al. utilisé cette approche ciblée pour identifier l'interaction de létalité synthétique entre le statut de ARID1A et les inhibiteurs EZH2 dans l' ovaire carcinomes à cellules claires 19. La découverte de CRISPR-cas9 technologie d'édition de gènes a également permis le développement d'écrans génétiques dans les cultures organoïdes et in vivo. Cependant, le tson approche dépend de disposer d' installations appropriées sur l'animal et peut être un coût prohibitif 20.
En conclusion, nous croyons que nous avons décrit un protocole que les modèles plus précisément les oxygène et de nutriments dégradés, qui sont caractéristiques du microenvironnement de la tumeur in vivo, permettant l'identification des cibles de cancer nouvelles ou validation robuste des objectifs établis. De plus, notre protocole peut être appliqué à tout type de lignée cellulaire qui forme sphéroïdes et peut donc être utilisée en routine dans le milieu de la recherche sur le cancer pour les écrans de siRNA à haut débit.
Déclarations de divulgation
Open Access fees were supported by Nexcelom Bioscience, LLC.
Remerciements
The authors would like to thank Fredrik Walberg (FACS and imaging facility, ICR) for assistance with microscopy. This work was funded by Breast Cancer Now. RN is the recipient of a Breast Cancer Now Career Development Fellowship (2011MaySF01).
matériels
| Name | Company | Catalog Number | Comments |
| Lullaby | Oz Biosciences | LL70500 | lipid-based transfection reagent |
| Viromer | Lipocalyx | VB-01LB-01 | virus-like polymer transfection reagent |
| Ultra-low attachment plate | Corning | CLS7007 | 96 well plate |
| Foil plate seals | ThermoFisher | AB-0626 | |
| Luminescent cell viability dye | Promega | G7570 | CellTitre-Glo |
| Pipette tips (200 μL) | Starlab | S1111-0806 | |
| Pipette tips (10 μL) | Starlab | S1111-3800 | |
| Pipette tips (1, 000 μL) | Starlab | S1122-1830 | |
| Serological pipettes (5 mL) | Sarstedt | 86.1253.025 | |
| Serological pipettes (10 mL) | Sarstedt | 86.1254.025 | |
| Serological pipettes (25 mL) | Sarstedt | 86.1685.020 | |
| RPMI Media | GIBCO | 11875-093 | |
| DMEM Media | GIBCO | 11965-084 | |
| Opti-MEM | GIBCO | 31985070 | |
| Feta bovine serum | GIBCO | 16140063 | |
| siRNA | Dharmacon | Cherry picked library | |
| Countess Cell Counter | ThermoFisher Scientific | AMQAX1000 | |
| Cell counting chamber slides | ThermoFisher Scientific | C10312 | |
| Celigo S | Nexcelom | contact company | |
| Victor X5 | Perkin Elmer | contact company | |
| Benchtop centrifuge | Various | ||
| Axiovert Inverted brightfield microscope | Zeiss | contact company | |
| Tissue culture CO2; Incubator | Various | ||
| Mulitichannel pipette | Various |
Références
- Seton-Rogers, S. E., et al. Cooperation of the ErbB2 receptor and transforming growth factor beta in induction of migration and invasion in mammary epithelial cells. Proc Natl Acad Sci U S A. 101 (5), 1257-1262 (2004).
- Debnath, J., Muthuswamy, S. K., Brugge, J. S. Morphogenesis and oncogenesis of MCF-10A mammary epithelial acini grown in three-dimensional basement membrane cultures. Methods. 30 (3), 256-268 (2003).
- Chia, S. K., et al. Prognostic significance of a novel hypoxia-regulated marker, carbonic anhydrase IX, in invasive breast carcinoma. J Clin Oncol. 19 (16), 3660-3668 (2001).
- Trastour, C., et al. HIF-1alpha and CA IX staining in invasive breast carcinomas: prognosis and treatment outcome. Int J Cancer. 120 (7), 1451-1458 (2007).
- Wilson, W. R., Hay, M. P. Targeting hypoxia in cancer therapy. Nat Rev Cancer. 11 (6), 393-410 (2011).
- Ros, S., et al. Functional metabolic screen identifies 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 4 as an important regulator of prostate cancer cell survival. Cancer Discov. 2 (4), 328-343 (2012).
- Baenke, F., et al. Functional screening identifies MCT4 as a key regulator of breast cancer cell metabolism and survival. J Pathol. 237 (2), 152-165 (2015).
- Schug, Z. T., et al. Acetyl-CoA synthetase 2 promotes acetate utilization and maintains cancer cell growth under metabolic stress. Cancer Cell. 27 (1), 57-71 (2015).
- Mashimo, T., et al. Acetate is a bioenergetic substrate for human glioblastoma and brain metastases. Cell. 159 (7), 1603-1614 (2014).
- Comerford, S. A., et al. Acetate dependence of tumors. Cell. 159 (7), 1591-1602 (2014).
- Vinci, M., Box, C., Eccles, S. A. Three-dimensional (3D) tumor spheroid invasion assay. J Vis Exp. (99), e52686 (2015).
- Vinci, M., Box, C., Zimmermann, M., Eccles, S. A. Tumor spheroid-based migration assays for evaluation of therapeutic agents. Methods Mol Biol. 986, 253-266 (2013).
- Barretina, J., et al. The Cancer Cell Line Encyclopedia enables predictive modelling of anticancer drug sensitivity. Nature. 483 (7391), 603-607 (2012).
- Maguire, S. L., et al. SF3B1 mutations constitute a novel therapeutic target in breast cancer. J Pathol. 235 (4), 571-580 (2015).
- Brough, R., et al. Functional viability profiles of breast cancer. Cancer Discov. 1 (3), 260-273 (2011).
- Ferber, E. C., et al. FOXO3a regulates reactive oxygen metabolism by inhibiting mitochondrial gene expression. Cell Death Differ. 19 (6), 968-979 (2012).
- Jensen, K. S., et al. FoxO3A promotes metabolic adaptation to hypoxia by antagonizing Myc function. EMBO J. 30 (22), 4554-4570 (2011).
- Ivascu, A., Kubbies, M. Diversity of cell-mediated adhesions in breast cancer spheroids. Int J Oncol. 31 (6), 1403-1413 (2007).
- Bitler, B. G., et al. Synthetic lethality by targeting EZH2 methyltransferase activity in ARID1A-mutated cancers. Nat Med. 21 (3), 231-238 (2015).
- Dow, L. E., et al. Inducible in vivo genome editing with CRISPR-Cas9. Nat Biotechnol. 33 (4), 390-394 (2015).
Réimpressions et Autorisations
Demande d’autorisation pour utiliser le texte ou les figures de cet article JoVE
Demande d’autorisationExplorer plus d’articles
This article has been published
Video Coming Soon
À PROPOS DE JoVE
Copyright © 2025 MyJoVE Corporation. Tous droits réservés.