
Method Article
糸球体疾患のマウス ・ モデルにおける腎機能の評価
要約
このプロトコルでは、糸球体疾患のマウス ・ モデルで実施する必要があります完全腎臓の仕事-アップについて説明します。糸球体疾患のすべてのマウスのモデルに適用することができます糸球体機能の詳細な機能・構造・機構解析を可能にするメソッド。
要約
ひと腎臓疾患を模倣するマウスモデルの使用はますます一般的になっています。我々 の研究は糖尿病性腎症とポドサイト固有 VEGF A ノックアウト マウスを用いて糸球体機能の評価に焦点を当ててください。したがって、このプロトコルは、糸球体疾患、腎臓および単一のマウスから得られる糸球体の機能に関する情報の膨大な量を有効にすることのこれらのマウスのモデルを評価するために我々 の研究室で使用される完全腎臓仕事アップについて説明します。糸球体の機能を評価するために文献の提示の方法と比較して本稿で説明したメソッドを使用して有効複数の側面から評価される完全に糸球体の表現型にします。このメソッドを使用して、研究者はモデルの腎臓の表現型を決定し、表現型を開発する理由とメカニズムを評価できます。病気のメカニズムに重要な情報は、これらのモデルの潜在的な治療手段を調べる場合に必要です。構造・超構造検討と同様、尿中アルブミン ・ クレアチニン比および個々 の糸球体水透磁率の測定による糸球体濾過障壁の詳細な機能評価を可能にする方法過ヨウ素酸シッフ染色、電子顕微鏡を使用しています。さらに、mRNA やタンパク質のレベルで遺伝子の調節不全の分析には、糸球体の機能の解明ができます。このプロトコルは、糸球体疾患のすべてのマウスのモデルに適用できる汎用が適応できる方法を紹介します。
概要
ひと腎臓疾患を模倣するマウスモデルの使用はますます一般的になっています。マウスのようなモデルは、自然発症高血圧ラット (SHR)1, ストレプトゾトシン (STZ) など自発的なモデル-誘発糖尿病ラットとマウス2と db/db タイプ II 糖尿病マウス3、遺伝子組み換えモデルプライマリ ポドサイト固有焦点区域糸球体硬化症 (FSGS) モデル4、ポドサイト固有血管内皮増殖因子 (VEGF A) ノックアウト (KO VEGF-A) モデル5とアルポート症候群モデルの6、および取得5/6 腎7など片側尿管閉塞 (UUO) モデル8モデル。これらのモデルにおける腎糸球体機能のさまざまな側面を評価するためにいくつかのテクニックがあります。本方法の目的は、包括的な作業を完全に糸球体の機能を評価するために腎臓病のマウス ・ モデルで実行する必要がありますを示すことです。
このメソッドを使用しての背後にある理論的根拠は、複数の側面から評価される完全に糸球体の表現が可能です。これは糸球体の透過性、タンパク質と水と糸球体の構造異常と Mrna やタンパク質の正常な糸球体機能のために不可欠の式/スプライシングの変化の両方評価含まれています。このメソッドを使用して、研究者はモデルの腎臓の表現型を決定し、表現型を開発する理由とメカニズムを評価することができます。これは、これらのモデルの潜在的な治療上の道を調べるときに必要なある病気のメカニズムに重要な情報です。
文学、尿中アルブミンの増加レベルで決まる糸球体疾患のマウスモデルで提示される一般的な現象です。しかし、糸球体の機能を決定する単一のメソッドが常に効果的なないことを示唆する証拠があります。尿中アルブミン排泄率または尿中アルブミン クレアチニン比 (uACR) 測定は、総腎機能および個々 の糸球体にだけ情報を提供します。以前の研究では同じ腎臓5,9,10から異なる糸球体透過性が変化する場合を示しています。さらに、個々 の糸球体の透過性の評価は、糸球体機能を評価するためのより敏感な方法個々 の糸球体の水の透過性を測る手法 (LpA/V は私) uACR9の測定よりも糸球体機能の変化により敏感になることを示しています。この試金は、蛋白尿、c57BL/6 背景11などに耐性のあるマウス モデルで有益です。メソッド本稿の利点は、それが水に個々 の糸球体透過性と同様、両方のアルブミン総腎透磁率を調べることです。
糸球体の構造異常の検討はしばしば電池など過ヨウ素酸シッフ (PAS)、ヒアリン、汚れや銀汚れによって評価されます。これらは、スコアリング法による腎疾患のレベルを評価する訓練された腎病理医を有効にします。すべての良い方法の糸球体のマクロ構造の変更がで常に観察されていませんが、急性腎障害は12をモデル化します。このメソッドは、上記の腎組織学手法を実施に加えで、糸球体の超構造する必要があります電子顕微鏡 (EM) で評価されるも提案します。染色糸球体は通常光学顕微鏡の下で比較的普通に見ることができます。ただし、EM、糸球体基底膜 (GBM) 幅の小さな変化で評価にポドサイト足プロセス卑下、内皮のピンホールとサブ ポドサイト領域カバレッジに分析されます。したがって、糸球体超構造と微細構造の両方が糸球体障害のメカニズムを決定する評価が不可欠です。
糸球体の構造異常を評価するとともに、さらに糸球体疾患のメカニズムを解明する mRNA とタンパク質の発現とスプライシングと同様にタンパク質の活性化 (例えば、リン酸化)、変更を検討すべき。糸球体疾患を見るときとか、例えば、KO/オーバー-expressing ポドサイト固有 VEGF-A KO マウス5のような糸球体の細胞で遺伝子はタンパクと mRNA の変化が内にのみ検討が重要なとき、糸球体細胞、腎全体ではないです。このプロトコルでは、糸球体マウス腎皮質から分離されたタンパク質・ RNA、分離法について説明します。これは疾患モデルの糸球体における蛋白質/mRNA 不全の特定の分析をことができます。
このプロトコルでは、糸球体疾患、腎臓および単一のマウスから得られる糸球体の機能に関する情報の膨大な量の有効化のマウス ・ モデルで実施する必要があります完全腎臓の仕事-アップについて説明します。糸球体疾患のすべてのマウスのモデルに適用することができます糸球体機能の詳細な機能・構造・機構解析を可能にするメソッド。
プロトコル
英国の法律およびローカル倫理委員会の承認に基づきすべての実験を行った。動物実験は、ブリストル研究倫理委員会の大学によって承認されました。
1 尿中アルブミン クレアチニン比 (uACR)
メモ: uACR はアルブミン GFB の透水性を評価するために使用されます。尿中アルブミンの存在は、クレアチニン尿流量の変動を制御するために正規化された GFB 全体にわたる透過性亢進を示します。蛋白尿、慢性腎臓病の一般的なマーカーです。
- 実験的エンドポイントまで尿実験のベースライン、定期的 (週 1 回、月 1 回) を収集します。
- 水と濃縮ダイエット マウス代謝ケージを設定します。静かな部屋で 6 h の個々 のケージにマウス (男性、高齢者 6-8 週間) を配置します。
- 空のケージからレギュラー住宅と収集の尿に戻りマウス。50 μ L の最小値が必要です。
注: マウスで与えられた時間で尿が生成されない場合は、別の日に暖かい部屋で繰り返します。 - 500 x g で 10 分間収集で尿に尿を遠心し、ポドサイト損失を評価するために土砂を保持します。この時点で、短期的には-20 ° C で尿を格納します。
- 生理食塩水 x Tris バッファー 1 (TBS pH 7.5) 1:10000 希薄、蛋白尿の程度によって 1: 500 で 1% ウシ血清アルブミン (BSA) に尿を希釈します。
注: 最後のボリュームにする必要があります > 400 μ L。 各時右の希釈を決定する最適化のポイント。 - マウスのアルブミンの酵素結合抗体法 (ELISA) が製造元の指示に従ってを用いた尿中アルブミン濃度を定量化します。
- 簡単に言えば、室温で 1 時間抗マウス アルブミン一次抗体 (0.5 M 重炭酸塩-炭酸塩 ph 9.6 10 μ g/mL) の 100 μ L の蛋白結合のために最適化されている elisa 用プレートをコートします。
- 5 回で 1 x TBS プラス 0.05% 井戸から洗って余分な抗体トゥイーン (pH 8.0)、室温で一晩ソリューション (1% BSA pH 8.0 で 1 x TBS) ブロックを 200 μ l 添加し、。
- 5 倍のプレートを洗って、基準の 100 μ L を追加 (シリアル希釈: 15.63 μ g/ml 1 %bsa、pH 8.0 で 1 x TBS に 2000年 μ G/ml)、空白、3 通の希釈サンプル。室温で 1 時間のままにして、5 回プレートを洗ってください。
- 各ウェル (1 %bsa、pH 8.0 で 1xTBS で 10 ng/mL) に HRP 検出抗体の 100 μ L を追加し、1 時間室温で孵化させなさい。
- 5 回プレートを洗浄し、各ウェルに酵素基質液を 100 μ l 添加を追加します。15 分のための開発、0.18 M 硫酸の 100 μ L の追加によって反作用を停止し暗闇の中でプレートを残して (H2SO4)。
注意: H24は腐食性です。
- 450 の吸光度でプレートを読み取ることによって各サンプルのアルブミン濃度を決定する nm。標準曲線を使用して、各サンプルで現在のアルブミンを定量化します。技術的な繰り返しの CV 値の 5% より大きい場合、それらのサンプルのアッセイを繰り返します。
- また、電気泳動法を用いた尿中アルブミン濃度を評価します。15 μ L の尿に蛋白質のサンプル読み込みバッファー x 4 の 5 μ L を追加します。95 ° C 10 分のサンプルを加熱、4-12% プレキャスト トリス ゲルにロードします。
- 100 V と製造元のプロトコルごとブルーで一晩染色ゲルを実行します。
- ゲルをイメージングした後デンシトメトリーを使用する相対倍変更アルブミン濃度を評価します。
注: 場合はアルブミンが予想される場合は、バンドは認められなかった、尿の蛋白質濃度を評価し、尿ゲルに追加のボリュームを調整します。
- DH2O 1:1、1:5、1:10 で生の尿サンプルを希釈します。
注: 最後のボリュームにする必要があります > 70 μ。 各サンプルの右の希釈を決定する最適化。 - キットの指示に従って化学物質分析法を用いた尿中クレアチニン濃度を定量化します。簡単に言えば、3 通 96 well プレートにクレアチニン基準、空白、または希釈尿の 20 μ L をロードします。
- 490 の吸光度でプレートを読み取ることによって各サンプルのクレアチニン濃度を決定する前に、と酸性溶液添加後の nm (注意、腐食性の皮膚との接触を避けるため)。
注: 吸光度値の差、各サンプルのクレアチニン濃度に正比例します。標準的な曲線は、基準からの世代です。技術的な繰り返しの CV 値の 5% より大きい場合、それらのサンプルのアッセイを繰り返します。 - UACR (μ g/mg) を生成します。グラフィカルな表現の各マウスのベースライン値データを正規化します。
2. 組織および血のコレクション
注: 腎臓と糸球体の組織は、腎疾患の構造、蛋白質、および mRNA の表現マーカーを評価するために使用できます。血液は、調整される腎疾患、糸球体のろ過能力の低下を示すことができるクレアチニンなどの腎機能のマーカーを評価するために使用できます。
- 次のソリューションの準備: 0.1 M ナトリウム cacodylate (pH 7.3)、リン酸緩衝生理食塩水 (PBS)、哺乳類リンゲル液 (115 mM 塩化ナトリウム (NaCl) 10 mM 酢酸ナトリウム (CH3COONa)、1.2 × 1 で 4% パラホルムアルデヒドで新鮮な 2.5% グルタルアルデヒドmM リン酸ナトリウム (Na2HPO4)、25 MM 重炭酸ナトリウム (NaHCO3)、(MgSO4) 硫酸マグネシウム 1.2 mM、1 mM 塩化カルシウム (CaCl2)、5.5 mM D (+) グルコース、pH 7.4) 1 %bsa と 1x PBS。
注意: 2.5% グルタルアルデヒド: 毒性、感作性、刺激;ヒューム キャビネットで使用します。0.1 M ナトリウム cacodylate: 有毒なヒューム キャビネットで使用。4 %pfa: 定着剤、ヒューム キャビネットで使用 - 以下の資料を準備: イソフルラン、小さなエチレンジアミン四酢酸 (EDTA)-血液チューブ、23-25 G 針、5 mL コーティング EDTA 注射器、10 mL ガラスバイアル、10 mL プラスチック容器、0.5 mL プラスチック チューブ、使い捨てティッシュ金型、ドライアイス、液体 Nをコーティング2、マウス外科的ツール、および最適な切削の媒体 (10 月)。
- イソフルランの商工会議所またはターミナルの麻酔注射の麻酔剤 (ペントバルビ タール、50 mg/kg などの同等のルートを使用して、足のパッドに針を刺すに対応マウスを行い深い麻酔下マウスを置きます腹腔内 [IP];タヴェルタン、240 mg/kg IP)、または二酸化炭素 (CO2) 露出 (75% CO225% O2)。
- 左心室に心臓穿刺を介してマウスを処分し、できるだけ多くの血液を収集します。4 h までの血の EDTA 被覆管に転送します。優先、マウスはない頚静脈の破裂に注意して頚部転位でカリングできます。
- 腹部と氷冷 1x PBS で洗浄から腎臓を解剖します。
- 皮質の糸球体を調べる腎皮質中の 1 つのポールを取り外して 1 mm3粉々 にカットします。深い傍髄質糸球体を調べると、髄質からの組織と同じテクニックを繰り返します。EM バイアルで 2.5% グルタルアルデヒド溶液 5 mL に配置します。4 ° C でのストア
注意: 2.5% グルタルアルデヒド溶液: 毒性、感作性、刺激;ヒューム キャビネットで使用します。
注: 最良の結果のための 1 ヶ月内のプロセス。 - 組織削除腎臓皮質と傍髄質糸球体が存在するように、修正の上から 3 番目 24 時間 70 %5 mL に転送 4 ° C で 4% パラホルムアルデヒドの 5 mL の EtOH のパラフィン包埋前に、の 24 時間。
注意: 4 %pfa: 定着剤、ヒューム キャビネットで使用 - 蛍光抗体法により, 皮質・髄質糸球体が存在する組織型と 10 月のコートになるように、腎臓の 3 分の 1 の場所を凍結し、-80 ° C で保存にドライアイスの場所
- タンパク質と RNA は、0.5 mL プラスチック チューブとスナップに腎皮質の場所 3 × 2 mm3個は液 N2でフリーズします。-80 ° C でストアRNA の長期の組織保存、RNA 安定化ソリューションの 5 巻で組織を置くし、-80 ° C で保存
- 糸球体の分離、残りの腎臓組織をスライスし、5 mL リンゲル液の哺乳類 1 %bsa を氷の上に配置します。すぐに糸球体をふるいを準備します。
3. 血漿クレアチニン
注: 血漿クレアチニン調整される腎疾患、糸球体のろ過能力の低下を示すことができます。プロトコルがここで説明はしませんが、血中尿素窒素 (BUN) も評価することができます。
- 4 ° C で 15 分間 500 x g で血液サンプルを遠心分離します。
- 格納できる-20 ° C で短期的にはこの時点でプラズマを収集します。
- 上記の指示に従って化学クレアチニンを用いたプロトコル 1.11 では尿中のクレアチニン血漿クレアチニン濃度を定量化します。
- 490 の吸光度でプレートを読み取ることによって各サンプルのクレアチニン濃度を決定する前に、と酸性溶液添加後の nm。
注: これらの吸光度の値の違いは、各サンプルのクレアチニン濃度に比例です。標準的な曲線は、基準からの世代です。技術的な繰り返しの CV 値の 5% より大きい場合、それらのサンプルのアッセイを繰り返します。
4. 糸球体の分離
注: 個々 の糸球体前のヴィヴォの透磁率と同様、特定の蛋白質の発現と mRNA 糸球体疾患マーカーを評価するために糸球体を分離できます。
- リンゲル液の哺乳類 1 %bsa は、腎臓の組織を取るそしてふるい分け法13標準を使用して糸球体を解剖します。簡単に言えば、70 の (下) μ m、100 μ m、125 μ m、175 μ m、250 μ m とガラス ビーカーに 425 μ m (最上位) ふるいをスタックします。
- 腎臓をマッシュ アップ、ふるい、氷冷哺乳類のリンゲル液を用いた 1 %bsa を押し通す 425 μ m にシリンジのプランジャーを使用します。腎臓のビット プッシュされるとトップのふるいを削除し、次に同じように進みます。100 だけまで繰り返し μ m、70 μ m ふるいのままです。
- 100 μ m、70 μ m によって保持される糸球体の収穫を転送 1 %bsa、氷の上で新鮮な哺乳類リンゲル液溶液 10 mL をふるい。
注: リンゲル液 mL あたり糸球体の数が少数の場合は、リンゲル液を使用して最後の 2 つのふるいから糸球体を収集の量を減らします。 - 2 つの個別のチューブ (2.5 mL) に糸球体と 1,000 x g で 4 ° C で 10 分間遠心を含む溶液 5 mL を削除します。上澄みを除去し、タンパク質と RNA の抽出、後日-80 ° C で保存する前に凍結する液体 N2の糸球体。
- 糸球体の LpA の測定のための 37 ° c の水浴の糸球体を含む残りの溶液を配置/V は私前のヴィヴォ。腎臓を削除する 3 h 以内に完了します。
5. 糸球体の水透過性 (LpA/V は私)
注: 糸球体 LpA/V私アッセイにより個々 の糸球体の透過性の前のヴィヴォ測定再現性のある方法で。糸球体の LpA の増加/V は私は、GFB の中断に腎疾患の暗示的であることを示します。
- 糸球体の LpA を設定/V は私リグ サーモンら10で説明したよう。セットアップの詳細な図は、図 1を参照してください。
- 次のソリューションの準備: リンゲル液の哺乳類 1 %bsa (pH 7.4) とリンゲル液の哺乳類 8 %bsa (pH 7.4)。37 ° c. に両方を暖める
- ガラス毛細管からマイクロ ピペットを引く (光学密度: 1.2 mm)。顕微鏡下でマイクロ ピペットを切断することによって 5-8 μ m 開口部先端を生成します。
- 使用する糸球体の LpA/V私はボーマン嚢や吸引を使用してマイクロ ピペットに管状の断片がないそのままの個々 の糸球体をキャッチするリグします。Oncometric 試金の詳細な要約は、サーモンら10で発見されます。簡単に言えば、糸球体がキャッチし、吸引ピペットに固定されて、顕微鏡下で糸球体のビデオ録画を開始します。
- まず、30 の 1 %bsa リンゲル液ソリューションにおける糸球体を平衡濃度 8 %bsa リンガー向け 10 に、perifusate を切り替える前に s s。Perifusate を 1 %bsa リンゲル液に切り替える、録音を停止します。
- 糸球体を洗い流すし、マウスあたり 10-15 糸球体のプロセスを繰り返します。Perifusate 流量が同一と高速にないことを確認 (10 mL/分) 糸球体構造を歪めるように。
- 球体の容積 (V私) に正規化された糸球体の水透過性 (LpA) を計算する糸球体の収縮の初期速度を測定します。サーモンら10の分析に関する詳細な情報を見つけることが。
6. 過ヨウ素酸シッフ (PAS) 染色
注: PAS 染色は、糸球体毛細血管ループの基底膜および尿細管上皮にハイライト表示されます。それは糸球体細胞、メサンギウム マトリックスと潜在的な拡張、および糸球体基底膜 (すなわち肥厚、凹凸) の潜在的な変更の詳細な可視化を実現。 にします。
- ポリ-L-リジン コーティング スライドに 5 μ m の厚みでミクロトームを用いたパラフィン包埋、PFA 固定腎皮質をセクションします。1 h. 確認の 37 ° C で乾燥セクションは任意のひだや、顕微鏡で形態を歪めることができる穴を含まない。
- キシレンの 2 回の孵化によってスライドを deparaffinize (注意、刺激; ヒューム キャビネットで使用) 3 分、100% EtOH 3 分、95%、70%、50% で、一度で二度 3 分ごとに、すべての部屋の温度でのエタノール。DH2o. でスライドを再水和物します。
- 周期酸溶液でスライドを孵化させなさい (注意、刺激; ヒューム キャビネットで使用) (1 g/dL) 5 分と、すすぎの dH2o. のいくつかの変更でスライドは 100 mL dH2O 常温コンテナーを使用します。
注意: 周期的な酸溶液: 刺激;ヒューム キャビネットで使用します。 - シフの試薬のスライドを孵化させなさい (Parasoaniline 塩酸 6 g/L とナトリウム亜硫酸 HCl 0.25 mol/L で 4%) 室温で 15 分間。5 分間流水でスライドを洗います。
- 3 のヘマトキシリンで対比染色 15 分間水道水を実行しているスライドを徹底的に洗浄する前に s。
注: いくつかの最適化は、ヘマトキシリン染色に最適な時間を決定する必要があります。 - 手順 6.2 deparaffinization プロトコルのリバースを使用してスライドを脱水します。キシレンと終了します。
- 空気乾燥スライドとキシレン ベース マウント メディアをマウントします。
- 糸球体構造を評価するために 400 倍の倍率での顕微鏡のイメージ。以下を評価: 肥厚し、糸球体基底膜の不規則性毛細血管ループ、線維組織、多発性硬化症、細胞増殖の崩壊 (内皮、ポドサイトとメサンギウム、または房を浸潤した炎症細胞)。
注: 糸球体病態生理学の包括的な評価、他腎臓の病変必要があります細管などに評価されました。
7. 透過型電子顕微鏡 (TEM)
注: TEM は、腎臓は、糸球体基底膜、ポドサイト足プロセスは光学顕微鏡で見えない血管内皮のピンホールなどの超構造異常の検討を使用できます。これは (すなわち、ない蛋白尿および主要な構造異常) 腎障害はそうは発音しないモデルで重要です。
- 2.5 %gluteraldehdye - 固定さいの目に切られた腎臓を取る、後 1% オスミウム四酸化 1 h. 洗浄 50 mL の 0.1 M cacodylate バッファー (pH 7.3) し、dH2O の 50 mL 用 (3 x 15 分変更) で修正します。
注意: 2.5% グルタルアルデヒド: 毒性、感作性、刺激;ヒューム キャビネットで使用します。0.1 M ナトリウム cacodylate: 有毒なヒューム キャビネットで使用 - エタノールの脱水してアラルダイト樹脂に埋め込みます。
- 50-100 nm の厚さの部分をカットし、3% 酢酸 (水様) ウランとレイノルズの鉛の混合水溶液で染色します。
- デジタル顕微鏡の 940 X で糸球体のいくつかの領域を引き継ぐ、1250 X と 6200 X 必ず足、GEnCs、糸球体基底膜、およびメサンギウム識別できます。
- ImageJ を使って盲目の糸球体を分析できます。各 6200 X 顕微鏡写真の尺度を設定するには、定規などの既知の距離の 2 つの点の間に線を描画します。分析に移動し、縮尺を設定、線の長さをピクセル単位で表示されます。既知の距離と測定単位を入力します。各パラメーターの測定には以下のプロトコルを使用します。
注: この分析には、マウスの 1 日あたり約が必要です。十分な検定力、マウス 3 から 3 腎糸球体から平均測定値を使用します。- 糸球体基底膜、6200 X 顕微鏡像上固定デジタル グリッド (10 x 10) を挿入し、グリッド線が、糸球体基底膜を越える時点で糸球体基底膜の厚さを測定します。直線ツールを使用して血管内皮細胞膜に垂直正接で基底ポドサイト足プロセス細胞膜に基底の内皮細胞膜から測定します。10 の個々 の測定値から各糸球体の平均の測定を決定します。
- 内皮開窓数 6200 X 顕微鏡写真で現在の GBM の長さを測定し、GBM の単位長さ当たりの内皮のピンホールの数をカウントします。糸球体あたり少なくとも 4 顕微鏡写真から平均を取る。
- ポドサイトのプロセスの幅を足、6200 X 顕微鏡像上固定デジタル グリッド (10 x 10) を挿入します。グリッド線を越えたポドサイト足プロセスの幅を測定します。糸球体基底膜; を満たす足プロセスの最も広い部分の幅を計測します。ポドサイトの基底膜の接線に垂直な線を確認します。10 の個々 の測定値から各糸球体の平均の測定を決定します。
- スリット幅ポドサイト, 6200 X 顕微鏡像上固定デジタル グリッド (10 x 10) を挿入します。グリッド線を越えるポドサイト スリット板の幅を測定します。これ足プロセスはポドサイト膜に膜から足の各プロセスの最も広い部分で、一緒に最も近いポイントです。ポドサイトの基底膜の接線に垂直な測定を確認します。10 の個々 の測定値から各糸球体の平均の測定を決定します。
- ポドサイトの数のプロセスを足、GBM 6200 X 顕微鏡写真の現在の長さを測定、糸球体基底膜の単位長さあたりのポドサイト足プロセスの数をカウントします。糸球体あたり少なくとも 4 顕微鏡写真から平均を取る。
- サブ ポドサイト空間範囲で、ニールらの詳細な方法を参照してください。14。
- 940 X 顕微鏡を使用して、目で異常な構造、預金、および浸潤の有無を糸球体を調べる。
8. ポドサイトと内皮マーカーの蛍光
注: 免疫染色は、糸球体疾患で折りたたむことができます内皮の毛管ループなどの蛋白質の表現パターンの可視化を使用できます。
- 切断する前に 2 時間-20 ° C で凍結腎臓を含む 10 月金型を配置します。腎臓の切断面は慎重によく指向ティッシュ セクションを有効にする 10 月金型の底面に対して配置を確認します。
- クライオスタットを使用して、セクション組織ポリ L リジンに 5 μ m の厚みでコーティング スライドです。
注: はひだや形態を歪めることができるティッシュ セクションの穴を確認します。 - クリオスタットから除去、修正スライド 4% PFA 10 分洗浄スライド dH2o. の 100 ml コンテナー 3 x 5 分
注意: 4 %pfa: 定着剤、ヒューム キャビネットで使用 - 、使用される抗体の量を最小限に抑えるには、疎水性ペンでセクションの周りを描画します。乾燥セクションをてはいけないです。
- ブロック ソリューション (3 %bsa と 1x PBS で 5% 正常な血清) 室温で 1 時間インキュベートします。
- ヒメアリ ブロック ソリューションを削除し、希釈した一次抗体 (ネフリン、podocin、または PECAM 1) 1: 250 (1 × PBS で 3 %bsa) でセクションを孵化させなさい。一晩、4 ° C で加湿チャンバーにスライドを配置します。湿気のある商工会議所が使用できない場合は、軽く抗体をコーティングしたスライドの上パラフィルムの小さなストリップを配置します。セクションを妨害しないよう次の日を削除するとき注意してください。
- 1x PBS でスライド 3 x 5 分を洗浄します。
- 適切な蛍光抗体希釈 1: 1000 (1 × PBS で 3 %bsa) 暗闇の中で部屋の温度で 2 時間で孵化させなさい。
- 1x PBS でスライド 3 x 5 分を洗浄します。蛍光 DAPI を含むメディアのマウントをマウントします。
- 糸球体を表示する 400 倍の倍率で蛍光顕微鏡でスライドをイメージします。これはバイアスを避けるために目がくらんでいるを確認します。
- 染色強度、糸球体領域と染色、すなわち、毛細血管ループ数のパターンに正規化を分析する使用 ImageJ は盲検の方法で糸球体領域に正規化します。
9. 蛋白質の抽出および西部にしみが付くこと
注: 西部のしみ腎疾患の調節不全に知られている式タンパク質を評価することができます。たとえば、podocin とネフリン発現の減少はポドサイトの損失を示します。
- 腎皮質およびふるわれた球体; から蛋白質を抽出します。プロトコルごとに同じで、組織量換散バッファーのボリュームを調整しました。
- NP 40 換散バッファー (150 mM NaCl、1% NP 40、50 ミリメートル トリス pH 8) 含んでいるプロテアーゼとホスファターゼ阻害剤を追加する前に、腎・糸球体氷の上を解凍します。30 のサンプルの均質 s。
- 氷上で 30 分間、一定の間隔で均質化されたサンプルをインキュベートします。
- 遠心分離機の 4 ° C で 15 分間 10,000 x g でサンプル
- 氷の上の新鮮なチューブに上清を除去します。
注: は、サンプルあたり約 1 mg タンパク質の回復を期待します。 - 標準的な 4 x Laemmli バッファーを使用して蛋白質を変化します。95 から 100 ° C で 5 分間混合物を沸騰させます。
- 腎糸球体細胞マーカー蛋白質(ネフリン、Podocin、PECAM 1 など)とリン酸化の表現とウエスタンブロット (を用いた疾患モデルの腎・糸球体に変更される知られている/仮説蛋白質の表現を評価します。標準的な方法。マフムードと陽15)。
注: プロトコルは、サイズや興味の蛋白質の豊富さによって異なります。
10. RNA の抽出およびポリメラーゼの連鎖反応 (PCR)
注: mRNA の発現解析遺伝子発現と選択的スプライシングの変化などの腎疾患の遺伝子を調整する方法を決定することができます。
- 腎皮質はまだ凍結されて、しながら徹底的に乳棒と乳鉢を使用してフェノール試薬 3 mL で挽きます。糸球体の抽出物を使用している場合フェノール試薬 1 mL を加えるし、30 のサンプルの均質 s。
注意: TRIzol 試薬: 刺激;ヒューム キャビネットで使用します。 - Chomczynski、サッキ16で説明したメソッドを使用して RNA の抽出を実行します。
注: 商業 RNA 抽出キット、この方法への代わりとして使用できます。 - 使用できるさまざまな方法の 1 つを使用して得られた RNA の質と量を評価します。RNA は避けようと-80 ° C で保存されますこの時点で。繰り返しを避けるため分かれるフリーズします。
注: このメソッドを新しい場合、は、クリア 28 s、18 s リボソーム バンドように agarose のゲルの RNA を実行する次の手順に進む前に RNA の品質をチェックします。このメソッドを使用して RNA の 2 に 5 μ g の間の回復を期待します。 - DNase は 37 ° C で 1 時間 RNA (RNase フリー水 10 μ L 加え DNase の 1 μ L、DNase バッファーの 1 μ L まで作るボリューム) の 1 μ g を扱う10 分の 65 ° c の DNase 停止液を 1 μ L で反応を停止します。
- オリゴ (dT) とランダムなプライマーの 0.5 μ L を追加します。70 の ° c で 10 分間インキュベートします。
- すぐに 5 分間氷の上を癒します。
- 以下を追加MMLV 逆転写酵素酵素 (400 U; DEPC H2O RT - コントロールのサンプルで置換) MMLV バッファー (1 x)、dNTP ミックス (0.5 mM)、リボヌクレアーゼ阻害剤 (40 U);DEPC 水で最大 50 μ L を作る。
- 95 ° C、5 分酵素を非アクティブ化する 1 h 37 ° C で反応混合物を孵化させなさい。
注: cDNA の高収率を生成するには、3 h までの 37 ° C で孵化させなさい。 - 使用できるさまざまな方法を使用して cDNA の質と量を評価します。
- 遺伝子の mRNA 発現とスプライシング パターンを評価するために PCR の使用は、糸球体疾患モデルの調節不全の仮説。プロトコルは興味の遺伝子によって異なります。
結果
尿は、野生型 (WT)、誘導ポドサイト固有 VEGF A ノックアウト (VEGF-A KO)、VEGF-A 島 X Neph VEGF165b マウスから代謝ケージを使用して収集した (表現、球体のヒト VEGF A165b アイソ フォーム VEGF-A KO マウス、構成方法)。VEGF-A KO のドキシサイクリン誘導後 0、4、10、14 週で尿中アルブミン クレアチニン比の測定、VEGF-A の KO マウスは WT 濾胞と比較して 10 週間によって進歩的な蛋白尿を開発しました。絶対値は、図 2 a、および図 2 bに各マウスの値の基準に正規化で見ることができます。しかし、VEGF-A X Neph の VEGF-A165b マウス (図 2) で蛋白尿は認めその VEGF の165を示す b は蛋白尿5モデルの保護。
糸球体 LpAV は私は WT、VEGF-A の島、VEGF-A X Neph の VEGF-A165b 腎臓からふるわれた個々 の糸球体で測定しました。例、糸球体のキャッチ方法と収縮の観察 8 %bsa を用いで3 a を図に表示される場合。この収縮し、糸球体の LpA を調べるため/V私の各糸球体 (図 3 b)。VEGF の KO マウスは、大幅に増加した糸球体 LpA/V私14 週間でポスト WT 制御糸球体と比較して VEGF KO 誘導。VEGF-A X Neph の VEGF-A165b マウス、高められた糸球体 LpA で下が 14 週5で VEGF A165b の過剰発現によって妨げられていない私V/。
PAS 染色腎臓皮質セクションの VEGF-A 島導入後 14 週 VEGF-A KO または VEGF-A X Neph の VEGF-A の165b マウス (図 4 a) 糸球体構造的な異常を明らかにしなかった。ただし、EM を介して糸球体微細構造の分析、VEGF-A の KO マウスいた増加の GBM 幅を開発、fenestrae 内皮細胞の数を減少、SPS カバレッジを減少、平均ポドサイトのスリット幅を増加 (図 4-4 階).平均ポドサイト足は、スリットの変わらずに残った (図 3B と 3 G) の幅と数を処理します。VEGF-A165b VEGF-A KO マウスでの過剰発現は、GBM への変更を防止し、スリット幅 (図 4 ・ 4 f)。ただし、VEGF-A165b には、変更された fenestrae 数と SPS カバレッジ (図 4 および 4E)5の影響はなかった。
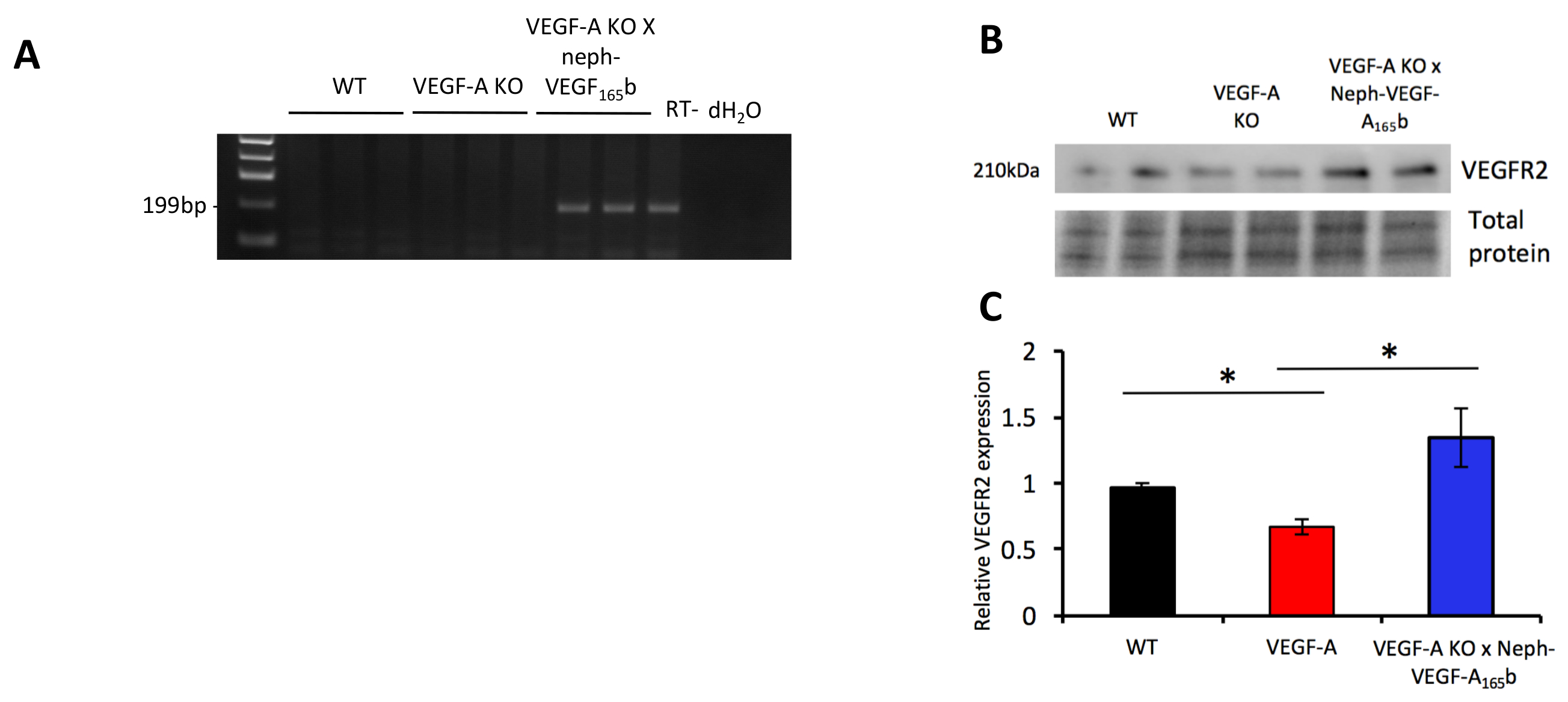
RT-PCR ふるわれた腎糸球体から抽出した RNA 上で実行は、ヒト VEGF A165b mRNA がのみに存在する (図 5 a) VEGF-A 島 X Neph-VEGF-A165b マウスを明らかにしました。VEGFR 2 糸球体蛋白発現は VEGF-A165b (の過剰発現によって防がれた VEGF-A KO マウスで減少する発見されたふるわれた腎糸球体および西部にしみが付くことによって蛋白質のレベルを評価するからタンパク質を抽出する場合図 5B , 5 C)5。

図 1。糸球体透過性 (LpA) リグの図式設定します。(A)糸球体がキャッチ安定性のためのマウントの上にクランプする(B)マイクロ ピペット吸引を使用してホルダー内に。(C)長方形 microslide。(D)ビデオ カメラで顕微鏡の 4 X の目的。(E) 1 %bsa 溶液 37 ° C に加温(F) 8 %bsa 溶液 37 ° C に加温(G)リモート タップ ベアリング 2 perifusate を含む行、急速な perifusate 交換を許可します。(H)し、糸球体を bathes microslide に向かって perifusate のルートです。この図の拡大版を表示するのにはここをクリックしてください。
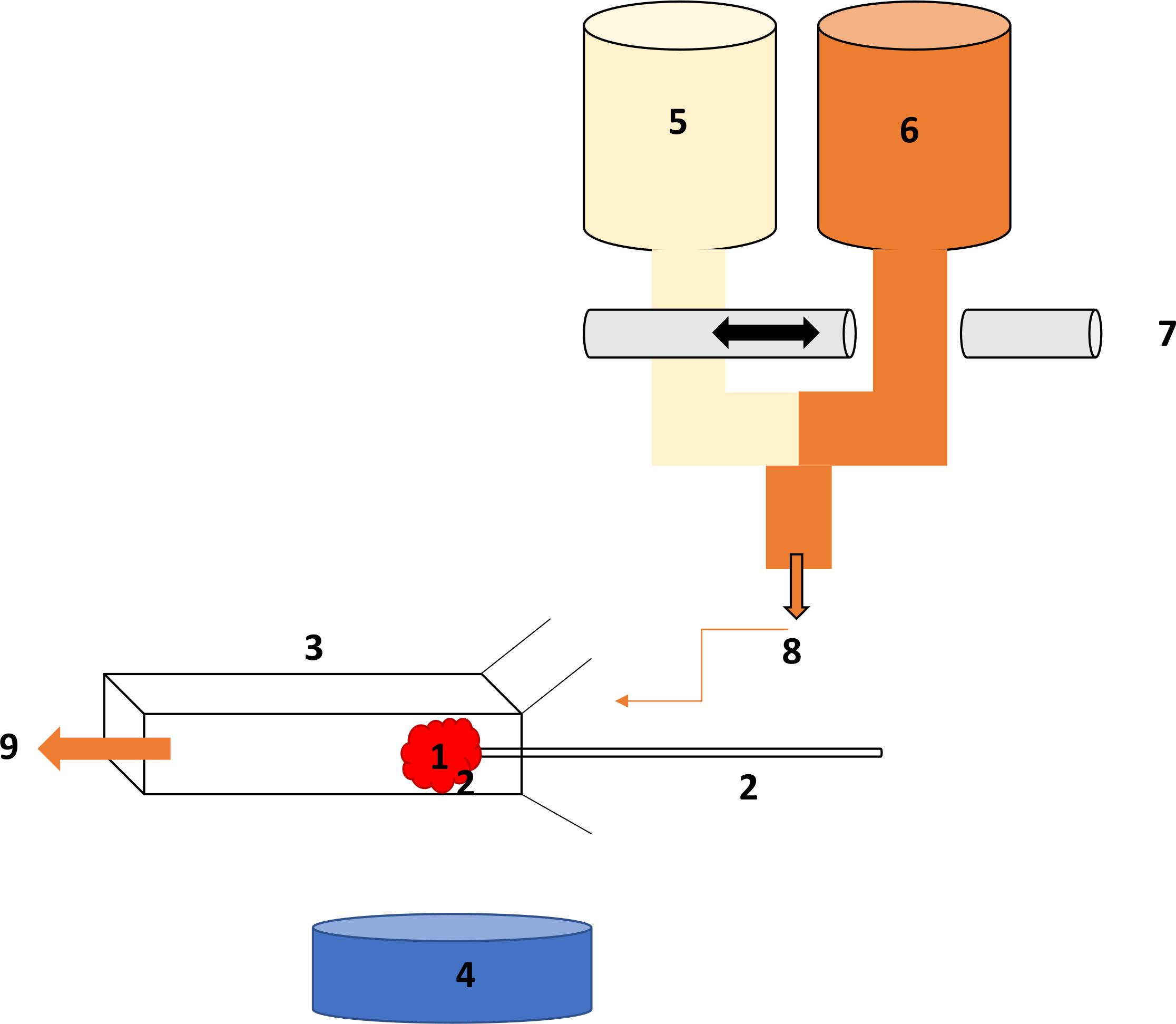{kind=link}

図 2。尿中アルブミン クレアチニン比。(A) WT、VEGF-A の島、VEGF-A X Neph の VEGF-A の165b マウスにおける VEGF-A の KO の誘導後 0、4、10、14 週間で uACR 値。(B)同じ uACR 値はそれぞれ個々 のマウスの基準値 (週 0) に正規化します。UACR 大幅増加 VEGF-A KO マウス週 10 と 14 WT 濾胞と比較して VEGF-A X Neph の VEGF-A165b マウスができなかった (* p < 0.05;二元、ペアの比較のための補正n = 4-12 マウス時間ポイントごと誤差範囲: [SEM] 平均値の標準誤差)。この図は、スティーブンスら5から変更されています。この図の拡大版を表示するのにはここをクリックしてください。
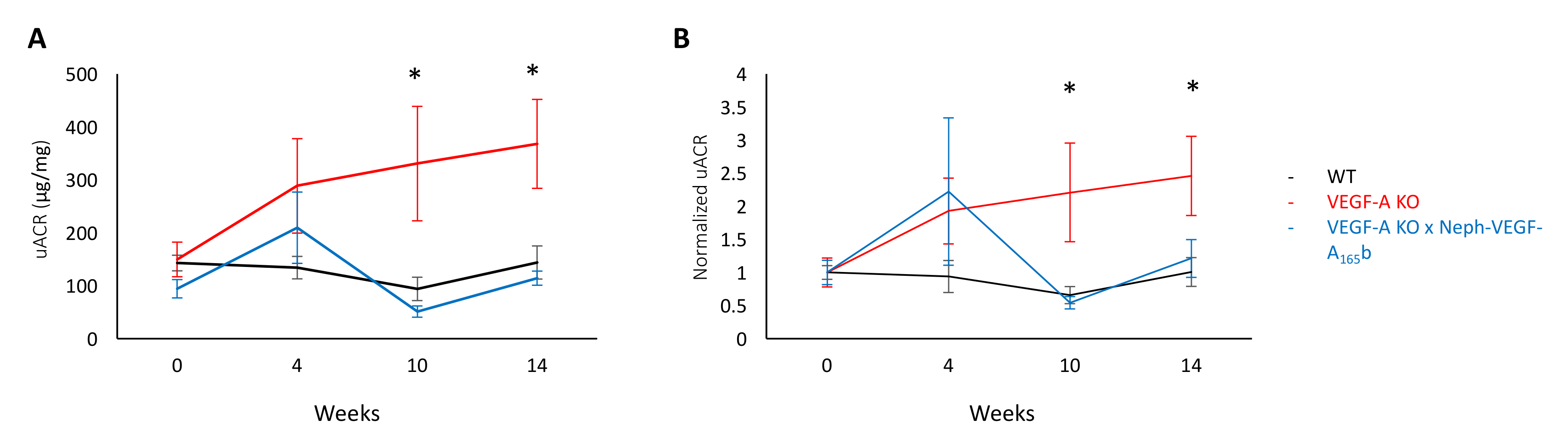{kind=link}

図 3。糸球体の水透過性の測定。(A)吸引; を介してマイクロ ピペットで糸球体をキャッチperifusate が 1 %bsa (Ai) から 8 %bsa (Aii) に切り替わるし、糸球体の収縮が観察されます。(B) 8 %bsa スイッチ前後の測定値決定される糸球体 LpA/V は私。(C) VEGF-A KO マウスを開発する高められた糸球体 LpA/V私14 週間でポスト VEGF-A KO WT コントロールと比較しての誘導。これは大幅 VEGF-A X Neph の VEGF-A165b マウスで妨げられない (* p < 0.05;一方通行 ANOVA、ボンフェローニ補正の組み合わせの比較n = 4-9 マウス, 15-30, 糸球体;誤差範囲: SEM)。この図の拡大版を表示するのにはここをクリックしてください。
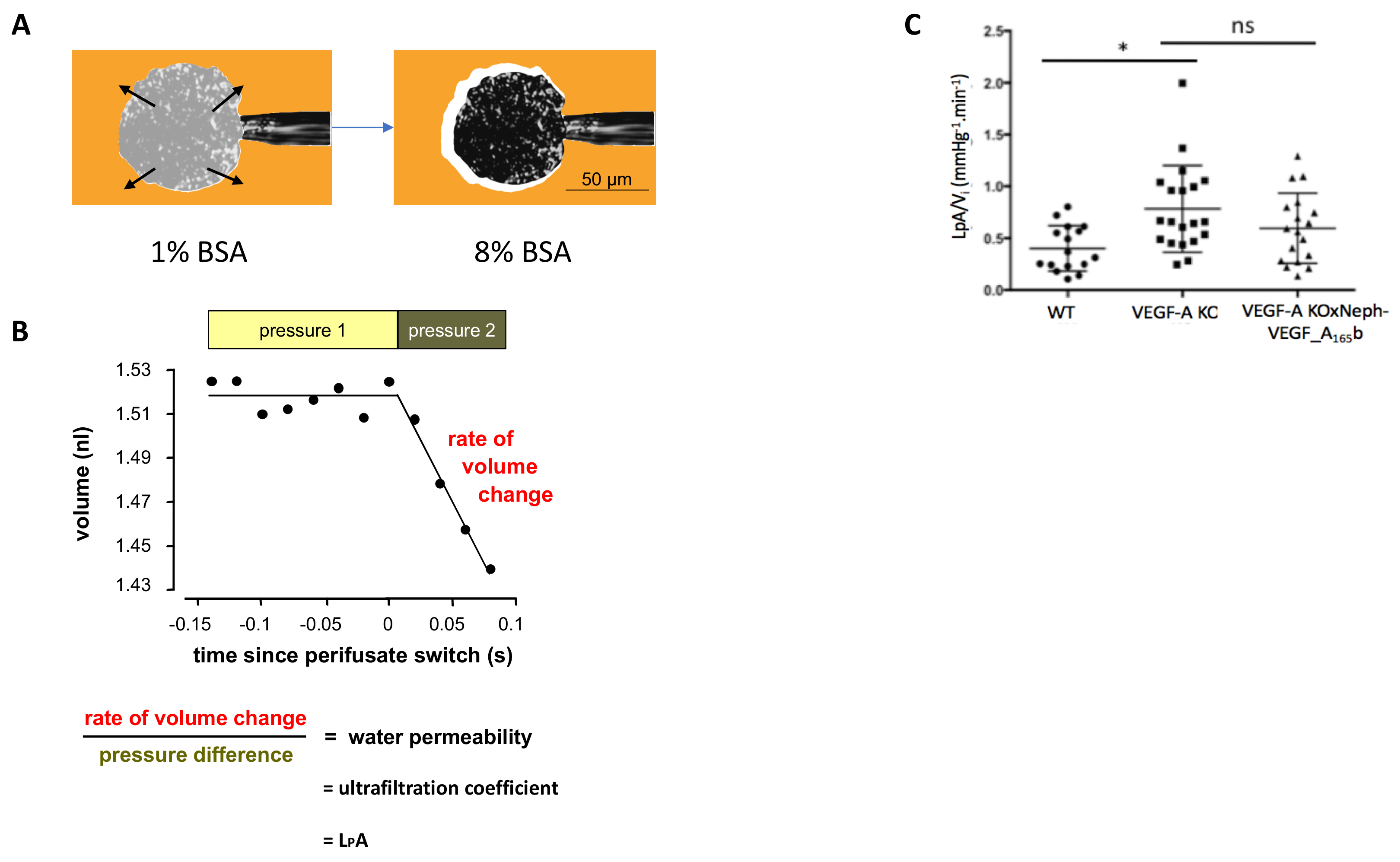{kind=link}

図 4。糸球体の構造解析。(A) PAS 陽性腎皮質は WT、VEGF-A の島、VEGF-A X Neph の VEGF-A165b マウス (Ai - iii) から糸球体の構造的な異常を示さなかった。EM は、VEGF-A 島糸球体 (Aiv - vi) の超構造異常を明らかにしました。(B)グループ間平均 FPW は変更しませんでした。(C)は糸球体基底膜増加 VEGF-A 島糸球体における VEGF A165b (D) fenestrae は VEGF-A165* (E) 、SPS 報道によって不変に残った VEGF-A 島糸球体の減少によって防がれました。いたも VEGF A165b (F)スリット幅平均は VEGF-A 島糸球体で増加したがそのまま残った、VEGF-A 島糸球体の減少を変更されなかった VEGF-A165b (G)スリット数によって防がれました。3 つのグループの間 (* p < 0.05;一方通行 ANOVA、ボンフェローニ補正の組み合わせの比較n = 3 のマウス, 9 糸球体;誤差範囲: SEM)。この図は、スティーブンスら5から変更されています。この図の拡大版を表示するのにはここをクリックしてください。
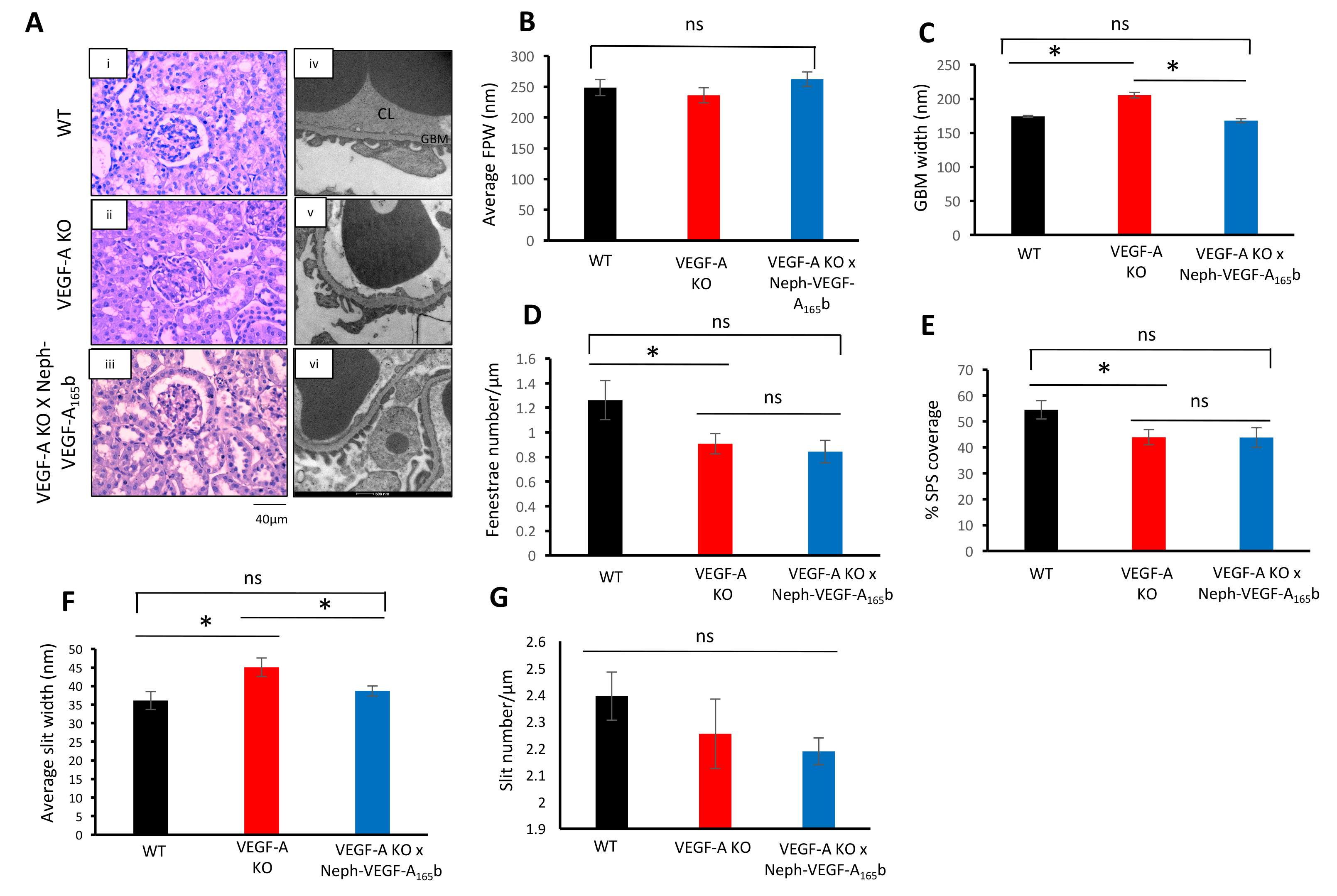{kind=link}

図 5。 マーカーの mRNA および蛋白質の表現。(A) RT-PCR ヒト VEGF A165b mRNA 発現のみ VEGF-A 島 X Neph-VEGF-A165b マウスからふるわれた球体で明らかにされたことを示した。(B)西部にしみが付くことを示した VEGFR 2 発現 VEGF-A 島糸球体でダウン規制 VEGF-A 島 X Neph-VEGF-A165b 糸球体で防がれた (* p < 0.05;一方通行 ANOVA、ボンフェローニ補正の組み合わせの比較n = 3-6 マウス;誤差範囲: SEM)。この図は、スティーブンスら5から変更されています。この図の拡大版を表示するのにはここをクリックしてください。
{kind=link}
ディスカッション
このプロトコルでは、糸球体疾患、腎臓および単一のマウスから得られる糸球体の機能に関する情報の膨大な量の有効化のマウス ・ モデルで実施する必要があります完全腎臓の仕事-アップについて説明します。腎臓の全体 (uACR および血漿クレアチニン測定)、透磁率の評価を含む、糸球体の機能の詳細な機能・構造・機構解析を可能にする各メソッドで重要な手順の透磁率個々 の球体 (糸球体 LpA/V は私)、構造変化 (PAS、ブルー、Trichrome、EM)、蛋白質のローカリゼーション (IF) と糸球体の遺伝子発現 (RT-PCR および西部にしみが付くこと) の検討。これらのメソッドは、腎疾患のマウスモデルの糸球体の機能を完全に把握するための鍵です。
GFB の透水性を評価する際、多くの研究は効果的な対策17,18として従来 uACR または 24 h アルブミン排泄率を使用するオプトインしています。これらのテクニックは、全体として GFB 透過性の評価をできるように、個々 の糸球体透過性評価と球体の間で変動のためそれはできません。以前の研究は、糸球体の LpA の測定を発見した/Vi GFB 透磁率5,9への変更のより敏感な指標となります。確かに、本稿で示した代表的な結果で 14 週に投稿 VEGF-A KO の誘導、VEGF-A 島 X Neph-VEGF-A165b マウス VEGF-A KO マウス; と比較して有意に低い uACRただし、この結果、糸球体の LpA に反映されません GFB 透磁率 (図 1および図 2)5V私計測、VEGF-A165b は大幅防ぐない増加/。これは腎臓の透水性と個々 の糸球体の透過性の両方を評価するために複数のアッセイを使用しての重要性を示しています。さらに、糸球体の LpA/V私oncometric アッセイは、同じ腎臓から個々 の糸球体の透過性が大きく異なることが、特に疾患モデル5,10,を示唆しています。19. 糸球体 LpA を測定する 1 つの制限/V私はそれすることができますのみ; 実験的なエンドポイントで実行されることしたがって、正規 uACR 測定実験のエンドポイントの指示を与える必要があります。
機能の表現型を評価するとともに本法はまた構造と微細構造の表現の評価を推奨します。これを行うことができます PAS、ヒアリンなど汚れやシルバーの汚れの選択を使用して各糸球体形態のさまざまな側面を評価します。これはしばしばマウス モデルの場合、糸球体疾患の急性のモデルでは、訓練された腎病理医でない限り、これらの汚れを使用して任意の主要な構造異常を検出することは困難することができます。したがって、EM の遂行が糸球体基底膜、内皮 fenestrae サイズと数、およびポドサイト特性などのパラメーターの定量化を可能にする GFB の微細構造を評価するために示唆されました。そのような測定を実行する最小限のトレーニングを必要とするでき、疾患モデルで影響を受けるセル-型/構造決定する捜査官。例では代表的な結果に VEGF-A KO マウスが発見された糸球体疾患の軽度のモデルであること、したがって、主要な構造異常が存在しない PAS 染色時に。ただし、ポドサイト固有 VEGF-A KO でした変化を誘発する糸球体基底膜、足細胞、内皮細胞糸球体の超構造5を調べるとき。残念なことに、EM 法に記載されているため、腎臓の準備 GFB19の透磁率に重大な影響があることも知られている内皮の糖衣の検出を有効にしません。糖衣の深さを正確に測定するために腎臓はずの内皮糖衣 1% アルシアン ブルーと 2.5% グルタルアルデヒド灌流固定 Olteanら19で説明されているようです。
機能的及び構造的表現型が評価され、一度異なる遺伝子および細道の式/アクティベーション パターン、糸球体では特に評価できます。事前の超構造評価の種類/糸球体構造関係、ポドサイトまたは内皮特異遺伝子/経路を検討すべきかどうかを示すセルに関するいくつかの情報を与えることができます。たとえば、VEGF-A KO マウスから代表的な結果、内皮 fenestrae 数の減少は、(図 3 D) を認められた;したがって、VEGF の経路に関与すると知られている血管内皮マーカーの糸球体蛋白発現を検討しました。VEGFR 2 (図 4 b)5。糸球体のタンパクの発現、に加えて局在も場合を使用して視覚化することができます。張ら20による調査で、ポドサイト固有 GLUT1 の発現は、共同 podocin と増加の GLUT1 をローカライズする場合、足で確認されました。
糸球体の機能を評価するために文献の提示の方法と比較して糸球体疾患のマウスモデルの腎機能を評価するために本稿で説明した方法を使用できますから完全に評価する糸球体の表現型複数の側面。このメソッドを使用して、研究者はモデルの腎臓の表現型を決定し、表現型を開発する理由とメカニズムを評価することができます。病気のメカニズムに重要な情報は、これらのモデルの潜在的な治療手段を調べる場合に必要です。このメソッドは、疾患表現型と潜在的な治療の評価における糸球体機能に将来の調査に簡単に適用できます。
結論としては、このジェネリックと適応プロトコルは、腎臓および単一のマウスから得られる糸球体の機能に関する情報の膨大な量の有効化、糸球体疾患のマウスモデルの仕事アップにおける腎臓をについて説明します。糸球体疾患のすべてのマウスのモデルに適用することができます糸球体機能の詳細な機能・構造・機構解析を可能にするメソッド。
開示事項
著者が明らかに何もありません。
謝辞
この作品は、英国心臓財団、リチャード ・ ブライト VEGF 研究トラストと MRC によって支持されました。
資料
| Name | Company | Catalog Number | Comments |
| Metabolic Cages | Harvard Apparatus | 52-6731 | |
| Tris buffered saline (10x) | Sigma-Aldrich | T5912-1L | |
| Bovine Serum Albumin | Sigma-Aldrich | A2058 | |
| Mouse Albumin ELISA Quantitation Set | Bethyl Laboratories | E90-134 | |
| TMB ELISA Substrate solution | ThermoFisher Scientific | 34028 | |
| Sulphuric acid | Sigma-Aldrich | 339741 | |
| SPECTRstar nano | BMG Labtech | SPECTRstar nano or equivalent | |
| RNAlater stabilisation solution | ThermoFisher Scientific | AM7020 | |
| 4-15% precast protein gel | BIORAD | 4568084 | |
| 4x Laaemmli Sample buffer | BIORAD | 161-0747 | |
| Mini Trans-Blot cell | BIORAD | 1703930 | |
| 10x Tris running buffer | BIORAD | 1610732 | |
| Coomassie Brilliant Blue Dye | ThermoFisher Scientific | 20278 | |
| Creatinine Companion | Exocell | 1012 Strip Plate | |
| Glutaraldehyde Solution | Sigma-Aldrich | G5882 | |
| Sodium Cacodylate | Sigma-Aldrich | C0250 | |
| 10x Phosphate Buffered Saline | ThermoFisher Scientific | AM9625 | |
| Sodium Chloride | Sigma-Aldrich | S7653 | |
| Sodium Acetate | Sigma-Aldrich | S2889 | |
| Sodium Phosphate | Sigma-Aldrich | 342483 | |
| Sodium Bicarbonate | Sigma-Aldrich | S5761 | |
| Magnesium Sulfate | Sigma-Aldrich | M2643 | |
| Calcium Chloride | Sigma-Aldrich | C5670 | |
| D(+)Glucose | Sigma-Aldrich | G8270 | |
| EDTA Blood Collection tubes | BD | 367835 | |
| 23-25G Needle | BD | PMC0735 | |
| EDTA | Sigma-Aldrich | E9884 | |
| 10 ml Glass Vial | Thomas Scientific | 0914X10 | |
| Falcon 10 ml Polypropylene Tubes | ThermoFisher Scientific | 10110101 | |
| 0.5 ml Tubes | ThermoFisher Scientific | 10681894 | |
| Disposable Tissue Molds | ThermoFisher Scientific | 22-363-553 | |
| Mouse Surgical Disection Kit | ThermoFisher Scientific | 13-005-204 | |
| Optimal Cutting Medium | ThermoFisher Scientific | 23-730-571 | |
| 4% Paraformaldehyde | ThermoFisher Scientific | AAJ19943K2 | |
| Glass Capillary Tubes | Harvard Apparatus | EC1 64-0770 | |
| Glomerular Permeability Rig | Built at the Univeristy of Bristol - not comercially available | Citation of LpA rig: Salmon et al. 2006; J. Physiol | |
| Stainless Steel Sieves | Cole-Parmer | WZ-59984 | |
| Periodic Acid-Schiff (PAS) Staining System | Sigma-Aldrich | 395B-1KT | |
| Hematoxylin | Sigma-Aldrich | H3136 | |
| Xylene | MerckMillipore | 108298 | |
| Poly-Prep Slides | Sigma-Aldrich | P0425-72EA | |
| Mounting Medium | ThermoFisher Scientific | 8030 | |
| Osmium tetroxide solution | Sigma-Aldrich | 75632 | |
| Aradite Resin | Agar Scientific | CY212 | |
| Uranyl Acetate | Agar Scientific | AGR1260A | |
| Lead Citrate | Agar Scientific | AGR1210 | |
| Cryostat | ThermoFisher Scientific | e.g. 957000H | |
| Hydrophobic Pen | Abcam | ab2601 | |
| Nephrin (1243-1256) Antibody | Acris | BP5030 | |
| Anti-Podocin | Sigma-Aldrich | P0372-200UL | |
| Anti-CD31 | BD Biosciences | 550274 | |
| Alexa Fluor Secondary Antibody | ThermoFisher Scientific | A32732 | |
| Vectashield Mounting Medium with DAPI | Vector Labs | H-1200 | |
| NP40 Cell Lysis Buffer | ThermoFisher Scientific | FNN0021 | |
| Halt Protease and Phosphatase Inhibitor Cocktail | ThermoFisher Scientific | 78437X4 | |
| 10x Transfer Buffer | BIORAD | 1610734 | |
| PVDF Membrane | ThermoFisher Scientific | LC2002 | |
| HRP-Conjugated Secondary Antibodies | Abcam | ab6721 | |
| ECF Substrate for Western Blotting | Fisher | 10713387 | |
| TRIzol | ThermoFisher Scientific | 15596018 | |
| Dnase I | New England Biolabs | M0303S | |
| M-MLV Reverse Transcriptase | New England Biolabs | M053S | |
| Oligo dT | ThermoFisher Scientific | 18418012 | |
| Random Primers | ThermoFisher Scientific | 48190011 | |
| dNTP | ThermoFisher Scientific | 18427088 | |
| Ribonuclease Inhibitor | ThermoFisher Scientific | 10777019 | |
| DEPC Water | ThermoFisher Scientific | AM9915G | |
| Fluorescent Light Miscroscope | Leica Microsystems | ||
| Image J Analysis Software | Image J | ||
| PCR Thermocycler | ThermoFisher Scientific | ||
| TEM Microscope | Britannica |
参考文献
- Humtstrom, M. Development of structural kidney damage in spontaneously hypertensive rats. Journal of Hypertension. 30 (6), 1087-1091 (2012).
- Kitada, M., Ogura, Y., Koya, D. Rodent models of diabetic nephropathy: their utility and limitations. International Journal of Nephrology and Renovascular Disease. 14 (9), 279-290 (2016).
- Tesch, G. H., Lim, A. K. Recent insights into diabetic renal injury form the db/db mouse model of type 2 diabetic nephropathy. American Journal of Physiology-Renal Physiology. 300 (2), F301-F310 (2011).
- Fogo, A. B. Animal models of FSGS: lessons for pathogenesis and treatment. Seminars in Nephrology. 23 (2), 161-171 (2003).
- Stevens, M., et al. VEGF-A165b protects against proteinuria in a mouse model with progressive depletion of all endogenous VEGF-A splice isoforms from the kidney. Journal of Physiology. 595 (19), 6281-6298 (2017).
- Cosgrove, D., Kalluri, R., Miner, J. H., Segal, Y., Borza, D. B. Choosing a mouse model to study the molecular pathobiology of Alport glomerulonephritis. Kindey International. 71 (7), 615-618 (2007).
- Kujal, P., Vernerova, Z. 5/6 nephrectomy as an experimental model of chronic renal failure and adaptation to reduced nephron number. Ceskoslenska Fysiologuie. 57 (4), 104-109 (2008).
- Chevalier, R. L., Forbes, M. S., Thornhill, B. A. Ureteral obstruction as a model of renal interstitial fibrosis and obstructive nephropathy. Kidney International. 75 (11), 1145-1152 (2009).
- Oltean, S., et al. VEGF165b overexpression restores normal glomerular water permeability in VEGF164-overexpressing adult mice. American Journal of Physiology-Renal Physiology. 303 (7), F1026-F1036 (2012).
- Salmon, A. H., Neal, C. R., Bates, D. O., Harper, S. J. Vascular endothelial growth factor increases the ultrafiltration coefficient in isolated intact Wistar rat glomeruli. Journal of Physiology. 570 (Pt 1), 141-156 (2006).
- Zheng, F., Striker, G. E., Esposito, C., Lupia, E., Striker, L. J. Strain differences rather than hyperglycemia determine the severity of glomerulosclerosis in mice. Kidney International. 54 (6), 1999-2007 (1998).
- Betz, B., Conway, B. R. An update on the use of animal models in diabetic nephropathy research. Current Diabetes Reports. 16 (18), (2016).
- Savin, V. J., Terreros, D. A. Filtration in single isolated mammalian glomeruli. Kidney International. 20 (2), 188-197 (1981).
- Neal, C. R., Crook, H., Bell, E., Harper, S. J., Bates, D. O. Three-dimensional reconstruction of glomeruli by electron microscopy reveals a distinct restrictive urinary subpodocyte space. Journal of the American Society of Nephrology. 16 (5), 1223-1235 (2005).
- Mahmood, T., Yang, P. C. Western blot: technique, theory, and trouble shooting. North American Journal of Medicine & Science. 4 (9), 429-434 (2012).
- Chomczynski, P., Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Analytical Biochemistry. 162 (1), 156-159 (1987).
- Adembri, C., et al. Sepsis induces albuminuria and alterations in the glomerular filtration barrier: a morphofunctional study in the rat. Critical Care. 15 (6), R277 (2011).
- Ma, S. T., Liu, D. L., Deng, J. J., Niu, R., Liu, R. B. Effect of arctiin on glomerular filtration barrier damage in STZ-induced diabetic nephropathy rats. Phytotherapy Research. 27 (10), 1474-1480 (2013).
- Oltean, S., et al. Vascular endothelial growth factor-A165b is protective and restores endothelial glycocalyx in diabetic nephropathy. Journal of the American Society of Nephrology. 26 (8), 1889-1904 (2015).
- Zhang, H., et al. Podocyte-specific overexpression of GLUT1 surprisingly reduces mesangial matric expansion in diabetic nephropathy in mice. American Journal of Physiology-Renal Physiology. 299 (1), F91-F98 (2010).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved