Method Article
Оценка функции почек в моделях мыши клубочковых болезни
В этой статье
Резюме
Этот протокол описывает полный почек работы вверх, которая должна осуществляться в моделях мыши клубочковых болезни. Методы позволяют для детального анализа функциональной, структурной и механистические клубочковых функции, которые могут быть применены ко всем моделям мыши клубочковых болезни.
Аннотация
Использование мышиных моделей имитировать человека почек становится все более распространенным. Наши исследования сосредоточены на оценке клубочковых функции в диабетической нефропатии и Подоцит конкретных VEGF-A-нокаут мышей; Таким образом этот протокол описывает полный почек работы вверх, используемые в нашей лаборатории для оценки этих моделей мыши клубочковых заболеваний, что позволяет огромное количество информации, касающейся почек и клубочковых функции будут получены из одной мыши. По сравнению с альтернативными методами, представлены в литературе для оценки клубочковых функции использование метода, описанного в настоящем документе позволяет клубочковых фенотип полностью оцениваться от нескольких аспектов. С помощью этого метода, исследователь может определить почек фенотип модели и оценить механизм о том, почему фенотипа развивается. Это жизненно важную информацию о механизме заболевания требуется при рассмотрении потенциальных терапевтических направлений в этих моделях. Методы позволяют для детальной функциональной оценки клубочковой фильтрации барьера путем измерения креатинина соотношение альбумин мочевыделительной и отдельных клубочковых водопроницаемости, а также как структурные, так и ультра-структурного обследования Использование периодической кислоты Шифф пятно и электронной микроскопии. Кроме того анализ генов dysregulated на уровне мРНК и белка позволяет механистический анализ клубочковых функции. Этот протокол определяет общий, но гибкой методы, которые могут быть применены ко всем моделям мыши клубочковых болезни.
Введение
Использование мышиных моделей имитировать человека почек становится все более распространенным. Такие мышиных модели включают спонтанные модели, такие как спонтанно гипертензивных крыс (ШРМ)1, Стрептозотоцин (СТЗ)-индуцированной сахарным диабетом крыс и мышей2, и/дБ тип II диабетических мышей3, генетически модифицированных моделей, таких как первичный Подоцит конкретных координационных сегментарный клубочковых склероз (FSGS) модели4, Подоцит конкретных Сосудистый эндотелиальный фактор роста ((VEGF-A)-нокаут VEGF-A KO) модель5, и синдром Алпорт моделей6и приобрела такие модели, как 5/6 нефрэктомия7 и односторонние обструкции мочеточника (UUO) модель8. Для того чтобы оценить различные аспекты клубочковых функции в этих моделях, доступны несколько методов. Цель этого документа метод является продемонстрировать всеобъемлющую работу вверх, которая должна быть выполнена в мыши модели болезни почек для того, чтобы полностью оценить клубочковых функции.
Обоснование использования этого метода является, что он позволяет клубочковых фенотип полностью оцениваться от нескольких аспектов. Это включает в себя оценку клубочковых проницаемости, как белок, так и воду, клубочковых структурные аномалии и изменения в выражение/сплайсинга мРНК и белков, необходимых для нормальной клубочковых функции. С помощью этого метода, исследователь может определить почек фенотип модели и оценить механизм о том, почему фенотипа развивается. Это жизненно важная информация о механизме заболевания, которое является обязательным при рассмотрении потенциальных терапевтических направлений в этих моделях.
В литературе это обычным явлением будет представлена модель мыши клубочковых болезни, где фенотипа определяется повышенный уровень альбумина в моче. Однако есть основания полагать, что один метод для определения клубочковых функция не всегда является эффективным; только измерение уровень экскреции альбумина мочи или соотношение альбумин мочевыделительной креатинина (uACR) предоставляет информацию, всего почечной функции, а не отдельных клубочков. Предыдущие исследования показали, что проницаемость может варьироваться в разных клубочков же почек5,9,10. Кроме того Оценка проницаемости отдельных клубочков является более чувствительным способом оценки клубочковых функции; Методика измерения индивидуальных клубочковых водопроницаемость (LpA / V,я) показал, чтобы быть более чувствительны к изменениям в клубочковых функции, чем измерения uACR9. Этот assay выгодно в мыши модели, которые устойчивы к протеинурии, например тех, на фоне c57BL/611. Преимуществом настоящего документа метод является, что она рассматривает как общая почечной проницаемость для альбумина, так и отдельных клубочковых проницаемости для воды.
Экспертиза клубочковых структурные аномалии часто оценивается батареи пятна такие периодические кислоты Шифф (ССА), trichrome, и серебряные пятна. Они позволяют патологоанатом подготовленных почек для оценки уровня почечной болезни через метод скоринга. Хотя все хорошие методы, изменения в структуре клубочковых макрос не всегда соблюдаются в острой почечной травмы модели12. Этот метод предлагает, что помимо проведения почечной гистологии методы, описанные выше, клубочковых ультра структуры также должны оцениваться через электронной микроскопии (EM). Окрашенных glomerulus может выглядеть относительно нормальной под микроскопом регулярных света; Однако после оценки с EM, небольшие изменения в ширину клубочковых базальной мембраны (GBM), анализируется Подоцит фут процесс самоотречение, эндотелиальные ярусе и охват пространства суб Подоцит. Таким образом важно, что клубочковых ультра структура и микро структура оценивается для определения механизма клубочковых дисфункции.
Помимо оценки клубочковых структурные аномалии, изменения в мРНК и выражения протеина и сращивания, а также активации белков (например, фосфорилирование), должны рассматриваться для дальнейшего выяснения механизмов клубочковых болезни. При взгляде на клубочковых болезнь, или, например, когда KO/над-expressing ген специально в клубочковых клеток, таких, как Подоцит конкретных VEGF-A KO мыши5, важно, что белок и мРНК изменения рассматриваются только в пределах клубочковых клеток, а не весь почек. Этот протокол описывает метод в котором клубочков изолированы от мыши почечной коры, и затем РНК белками изолированы. Это позволяет конкретного анализа белка/mRNA dysregulation клубочков модели болезни.
Этот протокол описывает полный почек работы вверх, которая должна осуществляться в моделях мыши клубочковых заболеваний, что позволяет огромное количество информации, касающейся почек и клубочковых функции будут получены из одной мыши. Методы позволяют для детального анализа функциональной, структурной и механистические клубочковых функции, которые могут быть применены ко всем моделям мыши клубочковых болезни.
протокол
Все эксперименты проводились в соответствии с законодательством Великобритании и местные этического комитета утверждения. Исследования на животных были утверждены университет Бристоль Комитета по этике исследований.
1. моче альбумина креатинина соотношение (uACR)
Примечание: UACR используется для оценки проницаемость GFB альбумина. Наличие альбумина в моче указывает на повышение проницаемости через GFB, который является нормализованным креатинина в элемент управления для изменения скорости потока мочи. Альбуминурия является общим маркером для хронической болезнью почек.
- Сбор мочи на экспериментальной базовой линии и на регулярной основе (один раз в неделю на один раз в месяц) вплоть до экспериментальной конечной точки.
- Настройка мыши метаболизм клеток с водой и обогащение диеты. Место мыши (мужчины, в возрасте 6-8 недель) в отдельных клетках для 6 h в тихой комнате.
- Возвращение мышей регулярные жилье и сбора мочи из пустых клеток. Требуется минимум 50 мкл.
Примечание: Если мышь не производят мочи в данное время, повторите на другой день в комнате с подогревом. - Центрифуга мочи на 500 x g 10 мин сбор мочи и сохранить осадков для оценки потери Подоцит. На данный момент магазин мочи при-20 ° C в краткосрочной перспективе.
- Разбавьте мочу в 1% бычьим сывороточным альбумином (БСА) в 1 x Tris буфер солевой раствор (рН 7,5 TBS) на 1: 500 для разбавления 1: 10000, в зависимости от тяжести альбуминурии.
Примечание: Объем конец должен быть > 400 мкл. оптимизировать определить правильный разрежения на каждом точку. - Количественного определения концентрации альбумина мочи с помощью мыши альбумина энзим связанный assay иммуносорбента (ELISA) в инструкции производителя.
- Вкратце пальто ELISA плита, которая оптимизирована для связывания белков, с 100 мкл основное антитело против мыши альбумина (10 мкг/мл 0,5 М карбонат бикарбонат рН 9,6) за 1 ч при комнатной температуре.
- Избыточные антитела мыть из скважин пять раз с 1 x TBS плюс 0,05% анимации (рН 8,0) и 200 мкл преграждая разрешение (1 x TBS с 1% BSA pH 8.0) на ночь при комнатной температуре.
- Вымойте тарелка пять раз и 100 мкл стандартов (последовательного растворения: 2000 мкг/мл 15.63 мкг/мл в 1 x TBS с 1% BSA, рН 8,0), пустые и разбавленных образцы в трех экземплярах. Оставить на 1 час при комнатной температуре, затем промойте пластины пять раз.
- 100 мкл HRP обнаружения антител для каждой скважины (10 нг/мл в 1xTBS с 1% BSA, рН 8.0) и инкубации при комнатной температуре в течение 1 ч.
- Вымойте тарелку пять раз и 100 мкл раствора субстрата энзима в каждой скважине. Оставьте пластину в темноте, чтобы разработать для 15 мин и остановить реакции, добавив 100 мкл 0,18 М серной кислоты (H2т-4).
Предупреждение: H2так4 коррозионные.
- Определить концентрацию альбумина каждого образца, читая пластину на поглощение 450 Нм. Использование стандартной кривой для количественного определения альбумина, присутствующих в каждом образце. Если технические повторяет значение больше, чем 5% CV, повторите пробу для этих образцов.
- Кроме того оцените концентрацию альбумина мочи с помощью электрофореза. 5 мкл 4 x буфер загрузки образца протеина, чтобы 15 мкл мочи. Тепла образцов до 95 ° C 10 мин и затем загрузить в гель трис сборный 4-12%.
- Побегите гель на 100 V и пятно на ночь Кумасси синим за производителя протокол.
- После визуализации гель, используйте денситометрия оценить концентрацию альбумина изменения раза относительно элемента управления.
Примечание: Если не полосы наблюдаются для альбумина, когда ожидалось, оценить концентрацию белка мочи и скорректировать объем мочи, добавляется гель.
- Разбавьте сырье мочи в dH2O 1:1, 1:5 и 1:10.
Примечание: Объем конец должен быть > 70 мкл. оптимизировать определить правильный разбавления каждого образца. - Количественно с помощью химического анализа в комплект инструкций концентрация мочи креатинина. Вкратце Загрузите 20 мкл креатинина стандартов, пустой, или разбавленных мочи до 96 хорошо пластины в трех экземплярах.
- Определение концентрации креатинина каждого образца, читая пластину на поглощение 490 Нм до и после добавления раствора кислоты (осторожно, коррозионные; избегать контакта с кожей).
Примечание: Разница между значениями оптической плотности прямо пропорциональна концентрации креатинина в каждом образце. Калибровочной кривой это поколение от стандартов. Если технические повторяет значение больше, чем 5% CV, повторите пробу для этих образцов. - Создание uACR (мкг/мг). Нормализовать данные базовой стоимости каждой мыши для графического представления.
2. ткани и крови коллекции
Примечание: Почки и клубочковых ткани может использоваться для оценки структурных, белок и мРНК выражение маркеры почечной болезни. Кровь может использоваться для оценки маркеры почечной функции, такие как креатинин, который может быть вверх регулируется в почечной недостаточности, указывающее уменьшение возможностей фильтрации клубочков.
- Подготовить следующие решения: свежие глютаральдегид 2,5% в 0,1 М натрия cacodylate (рН 7.3), параформальдегида 4% в 1 x фосфатный буфер (PBS), раствор Рингера млекопитающих (115 мм хлорида натрия (NaCl), ацетат натрия 10 мм (CH3COONa), 1.2 Мм фосфат натрия (Na2HPO4), бикарбонат натрия 25 мм (3NaHCO), сульфат магния 1,2 мм (4MgSO), хлорид кальция 1 мм (2CaCl), 5,5 мм D (+) глюкозы, рН 7,4) с 1% BSA и ПБС.
Предупреждение: 2,5% глютаральдегид: токсичные, сенсибилизатор, раздражающее; Используйте вытяжной шкаф. 0,1 cacodylate натрия М: токсичные, использование в вытяжной шкаф. 4% ПФА: фиксатор, использование в вытяжной шкаф - Подготовьте следующие материалы: изофлюрановая, небольшой Этилендиаминтетрауксусная кислота (ЭДТА)-покрытием Трубы крови, 23-25G иглы, шприцы 5 мл ЭДТА покрытием, стеклянные флаконы по 10 мл, Пластиковые флаконы по 10 мл, 0,5 мл пластиковых труб, располагаемого ткани плесени, сухого льда, жидкость N 2, мышь хирургические инструменты и оптимального раскроя среднего (OCT).
- Поместите курсор мыши под глубокой анестезии, которая проверяется мышь, не отвечающих на укол иглой для ног колодки, используя камеру изофлюрановая, или эквивалентные маршруты терминала анестезии как инъекции анестетиков (Пентобарбитал, 50 мг/кг Внутрибрюшинное [IP]; ««Авертен, IP 240 мг/кг), или воздействия диоксида углерода (CO2) (75% CO2/25% O2).
- Отбирать мыши через сердца прокол в левый желудочек и собирать столько крови, как это возможно. Переезд в ЭДТА покрытием крови трубки для до 4 ч. Если предпочтительнее, мышей может взятые через шейки матки дислокации с осторожностью, чтобы не разорваться яремной вены.
- Вскрыть из почки через живот и вымыть в холодной ПБС льда.
- Для изучения корковых клубочков, удалить один полюс почки коры и разрезать на3 части 1 мм. Для изучения глубокие juxta Медуллярная клубочков, повторите ту же технику с ткани из продолговатого мозга. Место в 5 мл 2,5% раствора глютаральдегид во флаконе EM стекла. Хранить при 4 ° C.
Предупреждение: 2,5% раствор глютаральдегид: токсичные, сенсибилизатор, раздражающее; использование в вытяжной шкаф
Примечание: процесс в течение 1 месяца для достижения наилучших результатов. - Для гистологии, удалить верхней трети почек, обеспечить коркового и мозгового juxta клубочков будет присутствовать и исправить в 5 мл параформальдегида 4% при температуре 4 ° C для 24 h. передачи до 5 мл 70% EtOH за 24 часа до встраивания в парафин.
Предупреждение: 4% ПФА: фиксатор, использование в вытяжной шкаф - Для иммунофлюоресценции, место треть почки, чтобы обеспечить коркового и мозгового клубочков будет присутствовать в ткани плесени и пальто в октября место на сухой лед заморозить и хранить при температуре-80 ° C.
- Для белка и РНК место 3 x 2 мм3 куски коры почек в 0,5 мл пластиковых трубок и оснастки заморозить в жидкости N2. Хранить при температуре-80 ° C. Для длительного хранения тканей для РНК место ткани в 5 томах РНК стабилизации раствора и хранить при температуре-80 ° C.
- Для изоляции клубочков нарезать остальные ткани почек и место в 5 мл раствора Рингера млекопитающих с 1% BSA на льду. Подготовка к сита клубочков немедленно.
3. плазмы креатинина
Примечание: Креатинина плазмы может быть вверх регулируется в почечной недостаточности, указывающее уменьшение возможностей фильтрации клубочков. Азота (БУЛОЧКИ) уровни мочевины в крови также могут быть оценены, хотя протокол не описано здесь.
- Центрифугуйте образцы крови на 500 g x 15 мин при 4 ° C.
- Сбор плазмы, которые могут храниться при температуре-20 ° C в краткосрочной перспективе на данный момент.
- Количественно с помощью химических креатинина пробирного инструкции выше для мочи креатинина в протоколе 1.11 концентрация креатинина плазмы.
- Определение концентрации креатинина каждого образца, читая пластину на поглощение 490 Нм до и после добавления раствора кислоты.
Примечание: Разница между этими значениями оптической плотности прямо пропорциональна концентрации креатинина в каждом образце. Калибровочной кривой это поколение от стандартов. Если технические повторяет значение больше, чем 5% CV, повторите пробу для этих образцов.
4. изоляция клубочков
Примечание: Клубочков могут быть изолированы для оценки проницаемость отдельных клубочков ex vivo, а также выражение определенного белка и мРНК маркеры клубочковых болезни.
- Возьмите почечной ткани, помещается в раствор Рингера-млекопитающих с 1% BSA и вскрыть клубочков, с помощью стандартного просеивание техника13. Вкратце стек 70 мкм (внизу), 100 мкм, 125 мкм, 175 мкм, 250 мкм и 425 мкм (сверху) сита на стеклянный стакан.
- Размять почки, используя поршень шприца, в 425 мкм сита и протолкнуть, используя лед холодной млекопитающих Рингера раствор с 1% BSA. Как протолкнула биты почки, удалить верхнего сита и приступить к делать то же самое на следующий. Повторите, пока только 100 мкм и 70 мкм сита остаются.
- Передача клубочковых урожай, удерживается 100 мкм и 70 мкм сита до 10 мл свежего млекопитающих Рингера раствор с 1% BSA, на льду.
Примечание: Если количество клубочков в раствор Рингера мл несколько, сократить объем раствора Рингера, используемые для сбора клубочков от последних двух сита. - Удаление 5 мл раствора, содержащего клубочков в два отдельных труб (по 2,5 мл) и центрифуги на 1000 x g 10 мин при 4 ° C. Удалить супернатант и оснастки заморозить клубочков в жидкости N2 прежде чем убрать на-80 ° C для белков и РНК добыча на более поздний срок.
- Поместите оставшийся раствор, содержащий клубочков в водяной бане при 37 ° C для измерения клубочковых LpA / V,яex vivo. Завершите в течение 3 ч удаления почки.
5. клубочковых водопроницаемость (LpA / V,я)
Примечание: Клубочковых LpA / Vя проба позволяет ex vivo измерения проницаемости отдельных клубочков в быстром воспроизводимый манере. Увеличение клубочковых LpA / V,я указывает нарушение GFB, который наводит на мысль о почечной болезни.
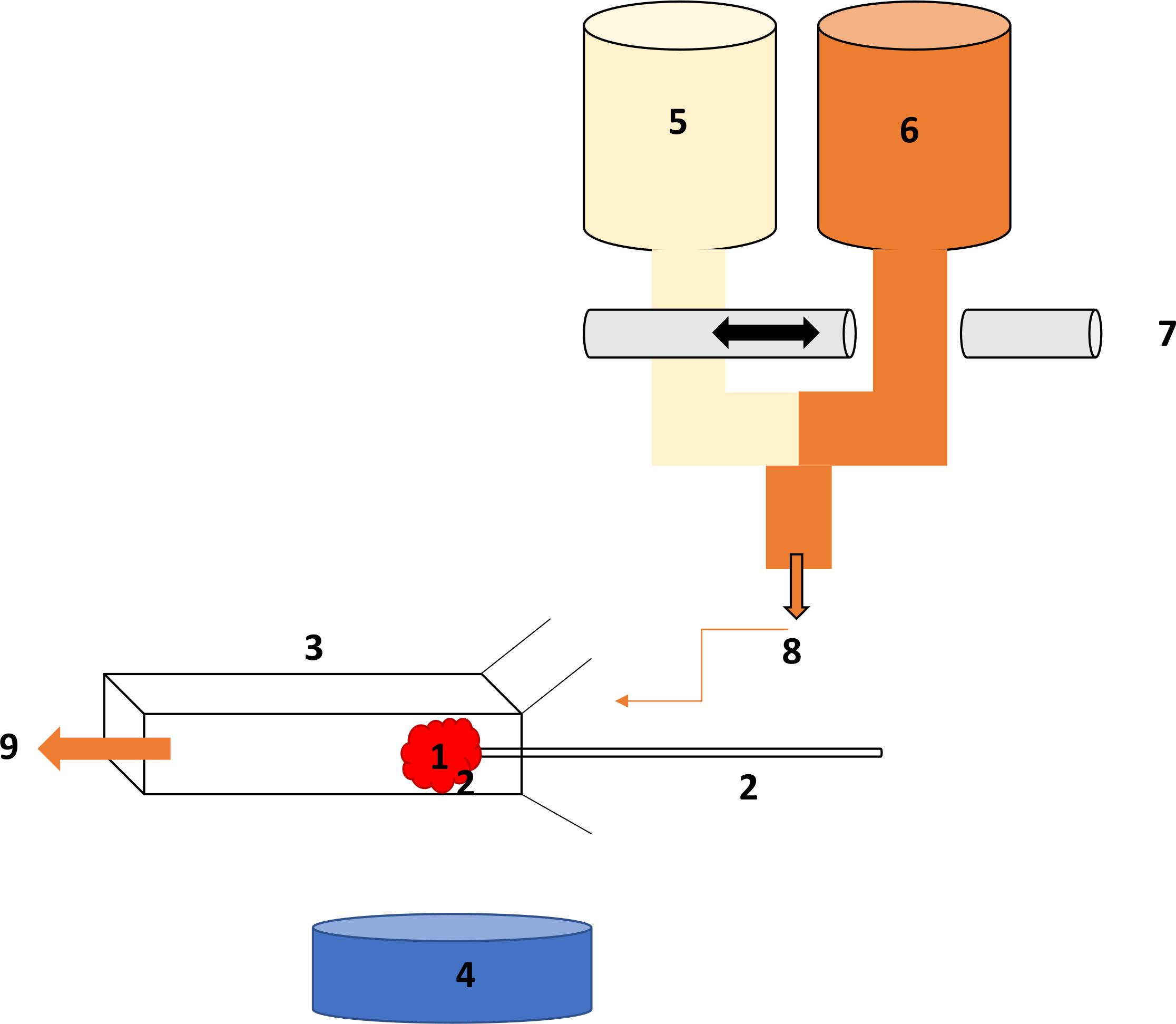
- Настройка клубочковых LpA / V,я установки, как описано в10лосося и др. Подробная схема настройки обратитесь к рис .
- Подготовить следующие решения: раствор Рингера-млекопитающих с 1% BSA (рН 7,4) и раствор Рингера-млекопитающих с 8% BSA (рН 7,4). Теплый и 37 ° C.
- Тянуть micropipettes из стеклянных капиллярных трубок (оптическая плотность: 1,2 мм). Генерировать кончик 5-8 мкм диафрагмы путем разрезания микропипеткой под микроскопом.
- Используйте клубочковых LpA / V,я сфальсифицировать поймать нетронутыми отдельных клубочков, которые свободны от Bowman's капсулу и трубчатых фрагменты на микропипеткой, используя всасывания. Подробное резюме oncometric assay находится в10лосося и др. Короче говоря после glomerulus поймали и закрепляться на всасывания микропипеткой, начните запись видео glomerulus под микроскопом.
- Во-первых, сбалансировать glomerulus в растворе Рингера BSA 1% для 30 s перед переключением perifusate концентрированный 8% BSA раствор Рингера для 10 s. Затем включите perifusate обратно в раствор 1% BSA Рингера и остановить запись.
- Смыть glomerulus и повторите этот процесс для 10-15 клубочков на мышь. Обеспечение скорости потока perifusate идентичны и не быстро (10 мл/мин), чтобы не исказить клубочковых структуры.
- Измерьте начальную скорость клубочковой усадки для вычисления проницаемости клубочковых воды (LpA) нормированный клубочковых тома (Vя). Подробная информация относительно анализа можно найти в10лосося и др.
6. периодические кислоты Шифф (ССА) пятно
Примечание: PAS пятно будет выделить подвале мембраны клубочковых капилляров петель и трубчатых эпителия. Это дает подробные визуализации клубочковых клеток, Мезангиальный матрицы и потенциал расширения и потенциальные изменения GBM (то есть, утолщение и нарушений).
- Раздел парафин врезанных, PFA-Исправлена почечной коры, используя микротом на 5 мкм толщина поли L-лизин покрытием слайды. Сухой при 37 ° C за 1 ч. обеспечить раздел не содержит любые складки или отверстий, которые могут исказить морфология под микроскопом.
- Deparaffinize слайды по инкубации дважды в ксилоле (осторожно, раздражающее; использование в кабинете дыма) 3 мин, дважды в 100% EtOH на 3 мин, а затем один раз в 95%, 70% и 50% EtOH 3 мин каждая, все при комнатной температуре. Повторно гидрата слайды в dH2O.
- Инкубируйте слайды в периодической кислоты раствор (осторожно, раздражающее; использование в вытяжной шкаф) (1 г/дл) для 5 минут, а затем промыть слайды в несколько изменений dH2O. используют контейнер с 100 мл dH2O при комнатной температуре.
Предупреждение: периодическая кислоты раствор: раздражающее; использование в вытяжной шкаф - Инкубируйте слайды в Шифф 's реагента (6 г/Л и натрия метабисульфит Parasoaniline HCl 4% HCl 0,25 моль/Л) для 15 мин при комнатной температуре. Вымойте слайды в проточной водой в течение 5 мин.
- Изображение с применением окрасок гематоксилин 3 s перед полосканием тщательно слайды в проточной водой в течение 15 мин.
Примечание: Некоторые оптимизации может потребоваться определить оптимальное время для окрашивания гематоксилином. - Обезвоживает слайдов, используя Обратный протокол депарафинизации в шаге 6.2. Закончить с ксилола.
- Воздух сухой слайды и горе с средствами массовой информации на основе ксилол монтажа.
- Изображение на световой микроскоп при 400-кратном оценить клубочковых структур. Оценить следующее: утолщение и нарушений GBM, рушится капиллярных петель, фиброзных тканей, склероз, пролиферации (эндотелия сосудов, Подоцит и Мезангиальный, или воспалительных клеток, проникнув клок).
Примечание: Для всеобъемлющей оценки клубочковых патофизиологии, поражений других в почках должно быть оцененным, таких, как трубочки.
7. просвечивающей электронной микроскопии (ТЕА)
Примечание: ТЕА позволяет изучение ультра-структурные аномалии в почках, например GBM, Подоцит ног процессы и эндотелиальных ярусе, которые не видны с световой микроскопии. Это важно в моделях, где почек ущерб произносится не так (то есть, не альбуминурия и основные структурные аномалии).
- Возьмите 2,5% gluteraldehdye - Исправлена кубиками почек и после исправления в 1% осмия тетраоксид за 1 ч. мыть в 50 мл 0,1 М cacodylate буфера (рН 7.3), а затем 50 мл dH2O (3 x 15 мин изменения).
Предупреждение: 2,5% глютаральдегид: токсичные, сенсибилизатор, раздражающее; Используйте вытяжной шкаф. 0,1 cacodylate натрия М: токсичные, использование в вытяжной шкаф - Обезвоживает с EtOH и внедрить в аралдита смолы.
- Вырезать секций на 50-100 Нм толщиной и пятно с 3% (водный) уранила ацетат и цитрата Рейнольдс свинца.
- Захватить несколько областей glomerulus в 940 X цифровой микроскопии, 1250 X и 6200 X чтобы podocytes, Генцс, GBM и mesangium можно определить.
- Для анализа ослепленный клубочков используйте ImageJ. Установите масштаб для каждого 6200 X Микрофотография путем рисования линии между двумя точками известных расстояния, таких как правитель. Перейти для анализа, а затем задайте шкала, где будет отображаться длину линии в пикселях. Введите известные расстояние и единиц измерения. Использование протоколов, перечисленных ниже для измерения каждого параметра.
Примечание: Этот анализ требует около 1 день мыши. Для надлежащей статистической мощности используйте средняя измерений от 3 клубочков от 3 мышей.- GBM вставить фиксированной цифровой сетки (10 x 10) за 6200 X Микрофотография и измерить толщину GBM в точке, где линии сетки пересекают GBM. Измерьте расстояние от базальной мембраны эндотелиальных клеток базальной Подоцит ног процесс клеточной мембраны в перпендикулярных касательной к эндотелиальной мембраны клетки, используя средство прямой линии. Определите средний измерения для каждого glomerulus из 10 отдельных измерений.
- Для числа эндотелиальной порозность измерить длину в 6200 X Микрофотография GBM и подсчитать количество эндотелиальных ярусе на единицу длины GBM. Длится в среднем от по крайней мере 4 микроскопии за glomerulus.
- Для Подоцит фут ширины процесса, вставить фиксированной цифровой сетки (10 x 10) над Микрофотография 6200 X. Измерьте ширину Подоцит ног процессов, которые пересекают линии сетки. Измерьте ширину в самой широкой части процесса ног, где он встречает GBM; Убедитесь, что линия перпендикулярна к касательной базальной мембраны Подоцит. Определите средний измерения для каждого glomerulus из 10 отдельных измерений.
- Для Подоцит щели шириной вставьте фиксированной цифровой сетки (10 x 10) над Микрофотография 6200 X. Измерения ширины диафрагмы Подоцит щели, которые пересекают линии сетки. Это та точка, где процессы ног ближе вместе, в самой широкой части каждого процесса ноги, от Подоцит мембраны для мембраны. Убедитесь, что измерение представляет собой перпендикуляр к касательной базальной мембраны Подоцит. Определите средний измерения для каждого glomerulus из 10 отдельных измерений.
- Количество Подоцит ног процессы, измерить длину в 6200 X Микрофотография GBM и подсчитать количество процессов Подоцит ног на единицу длины GBM. Длится в среднем от по крайней мере 4 микроскопии за glomerulus.
- Для охвата суб Подоцит пространства увидеть подробный метод в Нил и др. 14.
- С помощью 940 X микроскопии, изучите клубочков наличие ненормальной структуры, депозитов и инфильтраты, глаз.
8. иммунофлюоресценции для Подоцит и эндотелиальных маркеры
Примечание: Иммуноокрашивания позволяет визуализацию шаблонов выражение, белка, например эндотелия капилляров петли, которые можно свернуть в клубочковых болезни.
- Место OCT-формы, содержащие замороженных почек при-20 ° C за 2 ч до разрезания. Убедитесь, что поверхности среза почки тщательно помещены против нижней части OCT-прессформы для включения разделов хорошо ориентированным ткани.
- С помощью криостат, раздел ткани на 5 мкм толщина на поли L-лизин покрытием слайды.
Примечание: Убедитесь, что есть никаких складок или отверстия в разделе ткани, который может исказить морфологии. - После удаления из криостата, исправить слайды в 4% PFA для 10 min. мыть слайды 3 x 5 мин в контейнере с 100 мл dH2O.
Предупреждение: 4% ПФА: фиксатор, использование в вытяжной шкаф - Чтобы свести к минимуму количество антител используемых, нарисуйте вокруг секции с гидрофобным пера. Не позволяйте разделы сухой.
- Инкубируйте в блокировании решение (BSA 3% и 5% нормальной сыворотки в ПБС) за 1 ч при комнатной температуре.
- Удалять блокирующие решение с аспиратором и инкубировать секции с основного антитела (Nephrin, podocin или PECAM-1) разводят 1: 250 (3% BSA в однократном ПБС). Место слайды в увлажненные камере при температуре 4 ° C на ночь. Если не доступен влажной камере, слегка место небольшую полоску парафина на слайд, антителами. Будьте осторожны при удалении на следующий день, чтобы не нарушить секции.
- Вымойте слайды 3 x 5 мин в однократном ПБС.
- Проинкубируйте с соответствующим флуоресцентные вторичное антитело разбавления 1: 1000 (3% BSA в однократном ПБС) за 2 ч при комнатной температуре в темноте.
- Вымойте слайды 3 x 5 мин в однократном ПБС. Крепление с флуоресцентных монтаж средств массовой информации, содержащие DAPI.
- Изображения слайдов с флуоресцентный Микроскоп при увеличении 400 X для просмотра клубочков. Убедитесь, что это ослепленный избежать предвзятости.
- ImageJ использование для анализа интенсивность окрашивания, нормированный клубочковых области, и шаблон окрашивание, то есть, количество капиллярных петель нормированный клубочковых район, ослепленный образом.
9. белка добычу и западный Blotting
Примечание: Западный blotting позволяет нам оценить выражение белки, известно dysregulated в почечной болезни. Например снижение podocin и nephrin выражение указывает на потерю Подоцит.
- Экстракт белка от почечной коре и, котор фильтруют клубочков; Протокол является то же самое для каждого и объем буфера lysis корректируется на сумму ткани.
- Оттепель почек/клубочков на льду перед добавлением NP-40 лизис буфера содержащие ингибиторы протеазы и фосфатазы (150 мм NaCl, 1% NP-40, 50 мм трис pH 8). Однородный образца для 30 s.
- Инкубируйте гомогенизированных образцов на льду в течение 30 мин, vortexing на регулярной основе.
- Центрифугуйте образцы на 10000 x g 15 мин при 4 ° C.
- Удалите супернатант в свежий трубку на льду.
Примечание: ожидаете восстановления приблизительно 1 мг белка на сэмпл. - Денатурируйте белки, с использованием стандартной 4 буфера x Laemmli. Кипятите смесь на 95-100 ° C за 5 мин.
- Оценить выражение клубочковых клеток маркер белков (Nephrin, Podocin, PECAM-1, и т.д.) и фосфорилирования и экспрессию белков, известный/предположить, чтобы быть изменены в почки/клубочков модели болезни с помощью западных blotting) Стандартный метод; Махмуд и Ян15).
Примечание: Протокол будет варьироваться в зависимости от размера и обилием протеина интереса.
10. РНК добыча и полимеразной цепной реакции (ПЦР)
Примечание: анализ выражения мРНК позволяет нам определить, как гены, регулируются в почечной болезни, такие как изменения в экспрессии генов и альтернативного сплайсинга.
- Хотя коре почки по-прежнему заморожены, тщательно растереть в 3 мл реагента фенола с помощью пестика и минометов. При использовании клубочковых экстракты, добавьте 1 mL реагента фенола и гомогенизации образца для 30 s.
Предупреждение: Тризол реагента: раздражающее; использование в вытяжной шкаф - Выполните извлечение RNA, с помощью метода, описанного Chomczynski и Сакки16.
Примечание: Коммерческие РНК добыча комплекты доступны в качестве альтернативы для данного метода. - Оценить количество и качество РНК, полученные с помощью одного из различных доступных методов. РНК является aliquoted и хранятся при температуре-80 ° C на данный момент. Избежать повторения заморозить оттаивания.
Примечание: Если новое в этот метод, проверьте качество РНК перед переходом к следующему шагу, запустив РНК на геле агарозы для обеспечения четкой 28S и 18S рибосомных band. Ожидаете, чтобы восстановить между 2-5 мкг РНК с помощью этого метода. - DNase лечения 1 мкг РНК (сделать объем до 10 мкл с РНКазы свободной воды плюс 1 мкл DNase и 1 мкл буфера DNase) за 1 ч при 37 ° C. Остановите реакции с 1 мкл DNase стоп раствора при 65 ° C для 10 мин.
- 0.5 мкл oligo (dT) и случайных праймеров. Инкубируйте на 70 ° C на 10 мин.
- Немедленно утолите на льду на 5 мин.
- Добавьте следующее; MMLV фермента обратной транскриптазы (400 U; заменить DEPC H2O в RT - образец элемента управления), буфер (1 x), MMLV dNTP смеси (0,5 мм), а также рибонуклеаза ингибитора (40 ед); Сделайте до 50 мкл DEPC водой.
- Инкубируйте реакция смеси при 37 ° C за 1 ч, следуют 95 ° C за 5 мин для деактивации фермента.
Примечание: Для создания более высокую доходность cDNA, инкубации при 37 ° C до 3 ч. - Оценить количество и качество cDNA, используя различные методы.
- Использование ПЦР для оценки моделей выражение и сплайсинга мРНК генов предположить, чтобы быть dysregulated в модели клубочковых болезни. Протокол будет варьироваться в зависимости от гена интереса.
Результаты
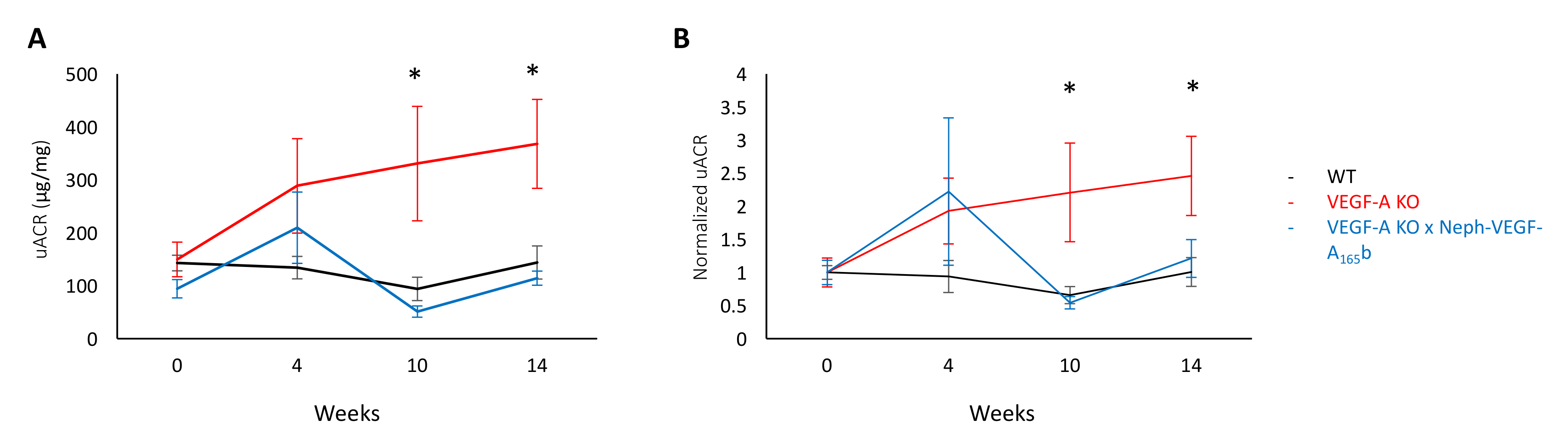
Моча была собрана с помощью метаболизм клеток от дикого типа (WT), индуцибельной Подоцит конкретных VEGF-A выбить (ко-VEGF-A) и VEGF-A KO X неф-VEGF165б мышей (VEGF-A KO мышей, которые чрезмерно экспресс изоформы b человека VEGF-А165в podocytes в Учредительный образом). После измерения креатинина соотношение альбумин мочевыделительной на 0, 4, 10 и 14 недель после индукции доксициклин VEGF-A KO VEGF-A KO мышей разработан 10 недель, по сравнению с WT помёте элементы прогрессивного альбуминурии. Абсолютные значения можно увидеть на рисунке 2A, и нормализованных к базовому значения каждой мыши на рисунке 2B. Однако альбуминурию в не наблюдается в VEGF-A X неф-VEGF-A165б мышей (рис. 2), указывающее, что VEGF-А165b защитные в модели альбуминурию5.
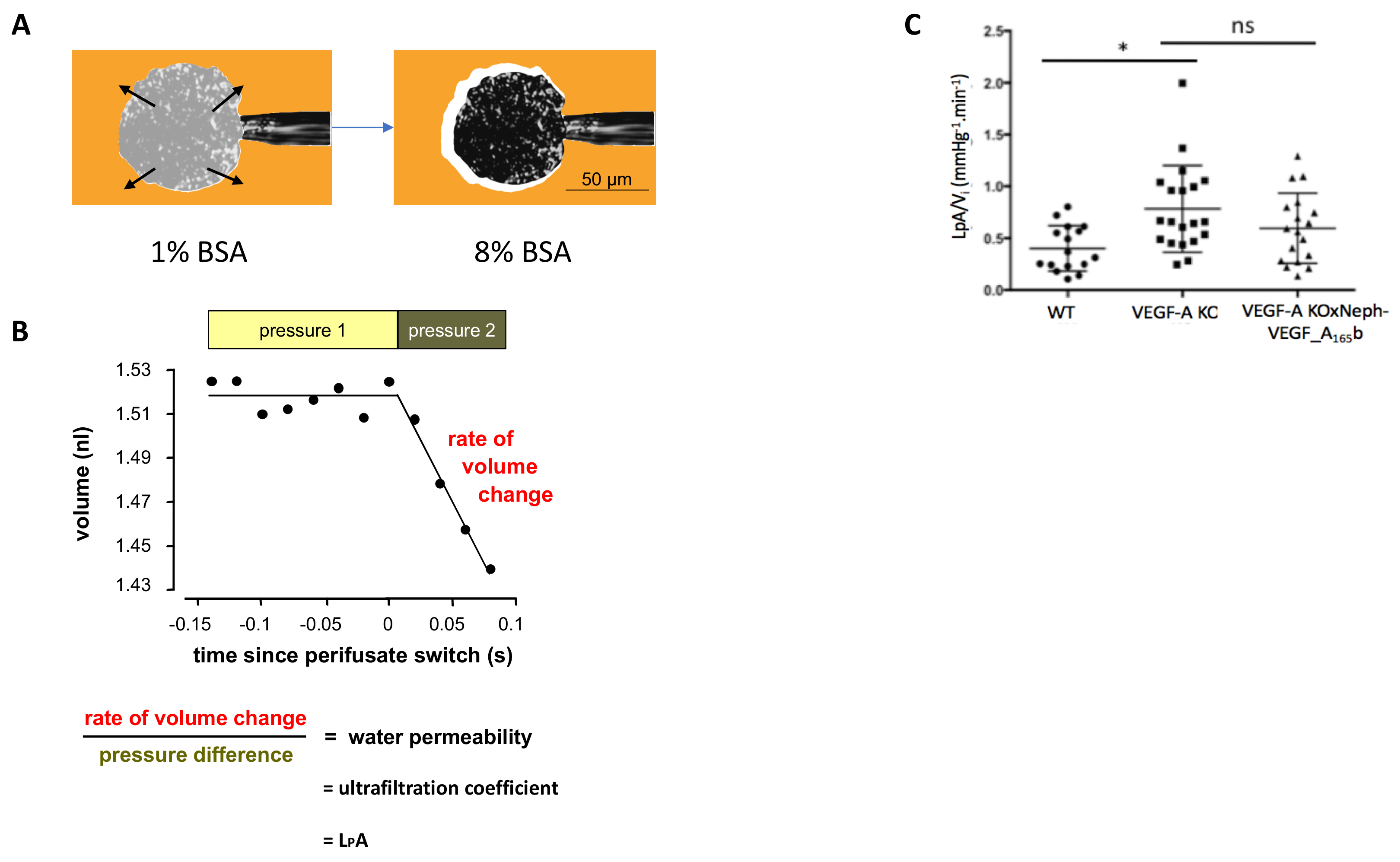
Клубочковых LpAVя была измерена в отдельных клубочков, котор фильтруют от WT, ко-VEGF-A и VEGF-A X неф-VEGF-A165б почки. Пример как поймали glomerulus и усадки наблюдается, когда perifused с 8% BSA показано на Рисунок 3A. Затем этот усадки используется для определения клубочковых LpA / Vя для каждого glomerulus (рис. 3B). VEGF-A KO мышей были значительно увеличили клубочковых LpA / Vя на 14 недель пост VEGF-KO индукции, по сравнению с WT управления клубочков. Хотя ниже в VEGF-A X неф-VEGF-A165б мышей, увеличение клубочковых LpA / V,я не была предотвращена гиперэкспрессия VEGF-А165б на 14 недель5.
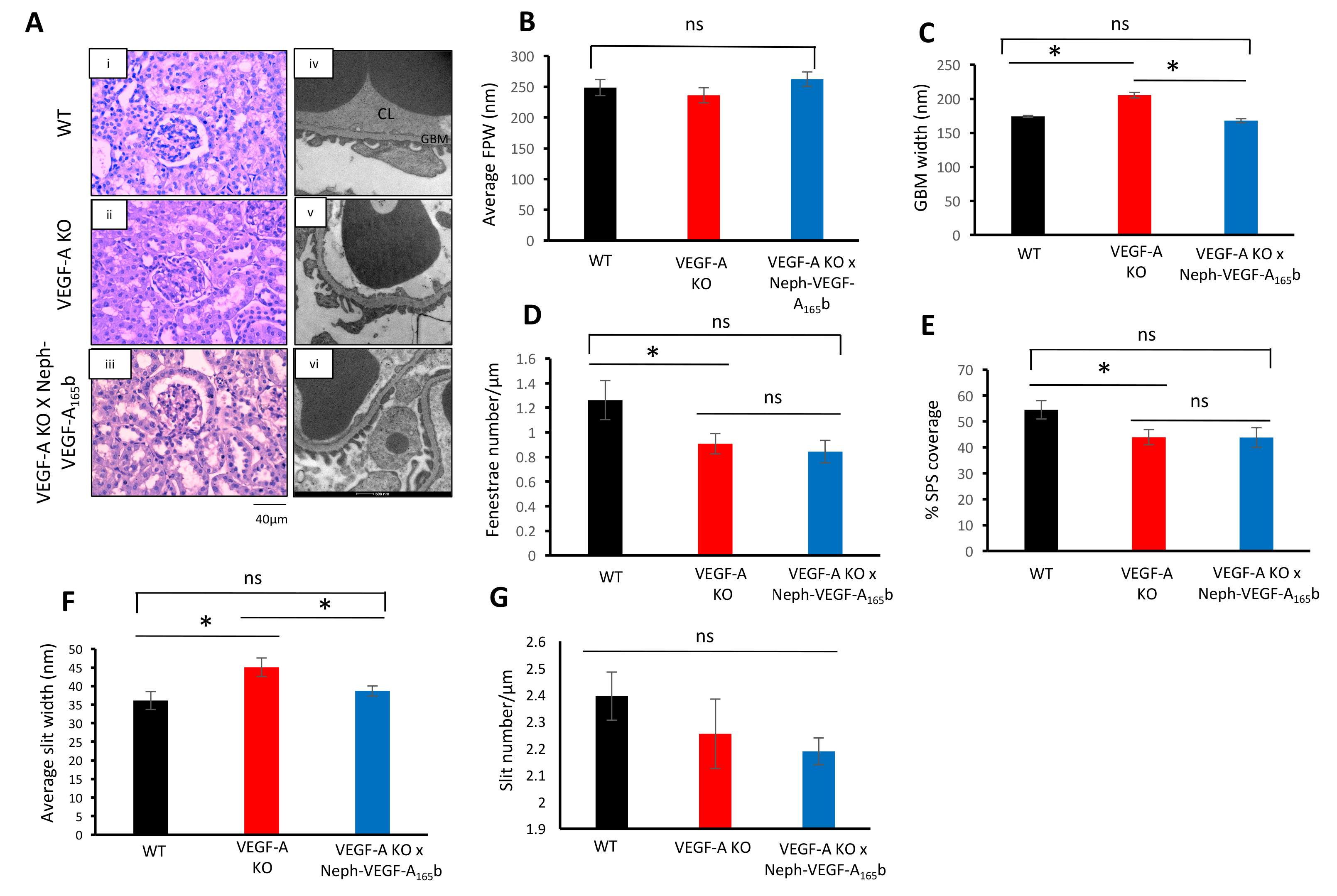
PAS окрашивание участков коры почки 14 недель после индукции VEGF-A KO не выявили какие клубочковых структурные аномалии в ко-VEGF-A или VEGF-A X неф-VEGF-A165б мышей (рис. 4A). Однако, после анализа структуры клубочковых ультра-через EM, VEGF-A KO мышей разработан увеличение ширины GBM, уменьшилось количество эндотелиальных окна, снизилась покрытие SPS и увеличилась средняя Подоцит щели шириной (рис. 4 c-4F ). Средняя Подоцит ног процесс ширину и количество щелей, осталась неизменной (рисунок 3B и 3 G). Гиперэкспрессия VEGF-А165б в VEGF-A KO мышей предотвратить изменения в GBM и щели шириной (Рисунок 4 c и 4F). Однако VEGF-А165б было не влияет на число измененных окна и SPS покрытия (Рисунок 4 d и 4E)5.
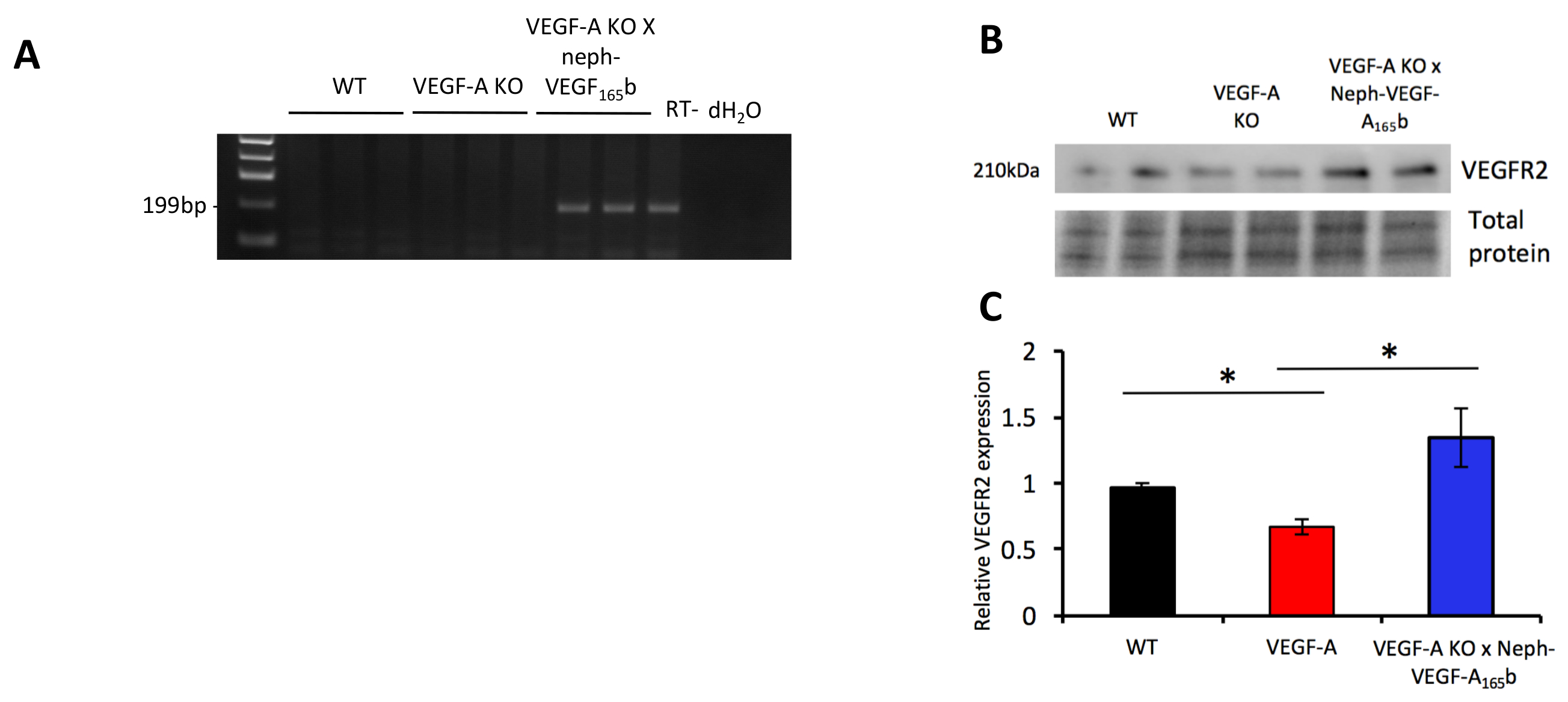
-ПЦР на РНК, извлеченные из фильтруют клубочков показали, что человека b165VEGF-A мРНК присутствует только в VEGF-A KO X неф-VEGF-A165б мышей (Рисунок 5A). При извлечении белка от фильтруют клубочков и оценки уровня белков через западный blotting, выражение клубочковых белка БЕСПРОГРЕССИВНУЮ-2 было установлено, уменьшилось в VEGF-A KO мышей, которая помешала гиперэкспрессия VEGF-А165б ( Рисунок 5B и 5 C)5.

Рисунок 1. Схематичных Настройка установки проницаемости клубочковых (LpA). (A) glomerulus ловится на (B) микропипеткой в держатель с помощью всасывания, который крепится на гору для стабильности. (C) прямоугольной микрослайдовому. (D) 4 X цель микроскопа с видео камерой. (E) BSA 1% раствор нагревают до 37 ° C. (F) решения 8% BSA прогреты до 37 ° C. (G) пульта нажмите подшипник две perifusate содержащих линии, которая позволяет быстрое perifusate обмен. (H) маршрут perifusate к микрослайдовому, который затем ванны glomerulus. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}

Рисунок 2. Соотношение альбумин мочевыделительной креатинина. (A) uACR значения на недели 0, 4, 10 и 14 после индукции VEGF-A KO в WT, ко-VEGF-A и VEGF-A X неф-VEGF-A165б мышей. (B) одинаковые значения uACR нормализовано к значению базового (неделя 0) каждого индивидуального мыши. UACR значительно увеличилась в VEGF-A KO мышей в недели, 10 и 14, по сравнению с WT помёте контроля, который был лишен в мышах b165VEGF-A X неф-VEGF-A (* p < 0,05; Двусторонний ANOVA, коррекции для сравнения между парами; n = 4-12 мышей на момент времени; планки погрешностей: среднеквадратичная ошибка среднего значения [МДж]). Эта цифра была изменена Stevens et al5. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}

Рисунок 3. Измерение проницаемости клубочковых воды. (A) glomerulus поймали на микропипеткой через всасывания; perifusate перешли от 1% BSA (Ai) до 8% BSA (Оки), и наблюдается клубочковых усадки. (B) измерений до и после 8% BSA переключения используются для определения клубочковых LpA / V,я. (C) VEGF-A KO мышей развивать увеличение клубочковых LpA / Vя на 14 недель пост индукции VEGF-A KO, по сравнению с WT элементов управления. Это не помешало значительно VEGF-A X неф-VEGF-A165б мышей (* p < 0,05; Один из способов ANOVA, Бонферрони коррекции для сравнения между парами; n = 4-9 мышей, 15-30 клубочков; планки погрешностей: SEM). Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}

Рисунок 4. Клубочковых структурный анализ. (A) PAS пятнать почечной коры не указывается каких-либо структурных отклонений в клубочков от WT, ко-VEGF-A и VEGF-A X неф-VEGF-A165б мышей (АИ - iii). ЕТ показали ультра-структурные аномалии в VEGF-A KO клубочков (АИВ - vi). (B) средняя РПЖ не изменился между группами. (C) GBM увеличилась в VEGF-A KO клубочков, которая помешала VEGF-А165б. (D) количество окна была снижена в VEGF-A KO клубочков, которые остались неизменными, VEGF-А165б. покрытие SPS (E) был сократилось в VEGF-A KO клубочков, которые также остаются неизменными VEGF-А165б. (F) в VEGF-A KO клубочков увеличилась средняя ширина щели, которая помешала VEGF-А165б. (G) щели номер был неизменным между тремя группами (* p < 0,05; Один из способов ANOVA, Бонферрони коррекции для сравнения между парами; n = 3 мышей, 9 клубочков; планки погрешностей: SEM). Эта цифра была изменена Stevens et al5. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}

Рисунок 5. выражение mRNA и белка маркеров. (A) RT-PCR показал, что человеческое выражение mRNA b165VEGF-А только проявляется в, котор фильтруют клубочков от VEGF-A KO X неф-VEGF-A165б мышей. (B) Западный blotting указал, что экспрессия белка БЕСПРОГРЕССИВНУЮ-2 регулируемых вниз в VEGF-A KO клубочков, который был лишен в VEGF-A KO X неф-VEGF-A165б клубочков (* p < 0,05; Один из способов ANOVA, Бонферрони коррекции для сравнения между парами; n = 3 - 6 мышей; планки погрешностей: SEM). Эта цифра была изменена Stevens et al5. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}
Обсуждение
Этот протокол описывает полный почек работы вверх, которая должна осуществляться в моделях мыши клубочковых заболеваний, что позволяет огромное количество информации, касающейся почек и клубочковых функции будут получены из одной мыши. Важнейшие шаги в каждом методе позволяют для детального анализа функциональной, структурной и механистические клубочковых функции, включая оценку проницаемости почек в целом (uACR и плазмы креатинина измерения), проницаемость Индивидуальные клубочков (клубочковых LpA / V,я), изучение структурных изменений (пас, Trichrome синий и Эм), белок локализации (КРП) и экспрессии генов клубочковых (RT-PCR и Западный blotting). Эти методы являются ключом к полной оценки клубочковых функции в модели мыши почечной болезни.
При оценке проницаемость GFB, многие исследования решили использовать обычных uACR или 24 h скорость экскреции альбумина в качестве эффективной меры17,18. Хотя эти методы позволяют оценку GFB проницаемости в целом, это не позволяет для оценки отдельных проницаемости клубочковых и вариации среди клубочков. Предыдущие исследования показали измерения клубочковых LpA / Vi более чувствительным мерой изменения GFB проницаемость5,9. Действительно в результатах представитель продемонстрировал в этой статье, в 14 недель поста индукции VEGF-A ко, VEGF-A KO X неф-VEGF-A165б мыши имеют значительно ниже uACR, по сравнению с VEGF-A KO мышей; Однако, этот результат не отражается в клубочковых LpA / Vя измерений, где VEGF-А165б не помешало значительно увеличивается в GFB проницаемость (рис. 1 и рис. 2)5. Это свидетельствует о важности использования нескольких анализов для оценки проницаемости почечных и проницаемость отдельных клубочков. Кроме того клубочковых LpA / Vя oncometric анализа предполагает, что проницаемость отдельных клубочков от же почек может сильно различаться, особенно в болезни модели5,10, 19. одним из ограничений для измерения клубочковых LpA / V,я это, что она может быть выполнена только на экспериментальной конечной точки; Таким образом дать указание экспериментальной конечной требуются регулярные uACR измерений.
Помимо оценки функциональной фенотип, нынешний метод также поощряет оценки структурных и ультраструктурная фенотипа. Это может быть сделано с помощью выбор пятна как PAS, trichrome и серебряные пятна; Каждый, для оценки различных аспектов клубочковых морфологии. В острый модели клубочковых болезни, которая часто бывает в моделях мыши, это может быть трудно обнаружить любые крупные структурные аномалии, с помощью этих пятен, если вы патологоанатом подготовленных почек. Таким образом проведение EM предлагается оценить Ультраструктура GFB, который позволяет количественного измерения параметров, таких как GBM, эндотелиальные окна размер и число и Подоцит характеристики. Такие размеры требуют минимальной подготовки для выполнения и позволяет следователю для определения ячеек типы/структуры пострадавших в модели болезни. В примере, показанном в результатах представительных, был найден VEGF-A KO мышь мягкая модель клубочковых болезни таким образом, без крупных структурные аномалии были представлены после окрашивания PAS. Однако ко-VEGF-A Подоцит конкретных вызвать изменения GBM, podocytes и эндотелиальных клеток при рассмотрении клубочковых ультра структура5. К сожалению подготовка почки для EM, описанные в рамках нынешнего метода не включить обнаружение эндотелиальной Гликокаликс, который известен также оказывать существенное воздействие на проницаемость GFB19. Для того, чтобы точно измерить глубину Гликокаликс, функции почек следует perfuse исправлена с глютаральдегид 2,5% с 1% Альциановый синий эндотелиальной Гликокаликс маркировки, как описано в Oltean et al19.
После того, как были оценены функциональные и структурные фенотип, выражение/активации модели различных генов и пути можно затем оценить специально клубочков. До ультра-структурной оценки могли бы дать некоторую информацию о камере типы/клубочковых структур, указывающее, следует ли Подоцит или эндотелиальной конкретных генов/пути. Например в результатах представитель от VEGF-A KO мышей, наблюдалось снижение числа эндотелиальной окна (рис. 3D); Таким образом было рассмотрено выражение клубочковых белка маркера эндотелиальной известно, участвующих в пути VEGF-A; БЕСПРОГРЕССИВНУЮ-2 (рис. 4B)5. Помимо выражения белков в клубочков их локализации можно изобразить с помощью если. В исследовании Zhang et al20Подоцит конкретных гиперэкспрессия GLUT1 было подтверждено в podocytes, если совместно локализации увеличение GLUT1 с podocin.
По сравнению с альтернативными методами, представлены в литературе для оценки клубочковых функции использование метода, описанного в настоящем документе для оценки функции почек в моделях мыши клубочковых заболевания позволяет клубочковых фенотип полностью оцениваться от несколько аспектов. С помощью этого метода, исследователь может определить почек фенотип модели и оценить механизм о том, почему фенотипа развивается. Это жизненно важную информацию о механизме заболевания требуется при рассмотрении потенциальных терапевтических направлений в этих моделях. Этот метод может легко применяться для будущих расследований клубочковых функции в оценке заболевания фенотипы и потенциальные терапии.
В заключение этот универсальный и адаптируемая протокол описывает полный почек работы вверх для моделей мыши клубочковых заболеваний, что позволяет огромное количество информации, касающейся почек и клубочковых функции будут получены из одной мыши. Методы позволяют для детального анализа функциональной, структурной и механистические клубочковых функции, которые могут быть применены ко всем моделям мыши клубочковых болезни.
Раскрытие информации
Авторы не имеют ничего сообщать.
Благодарности
Эта работа была поддержана Британского фонда сердца, Ричард Брайт VEGF исследований доверяют и MRC.
Материалы
| Name | Company | Catalog Number | Comments |
| Metabolic Cages | Harvard Apparatus | 52-6731 | |
| Tris buffered saline (10x) | Sigma-Aldrich | T5912-1L | |
| Bovine Serum Albumin | Sigma-Aldrich | A2058 | |
| Mouse Albumin ELISA Quantitation Set | Bethyl Laboratories | E90-134 | |
| TMB ELISA Substrate solution | ThermoFisher Scientific | 34028 | |
| Sulphuric acid | Sigma-Aldrich | 339741 | |
| SPECTRstar nano | BMG Labtech | SPECTRstar nano or equivalent | |
| RNAlater stabilisation solution | ThermoFisher Scientific | AM7020 | |
| 4-15% precast protein gel | BIORAD | 4568084 | |
| 4x Laaemmli Sample buffer | BIORAD | 161-0747 | |
| Mini Trans-Blot cell | BIORAD | 1703930 | |
| 10x Tris running buffer | BIORAD | 1610732 | |
| Coomassie Brilliant Blue Dye | ThermoFisher Scientific | 20278 | |
| Creatinine Companion | Exocell | 1012 Strip Plate | |
| Glutaraldehyde Solution | Sigma-Aldrich | G5882 | |
| Sodium Cacodylate | Sigma-Aldrich | C0250 | |
| 10x Phosphate Buffered Saline | ThermoFisher Scientific | AM9625 | |
| Sodium Chloride | Sigma-Aldrich | S7653 | |
| Sodium Acetate | Sigma-Aldrich | S2889 | |
| Sodium Phosphate | Sigma-Aldrich | 342483 | |
| Sodium Bicarbonate | Sigma-Aldrich | S5761 | |
| Magnesium Sulfate | Sigma-Aldrich | M2643 | |
| Calcium Chloride | Sigma-Aldrich | C5670 | |
| D(+)Glucose | Sigma-Aldrich | G8270 | |
| EDTA Blood Collection tubes | BD | 367835 | |
| 23-25G Needle | BD | PMC0735 | |
| EDTA | Sigma-Aldrich | E9884 | |
| 10 ml Glass Vial | Thomas Scientific | 0914X10 | |
| Falcon 10 ml Polypropylene Tubes | ThermoFisher Scientific | 10110101 | |
| 0.5 ml Tubes | ThermoFisher Scientific | 10681894 | |
| Disposable Tissue Molds | ThermoFisher Scientific | 22-363-553 | |
| Mouse Surgical Disection Kit | ThermoFisher Scientific | 13-005-204 | |
| Optimal Cutting Medium | ThermoFisher Scientific | 23-730-571 | |
| 4% Paraformaldehyde | ThermoFisher Scientific | AAJ19943K2 | |
| Glass Capillary Tubes | Harvard Apparatus | EC1 64-0770 | |
| Glomerular Permeability Rig | Built at the Univeristy of Bristol - not comercially available | Citation of LpA rig: Salmon et al. 2006; J. Physiol | |
| Stainless Steel Sieves | Cole-Parmer | WZ-59984 | |
| Periodic Acid-Schiff (PAS) Staining System | Sigma-Aldrich | 395B-1KT | |
| Hematoxylin | Sigma-Aldrich | H3136 | |
| Xylene | MerckMillipore | 108298 | |
| Poly-Prep Slides | Sigma-Aldrich | P0425-72EA | |
| Mounting Medium | ThermoFisher Scientific | 8030 | |
| Osmium tetroxide solution | Sigma-Aldrich | 75632 | |
| Aradite Resin | Agar Scientific | CY212 | |
| Uranyl Acetate | Agar Scientific | AGR1260A | |
| Lead Citrate | Agar Scientific | AGR1210 | |
| Cryostat | ThermoFisher Scientific | e.g. 957000H | |
| Hydrophobic Pen | Abcam | ab2601 | |
| Nephrin (1243-1256) Antibody | Acris | BP5030 | |
| Anti-Podocin | Sigma-Aldrich | P0372-200UL | |
| Anti-CD31 | BD Biosciences | 550274 | |
| Alexa Fluor Secondary Antibody | ThermoFisher Scientific | A32732 | |
| Vectashield Mounting Medium with DAPI | Vector Labs | H-1200 | |
| NP40 Cell Lysis Buffer | ThermoFisher Scientific | FNN0021 | |
| Halt Protease and Phosphatase Inhibitor Cocktail | ThermoFisher Scientific | 78437X4 | |
| 10x Transfer Buffer | BIORAD | 1610734 | |
| PVDF Membrane | ThermoFisher Scientific | LC2002 | |
| HRP-Conjugated Secondary Antibodies | Abcam | ab6721 | |
| ECF Substrate for Western Blotting | Fisher | 10713387 | |
| TRIzol | ThermoFisher Scientific | 15596018 | |
| Dnase I | New England Biolabs | M0303S | |
| M-MLV Reverse Transcriptase | New England Biolabs | M053S | |
| Oligo dT | ThermoFisher Scientific | 18418012 | |
| Random Primers | ThermoFisher Scientific | 48190011 | |
| dNTP | ThermoFisher Scientific | 18427088 | |
| Ribonuclease Inhibitor | ThermoFisher Scientific | 10777019 | |
| DEPC Water | ThermoFisher Scientific | AM9915G | |
| Fluorescent Light Miscroscope | Leica Microsystems | ||
| Image J Analysis Software | Image J | ||
| PCR Thermocycler | ThermoFisher Scientific | ||
| TEM Microscope | Britannica |
Ссылки
- Humtstrom, M. Development of structural kidney damage in spontaneously hypertensive rats. Journal of Hypertension. 30 (6), 1087-1091 (2012).
- Kitada, M., Ogura, Y., Koya, D. Rodent models of diabetic nephropathy: their utility and limitations. International Journal of Nephrology and Renovascular Disease. 14 (9), 279-290 (2016).
- Tesch, G. H., Lim, A. K. Recent insights into diabetic renal injury form the db/db mouse model of type 2 diabetic nephropathy. American Journal of Physiology-Renal Physiology. 300 (2), F301-F310 (2011).
- Fogo, A. B. Animal models of FSGS: lessons for pathogenesis and treatment. Seminars in Nephrology. 23 (2), 161-171 (2003).
- Stevens, M., et al. VEGF-A165b protects against proteinuria in a mouse model with progressive depletion of all endogenous VEGF-A splice isoforms from the kidney. Journal of Physiology. 595 (19), 6281-6298 (2017).
- Cosgrove, D., Kalluri, R., Miner, J. H., Segal, Y., Borza, D. B. Choosing a mouse model to study the molecular pathobiology of Alport glomerulonephritis. Kindey International. 71 (7), 615-618 (2007).
- Kujal, P., Vernerova, Z. 5/6 nephrectomy as an experimental model of chronic renal failure and adaptation to reduced nephron number. Ceskoslenska Fysiologuie. 57 (4), 104-109 (2008).
- Chevalier, R. L., Forbes, M. S., Thornhill, B. A. Ureteral obstruction as a model of renal interstitial fibrosis and obstructive nephropathy. Kidney International. 75 (11), 1145-1152 (2009).
- Oltean, S., et al. VEGF165b overexpression restores normal glomerular water permeability in VEGF164-overexpressing adult mice. American Journal of Physiology-Renal Physiology. 303 (7), F1026-F1036 (2012).
- Salmon, A. H., Neal, C. R., Bates, D. O., Harper, S. J. Vascular endothelial growth factor increases the ultrafiltration coefficient in isolated intact Wistar rat glomeruli. Journal of Physiology. 570 (Pt 1), 141-156 (2006).
- Zheng, F., Striker, G. E., Esposito, C., Lupia, E., Striker, L. J. Strain differences rather than hyperglycemia determine the severity of glomerulosclerosis in mice. Kidney International. 54 (6), 1999-2007 (1998).
- Betz, B., Conway, B. R. An update on the use of animal models in diabetic nephropathy research. Current Diabetes Reports. 16 (18), (2016).
- Savin, V. J., Terreros, D. A. Filtration in single isolated mammalian glomeruli. Kidney International. 20 (2), 188-197 (1981).
- Neal, C. R., Crook, H., Bell, E., Harper, S. J., Bates, D. O. Three-dimensional reconstruction of glomeruli by electron microscopy reveals a distinct restrictive urinary subpodocyte space. Journal of the American Society of Nephrology. 16 (5), 1223-1235 (2005).
- Mahmood, T., Yang, P. C. Western blot: technique, theory, and trouble shooting. North American Journal of Medicine & Science. 4 (9), 429-434 (2012).
- Chomczynski, P., Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Analytical Biochemistry. 162 (1), 156-159 (1987).
- Adembri, C., et al. Sepsis induces albuminuria and alterations in the glomerular filtration barrier: a morphofunctional study in the rat. Critical Care. 15 (6), R277 (2011).
- Ma, S. T., Liu, D. L., Deng, J. J., Niu, R., Liu, R. B. Effect of arctiin on glomerular filtration barrier damage in STZ-induced diabetic nephropathy rats. Phytotherapy Research. 27 (10), 1474-1480 (2013).
- Oltean, S., et al. Vascular endothelial growth factor-A165b is protective and restores endothelial glycocalyx in diabetic nephropathy. Journal of the American Society of Nephrology. 26 (8), 1889-1904 (2015).
- Zhang, H., et al. Podocyte-specific overexpression of GLUT1 surprisingly reduces mesangial matric expansion in diabetic nephropathy in mice. American Journal of Physiology-Renal Physiology. 299 (1), F91-F98 (2010).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены