
Method Article
細菌人工染色体cDNAクローンからの組換えジカウイルスの救出
要約
ジカウイルスの最近の流行は、ワクチンや治療戦略を開発するための逆遺伝的アプローチを確立することの重要性を強調しています。ここでは、ヒトサイトメガロウイルスの制御下で細菌人工染色体に組み立てられた全長cDNAクローンから感染性組換キンジカウイルスを早期に促進するプロトコルについて説明する。
要約
ジカウイルス(ZIKV)感染と神経性合併症との関連、および承認されたワクチンおよび/または抗ウイルス薬の欠如は、ZIKV生物学の研究と治療および/または予防的アプローチの開発。しかし、他のフラビウイルスと同様に、ZIKV全長感染性cDNAクローンの生成は、細菌の増幅中にウイルス配列の毒性のために妨げられてきた。この問題を克服するために、細菌人工染色体(BAC)の使用に基づく非伝統的なアプローチを開発しました。このアプローチを使用して、ZIKV株リオグランデドノルテナタール(ZIKV-RGN)の全長cDNAコピーは、4つの合成DNA断片から生成され、ヒトサイトメガロウイルス(CMV)の制御下で単一コピーpBeloBAC11プラスミドに組み立てられます。即時早期プロモーター。組み立てられたBAC cDNAクローンは細菌の伝播中に安定であり、感染性組換え(r)ZIKVはBAC cDNAクローンのトランスフェクション後にベロ細胞で回収される。ここで説明するプロトコルは、ZIKVおよび他の陽性鎖RNAウイルス、特に細菌の間に安定性の問題を有する大きなゲノムを持つものを含むフラビウイルスの感染性クローンの生成のための強力な技術を提供する伝播.
概要
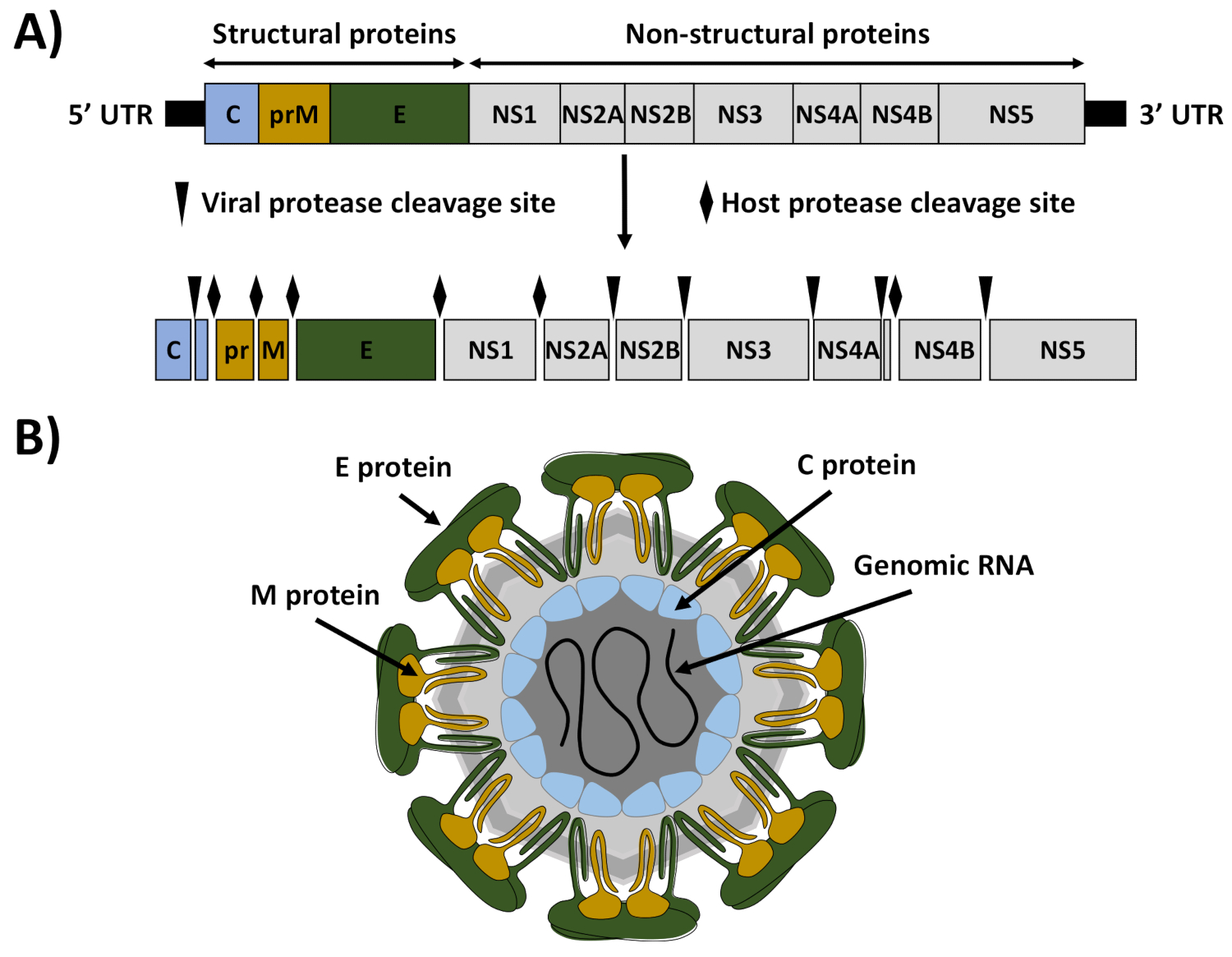
ZIKVは、現在世界的な公衆衛生上の緊急事態1を構成するフラビリダ科内のフラビウイルス属の蚊媒介メンバーである。他のフラビウイルスと同様に、ZIKVは、約10.8kb(図1)2の正の感覚、一本鎖RNA分子を含むイコサヘドラ状構造を有する封入されたRNAウイルスである。ウイルスゲノムは、ウイルスおよび細胞プロテアーゼによって処理される約3,423アミノ酸の大きな多タンパク質を、3つの構造タンパク質(カプシド[C]、膜/膜前膜[prM/M]、およびエンベロープ[E])の形成に関与する3つの構造タンパク質にコードする。ゲノム複製、ウイルス組み立て、宿主免疫応答の回避に関与する7つの非構造(NS2A、NS2B、NS3、NS4A、NS4B、NS5)のウイルス粒子および7個の非構造(NS)タンパク質(図1)3。
歴史的に、ZIKV感染は軽度の熱性疾患4、5と関連している。しかし、南米、中米、南太平洋、カリブ海諸国におけるZIKV感染の爆発的な最近のパンデミック、およびギラン・バレ症候群の発生との関連小頭症9、10、11、12、13は、歴史的認識を変更し、重要なヒト病原体としてのZIKVの関連性を増強した。この意味で、感染性cDNAクローンなどの分子ツールの開発は、ウイルス病因の研究と遺伝的に定義されたワクチンの開発、およびZIKV感染の治療のための抗ウイルス薬の同定を容易にする。他のフラビウイルスについて説明したように、ZIKV感染性クローンの生成は、ウイルスゲノム14における不可解な細菌プロモーターの存在により困難であり、その伝播中に有毒なウイルスタンパク質の漏出発現を可能にする。標準的な高コピー数プラスミド15、16、17を使用する細菌中のcDNAクローン。この毒性の問題を克服するために、いくつかの非伝統的なアプローチは、過去2年間で正常に実装されています18.これらには、低コピー数プラスミド19、20、有毒領域21、22、23を破壊するイントロンの挿入、cDNA断片のインビトロライゲーションの使用が含まれる。24歳,25, ウイルスゲノムに存在する不可解な細菌プロモーターの突然変異サイレンシング26,27, 感染性亜ゲノムアンプリコン (ISA)28,29, ギブソン組み立て方法30、および円形ポリメラーゼ拡張反応(CPER)31の使用。
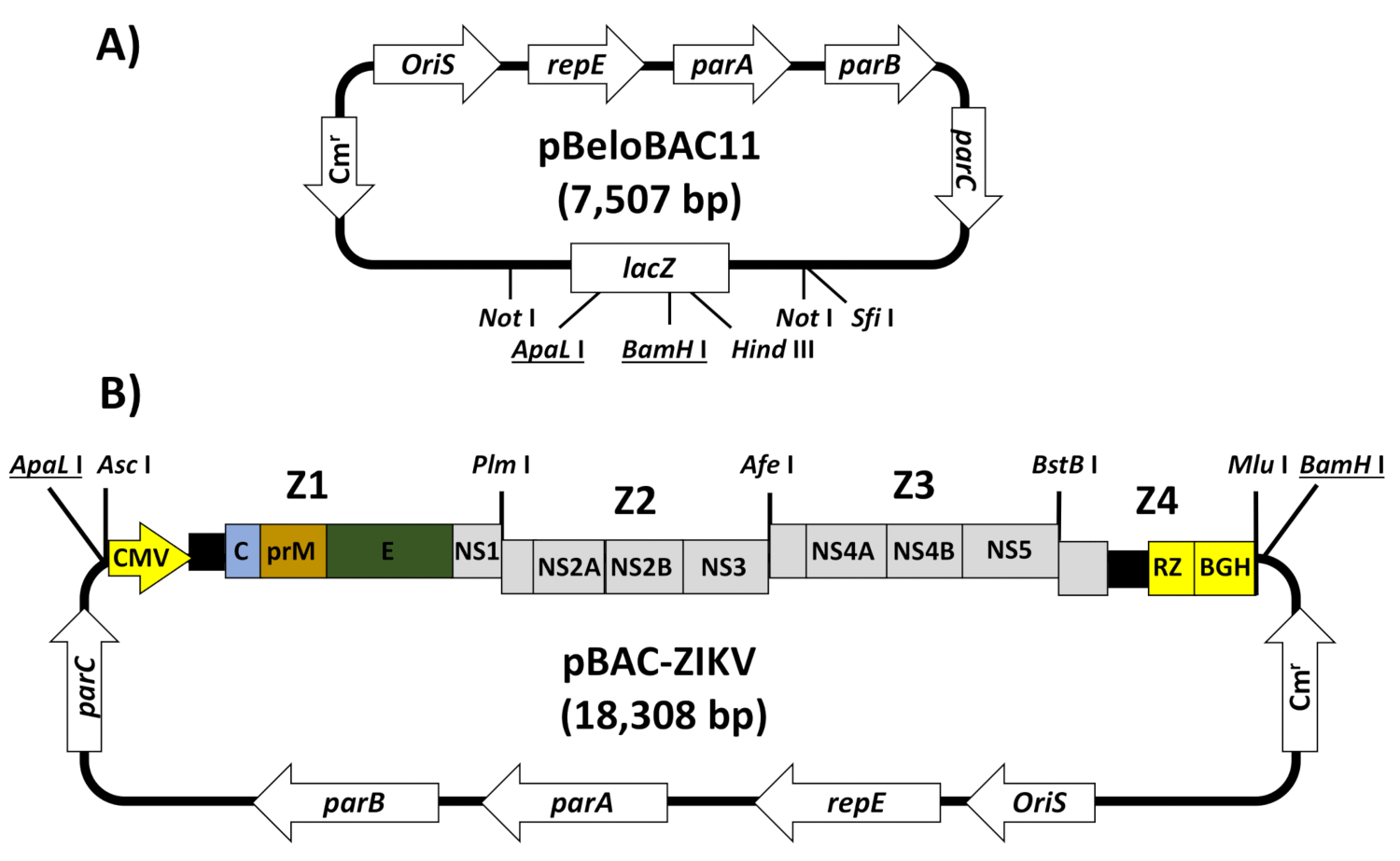
本明細書では、ZIKV株ZIKV-RGN13の全長cDNAクローンのエンジニアリングのための詳細なプロトコルについて説明し、毒性問題を克服するためにBACを使用し、およびVEroへのBAC cDNAクローンの直接トランスフェクションによる感染性rZIKVの救出について説明する。細胞32は、ヒトワクチン33の開発のために食品医薬品局(FDA)によって承認された細胞株である。このシステムでは、ウイルスゲノムの全長cDNAコピーをBACプラスミドpBeloBAC1134(図2A)に組み立て、エシェリヒア大腸菌F因子35に由来する低コピー数プラスミド(細胞当たり1~2部)、これは、細菌の伝播中にフラビウイルス配列の毒性を最小限に抑えます。ZIKVゲノムのcDNAは、ヒトCMV即時早期プロモーターの制御下でpBeloBAC11に組み立てられ、細胞RNAポリメラーゼIIによるトランスフェクト細胞の核におけるウイルス(v)RNAの発現を可能にし、肝炎によって3'末端に横たわるデルタウイルス(HDV)リボザイム(RZ)、続いてウシ成長ホルモン(BGH)終端およびポリアデニル化シグナルの配列が続き、ウイルスゲノムの本物の5'-および3'末端を有する合成RNAを産生する(図2B)。このcDNA起動システムは、キャップされたウイルスRNAの細胞内発現を生み出し、インビトロ転写ステップを必要とせずに感染性ZIKVの回復を可能にする。BACアプローチは、他のフラビウイルスおよび他の陽性鎖鎖RNAウイルス36、37、および他の陽性鎖ウイルスの安定した、完全に機能的なcDNAクローンの構築に適用可能な強力な方法論を提供する。38、39、40、41.
プロトコル
1. BACにおけるZIKV感染性cDNAクローンの構築
注:BACにおけるZIKVの組み立て戦略について、RGN株13(加盟番号KU527068)について説明する(図2)。
- プラスミドpBeloBAC11に存在しないウイルスゲノムに適切に間隔をあけた固有の制限部位を選択する(図2A)。
注:ZIKV-RGNでは、制限部位Pml I、Afe I、およびBstB I(ゲノム位置3,347、5,969、および9,127)がそれぞれ選択された。ウイルスゲノムに適切な制限部位が利用できない場合には、cDNA断片設計中にウイルスゲノムに無声ヌクレオチド変異を導入して新たな制限部位を生成する。 - 全長ゲノム(Z1〜Z4)にまたがる4つのcDNA断片を化学合成により生成し、5'末端およびHDV RZのCMVプロモーターによって側面付けされ、BGH終端、およびポリアデニル化配列を3'-end(図2B)で生成する。各フラグメントは、選択した制限サイトによって横たわっている必要があります (ステップ 1.1)。
注:フラグメント Z1 は、pBeloBac11 内の残りのフラグメントを複製するためのバックボーンとして使用されます。そのためには、ヒトCMVプロモーターを含む必要があり、ApaL I(pBeloBAC11でこの断片をクローンする)とAsc I(ウイルスゲノムに存在しない)によって5'-終端に横たわっていなければならない。.、Afe I、および BstB I)に続いて、Mlu I(ウイルスゲノムに存在しない)およびBamH I(pBeloBAC11でこの断片をクローンする)が続く。フラグメントZ4は、選択された最後の制限部位(BstB I)からウイルスゲノムの末端までのゲノム領域を含み、続いてHDV RZ、BGH終端、およびポリアデニル化配列、およびMlu I制限部位(図2B)を含む必要があります。あるいは、cDNA断片(Z1-Z4)は、標準的な逆転写酵素ポリメラーゼ連鎖反応(RT-PCR)と、特定のオリゴヌクレオチドを用いた重複するPcRの組み合わせによって生成され得る。 - 断片Z1からZ4の逐次クローニングにより感染性cDNAクローンをpBeloBAC11(図2B)に組み立てる。
- pBeloBAC11プラスミドとZ1フラグメントをApaL IおよびBamH Iで消化する。そのために、pBeloBAC11プラスミドの2μgまたはZ1フラグメントの1μgを10μLの10μLの反応バッファー、各酵素の20単位、および水を混合して、100 μLの最終体積に達する。
- エビアルカリホスファターゼ(SAP)を用いてBACベクターを脱リン酸化する。そのために、消化されたBACに2.5 μL(2.5単位)のSAPを加え、37°Cで1時間37°Cでインキュベートし、75°Cで15分間インキュベーションしてSAPを熱活性化させます。
- 10kbを超えるDNA断片の精製に最適化された市販のゲルクリーンアップキットを用いて、アガロースゲル電気泳動による脱リン酸化BACベクターおよびフラグメントZ1を精製する(材料の表を参照)。
- ライゲーション反応を行い、プラスミド(p)BAC-Z1を生成する。そのために、150ngの精製消化BACベクターを精製インサートと1:3のモル比を用いて精製インサートと混合し、10mM ATPを含む10x T4 DNAリゲスバッファーの1.5 μL、T4 DNAリゲスの1単位、および水を15μLの最終容積に混合する。
- ライゲーション混合物を16°Cで20時間インキュベートします。ライゲーション反応の制御として、挿入せずにライゲーション反応を並列に行う。65°Cでインキュベーションにより15分間リゲスを熱不活性化する。
注:鈍い終わりのDNの場合は、14°Cで20hのライゲーション反応をインキュベートする。BACベクター(図2A)の大きさに起因して、ライゲーション効率を高めるためには、従来のプラスミドを用いてライゲーションよりも多量のベクターとインサートを使用することが不可欠です。 - エレクトロポレーション(25 μF容量、2.5 kV、および100 Ω抵抗)によるライゲーション反応の2μL(ステップ1.3.5)を有する大腸菌DH10B電気容量細胞の50μLを変換し、0.2の間隔で間隔をあけた電極を装着した電極キュベットを使用して標準プロトコルに従います。
- 1 mLのSOC培地(2%トリプトン、0.5%酵母エキス、0.05%NaCl、2.5mM KCl、10mM MgCl 2、10mMMgSO 4、20mM[pH 7.0])で細胞を37°Cでインキュベートし、10°C(20°C)でインキュベートします(20)。12.5 μg/mLクロラムフェニコールを含むルリアブロス(LB)寒天プレートに細胞をプレートし、16時間37°Cでインキュベートします。
- 8~12個の細菌コロニーを選び、12.5μg/mLクロラムフェニコールを含む新鮮なLB寒天プレートにレプリカを作成し、特定のオリゴヌクレオチドを用いた直接PCR分析により正しい挿入物が含まれているかどうかをテストします。
- レプリカプレートから正のコロニーを選び、クロラムフェニコールの12.5μg/mLを含むLBの100mLで成長させ、市販のプラスミドミディキットを用いたアルカリ性リシス法によりBAC cDNAを単離し、大型の精製の推奨に従って低コピープラスミド(材料の表を参照)。
注:この方法で調製したBAC cDNAは、細菌ゲノムDNAの最大30%で汚染することができますが、制限分析、シーケンシング、クローニングを行うには品質に適しています。BACサイズに応じて、BACプラスミドの4〜6μgの収率が得られる。 - 制限分析により、クローンcDNAの遺伝的完全性を確認します。1時間37°CでAsc I及びPml Iを用いてpBAC-Z1プラスミドの500ngを消化し、ゲル電気泳動によるZ1断片の存在を確認した。望ましくない突然変異が導入されていないことをさらに確認するには、特定のオリゴヌクレオチドで挿入物を配列する。
- 選択された制限部位(Pml I、Afe I、BstB I、およびMlu I)を含むプラスミドpBAC-Z1から始まり、順次フラグメントZ2からZ4にクローンを作成し、全長感染性cDNAクローンpBAC-ZIKV(図2B)を生成し、同様の実験を行った。フラグメント Z1 (手順 1.3.1-1.3.10) に対して上記のようにアプローチします。
2. 感染性rZIKVの救助のための高純度pBAC-ZIKVの準備
注:感染性ウイルスのトランスフェクションおよび救助に適した超純型pBAC-ZIKV感染クローンの大規模な調製は、BAC精製のために特別に開発された市販キットを用いてアルカリ性リシスによって行われる(表参照) 材料)。キットには、細菌ゲノムDNA汚染を除去するATP依存性エキソヌクレアーゼ消化ステップが含まれている必要があり、塩化セシウム法で得られたものと同様の純度の高いBAC cDNAの単離を可能にする。
- pBAC-ZIKV感染性クローンを5mLのLB培地に含む大腸菌DH10Bの単一コロニーを37°Cで37°C(200-250rpm)で8時間成長させる。
- 2Lフラスコに12.5 μg/mLのクロラムフェニコールを含むLBの500 mLに細菌培養の1 mL(ステップ2.1)を加え、14-16h(0.6-0.8のOD600まで)37°Cで細胞を増殖させる。
注:素晴らしいスープ(TB)のような豊富なスープは、非常に高い細胞密度を生成することができ、その結果、BAC cDNAの収率が低く、純度が低くなります。 - BAC感染性cDNAクローンを、メーカーの仕様に従って、BAC精製用に特別に開発された市販キットを使用してアルカリリシスで精製します(材料の表を参照)。精製されたBAC cDNAを4°Cに保ちます。BACサイズに応じて、30μgの超純酸BAC cDNAクローンの収率を得ることができます。
注:過度の乾燥はBAC cDNAを溶解することが困難になるので、5分以上DNAペレットを乾燥させないようにしてください。BAC cDNAクローンのサイズが大きいため、プラスミド剪断を防ぐために渦やピペッティングを避けてください。
3. ベロ細胞のトランスフェクションによるBAC cDNAクローンからの感染性rZIKVの救出
注:感染性rZIKVは、市販のカチオン性脂質トランスフェクション試薬を用いて、pBAC-ZIKV cDNAクローンを用いたベロ細胞のトランスフェクションによって回収される(材料の表を参照)図3)
- トランスフェクションの前日、6ウェルプレート5 x 105ベロ細胞/成長培地(ダルベッコの改変イーグル培地[DMEM]は5%の胎児ウシ血清[FBS]、2mM L-グルタミン、1%非必須アミノ酸)を抗生物質なしで補充した。トランスフェクション時までに90%のコンフルエント細胞単層を上げる。
注:ウイルス救助を三循環で行うことをお勧めします。 -
室温で血清還元培地(材料表参照)を平衡化し、トランスフェクションサンプル毎に無菌マイクロフュージチューブ内のトランスフェクション混合物を以下のように調製する。
- 血清還元培地の250μLにBAC cDNAクローンの4 μgを加え、慎重に混ぜ合わせ、プラスミド剪断を防ぐために渦を避けます。
- 別のチューブでは、12μLのトランスフェクション試薬(1mg/mL)を血清還元培地の250μLで希釈し、渦で混合し、室温で5分間インキュベートする。
- 希釈されたBAC cDNAとトランスフェクション試薬(DNA:トランスフェクション試薬の1:3比)を組み合わせ、渦を避けながら慎重に混合し、室温で20〜30分間インキュベートします。
- BAC cDNA/トランスフェクション試薬混合物のインキュベーション期間中、抗生物質なしで成長培地でベロ細胞を洗浄し、抗生物質なしで新鮮な培地の1 mLで細胞を残します。
注:トランスフェクションプロセス中の抗生物質の添加は、トランスフェクション効率を低下させる可能性がある。 - BAC cDNA/トランスフェクション試薬混合物(ステップ3.2.3)の500 μLをVero細胞に滴下し、プレートを前後に揺さぶって混合し、5%CO2加湿インキュベーターで細胞を6時間37°Cでインキュベートします。
- トランスフェクション培地を取り除き、抗生物質(100 U/mLペニシリンおよび100 μg/mLストレプトマイシン)で2mLの新鮮な成長培地を追加し、37°Cで5%CO2加湿インキュベーターで細胞をインキュベートし、サイトパシー効果の誘導を毎日チェックします(CPE)).
注:ZIKV CPEはVero細胞で非常に顕著であり、組織培養上清に剥離して浮遊する丸みを帯びたビレフジエント細胞の存在によって特徴付げられる。 - 24時間ごとにベロ細胞組織培養上清(ステップ3.5)のアリコート(100μL)を収集し、新鮮なベロ細胞上の標準的なプラークアッセイを用いてZIKVの存在を評価してウイルス回収の効率を決定する(図4A)。
- トランスフェクションの4〜6日後、CPEが約50%〜75%(図4B)である場合、15mL円錐管および遠心分離機で組織培養上清を4°Cで10分間2,000xgで回収し、細胞破片を除去する。
- rZIKVを含む上清を収穫し、細胞ペレットを廃棄する。アリコートは凍結管で上清をアリコートし、さらに救助されたrZIKVの存在を確認するために-80 °Cでそれらを保存します。
4. 回収されたrZIKVの滴定
- 滴定の前日、12ウェルプレート上の種子2.5 x 105ベロ細胞/ウェル成長培地で滴定時までに90%のコンフルエント細胞単層を上昇させる。
注:回収されたrZIKVの滴定を三つトリケートで行うことはお勧めします。 - 冷凍庫から上清のアリコートを取り出し(ステップ3.8)、FBSなしで成長培地で連続10倍の希釈を行います。
- ベロ細胞をリン酸緩衝生理食塩水(PBS)で2倍洗浄し、各ウイルス希釈の200 μL/ウェルを三重に加えます。プレートを5%CO2加湿インキュベーターに37°Cで置き、90分間15分ごとに岩を打ちます。
- ウイルス接種を取り除き、2%FBS、1%DEAE-dextran、および0.6%寒天貴族を含む成長培地の2mLで細胞をオーバーレイし、3〜4日間5%CO2の下で37°Cでインキュベートする。
注:アガロース、メチルセルロース、または他のオーバーレイ媒体を使用することが可能です。適切なプラーク形成の時間は、使用されるオーバーレイとZIKV株によって異なります。 -
ZIKV感染細胞を室温でPBSで希釈した4%ホルムアルデヒドを1mL/ウェルで固定し、オーバーレイを除去し、室温で20%メタノールで0.1%の結晶バイオレットで15分間染色してウイルスプラークを可視化する。
- 結晶バイオレットを捨て、水で3倍洗い、プレートを乾燥させ、手動でプラークを数えます(図4C、左パネル)。ウイルスチターは、1ミリリットル当たりのプラーク形成単位(PFU/ml)として計算される。
-
あるいは、ウイルスプラークは、pan-flavivirus Eタンパク質マウスモノクローナル抗体(mAb)4G2(図4C、右パネル)による免疫染色により可視化することができる。
- 免疫染色によりZIKVプラークを評価するには、上述のようにオーバーレイ寒天の固定および除去後(ステップ4.5)、細胞をPBSで2倍に洗浄し、室温で15分間PBSで200 μL/ウェル0.5%のトリトン-X100をインキュベーションして透過させます。
- 透過性溶液を取り出し、細胞をPBSで3倍洗浄し、200μL/ウェルのブロッキング溶液(PBSで10%FBS)で室温で1時間ブロックします。
注:その他の標準的なブロッキングソリューション(例えば、PBSの2.5%BSA)は、このステップで使用することができる。 - ブロッキング溶液を除去し、37°Cで1時間のブロッキング溶液(1μg/mL)で希釈したパンフラビウイルスEタンパク質mAb 4G2の200μL/ウェルで細胞をインキュベートします。
注:他のmAbまたはポリクローナル抗体(pAb)は、ウイルスプラークの免疫染色および検出のために4G2の代わりに使用することができる。 - 細胞をPBSで3倍洗浄し、37°Cで1時間のブロッキング溶液中で希釈したビオチン化抗マウス二次抗体(メーカーの推奨に従って)の200μLでインキュベートします。
- 二次抗体を取り除き、PBSで細胞を4倍に洗浄し、メーカーの仕様に従ってアビジン/ビオチンベースのペルオキシダーゼキットを使用してウイルスプラークを可視化します(材料の表を参照)。
5. 成功したrZIKV救助の確認
注:駆出したウイルスの同一性をさらに確認するために、ZIKV Eタンパク質発現は、ZIKV Eタンパク質に特異的なマウスmAb 1176-56を用いて免疫蛍光によって分析される(図4D)。このmAbは、汎フラビウイルスEタンパク質mAb 4G2の状況とは対照的に、ZIKV Eタンパク質に特異的である(ステップ4.6.3)。あるいは、ウイルスのアイデンティティは、シーケンスによって確認することができる。
- 免疫蛍光分析の前日、種子は、感染時までに90%のコンフルエント細胞単層を上げるために成長培地で1 x 105ベロ細胞/ウェルを含む24ウェルプレートでスリップします。
- 細胞をPBSで2倍洗い、FBS(100 μL/ウェル)を含まない増殖培地で救助されたウイルスの0.5 PFU/細胞の多重感染(MOI)に感染します。プレートを37°Cで90分間インキュベートします。
- ウイルス吸着後、ウイルス接種を除去し、2%FBSで新鮮な成長培地の0.5mLを加え、37°Cで5%CO2加湿インキュベーターで細胞を48時間インキュベートする。
- 組織培養上清を取り除き、室温で20分間PBSで4%パラホルムアルデヒドの150μL/ウェルで細胞を固定し、室温で150μL/ウェル0.5%のトリトン-X100で細胞を透過化します。
- 透過性溶液を除去し、PBSで細胞を3倍に洗浄した後、室温で1時間の間に150μL/ウェルのブロッキング溶液で細胞をブロックする。
注:細胞スキャンは、代替ブロッキング溶液(例えば、PBSの2.5%BSA)でブロックされる。 - ブロッキング溶液を除去し、mAb 1176-56の100 μL/ウェルで細胞をインキュベートし、ZIKV Eタンパク質に特異的に、37°Cで1時間のブロッキング溶液(1μg/mL)で希釈する。
- 細胞をPBSで3倍洗浄し、Alexa Fluor 488-共役抗マウス二次抗体を100μL/ウェルで室温で1時間インキュベートし、ブロッキング溶液中で希釈した(メーカーの推奨に従って)ブロック溶液で細胞を広範囲に洗浄し、かつ、4',6'-diamidino-2-フェニリンドール(DAPI;1mg/mL)の150 μL/ウェルを室温で1:200に希釈し、室温で10分間浸入します。
- 細胞をPBSで3倍洗い、アンチフェード取り付け媒体にカバーリップを取り付け(材料の表を参照)、蛍光顕微鏡下でサンプルを分析します。
注:長期保存の場合は、光から保護された4°Cでサンプルを保管してください。
6. ウイルス株の増幅と生成
注:救出されたウイルスの同一性が確認されたら(セクション5)、Vero細胞上のウイルスを増幅し、さらなる研究のためにウイルスストックを生成する。
- 90%の合流点で100mm x 21 mmプレートでベロ細胞を成長させ、前述のように0.1 PFU/セルのMOIで感染させます。
- CPEが約75%(約48〜72時間の感染後)である場合は、50 mL円錐管で組織培養上清を採取し、4°Cで10分間2,000 x gで組織培養上清を回収し、細胞の破片を除去する。
- rZIKVを含む上清を収穫し、細胞ペレットを廃棄する。アリコートは、クライオチューブで上清をアリコートし、-80 °Cでそれらを保存します。
- 冷凍庫からウイルスアリコートを取り出し、前述のプラークアッセイでウイルス価火量を決定する(セクション4)。
結果
ここで説明するプロトコルは、いくつかのフラビウイルス配列に関連する毒性の問題を最小限に抑えるためにBACを使用して安定なZIKV全長cDNA感染クローンの生成を可能にする。BAC cDNAクローンからの感染性rZIKVの効率的な回収は、感受性ベロ細胞のトランスフェクション後に容易に達成することができる(図2)。このプロトコルを使用して、我々は、従来のクローニング方法とユニークな方法を使用して、BACプラスミドpBeloBAC1134に4つの重なり合うcDNA断片を順次クローニングすることにより、ZIKV株RGN 32の安定した全長cDNAクローンを生成しました。ウイルスゲノムに存在する制限部位(図2)。全長cDNAクローンは、トランスフェクト細胞の核内でvRNAの発現を可能にするためにヒトCMVプロモーターによって5'末端に横たわり、HDV RZによって3'末端に続いてBGHポリアデニル化および終端配列を、含むRNAを産生する。ウイルスゲノムの正確な3'-終わり(図2)。生成されたBAC cDNAクローンの細菌の安定性と、標準的な組換えDNA技術を用いて容易な操作が行われ、安定した全長cDNAクローンの迅速かつ信頼性の高い生成のためのBAC cDNAアプローチの可能性が強調されています。ZIKVおよび他のフラビウイルスまたは不安定なウイルスゲノムを有する陽性鎖RNAウイルス。
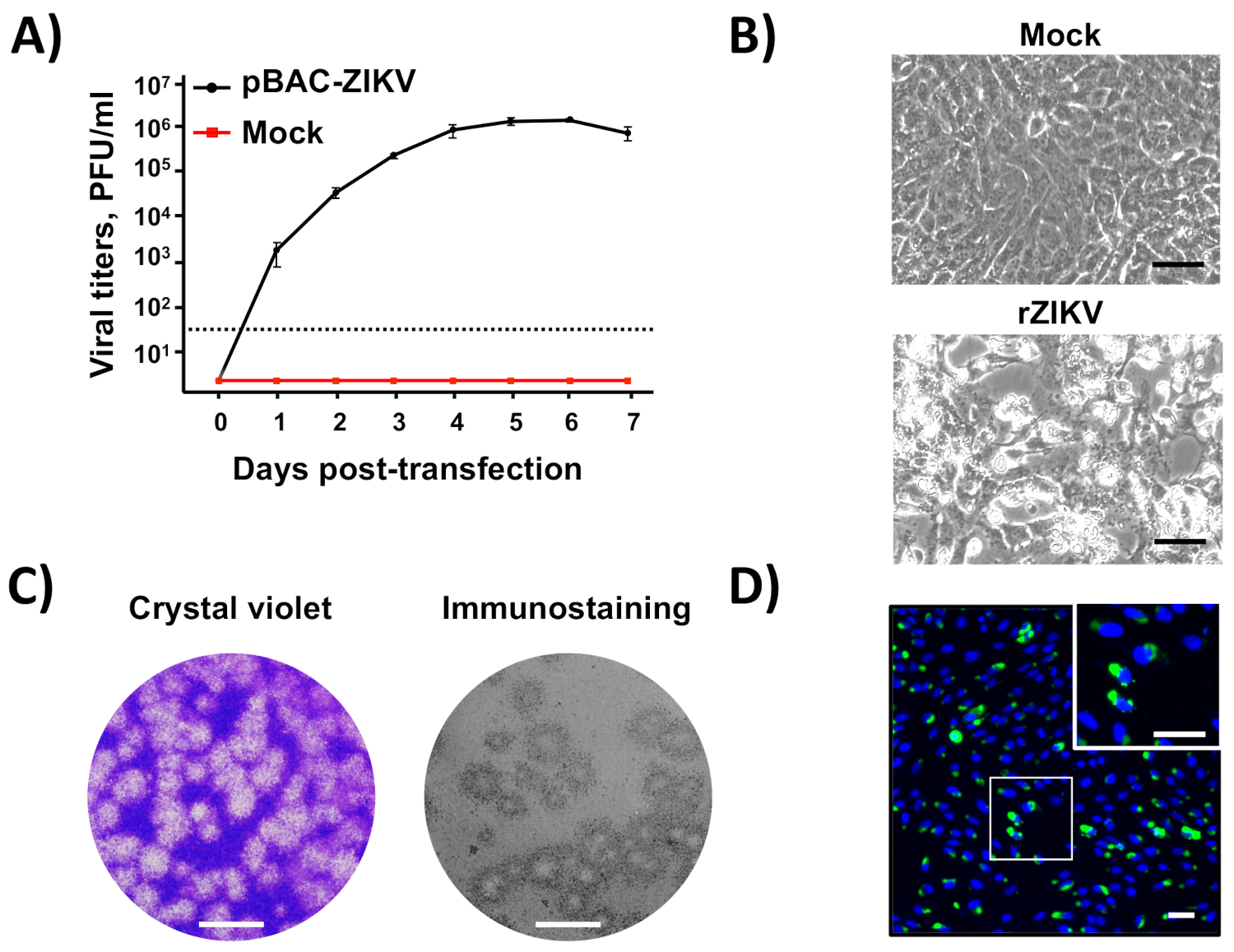
BAC cDNAクローンを組み立てた後(図2)、カチオン性リポソームを用いたBAC cDNAクローンを用いた感受性ベロ細胞の直接トランスフェクション後に感染性ウイルスを容易に回収できる(図3)。このcDNA起動システムは、キャップされたvRNAの細胞内発現を可能にし、インビトロ転写ステップを必要とせずに感染性ウイルスの回復を可能にした。このアプローチを用いて、トランスフェクション後5日間で106 PFU/mLより高い力価でrZIKV-RGNを救出することができました(図4A)。また、救助されたウイルスは、明確なCPE(図4B)を誘導し、約2mmの均質なプラーク(図4C)を生成し、その同一性をmAb特異的を用いてシーケンシング(図示せず)と免疫蛍光分析によって確認した。ZIKV Eタンパク質の場合、1176-56 (図 4D)インビトロデータは、回収されたrZIKV-RGNがベロ細胞および天然ZIKV単離32と比較してレベルに効率的に複製されたことを示す(データは示さない)。全体的に、これらの結果は、感染性rZIKVがBACで組み立てられた全長のcDNAクローンから救出できることを示している。

図 1: ZIKVゲノム組織とビリオン構造(A) ゲノム組織:ZIKVは、単一のポリタンパク質として翻訳される正の一本鎖RNAを含む。翻訳されたポリタンパク質をウイルス(矢印)および宿主(ダイヤモンド)プロテアーゼで切除し、構造タンパク質キャプシド(C、青)、マトリックス(M、茶色)、およびエンベロップ(E、緑色)、および7つの非構造タンパク質(NS1、NS2A、NS2B、NS3、NS4A、NS4B、NS5)を産生した。ウイルスゲノムの末端にある5'および3'未翻訳領域(UtR)は、黒い線で示されます。(B)ビリオン構造:ZIKVビリオンは、EおよびMタンパク質で飾られ、イコサヘドラ状構造を持つ脂質二重層に固定された。ウイルスエンベロープの下には、ウイルスゲノムRNAに関連するCタンパク質からなるウイルスヌクレオカプシドであった。この図は、アビラ・ペレスら18から適応されています.この図のより大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図 2: BACにおけるZIKV全長感染性cDNAクローンの組み立て。(A) pBeloBAC11 BACの概略表現:調節遺伝子parA、parB、parC、およびrepE、F因子複製起源(OriS)、クロラムフェニコール耐性遺伝子(Cm r)、およびlac Z遺伝子が示される。感染性ZIKV cDNAクローンの組み立てに使用される関連する制限部位には、下線が引かれています。(B) ZIKV全長感染性cDNAクローンをpBeloBAC11 BAC:4つの重なり合うDNA断片(Z1-Z4)を組み合わせ、ZIKVゲノム全体をカバーし(図1)、示された制限部位によって横たわって、化学的に生成したpBeloBAC11に合成し、順次クローン化して感染性ZIKV cDNAクローンpBAC-ZIKVを生成した。全長ZIKV感染性cDNAは、ヒトサイトメガロウイルス即時早期プロモーター(CMV)の制御下で組み立てられ、HDVリボザイム(RZ)およびウシ成長ホルモン(BGH)終端およびポリアデニル化配列によって3'末端に横たわっていた。ウイルス遺伝子および調節要素の頭字語は、図1に記載されているようにである。この図のより大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図 3: BAC cDNAクローンからrZIKVを生成するワークフロー。合流の90%におけるベロ細胞を、カチオン性リポソームを用いてZIKV全長感染性cDNAクローンpBAC-ZIKV(図2)を用いて単層にトランスフェクトした。4-6日後に、CPEが明らかになったとき、rZIKVを含む組織培養上清を採取し、ウイルスの存在について評価した(図4)。.この図のより大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}

図 4: rZIKVのリカバリとインビトロ特性。(A) BAC cDNAクローンからの感染性rZIKVの救助:合流の90%のベロ細胞(6ウェルプレートフォーマット、トリプリケート)を模擬トランスフェクトまたはpBAC-ZIKV(図3)の4μg/ウェルでトランスフェクションし、および移植後の指定日に、組織培養上清におけるウイルス力料は、プラークアッセイ(PFU/mL)によって決定した。誤差バーは、3つの異なるトランスフェクション実験からの標準偏差を示します。点線の黒い線は検出の限界を示します(50 PFU/mL)。(B) ウイルスCPE:90%合流(6ウェルプレートフォーマット、三価)のベロ細胞は、rZIKVで模擬感染(上)または感染(0.5 PFU/細胞のMOI)を有し、48h後感染時に、CPEの存在を光顕微鏡検査により評価した。スケールバー = 100 μm. (C)ウイルスプラークアッセイおよび免疫染色: 90%合流(12ウェルプレートフォーマット)のベロ細胞はrZIKVに感染し、72時間の感染後、ウイルスプラークは結晶紫色染色(左)または免疫染色によって可視化された(右)パンフラビウイルスEタンパク質mAb 4G2を用いて。スケールバー = 5mm. ( D) 免疫蛍光:90%合流(24ウェルプレートフォーマット、トリプリケート)のベロ細胞をrZIKVで感染させ(0.5 PFU/細胞のMOI)、48h後感染時に、mAb 1176-56を用いて免疫蛍光で分析し、ZIKVに特異的Eタンパク質。細胞核をDAPIで染色した。ZIKV感染ベロ細胞の代表的なマージ画像を示す。右上の白い正方形は、ZIKV感染ベロ細胞の拡大画像を表しています。スケールバー = 100 μm.この図のより大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}
ディスカッション
感染性cDNAクローンは、RNAウイルスの基礎研究やワクチンの開発および/または抗ウイルス戦略の同定に不可欠な分子ツールを構成します。しかし、フラビウイルスを含む多くの陽性鎖RNAウイルスの場合、標準的な高コピー数プラスミドを用いて細菌に伝播した場合、クローン化されたcDNAの不安定性のために感染性cDNAクローンの生成は困難である。ZIKVおよび他のフラビウイルスの場合、この不安定性は主にウイルスゲノム14、15、16、17に存在する不可解な細菌プロモーターからの毒性ウイルスタンパク質の漏出発現によるものである。.ここでは、毒性問題を克服するためにBACプラスミドpBeloBAC1134(図2A)を使用することに基づいて、単一のプラスミドとして安定なZIKV全長感染性cDNAクローンを生成するための代替かつ強力なプロトコルを説明する。CMVプロモーターは、トランスフェクト細胞の核におけるvRNAの発現を可能にし、HDV RZは正確な3'-終端を持つvRNAを生成する(図2B)。この方法を用いて、感受性ベロ細胞の直接トランスフェクション後の感染性rZIKVの効率的かつ信頼性の高い回復を可能にするZIKV株RGNの完全に安定した感染クローンの生成に成功しました(図3および図4)。
ここ数年、ZIKV感染性cDNAクローンに関連する不安定な問題を克服するために多大な努力がなされ、cDNA断片のインビトロライゲーションを含むいくつかのアプローチが18に成功しました24 ,25, 低コピープラスミド19,20, サイレント変異の導入による不可解な細菌プロモーターの不活性化 26,27, イントロン挿入21,22歳,23、 ギブソン組み立て方法30、 ISA 法28,29、 および CPER31の使用 .これらのアプローチは毒性の問題を克服し、ZIKV感染性cDNAクローンを生成するのに有用であるが、それらのいくつかは手間がかかり、ウイルスを減らすインビトロライゲーションおよび転写ステップの必要性を含むいくつかの欠点を提示する回復効率またはウイルスの適合性に影響を与える可能性のある不可解な細菌プロモーターを不活性化する多数の無声突然変異の導入。このプロトコルで説明するアプローチには、次のような利点があります。 i) BACプラスミドpBeloBAC1134は、厳密に制御された複製を有し、細胞当たりプラスミドのコピーを1つまたは2つ保持し、毒性を最小限に抑え、不安定なcDNの細菌の安定した維持を可能にする。ii)BACプラスミドの伝播および修飾は、大型のBAC-DNA断片および低コピープラスミドを操作するために、このプロトコルに記載されているわずかな修飾を考慮して、従来のプラスミドのそれらにほぼ類似している。ろよ、 キャッピングされたZIKV vRNAの細胞内発現およびインビトロ転写ステップを必要とせずに感染性ウイルスの回復。iv) 感染性rZIKVは、BAC cDNAクローンを用いて感受性細胞(例えば、ベロ)の直接トランスフェクション後に生成される。哺乳動物細胞におけるDNAトランスフェクションはRNAトランスフェクションよりも効率的であるため、BACアプローチによるウイルス回収効率はRNA転写物を用いて観察されたものよりも高く、培養細胞における通過数を減少させ、ウイルスストックを生成し、その結果、細胞培養適応による望ましくない突然変異の導入を制限する。
最後に、BACアプローチの可能性は、デングウイルス36を含む他のフラビウイルスの感染性cDNAクローンを設計するために、この方法(わずかな変更で)の成功によってサポートされ、および高い影響を及ぼすいくつかのコロナウイルスヒトおよび動物の健康、例えば透過性胃腸炎コロナウイルス37(TGEV)、ネコ感染性性性性性性性性性性性性性ウイルス38(FIPV)、ヒトコロナウイルスOC4339(HCoV-OC43)、重症急性呼吸器症候群コロナウイルス40(SARS-CoV)、および中東呼吸器症候群コロナウイルス41(MERS-CoV)、とりわけ。
ここで説明するプロトコルでは、考慮すべき重要な手順が 2 つあります。1つの重要な考慮事項は、BACプラスミドに存在しないウイルスゲノム中の適切な一意の制限部位を同定することです。十分な制限部位が利用できない場合、サイレントヌクレオチド変異の導入により、クローニング設計中に新しい制限部位を生成することができます。もう一つの重要な問題は、BACプラスミドが細胞あたり1つまたは2つのコピーにのみ存在し、したがって、細菌ゲノムDNAの高い汚染を有するBACプラスミドの低収率は、高および高のために設計された標準的なプロトコルを使用して得られるということです。中型コピー番号プラスミド。この潜在的な問題は、大量の培養量を使用して容易に克服し、BAC精製のために特別に開発された市販キットでBACプラスミドを精製します。
要約すると、我々は、これらの生物学の研究を容易にするために、他の陽性鎖RNAウイルスの安定した完全に機能的な感染性cDNAクローンを生成するために適応することができるBACの使用に基づいて強力なZIKV逆遺伝的アプローチを開発しました。ウイルスおよびワクチンの開発および/または抗ウイルス薬の同定を容易にする。
開示事項
著者は何も開示していない。
謝辞
著者らは、BAC cDNAクローン生成における彼女の技術的支援とスネザナ・ディミトロヴァがビデオの準備を手伝ってくれたことに感謝したいと思います。この研究は、スペイン経済競争力省(MINECO、助成番号BFU2016-79127-R)からF.A.T.と国立衛生研究所(NIH、助成番号1R21AI120500)からL.M.S.およびF.A.Tへの助成金によって一部支援されました。
資料
| Name | Company | Catalog Number | Comments |
| 1. Molecular Biology Reagents | |||
| Afe I | New England BioLabs | R0652S | 10,000 Units/mL |
| AmpliTaq DNA Polymerase | ThermoFisher Scientific (Applied Biosystems) | N8080161 | 5,000 Units/mL |
| ApaL I | New England BioLabs | R0507S | 10,000 units/mL |
| Asc I | New England BioLabs | R0558S | 10,000 Units/mL |
| BamH I | New England BioLabs | R0136S | 10,000 Units/mL |
| BstB I | New England BioLabs | R0519S | 20,000 Units/mL |
| Chloramphenicol | Sigma-Aldrich | C0378 | |
| ElectroMAX DH10B Cells | ThermoFisher Scientific (Invitrogen) | 18290015 | Electocompetent DH10B cells |
| Electroporation Cuvettes, 0.2 cm | Bio-Rad | 165-2086 | |
| Ethanol | Merck | 100983 | Flamable |
| Isopropanol | Merck | 109634 | Flamable |
| Large-Construct Kit (10) | QIAGEN | 12462 | For high-purity BAC preparation |
| LB Broth | ThermoFisher Scientific (Invitrogen) | 12780029 | Can be homemade as well |
| LB with Agar | ThermoFisher Scientific (Invitrogen) | 22700041 | Can be homemade as well |
| Methanol | Merck | 106009 | Flamable |
| Mlu I | New England BioLabs | R0198S | 10,000 Units/mL |
| Oligonucleotides | IDT | N/A | |
| Plasmid pBeloBAC11 | New England BioLabs | ER2420S (E4154S) | |
| Plasmid Midi Kit (25) | QIAGEN | 12143 | For midle-scale preparation of BAC plasmids |
| Pml I | New England BioLabs | R0532S | 20,000 Units/mL |
| Polypropylene tubes (10 mL) | DeltaLab | 175724 | Other commercial sources are acceptable |
| QIAEX II Gel Extraction Kit (150) | QIAGEN | 20021 | Gel-clean-up kit optimized for DNA fragments larger than 10 kb |
| Shrimp AlKaline Phosphatase (rSAP) | New England BioLabs | M0371S | 1,000 Units/mL |
| SOC Medium | ThermoFisher Scientific (Invitrogen) | 15544034 | Can be homemade as well |
| Synthesis of cDNA fragments | Bio Basic | N/A | |
| T4 DNA Ligase | Sigma-Aldrich (Roche) | 10481220001 | 1,000 Units/mL |
| 2. Cell Culture Reagents | |||
| 6-Well Plates | ThermoFisher Scientific (Nunc) | 140675 | |
| 12-Well Plates | ThermoFisher Scientific (Nunc) | 150628 | |
| 24-Well Plates | ThermoFisher Scientific (Nunc) | 142485 | |
| Agar Noble | VWR | 214230 | |
| Alexa Fluor 488 Conjugate ant-mouse secondary antibody | Varies | N/A | |
| Biotinylated Anti-Mouse Secondary Antibody | Varies | N/A | |
| Cell Culture Dishes (100 mm x 21 mm) | ThermoFisher Scientific (Nunc) | 172931 | |
| Conical Tubes (15 mL) | VWR | 525-0150 | |
| Conical Tubes (50 mL) | VWR | 525-0155 | |
| Crystal Violet | Sigma-Aldrich | C6158 | |
| DAPI | Sigma-Aldrich | D9542 | Toxic and carcinogenic |
| DEAE-Dextran | Sigma-Aldrich | D9885 | |
| DMEM | ThermoFisher Scientific (Gibco) | 11995065 | |
| Fetal Bovine Serum (FBS) | ThermoFisher Scientific (HyClone)) | SV30160.03 | |
| Formaldehyde | Sigma-Aldrich | F8775 | Toxic and carcinogenic |
| L-Glutamine | ThermoFisher Scientific (Gibco) | 25030081 | |
| Lipofectamine 2000 | ThermoFisher Scientific (Invitrogen) | 11668019 | Transfection reagent |
| Nonessential amino acids | ThermoFisher Scientific (Gibco) | 11140035 | |
| Opti-MEM I Reduced Serum Medium | ThermoFisher Scientific (Gibco) | 31985070 | Transfection medium |
| Pan-flavivirus E protein mAb 4G2 | BEI Resources | NR-50327 | |
| Paraformaldehyde | Electron Microscopy Sciences | 15710-S | Toxic and carcinogenic |
| PBS | ThermoFisher Scientific (Gibco) | 14190144 | |
| Penicillin/Streptomycin | ThermoFisher Scientific (Gibco) | 15140122 | |
| ProLong Gold Antifade Reagent | ThermoFisher Scientific (Invitrogen) | P10144 | |
| Triton-X-100 | Sigma-Aldrich | T8787 | |
| Vectastain ABC Kit | Vector Laboratories Inc | PK-4010 | Avidin/biotin-based peroxidase kit |
| Vero Cells | ATCC | CCL-81 | |
| ZIKV E Protein mAb 1176-56 | BioFront Technologies | BF-1176-56 | |
| 3. Equipment | |||
| Agarose Gel Electrophoresis System | Bio-Rad | 1704468 | Other commercial sources are acceptable |
| Class II Biosafety CO2 Cabinet | Varies | N/A | Other commercial sources are acceptable |
| Desktop Refrigrated Centrifuge | Varies | N/A | |
| Fluorescence Microscope | Varies | N/A | |
| High-speed Refrigrated Centrifuge | Varies | N/A | |
| MicroPulser Electroporator | Bio-Rad | 1652100 | Other machines are acceptable |
| SimpliAmp Thermal Cycler | ThermoFisher Scientific (Applied Biosystems) | A24811 | Other machines are acceptable |
| Vortexer | Varies | N/A |
参考文献
- Friedrich, M. J. WHO Calls Off Global Zika Emergency. JAMA. 317 (3), 246 (2017).
- Sirohi, D., et al. The 3.8 Å resolution cryo-EM structure of Zika virus. Science. 352 (6284), 467-470 (2016).
- Lindenbach, B. D., Murray, C. J., Thiel, H. J., Rice, C. M., Knipe, D. M., Howley, P. M. Flaviviridae. Fields Virology. , 712-748 (2013).
- Boeuf, P., Drummer, H. E., Richards, J. S., Scoullar, M. J., Beeson, J. G. The global threat of Zika virus to pregnancy: epidemiology, clinical perspectives, mechanisms, and impact. BMC Medicine. 14 (1), 112 (2016).
- Simpson, D. I. Zika virus infection in man. Transactions of The Royal Society of Tropical Medicine and Hygiene. 58, 335-338 (1964).
- Campos, G. S., Bandeira, A. C., Sardi, S. I. Zika Virus Outbreak, Bahia, Brazil. Emerging Infectious Diseases. 21 (10), 1885-1886 (2015).
- Cao-Lormeau, V. M., et al. Zika virus, French polynesia, South pacific, 2013. Emerging Infectious Diseases. 20 (6), 1085-1086 (2014).
- Faria, N. R., et al. Zika virus in the Americas: Early epidemiological and genetic findings. Science. 352 (6283), 345-349 (2016).
- Costello, A., et al. Defining the syndrome associated with congenital Zika virus infection. Bulletin of the World Health Organization. 94 (6), 406-406A (2016).
- Cugola, F. R., et al. The Brazilian Zika virus strain causes birth defects in experimental models. Nature. 534 (7606), 267-271 (2016).
- do Rosario, M. S., et al. Guillain-Barre Syndrome After Zika Virus Infection in Brazil. American Journal of Tropical Medicine and Hygiene. 95 (5), 1157-1160 (2016).
- Miner, J. J., et al. Zika Virus Infection during Pregnancy in Mice Causes Placental Damage and Fetal Demise. Cell. 165 (5), 1081-1091 (2016).
- Mlakar, J., et al. Zika Virus Associated with Microcephaly. The New England Journal of Medicine. 374 (10), 951-958 (2016).
- Li, D., Aaskov, J., Lott, W. B. Identification of a cryptic prokaryotic promoter within the cDNA encoding the 5' end of dengue virus RNA genome. PLOS ONE. 6 (3), e18197 (2011).
- Aubry, F., Nougairede, A., Gould, E. A., de Lamballerie, X. Flavivirus reverse genetic systems, construction techniques and applications: a historical perspective. Antiviral Research. 114, 67-85 (2015).
- Pu, S. Y., et al. Successful propagation of flavivirus infectious cDNAs by a novel method to reduce the cryptic bacterial promoter activity of virus genomes. Journal of Virology. 85 (6), 2927-2941 (2011).
- Ruggli, N., Rice, C. M. Functional cDNA clones of the Flaviviridae: strategies and applications. Advances in Virus Research. 53, 183-207 (1999).
- Avila-Perez, G., Nogales, A., Martin, V., Almazan, F., Martinez-Sobrido, L. Reverse Genetic Approaches for the Generation of Recombinant Zika Virus. Viruses. 10 (11), (2018).
- Annamalai, A. S., et al. Zika Virus Encoding Nonglycosylated Envelope Protein Is Attenuated and Defective in Neuroinvasion. Journal of Virology. 91 (23), (2017).
- Shan, C., et al. An Infectious cDNA Clone of Zika Virus to Study Viral Virulence Mosquito Transmission, and Antiviral Inhibitors. Cell Host & Microbe. 19 (6), 891-900 (2016).
- Liu, Z. Y., et al. Characterization of cis-Acting RNA Elements of Zika Virus by Using a Self-Splicing Ribozyme-Dependent Infectious Clone. Journal of Virology. 91 (21), (2017).
- Schwarz, M. C., et al. Rescue of the 1947 Zika Virus Prototype Strain with a Cytomegalovirus Promoter-Driven cDNA Clone. mSphere. 1 (5), (2016).
- Tsetsarkin, K. A., et al. A Full-Length Infectious cDNA Clone of Zika Virus from the 2015 Epidemic in Brazil as a Genetic Platform for Studies of Virus-Host Interactions and Vaccine Development. MBio. 7 (4), (2016).
- Deng, C. L., et al. Recovery of the Zika virus through an in vitro ligation approach. Journal of General Virology. 98 (7), 1739-1743 (2017).
- Widman, D. G., et al. A Reverse Genetics Platform That Spans the Zika Virus Family Tree. mBio. 8 (2), (2017).
- Munster, M., et al. A Reverse Genetics System for Zika Virus Based on a Simple Molecular Cloning Strategy. Viruses. 10 (7), (2018).
- Zhao, F., et al. Negligible contribution of M2634V substitution to ZIKV pathogenesis in AG6 mice revealed by a bacterial promoter activity reduced infectious clone. Scientific Reports. 8 (1), 10491 (2018).
- Atieh, T., Baronti, C., de Lamballerie, X., Nougairede, A. Simple reverse genetics systems for Asian and African Zika viruses. Scientific Reports. 6, 39384 (2016).
- Gadea, G., et al. A robust method for the rapid generation of recombinant Zika virus expressing the GFP reporter gene. Virology. 497 (Supplement C), 157-162 (2016).
- Weger-Lucarelli, J., et al. Development and Characterization of Recombinant Virus Generated from a New World Zika Virus Infectious Clone. Journal of Virology. 91 (1), (2017).
- Setoh, Y. X., et al. De Novo Generation and Characterization of New Zika Virus Isolate Using Sequence Data from a Microcephaly Case. mSphere. 2 (3), (2017).
- Marquez-Jurado, S., et al. An Alanine-to-Valine Substitution in the Residue 175 of Zika Virus NS2A Protein Affects Viral RNA Synthesis and Attenuates the Virus In Vivo. Viruses. 10 (10), (2018).
- Cheng, B. Y. H., Ortiz-Riaño, E., de la Torre, J. C., Martínez-Sobrido, L. Generation of Recombinant Arenavirus for Vaccine Development in FDA-Approved Vero Cells. Journal of Visualized Experiments. (78), (2013).
- Wang, K., Boysen, C., Shizuya, H., Simon, M. I., Hood, L. Complete nucleotide sequence of two generations of a bacterial artificial chromosome cloning vector. BioTechniques. 23 (6), 992-994 (1997).
- Shizuya, H., et al. Cloning and stable maintenance of 300-kilobase-pair fragments of human DNA in Escherichia coli using an F-factor-based vector. Proceedings of the National Academy of Sciences of the United States of America. 89 (18), 8794-8797 (1992).
- Usme-Ciro, J. A., Lopera, J. A., Enjuanes, L., Almazan, F., Gallego-Gomez, J. C. Development of a novel DNA-launched dengue virus type 2 infectious clone assembled in a bacterial artificial chromosome. Virus Research. 180, 12-22 (2014).
- Almazan, F., et al. Engineering the largest RNA virus genome as an infectious bacterial artificial chromosome. Proceedings of the National Academy of Sciences of the United States of America. 97 (10), 5516-5521 (2000).
- Balint, A., et al. Molecular characterization of feline infectious peritonitis virus strain DF-2 and studies of the role of ORF3abc in viral cell tropism. Journal of Virology. 86 (11), 6258-6267 (2012).
- St-Jean, J. R., et al. Recovery of a neurovirulent human coronavirus OC43 from an infectious cDNA clone. Journal of Virology. 80 (7), 3670-3674 (2006).
- Almazan, F., et al. Construction of a severe acute respiratory syndrome coronavirus infectious cDNA clone and a replicon to study coronavirus RNA synthesis. Journal of Virology. 80 (21), 10900-10906 (2006).
- Almazan, F., et al. Engineering a replication-competent, propagation-defective Middle East respiratory syndrome coronavirus as a vaccine candidate. mBio. 4 (5), e00650-e00613 (2013).
- Jamsai, D., et al. Targeted modification of a human beta-globin locus BAC clone using GET Recombination and an I-Scei counterselection cassette. Genomics. 82 (1), 68-77 (2003).
- Lee, E. C., et al. A highly efficient Escherichia coli-based chromosome engineering system adapted for recombinogenic targeting and subcloning of BAC DNA. Genomics. 73 (1), 56-65 (2001).
- Tischer, B. K., von Einem, J., Kaufer, B., Osterrieder, N. Two-step red-mediated recombination for versatile high-efficiency markerless DNA manipulation in Escherichia coli. BioTechniques. 40 (2), 191-197 (2006).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved