
Method Article
ショウジョウバエメラノガスターにおける軸索死における軸索とそのシナプスの形態学的および機能的評価
要約
ここでは、ショウジョウバエメラノガスターで3つの単純な傷害誘発軸索変性(軸索死)アッセイを行い、切断された軸索とそのシナプスの形態学的および機能的保存を評価するプロトコルを提供する。
要約
軸索変性は神経変性疾患の共有機能であり、神経系が機械的または化学的な力によって挑戦されるとき。しかし、軸索変性の根底にある分子メカニズムに対する我々の理解は依然として限られている。傷害誘発軸索変性は、切断された軸索が独自の分解(軸索死)を実行する方法を研究するための簡単なモデルとして機能します。近年、ハエから哺乳類への進化的に保存された軸索死シグナル伝達カスケードが同定されており、分離された軸索が傷害後に退化するために必要とされている。逆に、減衰した軸索死シグナル伝達は、切断された軸索およびそのシナプスの形態学的および機能的保存をもたらす。ここでは、軸索形態の観察、またはニューロン細胞体から切断された切断された軸索の軸索およびシナプス機能を、ショウジョウバエの果実飛行において観察することを可能にする3つの簡単かつ最近開発されたプロトコルを提示する。形態は翼で観察することができ、部分的な傷害は同じ神経束内の損傷していない対照軸索の軸索の並んで死亡をもたらす。あるいは、神経束全体がアンテナアブレーションによって引き起こされる軸索死を受ける脳でも軸索形態が観察される。切断された軸索とそのシナプスの機能的保存は、シナプス後のグルーミング行動と相まって単純な光遺伝学的アプローチによって評価することができる。高線機能喪失突然変異を用いた例と、過剰発現のdnmnatにより、いずれも数週間から数ヶ月間軸索死を遅延させることができる例を提示する。重要なことに、これらのプロトコルは、損傷を超えて使用することができます。それらは神経の維持因子、軸索輸送および軸索のミトコンドリアの特徴付けを促進する。
概要
ニューロンの形態学的完全性は、生涯を通じて持続的な神経系機能に不可欠です。ニューロン容積の大部分は軸索11,22によって取られる。したがって、特に長い軸索の生涯維持は、神経系にとって主要な生物学的および生体エネルギー的課題である。複数の軸索組み込みおよびグリア外因性支持機構が同定され、生涯にわたる軸索生存を保証している。彼らの障害は、神経系が病気で挑戦されている共通の特徴である軸索変性3、および機械的または化学的な力44、55をもたらす。しかし、軸索変性の根底にある分子メカニズムはいかなる文脈においても十分に理解されておらず、軸索損失を阻止するための効果的な治療法の開発は困難である。これらの神経学的状態に対する効果的な治療法の開発は、私たちの社会に大きな負担を生み出すため、重要です。
傷害誘発軸索変性は、切断された軸索が独自の分解を実行する方法を研究するための簡単なモデルとして機能します。1850年にアウグストゥス・ウォーラーにちなんで発見され、名前が付けられたウォレリア変性(WD)は、2つの異なる分子分離可能なプロセス7から成る傘用語です。第一に、軸索損傷後、細胞体から分離された軸索は、傷害8後1日以内に進化的に保存された軸索死シグナルカスケードを通じて、自らの自滅(軸索死)を積極的に実行する。第二に、周囲のグリアと専門の食細胞は、3〜5日以内に得られた軸索の破片を従事し、クリアします。軸索死シグナル伝達の減衰,は、,週99、10、11、1210,11のために保存されたままの切断軸索をもたらすが、グリア巻き込みの減衰は、生体内13、14、15,14で数週間持続する軸索の破片で最高潮に達する。1215
ハエ、マウス、ラットおよびゼブラフィッシュの研究は、軸索死シグナル伝達のいくつかの進化的に保存され、不可欠なメディエーターを明らかにした。軸索死変異体には、軸索死を受けることができない切断された軸索およびシナプスが含まれている。これらは、,細胞体サポート,,,,,,,179、10、12、13、16、17、18、19、20、21、22、2310の細胞体サポートがない場合、数週間、形態学的および機能的に保存されたままである。,1222,239131618192021これらのメディエーターの発見と特徴付けは、軸索死を実行する分子経路の定義につながった。重要なことに、軸索死シグナル伝達は、軸索が切断、粉砕または伸ばされたときに限らず活性化される24,,25;また、神経学的状態の別個の動物モデルの貢献者であるようです(例えば、軸索が傷害非依存的な様式で退化する4、まだ有益な結果の範囲を持つ4、8)。84したがって、軸索死が傷害後に軸索変性を実行する方法を理解することは、単純な傷害モデルを超えた洞察を提供するかもしれない。また、治療介入の標的を提供することもできる。
ショウジョウバエのメラノガスター (ショウジョウバエ)は、軸索死シグナル伝達のための非常に貴重なシステムであることが証明されています。ハエの研究は、4つの本質的に進化的に保存された軸索死遺伝子を明らかにした:ハイワイヤー(hiw)11、14、dnmnat11,1412、26、dsarm,26 dsarm10およびaxundead(軸)12。 dnmnatこれらのメディエーターの改変 - hiw 、dsarmとaxedの機能喪失突然変異、およびdnmnatの過剰発現 - 強力にハエの寿命のための軸索死をブロックします。切断された野生型軸索は1日以内に軸索死を受けるが、切断された軸索とそのシナプスはヒュウ、ダルムまたは軸を欠いているが、形態学的にだけでなく、機能的に数週間保存されたままである。高レベルのダンナを介して機能的保存も達成できるかどうかは未定である。
ここでは、細胞体サポートがない場合に軸索死(例えば、切断された軸索とそのシナプスの形態および機能)を研究するための3つの簡単で最近開発されたプロトコルを紹介します。我々は、アクソン死が、ヒヨー機能喪失突然変異(hiwΔ hiwN)で形態学的に保存される切断軸索をもたらす方法と、減弱された軸索およびシナプスが、少なくとも7日間機能的に保存され、徐引な軸索およびシナプスをもたらす方法を示す(dnmnatOE)。これらのプロトコルは、中央または末梢神経系(それぞれCNSおよびPNS)13,14,14のいずれかで個々の軸索およびシナプス形態の観察を可能にする一方、CNSにおける切断された軸索およびそれらのシナプスの機能的保存は、行動的読み出し12としてグルーミングと結合された単純な光遺伝学的セットアップの使用によって視覚化することができる。
プロトコル
PNSにおける軸索死時の軸索形態の観察
- 翼損傷:軸索束の部分的な損傷
- 5人の処女女性と5人の男性を右遺伝子型(図4A、P0世代)で使用し、室温(RT)で十字架を行います。0P0を 3 ~ 4 日ごとに新しいバイアルに渡します。毎日、閉鎖されたばかりの成人の子孫(F1世代)を収集し、7〜14日間年齢を重用します。
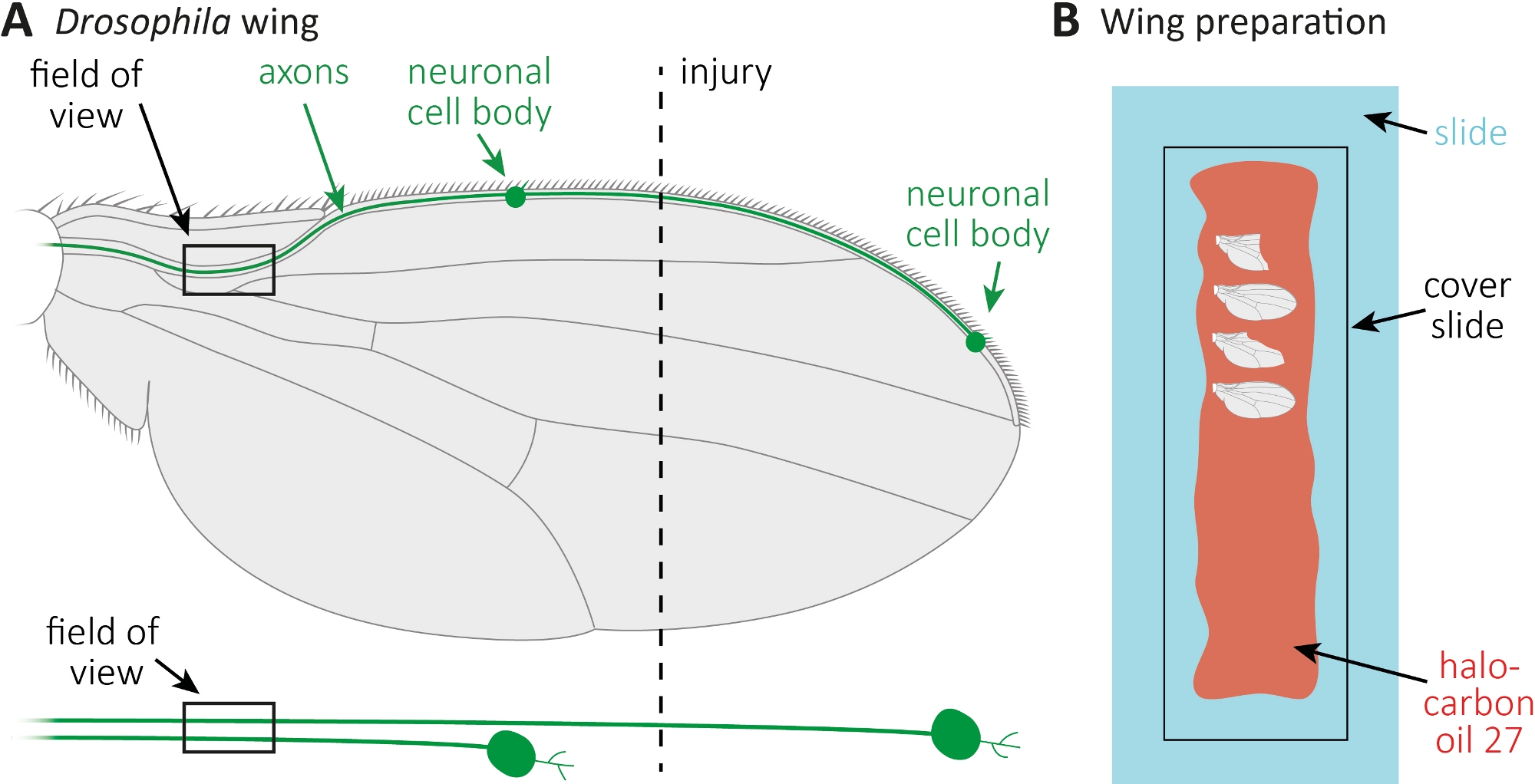
- CO2パッド上のハエを麻酔します。マイクロハサミを使用して、翼の中央に大まかに前翼静脈を切断する(図1A)。一方の翼を怪我に、もう1つの翼を年齢に合わせた怪我のないコントロールとして使用します。翼ごとに1つの怪我を適用し、十分な翼(約15の翼)を負傷させる必要があります。
注:翼全体を切り抜くことができますが、前翼静脈のみを切断するだけで十分です。これは翼の最も強い部分です。 - ハエを食物含有バイアルで回収する。
- 軸索の翼解剖と可視化
- ガラススライド全体に沿ってピペットを用いて10μLのハロカーボンオイル27を広げる(図1B)。
- 負傷した場合は、負傷したコントロールウィングを必要な時点(例えば、1日または7日の負傷後)に切り落とします。マイクロハサミを使ってカットし、ピンセットで翼をつかみます。最大4つの翼をハロカーボンオイル27(図1B)に取り付け、カバースライドで覆います。
- 回転ディスク顕微鏡を使用して、すぐに翼をイメージします。0.33 μmのステップサイズを持つZ軸に沿った一連の光学断面を取得し、zスタックを1つのファイルに圧縮して、その後の解析を行います。
注意:細胞のボディと軸索が収容されている前翼の静脈をつかむべきしないでください。中央の翼をつかむ。翼の組織は固定されていません。翼を取り付けるまでの時間を8分以内にイメージングするようにしてください。

図1:翼の軸索死時の軸索形態の観察(A)2つのまばらなGFP標識感覚ニューロンを持つスケマティックフライウィング(以下にも個別に示す)。負傷部位と観察分野が示されている。(B) 翼イメージング用の回路図設定。負傷および無傷のコントロール翼(灰色)は、ガラススライド(水色)にハロカーボンオイル27(赤)に取り付けられ、カバースライド(黒)で覆われています。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}
2. CNSにおける軸索死時の軸索とシナプス形態の観察
- アンテナアブレーション:軸索バンドル全体の損傷
- 右の遺伝子型から5人の処女女性と5人の男性(図5A、P0世代)を使用してRTでクロス0を行います。毎日、閉鎖されたばかりの成人子孫(F1世代)を収集し、7歳から14日間の年齢をとらせます。
- CO2パッド上のハエを麻酔します。ピンセットを使用して、一方的なアブレーションのために右の3番目のアンテナセグメントをアブレーションします。または左右の 3番目のアンテナ セグメントの両方を両側アブレーションに使用します (図 2A-C)。これは、GFP標識ニューロン細胞体を除去し、軸索投影はCNSに残る。
注: アンテナ アブレーションは軸索バンドル全体を切断します。一方的なアブレーションが行われる場合、対側側側(未アパックアンテナ)の軸索束が内部制御として機能します。十分なアンテナアブレーション(約15匹)を実行してください。 - ハエを食物含有バイアルで回収する。
- 脳の解剖と軸索の可視化
- シリコーンエラストマーベース(9 mL)と硬化剤(1 mL)を10:1の体積比で混合します。各5 mL混合物を35mmの組織培養プレートに移し、一晩のヒュームフードで穏やかな攪拌と混合することによって導入される空気を減らします。混合物は24時間以内に固化する。
注: 解剖プレートは1回だけ準備する必要があり、複数回使用することができます。 - CO2パッド上のハエを麻酔し、必要な時点(例えば、アンテナアブレーションの1または7日後)に2つのピンセットを使用して成人の頭部を切断する。1つのトゥイザーを使用して首をつかみ、もう1つのトゥイザーを使用して胸郭を固定します。首をそっと引っ張り、胸郭から頭を離します。
注:目的の数が達成されるまで、CO2パッドに首を切られたヘッドを残しますが、30分以内に次のステップに進んでください。 - 4%パラホルムアルデヒド(PFA)を含む固定溶液1mLを含む1.5mLマイクロ遠心分離チューブに、固定溶液に浸したピンセットを使用してリン酸緩衝生理食塩水(PBS)に0.1%トリトンX-100を入れます。
注:フライヘッドは濡れたピンセットによく付きます。マイクロ遠心チューブに容易にすべてのヘッドを移すことを可能にする。 - RTで穏やかな攪拌で20分間ヘッドを固定します。氷の上にマイクロ遠心分離管を置くと、ヘッドはマイクロ遠心分離管の底に引き寄せられます。ピペットで上澄み液を取り出し、RTで穏やかな攪拌を伴うPBSで0.1%トリトンX-100を含む洗浄バッファーの1 mLで5つの2分洗浄でこの手順を繰り返し、残留固定溶液を除去する。
注:成人ショウジョウバエの脳を解剖する方法に関するビデオは、容易に利用可能です27. - ガラスピペットでヘッドを洗浄バッファーで満たされた解剖プレートに移します。1つのトゥイザーを使用して、他のトゥイザーで頭を保持しながら、頭からプロボシスをつかんで引っ張ります。これは、プロボシスが外骨格に取り付けられた穴を残します。
- 穴と各複合眼の間の外骨格を除去するために2つのピンセットを使用してください。これは、両方のピンセットで頭の構造を開き、内の脳を穏やかに掻き出すことを可能にします。
- 気管や脂肪を取り除くことによって、各脳を洗浄します(図2D,上)。脳が洗浄されたら、氷の上に洗浄バッファーの1 mLを含む新しいマイクロ遠心チューブに入れます。
メモ:損傷または失われた光葉は、脳の中心にある嗅覚ローブに影響を与えない(図2D、上)。 - すべての脳が収集され、マイクロ遠心チューブの底に蓄積されたら、洗浄バッファーを固定溶液の1 mLに置き換えます。RTでロッキングで10分間脳を固定し、RTでロッキングして洗浄バッファの1mLに5つの2分の洗浄が続きます。
- 一次抗体(1:500)を4°Cで揺らしながら一晩洗浄バッファーに適用し、RTでロッキングして1mLの洗浄バッファーを使用して2時間にわたって10回の洗浄を行います。
- RTで揺れる洗浄バッファー2時間に二次抗体(1:500)を塗布し、マイクロ遠心分離管をアルミニウム箔で包んで光を遮断する。手順の残りの部分については、マイクロ遠心チューブをアルミホイルで覆ったままにしておきます。RTで揺れ動いて2時間にわたって1mLの洗浄バッファーで10の洗浄を適用します。
- 上清を取り除き、マイクロ遠心チューブの脳を覆うためにアンチフェード試薬の一滴を使用してください。4°Cで少なくとも30分間脳をインキュベートしてから、取り付けとイメージングの準備をします。
- カバー スライドを用意し、ラボ用テープを貼り付け、テープから "T" のような形を切り取ります (図 2D、下)。得られた空間は、脳含有抗フェード試薬28が両方のチャンバに配管される領域として機能する。
注:チップの開口部を広げるために、チップの3mmが切断されている20-200 μLピペットチップを使用してください。これは、脳を含む抗フェード試薬をピペットすることが可能になります.慎重にカバースライドで脳をカバーします。 - 粘土を使用して、2つの小さな偶数ロールを準備します。粘土ロールがガラススライドの高さより高くないことを確認します。粘土ロールをガラススライド(図2D、底)に貼り付けます。粘土ロールの上に脳を含むカバースライドサンドイッチを置きます。
注:GFPラベル付き軸索とそのシナプスは、脳の前部にあります。したがって、それらを正面からイメージ化する方が簡単です。しかし、脳は上向きになるか、カバースライドサンドイッチの上を下に向きます。粘土ロールはサンドイッチホルダーとして機能し、イメージング中にサンドイッチを逆さまに反転させることができます。これにより、すべての脳から正面から画像を取得することが可能になります。 - 共焦点顕微鏡を使用してz軸に沿った一連の光学断面を1.0 μmのステップサイズで取得し、zスタックを1つのファイルに圧縮して後の分析を行い、残りの軸索投影の数を評価します。
- シリコーンエラストマーベース(9 mL)と硬化剤(1 mL)を10:1の体積比で混合します。各5 mL混合物を35mmの組織培養プレートに移し、一晩のヒュームフードで穏やかな攪拌と混合することによって導入される空気を減らします。混合物は24時間以内に固化する。

図2:脳内の軸索死時の軸索とシナプスの形態の観察(A) GFP ラベルのセルボディ、軸索、シナプスを持つ概略フライヘッドの側面図。(B) GPF標識嗅覚受容体ニューロンとその軸索およびシナプスの高倍率の正面図。セルボディは3番目のアンテナセグメントに収納され、軸索はCNSに投影されます。軸索は、左嗅糸葉の糸球体にシナプスを形成し、中線を越え、逆側嗅覚葉に糸球体にシナプスを形成する。(C)片側アンテナアブレーションを持つフライヘッドの例。トップ:怪我のないコントロール。中央: 3番目のアンテナ セグメントのアブレーション。下: 2番目の (したがって、3rd)アンテナ セグメントのアブレーション。(D) 脳の準備.上:視野に示された嗅覚葉と軸索突起を有する模式的に解剖されたフライ脳。下:脳イメージングのための回路図設定。2つの粘土ロール(緑)はガラススライド(水色)に取り付けられ、フライブレイン(グレー)を含むカバースライドサンドイッチを運びます。脳はアンチフェード試薬(紫色)に取り付けられ、ラボテープ(オレンジ)で囲まれ、2枚のカバースライド(黒)で覆われています。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}
3. アキソンとシナプス機能の読み出しとして光遺伝学によって誘導されるグルーミング
- 光遺伝学的セットアップ
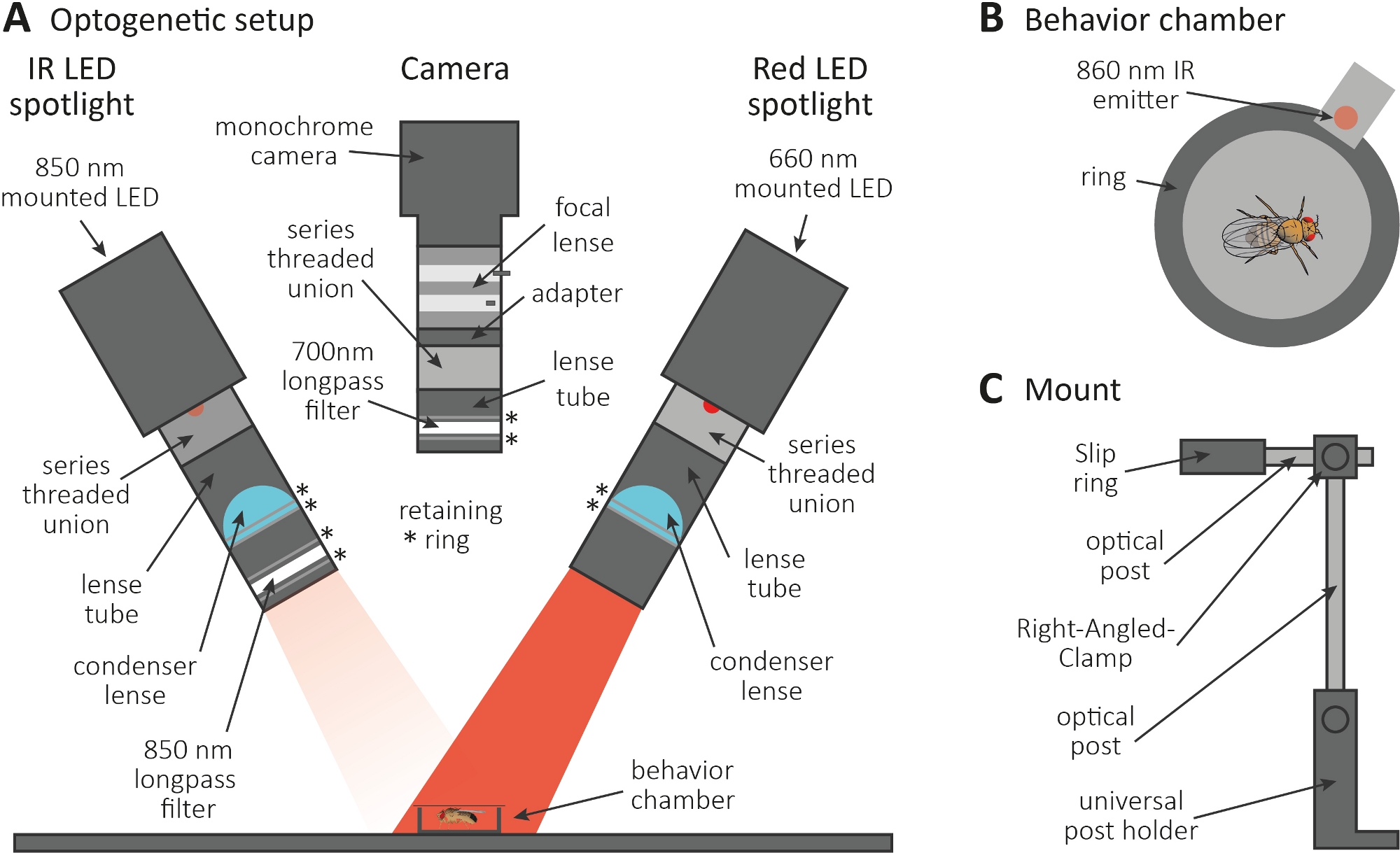
- 暗い部屋で光遺伝学的実験を行う。セットアップが暗闇の中でハエを照らす850 nm赤外線(IR)LEDスポットライト(図3A)、CsChrimsonを表現するニューロンを活性化する660 nmの赤色LEDスポットライト、および700 nmロングパスフィルタを備えたモノクロカメラで構成され、赤色の光が点滅するのを防ぎます。A
- 3Dプリンタを使用して、直径1cmの小さな円形の挙動チャンバーを生成し、カバースライドで覆い、チャンバーの隣にある赤色のLEDスポットライトに結合した860 nmのエミッタを配置します(図3B)。
注: エミッタは、赤い LED スポットライトがオンになっているときに、ニューロンをアクティブにするタイミングを示します。 - LEDスポットライトとカメラをチャンバーの上に取り付けます(図3A、C)。
- 10の間に10のHzの点滅によってニューロンを活性化する。活性化の期間は、実験計画に従って調整することができる。
- 光遺伝学のためのハエの準備
- 電子レンジで食べ物を溶かします。食品を冷却した後、固化する前に、エタノール(EtOH)のすべてのトランスレティナルを200μMの最終濃度に加え、すぐに空のバイアルに注ぎます。
注:ホットフードにすべての経常食を加えないように、これはあまり効率的な光遺伝学をもたらす可能性があります。 - プラグまたは綿のボールで固化食品を含むバイアルをカバーします。アルミホイルでバイアルを包みます。次に、食品含有バイアルを暗い冷たい部屋に保管します。
- 右の遺伝子型の5人の処女女性と5人の男性(図6A、世代P0)を使用してRTで十0字架を行います。毎日のベースで、差し込みたばかりの成人子孫(世代F1)を収集し、フライフードで200 μMすべてのトランスレチナルを含むアルミニウムで覆われたバイアルで7〜14日間老化させてください。
- ハエを食品を含むバイアルから、食べ物のない空のバイアルにタップして収集します。約30sの氷を含む水でバイアルを冷やすと、ハエは眠りに落ちるでしょう。個々のハエをカバースライドで覆われた小さなチャンバーに急速に入れます(図3B)。
注:ハエが暖かければすぐに目が覚めます。個々のハエをそれぞれ単一のチャンバーに素早く広げるのが重要です。CO2パッドを避けてハエを麻酔し、これは彼らの行動に影響を与えます。 - 光遺伝学を実行して、アンテナグルーミングを引き出します。ここで、プロトコルは、赤い光が存在しない30秒、10 Hzで10秒,の赤信号暴露が続く。この手順を合計で3回繰り返し、その後に赤い光が12、29、30,29の間隔を追加して30秒続く。
注: このプロトコルは、実験的な設定に従って調整できます。 - CO2パッド上の各チャンバーから個々のハエを収集します。触角損傷にさらします。左と右の2ndアンテナセグメントの両方をアスレートします(図2C)。これは、ジョンストンの器官(JO)ニューロンの細胞体を除去し、軸索突起はCNSに残る。200 μM全てのトランスレチナルを含むアルミニウムで覆われたバイアルでハエを回収します。
注:光遺伝学によって誘発されるアンテナグルーミングの場合、感覚ニューロン細胞体は第2のアンテナセグメントに収容される(図2C)。 - 対応する時点(例えば、7日間のポストアンテナアブレーション)で、被験者は別のグルーミングアッセイに飛ぶ(ステップ3.2.4に戻る)。
- 電子レンジで食べ物を溶かします。食品を冷却した後、固化する前に、エタノール(EtOH)のすべてのトランスレティナルを200μMの最終濃度に加え、すぐに空のバイアルに注ぎます。

図3:軸索およびシナプス機能の読み出しとしてグルーミングを誘導する光遺伝学的セットアップ。(A) 光遺伝学に必要な組み立て成分の図。赤外線(IR)LEDスポットライト、カメラおよび赤色LEDスポットライト(それぞれ左から右へ)。詳細な説明を含むコンポーネントは、一覧の一覧を参照してください。(B) IR エミッタを含む動作チャンバのトップビューの図は、赤色の LED スポットライトの活性化を示す。(C) 単一マウント設定の図。2 つの LED スポットライトとカメラにそれぞれ 3 台のマウント設定が必要です。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}
結果
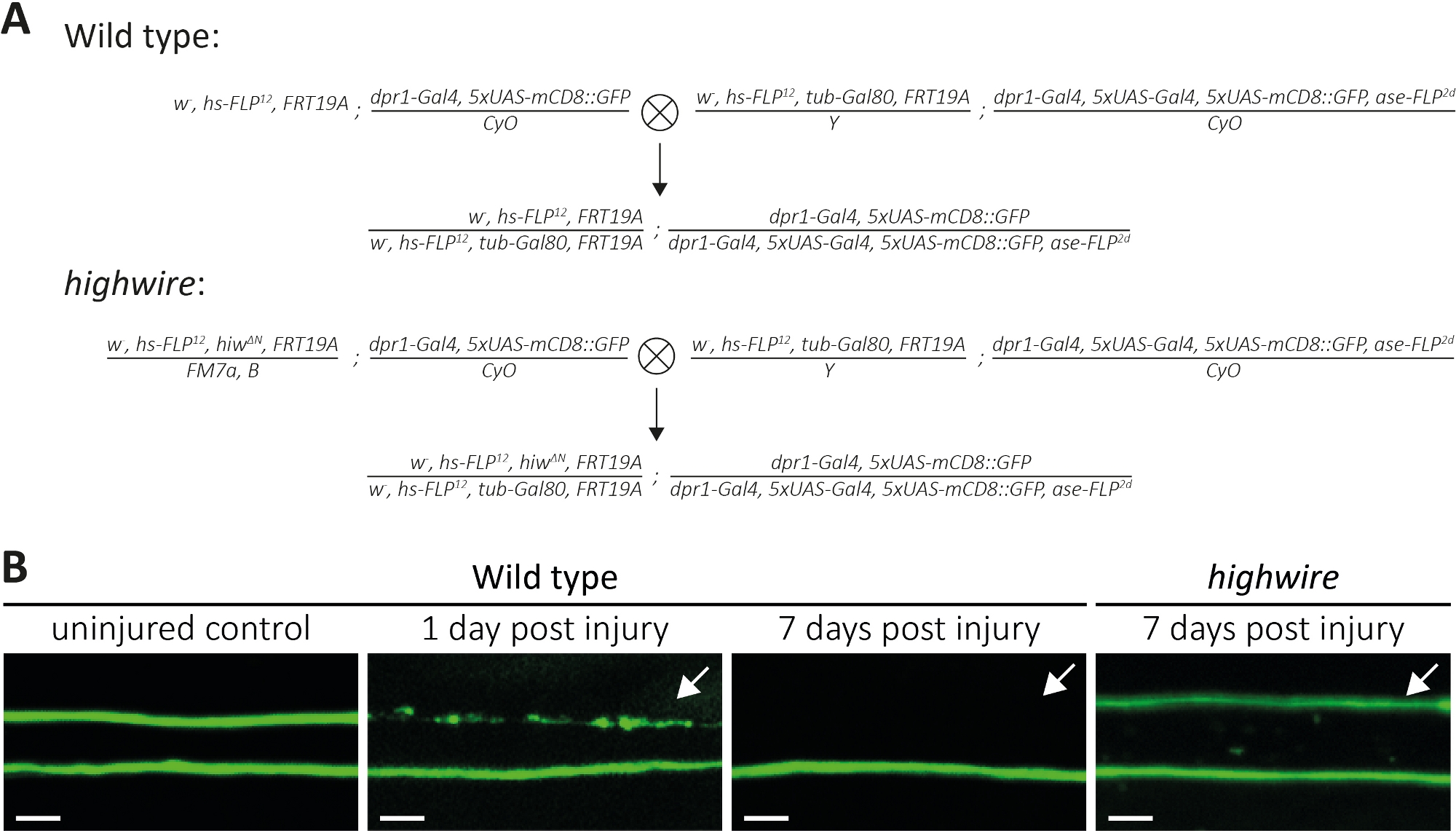
以上、切断された軸索とそのシナプスの形態と機能を研究する3つの方法について述べた。最初の方法はPNSの個々の軸索の高解像観察を可能にする。この場合、MARCM 技法14、31,によって生成されたクローンが必要です。ここでは、クロスを行って、野生型およびハイワイヤミュータントMARCMクローンを生成しました(図4A)。翼の中央に単純な切り傷は、遠位(例えば、翼の外側)に収容されたニューロンの軸索損傷を誘発し、近位ニューロン(例えば、切断部位と胸部の間)は損傷を受けていない。このアプローチにより、同じ神経束内の損傷のないコントロール軸索の軸索の軸索死を並べて観察することが可能になります(図1A、図4B)。ここでは、GFP標識クローンの数が少ない遺伝的背景を用いた(例えば、各実験14の2つ)。野生型軸索の傷害後1日および7日目の例を示し、制御軸索、軸索死を受けている軸索、および周囲のグリアによってクリアされる軸索断片の例をそれぞれ提供する。さらに、ハイワイヤー変異体で軸索損傷を繰り返し、怪我の7日後に結果を分析した。
無傷の制御翼は2つの野生型クローンを収容し、したがって2つのGFP標識された野生型軸索(図4B、野生型、無傷の制御)を収容する。マイクロハサミを使用して翼の真ん中を切断した1日後、細胞体が切断部位に遠位するGFP標識軸索では軸索死が誘発され、近位に収容された細胞体からの軸索は同じ神経束内の内部制御として機能する(図4B、野生型、1日後損傷)。矢印で示された上部の軸索破片の痕跡に注意してください。軸索損傷の7日後、GFP標識軸索は周囲のグリアによって取り除かれ、GFP標識された損傷していないコントロール軸索は神経束に残る(図4B、野生型、7日後の傷害、矢)。対照的に、7日間切断されたハイワイヤー変異軸索は形態学的に保存されたままであり、以前の知見11、14(図414 B、ハイワイヤー、傷害後7日、矢印)と一致する。Bこれらの結果は、ショウジョウバエ翼の強力な視覚的解像度を示す。軸索死は、同じ神経束中の損傷のないコントロールの並んで観察することができる。野生型軸索は傷害後1日以内に軸索死を受け、得られた破片は7日以内に取り除かれ、軸索死不全のハイワイヤー変異体は形態学的に7日間保存されたままである。

図4:翼におけるGFP標識感覚ニューロン軸索の軸索死を研究するアプローチ(A) スケマティッククロスは、翼内に野生型およびハイワイヤクローンを生成します(それぞれP0とF1生成)。ヴァージンの女性は左側、男性は右側にあります。遺伝子型の詳細については、材料表を参照してください。(B) 制御および負傷したGFP標識軸索の例。視野は(図1A)に示されています。左から:野生型コントロール軸索、野生型軸索1日後損傷、野生型軸索は傷害後7日、ハイワイヤー変異軸索は負傷後7日、それぞれ傷害後7日。矢印は、切断された軸索、スケールバー= 5 μmを示します。
{kind=link}
2番目の方法は、左右のアンテナに収容されたニューロンに属するシナプスを形成するCNSに投影される軸索束全体を視覚化する方法を説明する(図2A-C)。ここでは、クロスを行って、野生型およびハイワイヤミュータントMARCMクローンを生成しました(図5A)。怪我をしていない、GFPラベルの軸索とそのシナプスは、怪我がない場合、数日から数週間にわたって視覚化することができます(図5B、野生タイプ、無傷のコントロール)。あるいは、動物は3番目のアンテナセグメントアブレーションを受けることができ、切断されたGFP標識軸索とそのシナプスは、時間から数日にわたる時間経過中に観察することができる。この時点で軸索とそのシナプスが軸索死を受け、結果として生じる破片がグリア周囲によって取り除かれたため、アンテナアブレーション後7日間に焦点を当てました。右のアンテナの一方的なアブレーションが行われる場合、右軸索束が切断され、分解され、結果として生じる破片は、傷害の7日後(図5B、野生型、一方的なアブレーション、7日間の傷害後、矢)、以前の知見と一致する。あるいは、右と左のアンテナの両方をアブラプし、両方の軸索束を切断することができ、怪我の7日後、軸索とそのシナプスが消失した(図5B、野生型、両側アブレーション、7日後の怪我、矢)。対照的に、ハイワイヤー変異体における右のアンテナの一方的なアブレーションは、損傷後7日間保存された切断軸索をもたらし、以前の知見11、14(図511B、14ハイワイヤー、一方的なアブレーション、7日後の傷害、矢)と一致する。Bこれらの結果は、切断された野生型軸索が軸索死を受け、結果として生じる破片が7日以内に取り除かれ、軸索死不十分なハイワイヤー突然変異体が軸索死を受けず、形態学的に7日間保存されることを示している。

図5:脳内のGFP標識感覚ニューロン軸索の軸索死を研究するアプローチ(A) スケマティッククロスは、脳内で野生型およびハイワイヤークローンを生成する(P0とF1生成、それぞれ)。ヴァージンの女性は左側、男性は右側にあります。遺伝子型の詳細については、材料表を参照してください。(B) 制御および負傷したGFP標識軸索の例。左から:野生型コントロールの傷つかず、野生型7日後の一方的なアンテナアブレーション、野生型7日後の両側アンテナアブレーション、ハイワイヤー変異体は、それぞれ一方的なアンテナアブレーションの後7日。矢印は、切断された軸索バンドルを示し、スケールバー= 10 μm。この図のより大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}
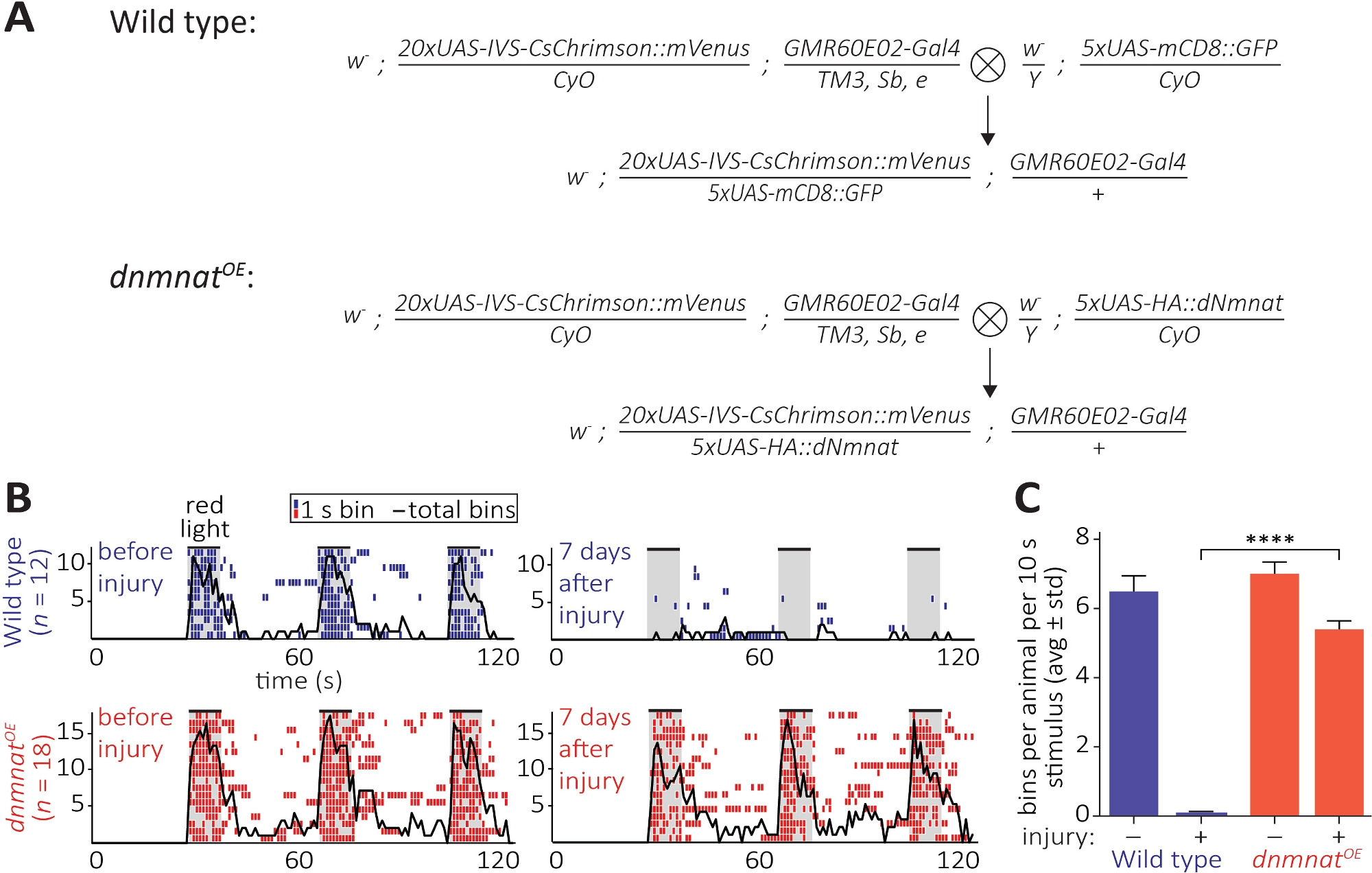
第3の方法はCNSの切断された軸索およびそれらのシナプスの機能保存の観察を可能にする。これは、アンテナグルーミングを誘導するのに十分な2ndアンテナセグメントに収容されたJOニューロンのサブセットの操作に依存しています。JOニューロンにおける赤シフトチャネルロドプシン(CsChrimson)の発現は、全てのトランスレチナルの栄養補給と組み合わされ、赤光暴露時のシナプス後のグルーミング挙動を12,30,30に単純に惹き起こすのに十分である。ここでは、交配を行って野生型JOニューロンを生成し、JOニューロンは過剰発現するdnmnat(dnmnat dnmnatOE)(図6A)を生成した。アキソン死(dnmnatOE)を有するJOニューロンを含む野生型ハエまたはハエは、いずれも傷害前に強力なグルーミング行動を有する。しかし、傷害後7日(例えば、第2のアンテナセグメントの両側アブレーション)、グルーミングは、傷害誘発軸索およびシナプス変性のために野生型ハエの光遺伝学によって引き出されないが、アクソン死が減少している動物はグルーミングを続けている(図6B、映画1、2)。したがって、減衰した軸索死は、切断された軸索とそのシナプスを7日間機能的に保存することができる。

図6:軸索とシナプス機能をアキソトミーの後に可視化するアプローチ(A)スケマティッククロスは、野生型を生成し、Dnmnat過剰発現JO感覚ニューロン(P0とF1生成、それぞれ)。ヴァージンの女性は左側、男性は右側にあります。遺伝子型の詳細については、材料表を参照してください。(B) 光遺伝学によって誘導されるグルーミング行動の個別のエトグラム。トップ:野生型の個々のエチグラムは、怪我の前と7日後に飛ぶ(青)。下:ハエの個々のエトグラムは、傷害の前および7日後のJOニューロンにおける過剰発現するドミンナット(dnmnatOE)(赤)である。各ビンは、1 s 内の少なくとも 1 つのグルーミング動作を示します。黒い線は、すべてのビンの合計を示します。(C) グルーミング動作の定量化。データは平均±標準偏差、p > 0.001 (一方向の分散分析、Tukeyの事後検定との多重比較) として示されます。この図の大きなバージョンを表示するには、ここをクリックしてください。
{kind=link}
動画1:アンテナアブレーションの前後7日前の光遺伝学によって引き出された野生型のグルーミング行動を代表する。こちらをクリックして、このビデオをダウンロードしてください。
動画2:触角切り前および7日前にJOニューロンの経皮を過剰発現するハエの光遺伝学によって引き起こされる代表的なグルーミング挙動。こちらをクリックして、このビデオをダウンロードしてください。
ディスカッション
ここで説明するプロトコルは、ショウジョウバエの細胞体から分離された軸索とそのシナプスの機能だけでなく、形態の堅牢で再現可能な観察を可能にする。翼アッセイは、PNS14における損傷のないコントロール軸索の軸索の軸索死亡の横並びの観察を容易にし、触理アッセイはGFP標識軸索とそのシナプスの神経束全体の観察を容易にし、脳内の形態および機能の両方を評価する(CNS)12。実験を設計する際に考慮する必要がある形態学を研究するための各アプローチには、重要なステップと特定の利点があります。
翼のPNSにおける軸索形態を観察するために、実験は、翼の透明性のために容易に行うことができる:それは解剖および免疫組織化学をバイパスすることを可能にする。しかし、固定の欠如のために、翼は14を取り付けた直後にイメージ化する必要があります。現在、2つの異なるGal4ドライバが頻繁に使用されています, ok371Gal4またはdpr1Gal4のいずれかであり、両方の参照は、変性14、26,26を定量化するための明確なアプローチを提供しています。軸索形態の分解能は前例がないので、「抑制可能な細胞マーカーを用いたモザイク解析(MARCM)」14,31を用いることで、いくつかのニューロンのまばらな標識が推奨される。14,31対照的に、シナプスの観察は翼では不可能であり、ハエの胸郭の内側の腹側神経コードに位置する。さらに、追加の軸索マーカーは免疫組織化学では視覚化できません:ワックス状のキューティクルは、下層組織への固定剤および抗体の拡散を不可能にします。
CNSで軸索とシナプスの形態を観察するには、脳の切除を行う必要があります。それらは、免疫神学的化学の使用によって追加の軸索およびシナプスマーカーを視覚化する利点を提供し、かつシナプスは、同じ視野10、13,13の軸索と並んで観察することができる。特徴付けられた嗅覚受容体ニューロン(ORN)Gal4ドライバの大規模なコレクションは、容易に利用できる32、そして頻繁に、OR22aGal4が選択のドライバである。アンテナアブレーションの場合、OR22aニューロンの細胞体は3番目のセグメントに収容されている(図2B)。蛍光強度ベースの定量化は、軸索またはシナプス13の変性を定量するために使用される。逆に、脳の解剖や抗体染色による実験時間が必要です。
奇数術後の軸索およびシナプス機能を可視化するために、光遺伝学は、触角グルーミングを引き起こすために使用される:それは、切断された軸索とそのシナプス12の機能的保存のための読み出しとして機能する。グルーミング回路と対応する感覚、インター・アンド・モーターニューロンGal4ドライバは、29,30,30を徹底的に記述されている。GMR60E02Gal4はジョンストンの器官(JO)感覚ニューロンのサブセットにラベルを付け、グルーミング29,30,30に十分である。アンテナアブレーションの場合、JOニューロンの細胞体は第2のアンテナセグメントに収容される(図2B)。光遺伝学的なセットアップは、容易にゼロから構築することができ、または既存のセットアップを調整することができます。重要なのは、暗い部屋で実験を行う必要があり、赤外線(IR)LEDスポットライトで視覚化されたハエです。CsChrimsonをチャネルとして使用する場合、JOニューロン29を活性化するために、すべての経常在性と赤色LEDスポットライトを食品に供給することが重要です。あるいは、青色光感受性チャンネルおよび青色LEDスポットライト、またはTrpA1チャネルおよび温度をニューロン活性化29、33,33に使用することができる。グルーミング動作の定量化は、すでに12,29,29に記載されています。
これらのアッセイを使用して軸索死を具体的に研究する場合、形態学的または機能的保存の表現型は時間の経過とともに堅牢でなければならないことに注意することが重要です。軸索死が形態学的保存34,35において一貫した、あまり顕著でない表現型をもたらす場合があり、35そのような表現型が機能的保存に変換されるかどうかはまだ決定されていない。
神経が負傷した11,23,23ではなく神経が押しつぶされたショウジョウバエ幼虫の発達中に神経細胞にも軸索死の型が認められている。ここでは、特に開発を完了した成体ショウジョウバエニューロンに焦点を当てた。この文脈では、RNA干渉36、または組織特異的なCRISPR/Cas937の使用を容易に実施することができる。重要なことに、上記の技術は、軸索死独立した文脈で使用することができる:彼らは神経学的維持因子38の特徴付けを促進し、軸索輸送39、加齢に依存する軸索ミトコンドリア変化40、および軸索ミトコンドリア41の形態を有する。
開示事項
著者らは、開示するものは何もないと宣言している。
謝辞
ノイコム研究所全体の貢献に感謝します。この研究は、スイス国立科学財団(SNSF)助教授賞(助成176855)、パラプレジア国際研究財団(IRP、助成金P180)、SNSFスパーク(グラント190919)、ローザンヌ大学の支援によって支援されました。LJNに基礎神経科学科(エタット・ド・ヴォー)
資料
| Name | Company | Catalog Number | Comments |
| Tweezers (high precision, ultra fine) | EMS | 78520-5 | Antennal ablation |
| MicroPoint Scissors (5-mm cutting edge) | EMS | 72933-04 | Wing injury |
| 1.5 mL microcentrifuge tube | Eppendorf | 30120086.0000 | |
| 35mm tissue culture dish | Sarstedt | 83.3900 | |
| Cover Slips, Thickness 1 | Thermo Scientific™ | BB02400600A113MNT0 | |
| Superfrost Microscope Slides | Thermo Scientific™ | AA00008032E00MNT10 | |
| High-Sensitivity USB 2.0 CMOS Camera, 1280 x 1024, Global Shutter | Thorlabs | DCC1240M | Camera setup |
| SM1 Retaining Ring for Ø1" Lens Tubes and Mounts | Thorlabs | SM1RR | |
| 25mm 1/1.2" C mount Lens | Tamron | M112FM25 | |
| Adapter with External M27 x 0.5 Threads and Internal SM1 Threads | Thorlabs | SM1A36 | |
| Aspheric Condenser Lens, Ø25 mm, f=20.1 mm, NA=0.60, ARC: 650-1050 nm | Thorlabs | ACL2520U-B | |
| Ø25.0 mm Premium Longpass Filter, Cut-On Wavelength: 700 nm | Thorlabs | FELH0700 | |
| SM1 (1.035"-40) Coupler, External Threads, 0.5" Long | Thorlabs | SM1T2 | |
| SM1 Lens Tube Without External Threads, 1" Long, Two Retaining Rings Included | Thorlabs | SM1M10 | |
| 850 nm, 900 mW (Min) Mounted LED, 1200 mA | Thorlabs | M850L3 | IR LED spotlight |
| SM1 (1.035"-40) Coupler, External Threads, 0.5" Long | Thorlabs | SM1T2 | |
| SM1 Lens Tube Without External Threads, 2" Long, Two Retaining Rings Included | Thorlabs | SM1M20 | |
| Aspheric Condenser Lens, Ø25 mm, f=20.1 mm, NA=0.60, ARC: 650-1050 nm | Thorlabs | ACL2520U-B | |
| Ø25.0 mm Premium Longpass Filter, Cut-On Wavelength: 850 nm | Thorlabs | FELH0850 | |
| SM1 Retaining Ring for Ø1" Lens Tubes and Mounts | Thorlabs | SM1RR | |
| 660 nm, 940 mW (Min) Mounted LED, 1200 mA | Thorlabs | M660L4 | Red LED spotlight |
| Aspheric Condenser Lens, Ø25 mm, f=20.1 mm, NA=0.60, ARC: 650-1050 nm | Thorlabs | ACL2520U-B | |
| SM1 (1.035"-40) Coupler, External Threads, 0.5" Long | Thorlabs | SM1T2 | |
| SM1 Lens Tube Without External Threads, 2" Long, Two Retaining Rings Included | Thorlabs | SM1M20 | |
| 15 V, 2.4 A Power Supply Unit with 3.5 mm Jack Connector for One K- or T-Cube | Thorlabs | KPS101 | LED control |
| T-Cube LED Driver, 1200 mA Max Drive Current | Thorlabs | LEDD1B | |
| 150 mm x 300 mm x 12.7 mm Aluminum Breadboard, M6 Double-Density Taps | Thorlabs | MB1530/M | Mount base |
| Ø12.7 mm Universal Post Holder, Spring-Loaded Locking Thumbscrew, L = 75 mm | Thorlabs | UPH75/M | Mount, 3x (IR LED, red LED, cam) |
| Ø1.20" Slip Ring for SM1 Lens Tubes and C-Mount Extension Tubes, M4 Tap | Thorlabs | SM1RC/M | |
| Ø12.7 mm Optical Post, SS, M4 Setscrew, M6 Tap, L = 150 mm | Thorlabs | TR150/M | |
| Ø12.7 mm Optical Post, SS, M4 Setscrew, M6 Tap, L = 40 mm | Thorlabs | TR40/M | |
| Right-Angle Clamp for Ø1/2" Posts, 5 mm Hex | Thorlabs | RA90/M | |
| M6 x 1.0 Stainless Steel Cap Screw, 16 mm Long, Pack of 25 | Thorlabs | SH6MS16 | screws for mount onto base |
| USB-6001 14-Bit 20 kS/s Multifunction I/O and NI-DAQmx | National Instruments | 782604-01 | Red LED spotlight controller |
| 20k Ohm 1 Gang Linear Panel Mount Potentiometer | TT Electronics/BI | P230-2EC22BR20K | fintuner for indicator |
| IR (860nm) emitter, 100 mA radial | Osram | 475-1365-ND | Red light indicator |
| cable | - | - | Misc |
| All-trans retinal | Sigma | R2625 | |
| Ethanol absolute | Vwr | 20821.296 | |
| Halocarbon Oil 27 | Sigma | H8773 | |
| Mowiol | Merk | 81381 | |
| Paraformaldehyde | Sigma | F8775 | |
| Phosphate buffered saline (PBS) | Sigma | P5493 | |
| Sylgard 184 silicone elastomer base | Dow Corning Corp | 4019862 | |
| Sylgard 184 silicone elastomer curing agent | Dow Corning Corp | 4019862 | |
| Triton X-100 | Sigma | T8787 | |
| Chicken anti-GFP antibodies | Rockland | 600-901-215 | Antibodies |
| Goat Dylight anti-Chicken | Abcam | ab96947 | |
| FM7a, B | BDSC | RRID:BDSC_785 | X chromosome |
| FRT19A[hs-neo] | BDSC | RRID:BDSC_1709 | |
| hiw[ΔN] | BDSC | RRID:BDSC_51637 | |
| hs-FLP[12] | BDSC | RRID:BDSC_1929 | |
| tub-Gal80[LL1] | BDSC | RRID:BDSC_5132 | |
| w[1118] | BDSC | RRID:BDSC_3605 | |
| 20xUAS-IVS-CsChrimson::mVenus | BDSC | RRID:BDSC_55135 | 2nd chromosome |
| 5xUAS-Gal4[12B] | Kyoto | RRID:Kyoto_108492 | |
| 5xUAS-HA::dnmnat | BDSC | RRID:BDSC_39702 | |
| 5xUAS-mCD8::GFP[LL5] | BDSC | RRID:BDSC_5134 | |
| ase-FLP[2d] | Freeman laboratory | Neukomm et al., 2014 (PNAS) | |
| CyO | BDSC | RRID:BDSC_2555 | |
| dpr1-Gal4 | BDSC | RRID:BDSC_25083 | |
| OR22a-Gal4 | BDSC | RRID:BDSC_9952 | |
| ey-FLP[6] | BDSC | RRID:BDSC_5577 | 3rd chromosome |
| GMR60E02-Gal4 | BDSC | RRID:BDSC_39250 | |
| TM3,Sb,e | BDSC | RRID:BDSC_3644 |
参考文献
- Matsuda, W., et al. Single Nigrostriatal Dopaminergic Neurons Form Widely Spread and Highly Dense Axonal Arborizations in the Neostriatum. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience. 29 (2), 444-453 (2009).
- Wedel, M. J. A Monument of Inefficiency: The Presumed Course of the Recurrent Laryngeal Nerve in Sauropod Dinosaurs. Acta Palaeontologica Polonica. 57 (2), 251-256 (2012).
- Mariano, V., Domínguez-Iturza, N., Neukomm, L. J., Bagni, C. Maintenance mechanisms of circuit-integrated axons. Current Opinion in Neurobiology. 53, 162-173 (2018).
- Conforti, L., Gilley, J., Coleman, M. P. Wallerian degeneration: an emerging axon death pathway linking injury and disease. Nature reviews Neuroscience. 15 (6), 394-409 (2014).
- Neukomm, L. J., Freeman, M. R. Diverse cellular and molecular modes of axon degeneration. Trends in Cell Biology. 24 (9), 515-523 (2014).
- Gustavsson, A., et al. Cost of disorders of the brain in Europe 2010. European Neuropsychopharmacology: The Journal of the European College of Neuropsychopharmacology. 21 (10), 718-779 (2011).
- Waller, A. Experiments on the Section of the Glossopharyngeal and Hypoglossal Nerves of the Frog, and Observations of the Alterations Produced Thereby in the Structure of Their Primitive Fibres. Philosophical Transactions of the Royal Society of London. 140, 423-429 (1850).
- Rosell, A. L., Neukomm, L. J. Axon death signalling in Wallerian degeneration among species and in disease. Open Biology. 9 (8), 190118(2019).
- Mack, T. G., et al. Wallerian degeneration of injured axons and synapses is delayed by a Ube4b/Nmnat chimeric gene. Nature Neuroscience. 4 (12), 1199-1206 (2001).
- Osterloh, J. M., et al. dSarm/Sarm1 is required for activation of an injury-induced axon death pathway. Science. 337 (6093), New York, NY. 481-484 (2012).
- Xiong, X., et al. The highwire ubiquitin ligase promotes axonal degeneration by tuning levels of nmnat protein. PLoS Biology. 10 (12), 1001440(2012).
- Neukomm, L. J., et al. Axon Death Pathways Converge on Axundead to Promote Functional and Structural Axon Disassembly. Neuron. 95 (1), 78-91 (2017).
- MacDonald, J. M., et al. The Drosophila cell corpse engulfment receptor Draper mediates glial clearance of severed axons. Neuron. 50 (6), 869-881 (2006).
- Neukomm, L. J., Burdett, T. C., Gonzalez, M. A., Zuchner, S., Freeman, M. R. Rapid in vivo forward genetic approach for identifying axon death genes in Drosophila. Proceedings of the National Academy of Sciences of the United States of America. 111 (27), 9965-9970 (2014).
- Lu, T. Y., et al. Axon degeneration induces glial responses through Draper-TRAF4-JNK signalling. Nature Communications. 8, 14355(2017).
- Lunn, E. R., Perry, V. H., Brown, M. C., Rosen, H., Gordon, S. Absence of Wallerian Degeneration does not Hinder Regeneration in Peripheral Nerve. The European Journal of Neuroscience. 1 (1), 27-33 (1989).
- Adalbert, R., et al. A rat model of slow Wallerian degeneration (Wld(S)) with improved preservation of neuromuscular synapses. The European Journal of Neuroscience. 21 (1), 271-277 (2005).
- Martin, S. M., O'Brien, G. S., Portera-Cailliau, C., Sagasti, A. Wallerian degeneration of zebrafish trigeminal axons in the skin is required for regeneration and developmental pruning. Development. 137 (23), Cambridge, England. 3985-3994 (2010).
- Feng, Y., et al. Overexpression of Wld(S) or Nmnat2 in Mauthner Cells by Single-Cell Electroporation Delays Axon Degeneration in Live Zebrafish. Journal of Neuroscience Research. 88 (15), 3319-3327 (2010).
- Gilley, J., Coleman, M. P. Endogenous Nmnat2 is an essential survival factor for maintenance of healthy axons. PLoS Biology. 8 (1), 1000300(2010).
- Babetto, E., Beirowski, B., Russler, E. V., Milbrandt, J., DiAntonio, A. The Phr1 ubiquitin ligase promotes injury-induced axon self-destruction. Cell Reports. 3 (5), 1422-1429 (2013).
- Gerdts, J., Summers, D. W., Sasaki, Y., DiAntonio, A., Milbrandt, J. Sarm1-mediated axon degeneration requires both SAM and TIR interactions. The Journal of Neuroscience. 33 (33), 13569-13580 (2013).
- Gerdts, J., Brace, E. J., Sasaki, Y., DiAntonio, A., Milbrandt, J. SARM1 activation triggers axon degeneration locally via NAD+ destruction. Science. 348 (6233), New York, NY. 453-457 (2015).
- Bridge, P. M., et al. Nerve crush injuries--a model for axonotmesis. Experimental Neurology. 127 (2), 284-290 (1994).
- Maxwell, W. L., Bartlett, E., Morgan, H. Wallerian Degeneration in the Optic Nerve Stretch-Injury Model of Traumatic Brain Injury: A Stereological Analysis. Journal of Neurotrauma. 32 (11), 780-790 (2015).
- Fang, Y., Soares, L., Teng, X., Geary, M., Bonini, N. M. A novel Drosophila model of nerve injury reveals an essential role of Nmnat in maintaining axonal integrity. Current Biology. 22 (7), 590-595 (2012).
- Janelia Farm Adult Drosophila Brain Dissection. , Available from: https://www.janelia.org/project-team/flylight/protocols (2015).
- Cold Spring Harbor. Mowiol mounting medium. Cold Spring Harbor Protocols. 2006 (1), (2006).
- Seeds, A. M., et al. A suppression hierarchy among competing motor programs drives sequential grooming in Drosophila. eLife. 3, 02951(2014).
- Hampel, S., Franconville, R., Simpson, J. H., Seeds, A. M. A neural command circuit for grooming movement control. eLife. 4, 08758(2015).
- Lee, T., Luo, L. Mosaic analysis with a repressible cell marker for studies of gene function in neuronal morphogenesis. Neuron. 22 (3), 451-461 (1999).
- Vosshall, L. B., Wong, A. M., Axel, R. An olfactory sensory map in the fly brain. Cell. 102 (2), 147-159 (2000).
- Hampel, S., McKellar, C. E., Simpson, J. H., Seeds, A. M. Simultaneous activation of parallel sensory pathways promotes a grooming sequence in Drosophila. eLife. 6, (2017).
- Farley, J. E., et al. Transcription factor Pebbled/RREB1 regulates injury-induced axon degeneration. Proceedings of the National Academy of Sciences of the United States of America. 23 (6), (2018).
- Wang, H., et al. Rapid depletion of ESCRT protein Vps4 underlies injury-induced autophagic impediment and Wallerian degeneration. Science Advances. 5 (2), 4971(2019).
- Dietzl, G., et al. A genome-wide transgenic RNAi library for conditional gene inactivation in Drosophila. Nature. 448 (7150), 151-156 (2007).
- Port, F., et al. A large-scale resource for tissue-specific CRISPR mutagenesis in Drosophila. bioRxiv. 102, 636076(2019).
- Vagnoni, A., Hoffmann, P. C., Bullock, S. L. Reducing Lissencephaly-1 levels augments mitochondrial transport and has a protective effect in adult Drosophila neurons. Journal of Cell Science. 129 (1), 178-190 (2016).
- Vagnoni, A., Bullock, S. L. A cAMP/PKA/Kinesin-1 Axis Promotes the Axonal Transport of Mitochondria in Aging Drosophila Neurons. Current Biology. 28 (8), 1265-1272 (2018).
- Cao, X., et al. In vivo imaging reveals mitophagy independence in the maintenance of axonal mitochondria during normal aging. Aging Cell. 16 (5), 1180-1190 (2017).
- Smith, G. A., et al. Glutathione S-Transferase Regulates Mitochondrial Populations in Axons through Increased Glutathione Oxidation. Neuron. 103 (1), 52-65 (2019).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved