Method Article
全反射蛍光(TIRF)顕微鏡を用いた脂質二重膜の横方向移動度とイオンチャネル活性の1分子イメージング(英語)
要約
このプロトコルでは、TIRF顕微鏡を使用して個々のイオンチャネルを追跡し、支持された脂質膜におけるそれらの活性を決定し、それによって側方膜の動きとチャネル機能の間の相互作用を定義する方法について説明します。メンブレンの準備、データの記録、および結果の分析方法について説明します。
要約
高解像度イメージング技術により、多くのイオンチャネルは静的ではなく、細孔形成と補助サブユニットの一時的な会合、横方向拡散、他のタンパク質とのクラスタリングなど、非常に動的なプロセスを受けることが示されています。しかし、横方向の拡散と機能の関係はよくわかっていません。この問題にアプローチするために、全反射蛍光(TIRF)顕微鏡を使用して、支持された脂質膜内の個々のチャネルの横方向の移動度と活性をどのように監視および相関させることができるかについて説明します。膜は、液滴界面二重層(DIB)技術を使用して極薄ヒドロゲル基板上に製造されます。他のタイプのモデルメンブレンと比較して、これらのメンブレンは機械的に堅牢であり、高感度の分析技術に適しているという利点があります。このプロトコルは、膜に近接したCa2+感受性色素の蛍光発光を観察することにより、単一チャネルを通るCa2+イオンフラックスを測定します。従来の単一分子追跡アプローチとは対照的に、膜内のラテラルムーブメントと機能を妨げる可能性のある蛍光融合タンパク質または標識は必要ありません。タンパク質の立体構造変化に関連するイオンフラックスの変化の可能性は、膜内のタンパク質の横方向の動きにのみ起因します。代表的な結果は、ミトコンドリアタンパク質転座チャネルTOM−CCおよび細菌チャネルOmpFを用いて示される。OmpFとは対照的に、TOM-CCのゲーティングは分子の閉じ込めと横方向拡散の性質に非常に敏感です。したがって、支持された液滴界面二重層は、横方向の拡散とイオンチャネルの機能との間のリンクを特徴付けるための強力なツールです。
概要
本プロトコルは、ポリマー支持液滴界面二重層(DIB)膜における膜タンパク質の膜移動度とイオンチャネル透過性との間の相関を研究する方法を説明することを目的とする1,2,3。
この技術は、単一粒子追跡4,5、蛍光相関分光法6,7、高速原子間力顕微鏡8,9,10などの印象的な一連の高度な光学および表面分析ツールを補完します。これらは、膜ベースの反応に影響を与える膜の動的な組成と構造に関する貴重な洞察を提供します11,12,13。タンパク質の移動と横方向の拡散は膜内のタンパク質の局所密度に依存しますが、個々のタンパク質分子は脂質ラフト14およびタンパク質間相互作用15,16によってもトラップできます。膜から細胞外環境または細胞質ゾルに突き出ているタンパク質ドメインに応じて、タンパク質の移動性は、高度に可動性から完全に不動性まで変化し得る。しかしながら、膜およびその周辺構造の複雑さのために、側方移動性の性質とタンパク質機能との間の相互作用を解読することはしばしば困難である17。
DIB膜は、膜タンパク質の生物物理学的単一分子分析のための効率的なプラットフォームであることが証明されています18,19,20,21,22。それらは、脂質/油相中の水性液滴とヒドロゲル担持基質との接触による脂質自己組織化によって形成されます。一般的に使用される支持脂質二重層(SLB)1、23、24、25と同様に、DIBは、適切なリガンドで官能化された場合、ポリマーマトリックスへのタンパク質の一時的または恒久的な結合による横方向の移動性の局所的な調節を可能にする17。後者は、不均一なタンパク質分布を有する細胞膜における生化学的プロセスのモデルシステムとして役立ち得る10。
ここで説明する実験的アプローチは、Tirf顕微鏡を用いてメンブレン2,22に近接した個々のチャネルを通るCa2+イオンフラックスを測定するために、Ca2+感受性色素の蛍光に依存する。この光学的アプローチは、サンプルの照明を膜に近い距離に制限し、エバネッセント励起光の物理的特性により、蛍光バックグラウンドの大幅な減少につながります。後者は、単一分子の検出に高い空間分解能と時間分解能が必要な場合の前提条件です。古典的な電気生理学的方法26,27とは対照的に、個々のチャネルを通るイオンフラックスを研究するために膜電圧は必要ありません。さらに、この方法は、膜中のチャネルの側方移動を妨げる可能性のある蛍光色素または分子による標識を必要としない。
この方法は、古典的な電気生理学を使用せずに、膜に埋め込まれたタンパク質チャネルを単一分子レベルで研究する場合に特に有用です。Neurospora crassa28,29,30のミトコンドリアタンパク質伝導チャネルTOM-CCと、大腸菌17,31の外膜を横切る低分子の親水性分子の拡散をサポートするOmpFを使用して、2つのタンパク質の膜移動度とチャネル活性を研究し、相関させる方法を示します。このアプローチは、TOM-CCおよびOmpFに最適化されていますが、他のタンパク質チャネルにも容易に適用できることを示唆しています。
プロトコル
1.タンパク質生産
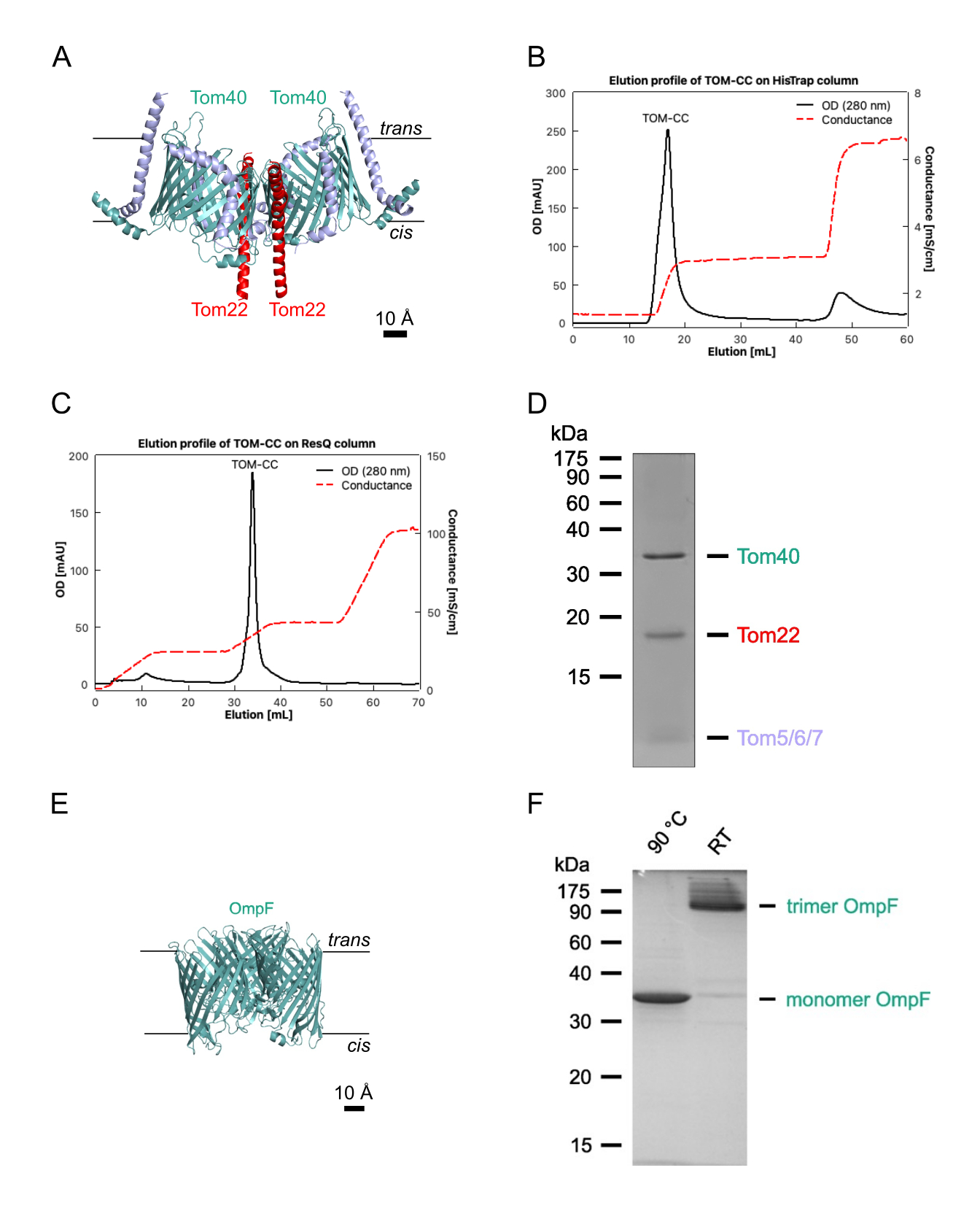
注:このセクションでは、LamBとOmpCを欠く大腸菌BE BL21(DE3)omp631,32からOmpFを分離する手順と、ニューロスポラクラッサからTOMコア複合体を分離する手順について説明します(図1)28,29。後者は、TOMサブユニットTom22の6xHis標識形態を含むN.クラッサ株28から単離されたミトコンドリアを必要とし(図1A)、これは28に記載のように単離することができる。以下のプロトコルは、通常、1〜2 mgのN.クラッサTOM-CCおよび10 mgの大腸菌OmpFを生成します。量を調整する場合は、タンパク質/界面活性剤の比率を正確に維持することが重要です。特に指定のない限り、すべてのステップは4°Cで実行する必要があります。
- TOMコア複合体の単離
- Bauseweinら30によると、約1.5 kg(湿重量)菌糸からの差動沈降によって得られた精製 N.クラッサ ミトコンドリア(2 gのタンパク質)を、20 mM Tris-HCl(pH 8.5)、1%(w/v)DDM、20%(v/v)グリセロール、300 mM NaCl、20 mMイミダゾール、および1 mMフェニルメチルスルホニルフルオリド(PMSF)中のタンパク質濃度10 mg/mLで4°Cで30分間可溶化します。
注意:PMSFは有毒です。適切な個人用保護具を着用してください。 - 可溶化ミトコンドリアを超遠心管に移し、130,000 x g で4°Cで40分間超遠心し、標準グレードのろ紙でろ過することにより、可溶化膜タンパク質から非可溶化膜を分離します。
- 自動タンパク質精製システムを使用して、充填済みのNi-NTAカラム(容量5 mL)を約5カラム容量(CV)のバッファーA1(表1)で平衡化します。Ni-NTAカラムは、すべてのステップで1 mL/minの一定流量で稼働させます。タンパク質が2 g未満のミトコンドリアから単離されている場合は、低容量カラム(1 mL)を使用してください。
- 可溶化タンパク質サンプルを Ni-NTA カラムに 1 mL/分の流速でロードします。
- Ni-NTAカラムを5 CVのバッファーA1(表1)で洗浄し、結合していないタンパク質を除去します。
- Hisタグ付きTOM-CCを30%バッファーA2で溶出し(表1;300 mMイミダゾール)、280 nmクロマトグラムに現れるタンパク質ピークを収集します(図1B)。
- TOM-CCをさらに精製するには、自動タンパク質精製システムを使用して、充填済みの陰イオン交換カラム(1 mL)をバッファーB1、B2、およびB1(表1)のそれぞれ5 CVで平衡化します。陰イオン交換カラムは、すべてのステップで1 mL/minの一定流量で運転します。
- TOM-CCピーク画分(ステップ1.1.6)を陰イオン交換カラム(流速1 mL/分)にロードします。
- 5 CVのバッファーB1(表1)および0%〜20%バッファーB2の線形塩勾配(表1)でカラムを洗浄することにより、未結合タンパク質を除去します。
- 20%-35%バッファーB2の線形塩勾配でTOM-CCを溶出し、280 nmクロマトグラムに現れるタンパク質ピーク画分を収集します(図1C)。
- SDS-PAGE(図1D)でサンプルの純度を評価し、メーカーのプロトコルに従って市販のタンパク質アッセイを使用してタンパク質濃度を決定します( 材料表を参照)。
- タンパク質サンプルを液体窒素で凍結し、さらに使用するまで-80°Cで保存します。
注意: 液体窒素は換気の良い場所で取り扱う必要があります。適切な個人用保護具を着用してください。
- Bauseweinら30によると、約1.5 kg(湿重量)菌糸からの差動沈降によって得られた精製 N.クラッサ ミトコンドリア(2 gのタンパク質)を、20 mM Tris-HCl(pH 8.5)、1%(w/v)DDM、20%(v/v)グリセロール、300 mM NaCl、20 mMイミダゾール、および1 mMフェニルメチルスルホニルフルオリド(PMSF)中のタンパク質濃度10 mg/mLで4°Cで30分間可溶化します。
- OmpFの分離
- LamBおよびOmpC32を欠く大腸菌株BE BL21(DE3) omp6を、無菌条件下で凍結グリセロールストックから回収し、Luria-Bertani(LB)寒天プレートにストリークします(表2)。これを行うには、サンプルをプレート全体に均等に徐々に広げます。
- サンプルを寒天に5分間浸した後、蓋を閉めた状態でプレートを反転させ、37°Cで一晩インキュベートします。
- 単一の 大腸菌 コロニーを選択し、滅菌爪楊枝を使用してこの単一コロニーに7.5 mLのLB培地(表2)を接種し、37°Cで攪拌(170 rpm)しながら一晩(14時間)増殖させます。
- 滅菌ピペットで2 x 1 mLの 大腸菌 細胞を2 x 500 mLのLB培地に移し(表2)、振とう機で攪拌(~170 rpm)しながら37°Cで一晩(~14時間)増殖させます。
- 5,000 x g で4°Cで20分間遠心分離して細胞を回収し、ペレットを液体窒素で凍結し、さらに使用するまで-80°Cで保存します。
注:細胞ペレットの湿重量は、通常、培養液1 Lあたり5 gです。この時点で、プロトコルを一時停止できます。 - 細胞(2 g)を20 mLの溶解バッファーC1(表2)に解凍して再懸濁し、製造元の指示に従って、懸濁液を予冷(4°C)高圧細胞破壊システムに1,000 psiで3回通します。
注意: フレンチプレスを使用すると、重傷を負う可能性があります。使用するセルの圧力制限を超えないようにしてください。適切な個人用保護具を着用してください。 - 4,000 x g で15分間遠心分離することにより、ライセートから壊れていない細胞を取り除き、上清を回収します。
- 100,000 x g で1時間の超遠心分離によって膜を収集します。
- メンブレンペレットを10 mLのバッファーC2(表2)に再懸濁し、ボールベアリングガラスホモジナイザーを使用して等容量のSDSバッファーC3(表2)と混合します。
注意:バッファーC3中のβ-メルカプトエタノールは有毒です。関連するすべての安全規制に従ってください。 - 懸濁液を50°Cの水浴中で30分間インキュベートします。
- サンプルを100,000 x g で20°Cで1時間遠心分離します。
- ボールベアリングガラスホモジナイザーを用いてメンブレンペレットを10mLのSDSバッファーC4(表2)に再懸濁し、懸濁液を37°Cの水浴中で30分間インキュベートします。
注:ペレットを再懸濁できない場合は、SDSバッファーC4を20mLの容量まで追加します。 - サンプルを100,000 x g で20°Cで30分間遠心分離し、上清を回収します。
- 上清をオクチルポリオキシエチレン(オクチルPOE)と最終洗剤濃度0.5%(w / v)まで混合し、サンプルをバッファーC5(表2)に対して4°Cで24時間透析します。この目的のために、製造元の指示に従って、20 kDaのカットオフの透析チューブを使用し、サンプルを透析チューブまたはデバイスに入れます。
注:他の分子量のタンパク質を透析する場合は、透析チューブの切断を調整する必要があります。 - SDS-PAGEでサンプルの純度を評価し、メーカーのプロトコル(材料表)に従って、市販のタンパク質アッセイを使用してタンパク質濃度を決定します。
注:95°Cに加熱されたサンプルは単量体OmpFを示します。加熱されていないサンプルは、三量体形態のOmpFを示しています(図1F)。 - ショックフリーズ透析OmpFを液体窒素中のアリコートで行い、さらに使用するまで-20°Cで保存します。
注意: 液体窒素は換気の良い場所で取り扱う必要があります。適切な個人用保護具を着用してください。
2. DIB膜中のイオンチャネルの光シングルチャンネル記録
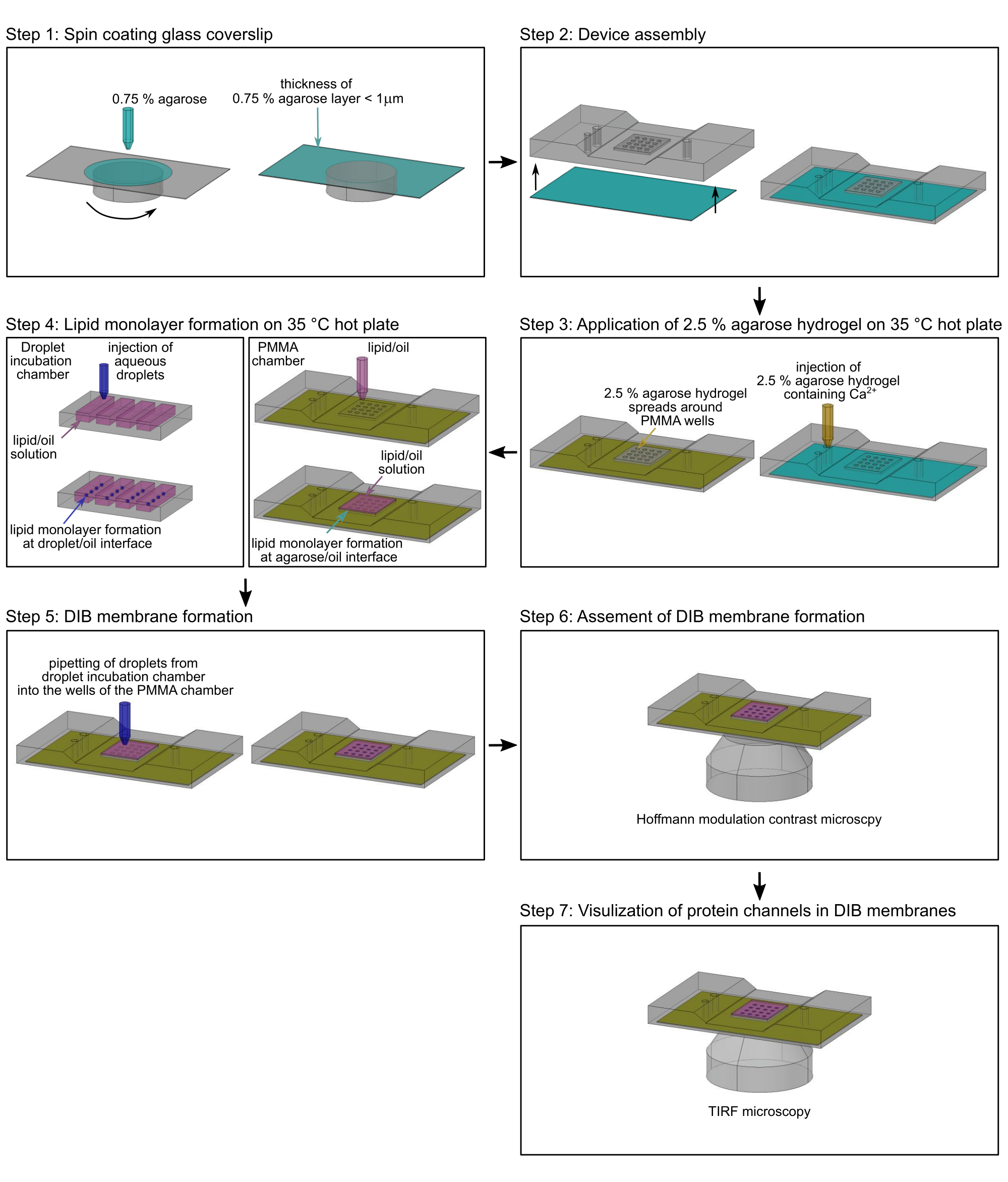
注:このセクションでは、微細加工ポリメチルメタクリレート(PMMA)チャンバー2 でDIBメンブレンを調製して、単一イオンチャネル17を通る横方向のタンパク質移動とイオンフラックスを監視する手順について説明します。チャンバーを製造するための寸法および正確な図面は、Lepthinら2に見出すことができる。 図2 は、PMMAチャンバー2 の組み立てとDIB膜の形成の概要を示しています。特に断らない限り、全ての工程は室温(RT)で行われる。図3は、DIBメンブレンの概略 図 と、単一チャネルタンパク質を通るCa2+ フラックスを使用して、メンブレン内の動きとチャネルの開閉状態の両方を監視する方法を示しています。
- 脂質の調製
- クロロホルムに溶解した1,2-ジフィタノイル-sn-グリセロ-3-ホスホコリン(25 mg/mL DPhPC)を含む脂質ストック溶液を-20°Cの冷凍庫から取り出し、室温まで昇温した。この目的のためには、一般に、サンプルを手で数分間ゆっくりと温めるだけで十分です。
注意: クロロホルムは有毒です。適切な個人用保護具を着用し、ドラフトの下でクロロホルムを含むすべての手順を実行してください。 - 380 μLのDPhPCストック溶液(25 mg/mL DPhPC)をガラスバイアルに移します。ゴム製のシーリングが付いたピペットは避けてください。代わりに、ステンレス鋼のプランジャーを備えたマイクロリットルのガラスシリンジを使用してください。
- 脂質原液を取り扱いた後、脂質原液をArまたはN2 ガスで重ねて脂質酸化を防止する。脂質が溶解している有機溶媒の蒸発やバイアルからの溶媒スプレーの飛散を避けるために、可能な限り低いガス流量を使用してください。有機溶媒を含む脂質を含むガラスバイアルの蓋の内側がポリテトラフルオロエチレン(PTFE)でコーティングされていることを確認してください。
- 脂質サンプルをN2の流れ下で乾燥させ(ステップ2.1.2)、オイルフリー真空ポンプを使用して真空下(2.0mbar)で一晩脂質サンプルから残りの有機溶媒を除去します。
- マイクロリットルガラスシリンジを使用して等量のヘキサデカンとシリコーンオイル(各500 μL)を加えて、脂質フィルムをヘキサデカン/シリコーンオイル溶液に溶解し、最終脂質濃度9.5 mg/mLにします。
注:脂質溶液はRTで数週間安定です。
- クロロホルムに溶解した1,2-ジフィタノイル-sn-グリセロ-3-ホスホコリン(25 mg/mL DPhPC)を含む脂質ストック溶液を-20°Cの冷凍庫から取り出し、室温まで昇温した。この目的のためには、一般に、サンプルを手で数分間ゆっくりと温めるだけで十分です。
- アガロースヒドロゲルの調製
- 二重脱イオン水中の低融点アガロース溶液(0.75%[w/v])約1 mLを調製し、ガラスカバーガラスのスピンコーティングに使用する加熱ブロックで85°Cに20分間加熱します。
注:アガロース溶液はRTで数週間保持でき、数回再加熱できます。 - 0.66 M CaCl 2および8.8 mM HEPES(pH 7.2)中の低融点2.5 %(w/v)アガロース溶液を調製し、加熱ブロックで85°Cに20分間加熱します。アガロースがよく溶けていることを確認してください。
注:アガロース溶液はRTで数週間保持でき、スピンコーティングに使用されるアガロース(ステップ2.2.1)と同様に数回再加熱できます。
- 二重脱イオン水中の低融点アガロース溶液(0.75%[w/v])約1 mLを調製し、ガラスカバーガラスのスピンコーティングに使用する加熱ブロックで85°Cに20分間加熱します。
- ヒドロゲルヘキサデカン/シリコーンオイル界面におけるポリメチルメタクリレート(PMMA)チャンバーアセンブリと脂質単分子膜形成
- ガラスカバースリップ(40 mm x 24 mm x 0.13 mm)をステンレス鋼のカバースリップホルダーに入れ、超音波洗浄機のアセトンでガラスビーカーで10分間洗浄します。カバーガラスを沈めるのに十分なアセトンを使用し、ビーカーをガラス板でゆるく覆い、洗浄プロセス中の蒸気の漏れを最小限に抑える無圧の閉鎖システムを作成します。
注意: アセトンは引火点の低い可燃性の高い溶剤です。蒸気/空気混合物は爆発性です。換気の良い場所で超音波洗浄機を操作してください。適切な個人用保護具を着用し、公式の安全上の注意(裸火や火花がないなど)を遵守してください。 - ガラスカバーガラスを二重脱イオン水ですすぎ、N2の流れの下で乾燥させます。
注意: この方法で洗浄されたカバーガラスは、数週間保管できます。 - カバーガラスをさらに洗浄し、プラズマクリーナーで酸素(0.5 mbar)で5分間親水化します。
注:プラズマ洗浄の親水性効果は時間の経過とともに低下するため、アガロース溶液でスピンコーティングする直前にこのステップを実行してください。 - プラズマ処理したカバーガラスをスピンコーターに取り付けます(図2、ステップ1)。
- 200 μLのピペットを使用して、140 μLの加熱した0.75%(w/v)低融点アガロース(ステップ2.2.1)を3,000 rpmで30秒間ゆっくりと添加することにより、カバーガラスをサブマイクロメートルの厚さのアガロースフィルムでコーティングします(図2、ステップ1)。
注:アガロースフィルムの厚さは、17で説明されているように、原子間力顕微鏡(AFM)によって決定できます。 - アガロースヒドロゲルの薄層を有するスピンコートカバーガラスを直ちにPMMAチャンバー2の下側に取り付ける。アガロースヒドロゲルが上を向いていることを確認します(図2、ステップ2)。
- カバーガラスの端を透明な粘着テープでPMMAマイクロマシニングデバイスに固定します。
- PMMAデバイスを35°Cに加熱したホットプレートに置きます(図2、ステップ3)。
- 200 μLの2.5%アガロース溶液(ステップ2.2.2)をピペットでチャンバーの入口に注意深く注ぎ(図2、ステップ3)、スピンコーティングによって塗布された薄いヒドロゲルが2.5%アガロース溶液のバッファーと平衡化し、水和したままになるようにします。2.5%アガロース溶液がスピンコーティングされたアガロースヒドロゲルの周囲に広がっていることを確認し、PMMAチャンバーを加熱プレートに軽く押し付けることで回避できます(図3A)。
- 直ちにPMMAチャンバーのウェル(図2、ステップ4)を合計60 μLの脂質/オイル溶液で覆い(ステップ2.1.5)、アガロース-オイル界面で脂質単層の形成を開始し、PMMAチャンバーのウェル内のスピンコーティングアガロースの脱水を回避します。デバイスを35°Cのホットプレートに約2時間保持します。
注:以前に発表された研究2によると、PMMAチャンバー内の円形ウェルの直径は0.5 mm、深さは1.8 mmです。
- ガラスカバースリップ(40 mm x 24 mm x 0.13 mm)をステンレス鋼のカバースリップホルダーに入れ、超音波洗浄機のアセトンでガラスビーカーで10分間洗浄します。カバーガラスを沈めるのに十分なアセトンを使用し、ビーカーをガラス板でゆるく覆い、洗浄プロセス中の蒸気の漏れを最小限に抑える無圧の閉鎖システムを作成します。
- ヘキサデカン/シリコーンオイル溶液中の脂質被覆水性液滴の調製
- 20 μLの脂質ヘキサデカン/シリコーンオイル溶液(ステップ2.1.5)を、液滴インキュベーションチャンバー内のいくつかの微細加工ウェルのそれぞれに入れます(図2、ステップ4)。脂質ヘキサデカン/シリコーンオイル溶液に適した容器は、薄いPMMAプレート2の40mm x 30mm x 3.5mmの小さなくぼみです。
- 垂直または水平のマイクロピペットプーラー(パッチピペットや鋭利なガラス電極の製造に使用)を使用して、先端開口部直径20μmのマイクロキャピラリーガラスニードルを準備します。7.5倍から35倍のズーム範囲で、低倍率で双眼実体顕微鏡下でマイクロキャピラリーガラス針の開口部直径を推定します。
注意: ガラスキャピラリーを引っ張る前の予熱モードの設定、引っ張るための加熱モード、および牽引力は、引っ張り装置のメーカーの仕様とキャピラリーのタイプに応じて、事前に実験的に決定する必要があります。 - マイクロキャピラリーガラスニードルに、8.8 mM HEPES(pH 7.2)、7 μM Fluo-8、400 μM EDTA、1.32 M KCl、および30 nM TOMコアコンプレックスを含む5 μLの水性注入液、またはマイクロキャピラリーピペットチップを使用して20 nM OmpFを充填します。多価金属イオン(Ca2+)と結合するキレート樹脂で予め処理した二重蒸留水のみを使用して水性注入液を調製する。
- マイクロキャピラリーガラスニードルを水性注入液とともにピエゾ駆動のナノインジェクターに取り付けます。
- ナノインジェクターを使用して、8.8 mM HEPES(pH 7.2)、7 μM Fluo-8、400 μM EDTA、1.32 M KCl、および30 nM TOMコアコンプレックス(ステップ1.1.12)、または20 nM OmpF(ステップ1.2.16)を含む100〜200 nLの水性液滴(図2、ステップ4)を、脂質ヘキサデカン/シリコーンオイル溶液(2.1.5を参照)で満たされた液滴インキュベーションチャンバー内のウェルに注入します。液滴を数ミリメートル(>10 mm)離してウェルに入れて、液滴が互いに接触しないようにしてください。それ以外の場合、脂質単層がオイル/バッファー界面でまだ形成されていない場合、それらはより大きな液滴に融合します。
注:クラスキャピラリーとナノインジェクターが利用できない場合は、2の説明に従って、0.1 μL〜0.5 μLの容量のシングルチャンネルマイクロリットルピペットを使用して液滴を手動で調製することもできます。ただし、この場合、液滴量はそれほど正確ではありません。 - 35°Cに加熱したホットプレート上でPMMAおよび液滴インキュベーションチャンバー(図2、ステップ4)を維持することにより、液滴/油界面に脂質単層を約2時間形成できます。
- DIB膜における単一イオンチャネルの調製とイメージング
- 10 μLの使い捨てポリプロピレンチップを備えたシングルチャンネルマイクロリットルピペットを使用して、実体顕微鏡下で液滴インキュベーションチャンバーのウェルからPMMAチャンバーのウェルに個々の水性液滴を手動で移します(図2、ステップ5)。液滴をヒドロゲル-油界面に形成された脂質単層上に約5分間沈め、液滴とアガロースヒドロゲルの間に脂質二重層(図3)を形成します。
- DIBメンブレンを備えたPMMAチャンバーを倒立光学顕微鏡のサンプルホルダーに取り付け、10倍ホフマン変調コントラスト対物レンズを使用してメンブレン形成を評価します(図2、ステップ6)。DIB膜形成は、液滴とヒドロゲルとの間の界面における透明な白色リングによって示される(図4A)。壊れたDIBメンブレンには、このリングは表示されません(図4B)。
注:あるいは、DIB膜は、位相差または微分干渉コントラスト(DIC)顕微鏡によって10倍の倍率で視覚化することができます。 - DIB膜が形成されている場合は、PMMAチャンバーを、従来の落射蛍光照明用光源、488 nmレーザー(Pmax = 100 mW)、および裏面照射型電子増倍CCDカメラ(512 x 512ピクセル、>95%QE)を備えたTIRF顕微鏡(図2、ステップ7)のサンプルホルダーに取り付けて、~0.16 μmのピクセルサイズを実現します。
- GFPフィルターセットを使用して、高輝度光源による落射蛍光照明下で、10倍の倍率対物レンズでDIB膜の端に焦点を合わせます。
- 液滴中の蛍光色素Fluo-8の弱いバックグラウンド蛍光を可視化できるGFPフィルターセットを使用して、100倍/N.A. 1.49アポクロマートオイルTIRF対物レンズを使用して、DIB膜の同じエッジを高倍率でファインフォーカスします(図4C)。
- フィルタ設定をGFPからクワッドバンドTIRFフィルタセットに変更します。
- 488 nmレーザーのスイッチを入れ、対物レンズのレーザーの強度を8 mW〜10 mWの値に設定します。
注:対物レンズでのレーザー強度に関する定量的な情報がないことが多いため、レーザー強度設定は、メーカーの仕様に従って、フォトダイオードセンサーを備えた光レーザーパワーメーターを使用して、レンズでの個別の測定で事前に校正する必要があります。
注意: レーザーの安全な操作を確実にするために、オペレーターはレーザー放射の潜在的な危険性と事故防止規則に注意する必要があります。 - 単一イオンチャネルを可視化するには、TIRF角度とEMCCDカメラゲインを調整し(EMゲイン乗数設定:285など)、DIBメンブレンの開いたイオンチャネルが暗い背景に高コントラストの蛍光スポットとして表示され(図4D-G)、目視検査時にS/G比が最大になるようにします。単一イオンチャネルを通るCa2+フラックスに対応するスポットが焦点を合わせたままで丸い形をしており、中央が高強度で、周辺に向かって徐々に減少することを確認します。チャネルのないメンブレンはバックグラウンド蛍光のみを示します(図4C)。
- 個々のチャネルの動きと開閉活動の経時を記録する前に、蛍光スポットに焦点が合っていることを確認し、イオンチャネルがDIB膜に再構成され、膜面内で横方向に移動していることを確認します。そうでない場合は、蛍光分子が膜近くのレーザーによって生成されたエバネッセント場に出入りしていることを示している可能性があります。
- 位置の適切な追跡と個々のイオンチャネルの開閉状態のモニタリングを可能にする一連の膜画像を記録します。横方向の移動度の種類(例えば、自由運動対過渡的な停泊地[参考のために、16を参照])およびチャネル活動の状態(例えば、開いたか閉じているか)を決定するには、十分に長い(例えば、30秒から1分)そして十分にサンプリングされた軌道(例えば、48 s-1のフレームレート)を取得する。
注:チャネリング拡散、制限拡散、またはホップ拡散16 の観測には、サンプリングレートが限られた境界内の無制限の拡散を測定するのに十分な速さが必要です。さらに、データのサンプリング時間が、制約なし拡散から制約拡散への遷移を測定するのに十分長いことを確認してください(詳細については、Jacobsonら16を参照)。
- 画像およびデータ処理
注:市販のソフトウェアパッケージに基づくオープンソースの画像処理パッケージ33 または自作ルーチン17 を使用して、DIB内の個々のイオンチャネルの時空間ダイナミクスを解析することができる。横方向の膜移動度を個々のチャネルを通るイオンフラックスと相関させるには、フィルターアルゴリズムを適用しないでください。- 漂白のための正しい画像時系列は、自己書き込みルーチンを用いて標準手順34を適用し、フル画像の平均フレーム強度
 をフィ
をフィ ッティングすることによって、ここでtはフレームインデックス(時間)であり、kはフィッティングパラメータである。次に、に従って時系列
ッティングすることによって、ここでtはフレームインデックス(時間)であり、kはフィッティングパラメータである。次に、に従って時系列 を修正します ここで、I(t)は強度です。あるいは、初期データ分析のために、カメラのスポットおよびピクセルサイズ(例えば、50ピクセル)に応じて、実装されたローリングボールアルゴリズム33およびローリングボール半径を有するオープンソースソフトウェアを使用してバックグラウンド補正を実行する。
を修正します ここで、I(t)は強度です。あるいは、初期データ分析のために、カメラのスポットおよびピクセルサイズ(例えば、50ピクセル)に応じて、実装されたローリングボールアルゴリズム33およびローリングボール半径を有するオープンソースソフトウェアを使用してバックグラウンド補正を実行する。 - 所定の時間tにおけるI(t)を、に従って
 可能な局所照明勾配を説明する平面傾斜を有する2次元ガウス関数にフィッティングすることにより、定義された関心領域(ROI)(例えば、30 x 30ピクセル)内の蛍光スポットの空間位置および振幅を決定する。ここで、x=(x,y)は蛍光強度情報によるROIであり、Aおよびσはガウスの振幅および幅であり、pkはROIのバックグラウンド強度を特徴付けるパラメータであり、μ=(x0,y0)はガウス分布の位置を定義する。 個々のチャネルを通るイオンフラックスは、パラメータAによって与えられる。チャネルの軌道はxで与えられ、チャネルの横方向の移動度を決定することができます。あるいは、35、36に記載されるようなオープンソースプラットフォーム33およびプラグインを使用して個々のスポットの位置および強度を決定することができる。EMCCDカメラ読み出しを光子数38に変換した後の位置精度37を推定する。
可能な局所照明勾配を説明する平面傾斜を有する2次元ガウス関数にフィッティングすることにより、定義された関心領域(ROI)(例えば、30 x 30ピクセル)内の蛍光スポットの空間位置および振幅を決定する。ここで、x=(x,y)は蛍光強度情報によるROIであり、Aおよびσはガウスの振幅および幅であり、pkはROIのバックグラウンド強度を特徴付けるパラメータであり、μ=(x0,y0)はガウス分布の位置を定義する。 個々のチャネルを通るイオンフラックスは、パラメータAによって与えられる。チャネルの軌道はxで与えられ、チャネルの横方向の移動度を決定することができます。あるいは、35、36に記載されるようなオープンソースプラットフォーム33およびプラグインを使用して個々のスポットの位置および強度を決定することができる。EMCCDカメラ読み出しを光子数38に変換した後の位置精度37を推定する。 - 時間に対する蛍光振幅Aおよび対応する軌跡xをプロットして、チャネル拡散率とチャネルを通るイオンフラックスとの間の可能な相関を決定する。これは、任意のグラフィックソフトウェアで実行します。自由横流路移動の場合、時間遅延τの線形回帰により横方向拡散係数Dを求め、xからスポットの平均二乗変位を算出する
 。
。
注:DIBメンブレン内を自由に移動するTOM-CCおよびOmpFの典型的な拡散係数17は、0.5〜1.5μm2 s-1の範囲です。拡散定数が0.01mm2・s-1未満の分子は固定化17と定義される。
- 漂白のための正しい画像時系列は、自己書き込みルーチンを用いて標準手順34を適用し、フル画像の平均フレーム強度
結果
リアルタイムの電極フリー光学シングルチャンネル記録は、DIB膜におけるタンパク質の横方向の動きと個々のイオンチャネルの機能との間の相互作用を明らかにします。ミトコンドリアTOMコア複合体(図1A)のDIB膜への再構成(図4D)は、横方向の移動度とイオン透過性(図5A)の間の強い時間的相関を示しています。TOM-CCゲーティングは、横方向の動きのモードに敏感であるように見えます17。移動チャネルは、細孔を通るCa2+フラックスと高い蛍光点強度を示します。トラップされた非移動分子は、低および中程度の蛍光強度を示します。ミトコンドリアTOM-CCの一般的なタンパク質輸入孔について、この単一分子アプローチは、横方向の移動度とイオン透過性の間に強い時間的相関を明らかにし、TOM-CCチャネルが機械感受性特性を有することを示唆した17。自由に動くTOM-CC分子の横方向の停止は、TOM-CCチャネルの部分的または完全な閉鎖を伴う。膜にほぼ完全に埋め込まれているOmpF(図1Eおよび図4F)でDIB膜をイメージングすると、ストップアンドゴー効果は見られません(図5B)。OmpFのランダムな停止は、強度の変化、したがってその細孔の閉鎖とは関連していません。蛍光シグナル38に基づいて、個々のチャネルの位置精度を、5〜10nmの範囲で推定することができる。しかしながら、例えば、120nmの二項平均変位を有する中間状態のTOM-CC分子について示されるように、アガロースヒドロゲルとの可動固定のためにチャネルがわずかにぐらつく場合、この精度は達成できないことに注意すべきである(図5A)。

図1:TOM-CCの分離。 (A)N. crassa TOM-CC30,39のクライオEM構造。6xHisタグを有するTom22を含むN.クラッサ株由来のミトコンドリアをDDMに可溶化し、Ni-NTAアフィニティークロマトグラフィー(B)および陰イオン交換クロマトグラフィー(C)に供した。(D) 孤立した TOM-CC の SDS-PAGE(E)精製大腸菌OmpFの結晶構造(PDB、1OPF)および(F)SDS-PAGE。この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

図2:PMMAチャンバーアセンブリのフローチャート。 ステップ1:ガラスカバーガラスをアガロースヒドロゲルでスピンコーティングします。ステップ2:スピンコーティングされたカバーガラスをカスタムメイドのPMMA顕微鏡チャンバーに取り付けます。ステップ3:追加の低溶融アガロースを35°Cのホットプレート上のPMMAチャンバーの入口ポートに追加します。ステップ4:脂質単層は、バッファー/オイル界面(左)およびアガロースヒドロゲル/オイル界面(右)の水性液滴の周りに形成されます。ステップ5:個々の水性液滴をPMMAチャンバーのウェルにピペットで入れ、2つの脂質単層が接触すると脂質二重層を形成します。ステップ6:DIB膜の形成は、ホフマン変調コントラスト顕微鏡によって検証されます。ステップ7:イオンチャネルが挿入されたDIB膜の選択された領域の画像をTIRF顕微鏡で取得します。緑:0.75%アガロース;黄色:Ca2+ イオンを含む2.5%アガロース;マゼンタ:脂質/油相;濃い青:Ca2+感受性色素およびタンパク質を含む水性液滴バッファー。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

図3:実験のセットアップ。 (A)PMMAウェル内のDIB膜の模式図。二層膜は0.75%の極薄アガロースフィルム上に置かれており、トランス中のCa2+感受性蛍光色素(Fluo-8)を使用して、イオンチャネルを通るCa2+フラックスのTIRFイメージングを経時的に行うことができます。(B)Ca2+フラックスは、シスからトランスへの浸透圧によってのみ制御されます。 これにより、膜内の位置の決定とチャネルの開閉状態の両方が可能になります。ここに示すチャネルは、N. crassa ミトコンドリア30のタンパク質伝導性チャネルTOM-CCである。(C) 単一の TOM-CC チャネルの軌跡。緑:移動チャネル。黄色:移動していないチャネル。この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

図4:DIBメンブレンにおけるTOM-CCおよびOmpFのイメージング 。 (a)ヒドロゲルと液滴との間の二層接触面積を示すホフマン変調コントラスト顕微鏡により画像化されたDIB膜。(B)(A)のように撮像された壊れたDIB膜。矢印、PMMAチャンバーの端。(C)タンパク質チャネルのないTIRF顕微鏡によって画像化されたDIB膜。(d)再構成されたTOM−CCを有するDIB膜を、TIRF顕微鏡により画像化する。白い四角形は、高強度(SH)、中強度(SI)、および低強度(SL)のスポットを示します。(E)(A)でマークした3つのスポットの蛍光強度プロファイルを2次元ガウス関数に当てはめると、個々のTOM-CCの位置とチャネルを通るCa2+ フラックスが明らかになります。(f)再構成されたOmpFを有するDIB膜。(G)(F)でマークされた蛍光スポットのガウスフィット。2細孔βバレルタンパク質複合体TOM-CCとは対照的に、3細孔βバレルOmpFは1つの透過状態のみを明らかにします。ピクセルサイズ、0.16μm。この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}

図5:チャネル活性はTOM-CCの横方向の移動度と相関 する。 (A)TOM-CCの蛍光振幅トレース(上)と対応する軌跡(下)は、TOM-CCの開閉チャネル活性が複合体の側方膜移動度と相関していることを示しています。軌跡には、3 つの透過性状態が表示されます。緑:完全に開いた状態。黄色:中間透過状態。赤:閉じたチャネル状態。赤い星:中間状態のTOM-CCは、その平均位置の周りで約±60 nmぐらつきます。全開状態および中間状態における蛍光シグナルに基づく位置精度37 は、5nm〜10nmの範囲である。(B)OmpFの蛍光振幅トレース(上)と対応する軌跡(下)。OmpFは、動いているか閉じ込められているかに関係なく、1つの強度レベルのみを明らかにします。トラップされた分子の期間に対応する軌道セグメントは灰色でマークされています。 この図の拡大版を表示するには、ここをクリックしてください。
{kind=link}
| バッファ | 試薬濃度 | 容積 | ||
| A1* | 20 mM トリス塩酸塩 pH 8.5、0.1 % (w/v) n-ドデシル-β-D-マルトシド (DDM)、10% (v/v) グリセロール、300 mM NaCl、および 1 mM フェニルメチルスルホニルフルオリド (PMSF) | 100ミリリットル | ||
| A2* | 20 mM トリス塩酸塩 pH 8.5、0.1% (w/v) DDM、10% (v/v) グリセロール、1 M イミダゾールおよび 1 mM PMSF | 100ミリリットル | ||
| B1* | 20 mM HEPES pH 7.2, 0.1% (w/v) DDM, 2% (v/v) ジメチルスルホキシド (DMSO) | 100ミリリットル | ||
| B2* | 20 mM HEPES pH 7.2, 0.1% (w/v) DDM, 1 M KCl, 2% (v/v) ジメチルスルホキシド (DMSO) | 100ミリリットル | ||
| * 使用前に0.22μmのフィルターを脱気して通過させてください。 | ||||
表1:TOM-CC分離用のバッファー溶液。
| バッファ | 試薬濃度 | 容積 | ||
| ポンド* | 1%(w / v)トリプトン、1%(w / v)NaClおよび0.5%(w / v)酵母エキス | 1100ミリリットル | ||
| C1クラス | 2 mM MgCl2、および~750ユニットのDNAseおよび50 mM トリス塩酸塩 pH 7.5 | 20ミリリットル | ||
| C2 クラス | 50 mM トリス塩酸塩、pH 7.5 | 50ミリリットル | ||
| C3クラス | 4%ドデシル硫酸ナトリウム(SDS)、2 mM β-メルカプトエタノールおよび50 mM トリス塩酸塩 pH 7.5 | 50ミリリットル | ||
| C4クラス | 2% (標準/v) SDS、500 mM NaClおよび 50 mM トリス塩酸塩 pH 7.5 | 50ミリリットル | ||
| C5クラス | 0.5% (w/v) オクチルポリオキシエチレン (オクチル POE), 1 mM EDTA および 20 mM トリス pH 8.5 | 1000ミリリットル | ||
| ※滅菌してからご使用ください。 | ||||
表2:OmpF単離用のバッファー溶液。
ディスカッション
ここで紹介するプロトコルは、単一分子TIRF顕微鏡を使用して、横方向イオンチャネルの動きとチャネル機能の間の相互作用を研究するためのDIBメンブレンの使用の概要を提供します。可能な限り最良のデータを得るためには、十分に分析できる個々の粒子の時系列を得るために、できるだけ多くの十分に分離されたチャネルを有する安定したDIB膜の調製が不可欠です。
最適化すべき重要なパラメータには、脂質の選択、油相中の脂質濃度、水性液滴中のタンパク質および界面活性剤濃度が含まれます。使用される脂質は、低温で明確な相転移を示さないという点で珍しいものです。DPhPCは、安定な膜系40を産生するために一般的に使用される脂質である。原則として、低温で流体環境を維持する脂質は、この用途に適している可能性があります。さらに、脂質は酸化に敏感であってはなりません。液滴中の洗剤濃度は、膜の破裂を避けるためにできるだけ低くする必要があります。膜タンパク質が沈殿しないことを考えると、安定した膜および良好なタンパク質取り込み率は、一般に臨界ミセル濃度(cmc)未満の界面活性剤濃度で達成される。
DIB膜が特定の界面活性剤21,41に耐えられない場合、またはタンパク質が低界面活性剤溶液からDIB膜に統合されない場合、タンパク質チャネルは最初に小さな単層脂質小胞(SUV)に再構成され、次に、大腸菌MscL42で成功裏に示されているように、液滴側からDIB膜に融合されます。.油相の脂質濃度が低すぎるためにDIB膜が形成されない場合があります。DIB膜の破裂を防ぐために、ヒドロゲルと液滴の間の浸透圧は、シスからトランスへのCa2+フラックスに過度に影響を与えることなく正確にバランスが取れていなければならないことにも注意する必要があります。最適化されたアガロースの厚さとメッシュサイズは、膜タンパク質の拡散を観察するために重要であるように思われます。アガロース層の乾燥は避けてください。厚さは、原子間力顕微鏡17を用いて決定することができる。スピンコーティング中のアガロース濃度、体積、回転速度を変えることで、ヒドロゲルのメッシュサイズと厚さを最適化することができます。ただし、ヒドロゲル層の厚さは画像のコントラストに影響することに注意してください。DIB中の膜タンパク質を捕捉するために、アガロースヒドロゲルをカスタム合成された非架橋のNi-NTA修飾低融点アガロースで置き換えて、His-tag17を介してそれらをトラップすることができます。過度に高い蛍光バックグラウンドは、多くの場合、DIB膜の破裂によって引き起こされます。これは、Ca2+感受性色素がヒドロゲルに拡散するため、マルチウェルチャンバーで特に問題になります。この場合、隣接する井戸は避けてください。膜上のCa2+感受性色素の蛍光漂白は、TIRFエバネッセント場の外側の液滴の大部分(図3A)中の非励起色素によって交換されるため、重要な制限因子ではないはずです。タンパク質の局在精度は、スポットのフィッティング精度とピクセルサイズによって決まります。
弱い蛍光シグナルは、チャネルを通る低いCa2+フラックスによって引き起こされる可能性があります。考えられる理由には、(i)不正確なTIRF設定(例えば、レーザー強度)、(ii)膜を横切る浸透圧Ca2+圧力、または(iii)チャネルの固有のCa2+透過性が低すぎることが含まれる。最初の問題に対処するには、レーザー強度、TIRF角度、およびカメラゲインを最適化する必要があります。後者の2つの問題は、膜2、43を横切る電位の印加によって克服することができる。ただし、電気的影響が実際には電圧制御されていないリガンド依存性または機械感受性イオンチャネルのチャネル開口に影響を与える可能性があるため、外部電圧を印加すると結果が歪む可能性があります。このようなチャネルの例は、ミトコンドリアタンパク質トランスロカーゼTOM−CC 27、およびそのチャネル形成サブユニットTom4026、44、45、46である。最後に、所望の機能性を達成するために膜タンパク質を特定の方向にDIB膜に挿入することはトリッキーであり、定量的研究はまれであることに注意すべきである47,48。場合によっては、組み込まれたタンパク質の配向はランダムである。特定の膜タンパク質は膜の片側だけで活性化されるため、これは膜タンパク質の研究にとって深刻な問題です。
TIRF顕微鏡は、平面支持膜における単一分子イベントに対処するための強力な方法です49。例としては、α-ヘモリシン50、ペルフリンゴライシンO51、OmpG52などのチャネルタンパク質の組み立ておよび折り畳み経路の解明が含まれます。これらの研究には、追加の技術としてFRETが含まれていました。加えて、機械感受性イオンチャネルMscLの活性化は、電流測定を用いた担持されたDIB二重層42の機械的刺激によって以前に研究されてきた。この研究に基づいて、将来の研究では、ここで説明するプラットフォームを単一分子FRET実験と組み合わせて、光学的に単一分子レベルで機械受容チャネルに対処することができます17。液滴への緩衝液の注入、内部DIB単層の伸張、または下にあるヒドロゲルへの個々のチャネルの標的結合は、MscLおよびMscS、2細孔ドメインK+チャネル、TREK-1、 TREK-2、およびTRAAK、およびPIEZO(レビューについては、53を参照されたい)だけでなく、タッチセンシティブイオンチャネルNOMPC54、55について示されているように、細胞細胞骨格への局所結合も含む。
開示事項
利益相反はないと宣言します。
謝辞
タンパク質の準備に協力してくれたベアテ・ニッチュケと、洞察に満ちた議論をしてくれたロビン・ゴーシュ、マイケル・シュヴァイケルト(シュトゥットガルト)、マクシミラン・ウルブリッヒ(フライブルク)に感謝します。この研究は、シュトゥットガルト研究センターシステム生物学(SRCSB)とバーデンヴュルテンベルク財団からの助成金(BiofMO-6からSN)の支援を受けました。
資料
| Name | Company | Catalog Number | Comments |
| 1,2-diphytanoyl-sn-glycero-3-phosphocholine | Avanti Polar Lipids | 850356C | |
| 100x Oil objective Apochromat N.A. 1.49 | Nikon | MRD01991 | TIRF microscope |
| 10x Hoffmann modulation contrast objective NA 0.25 | Nikon | Microscope to assess DIB membrane formation | |
| 10x objective N.A. 0.25 | Nikon | MRL00102 | TIRF microscope |
| 40x Hoffmann modulation contrast objective NA 0.55 | Nikon | Microscope to assess DIB membrane formation | |
| 488 nm laser, 100 mW | Visitron | TIRF microscope | |
| Adhesive tape | |||
| Äkta pure | Cytiva | Protein purification system | |
| Bradford assay kit, Pierce | Thermo Fisher | 23236 | |
| CaCl2 | Roth | 5239.2 | |
| Chelax 100 resin | Biorad | 143-2832 | |
| Chloroform | Sigma-Aldrich | MC1024452500 | |
| Dialysis cassettes Slide-A-Lyzer 20 k MWCO | Thermo Fisher | 87735 | |
| DIB chamber | Custom made | PMMA chamber for DIB membranes | |
| Digital power meter and energy console | Thorlabs | PM100D | Laser power meter |
| Dimethyl sulfoxide | Roth | 4720.1 | |
| Double distilled H2O | |||
| Eclipse TS 100 Hoffmann modulation contrast microscope | Nikon | Microscope to assess DIB membrane formation | |
| EDTA | Roth | 8042.2 | |
| EMCCD camera iXon Ultra 897 | Andor | TIRF microscope | |
| Ethanol | Sigma-Aldrich | 32205-M | |
| Fixed angle rotor Ti70 | Beckman Coulter | ||
| Fluo-8, CalciFluorTM | Santa Cruz Biotechnology | SC-362561 | Ca2+-sensitive dye |
| French press cell disruption homogenizer | Igneus | Igneus 40000 psi | |
| GFP filter | AHF | Filter seeting used for excitation of DIB-membranes by epifluorescence with white light source | |
| Glass capillaries | World Precision Instruments | 4878 | |
| Glass coverslips 40 mm x 24 mm x 0.13 mm | Roth | 1870.2 | |
| Glycerol | Roth | 3783.2 | |
| Hamilton syringe 10 mL | Roth | X033.1 | |
| Hamilton syringe 100 mL | Roth | X049.1 | |
| Hamilton syringe 500 mL | Roth | EY49.1 | |
| Heating block | Eppendorf | Thermomixer comfort | |
| Heating plate | Minitube | HT200 | |
| Hepes | Roth | 9205.3 | |
| Hexadcane | Sigma-Aldrich | 296317 | |
| His Trap HP 1 mL | Cytiva | 29051021 | Ni-NTA column |
| Imidazole | Sigma-Aldrich | 1.04716.1000 | |
| KCl | Honeywell | 10314243 | |
| KLM spin coater | Schaefer Tec | SCV-10 | |
| List medical L/M-3P-A vertical pipette puller | Artisan Technology Group | 57761-1 | |
| Low melting point agarose | Sigma-Aldrich | A9414 | |
| M8 Stereomicroscope | Wild | Stereomicrosope | |
| Matlab | MathWorks | R2022a | |
| Methanol | Sigma-Aldrich | 34860 | |
| MicroFil pipette tips | World Precision Instruments | MF34G-5 | |
| N2 gas | |||
| NaCl | Roth | 3957.1 | |
| Nanoliter 2010 injector | World Precision Instruments | Nanoliter 2010 | |
| n-dodecyl-b-D-maltoside | Glycon Biochemicals | D97002-C | |
| Ni-NTA agarose, non-crosslinked | Cube Biotech | 124115393 | Custom made |
| NIS-Elements AR software | Nikon | MQS31100/MQS42560/MQS42580/MQS42780/MQS41930 | Imaging software |
| n-octyl-polyoxyethylene | Sigma-Aldrich | 40530 | |
| O2 gas | |||
| Phenylmethylsulfonyl fluoride | Roth | 6367.3 | |
| Photodiode sensor Si, 400 - 1100 nm, 500 mW | Thorlabs | S130C | Sensor for laser power meter |
| Plasma cleaner | Diener Electronics | Zepto | |
| Preparative ultracentrifuge Optima | Beckman Coulter | ||
| Quad-band TIRF-filter 446/523/600/677 HC | AHF | Filter setting used for excitation of DIB-membranes with 488 nm laser | |
| Resource Q 1 mL | Cytiva | 17117701 | Anion exchange column |
| Silicon oil AR 20 | Sigma-Aldrich | 10836 | |
| Sodium dodecyl sulfate | Roth | 2326.2 | |
| Super LoLux camera | JVC | Stereomicrosope | |
| Thermoshaker | Gerhardt | THL 500/1 | |
| Ti-E Fluorescence microscope | Nikon | MEA53100 | |
| Tris-HCl | Sigma-Aldrich | 9090.3 | |
| Tryptone | Roth | 8952.2 | |
| Ultrasonic bath | Bandelin Sonorex | RK 100 | |
| Vaccum pump | Vacuubrand | MD 4C NT | |
| White light source for epifluorescence illumination (100 W) | Nikon | MBF72655 | TIRF microscope |
| Yeast extract | Roth | 2363.2 | |
| β-mercaptoethanol | Sigma-Aldrich | M3148 |
参考文献
- Tanaka, M., Sackmann, E. Polymer-supported membranes as models of the cell surface. Nature. 437 (7059), 656-663 (2005).
- Leptihn, S., et al. Constructing droplet interface bilayers from the contact of aqueous droplets in oil. Nature Protocols. 8 (6), 1048-1057 (2013).
- Thompson, J. R., Heron, A. J., Santoso, Y., Wallace, M. I. Enhanced stability and fluidity in droplet on hydrogel bilayers for measuring membrane protein diffusion. Nano Letters. 7 (12), 3875-3878 (2007).
- Kusumi, A., et al. Paradigm shift of the plasma membrane concept from the two-dimensional continuum fluid to the partitioned fluid: high-speed single-molecule tracking of membrane molecules. Annual Review of Biophysics and Biomolecular Structure. 34 (1), 351-378 (2005).
- Qian, H., Sheetz, M. P., Elson, E. L. Single particle tracking. Analysis of diffusion and flow in two-dimensional systems. Biophysical Journal. 60 (4), 910-921 (1991).
- Kusumi, A., Shirai, Y. M., Koyama-Honda, I., Suzuki, K. G. N., Fujiwara, T. K. Hierarchical organization of the plasma membrane: Investigations by single-molecule tracking vs. fluorescence correlation spectroscopy. FEBS Letters. 584 (9), 1814-1823 (2010).
- Betaneli, V., Schwille, P. Fluorescence correlation spectroscopy to examine protein-lipid interactions in membranes. Methods in Molecular Biology. 974, 253-278 (2013).
- Ando, T., et al. A high-speed atomic force microscope for studying biological macromolecules. Proceedings of the National Academy of Sciences. 98 (22), 12468-12472 (2001).
- Casuso, I., et al. Characterization of the motion of membrane proteins using high-speed atomic force microscopy. Nature Nanotechnology. 7 (8), 525-529 (2012).
- Karner, A., et al. Tuning membrane protein mobility by confinement into nanodomains. Nature Nanotechnology. 12 (3), 260-266 (2017).
- Rajendran, L., Simons, K. Lipid rafts and membrane dynamics. Journal of Cell Science. 118 (6), 1099-1102 (2005).
- Schink, K. O., Tan, K. -. W., Stenmark, H. Phosphoinositides in control of membrane dynamics. Annual Review of Cell and Developmental Biology. 32 (1), 143-171 (2016).
- Schafer, D. A. Coupling actin dynamics and membrane dynamics during endocytosis. Current Opinion in Cell Biology. 14 (1), 76-81 (2002).
- Lingwood, D., Ries, J., Schwille, P., Simons, K. Plasma membranes are poised for activation of raft phase coalescence at physiological temperature. Proceedings of the National Academy of Sciences. 105 (29), 10005-10010 (2008).
- Engelman, D. M. Membranes are more mosaic than fluid. Nature. 438 (7068), 578-580 (2005).
- Jacobson, K., Liu, P., Lagerholm, B. C. The lateral organization and mobility of plasma membrane components. Cell. 177 (4), 806-819 (2019).
- Wang, S., et al. Spatiotemporal stop-and-go dynamics of the mitochondrial TOM core complex correlates with channel activity. Communications Biology. 5 (1), 471 (2022).
- Ide, T., Yanagida, T. An artificial lipid bilayer formed on an agarose-coated glass for simultaneous electrical and optical measurement of single ion channels. Biochemical and Biophysical Research Communications. 265 (2), 595-599 (1999).
- Funakoshi, K., Suzuki, H., Takeuchi, S. Lipid bilayer formation by contacting monolayers in a microfluidic device for membrane protein analysis. Analytical Chemistry. 78 (24), 8169-8174 (2006).
- Heron, A. J., Thompson, J. R., Mason, A. E., Wallace, M. I. Direct detection of membrane channels from gels using water-in-oil droplet bilayers. Journal of the American Chemical Society. 129 (51), 16042-16047 (2007).
- Bayley, H., et al. Droplet interface bilayers. Molecular BioSystems. 4 (12), 1191-1208 (2008).
- Huang, S., Romero-Ruiz, M., Castell, O. K., Bayley, H., Wallace, M. I. High-throughput optical sensing of nucleic acids in a nanopore array. Nature Nanotechnology. 10 (11), 986-991 (2015).
- Sackmann, E. Supported membranes: scientific and practical applications. Science. 271 (5245), 43-48 (1996).
- Kiessling, V., Yang, S. -. T., Tamm, L. K. Supported lipid bilayers as models for studying membrane domains. Current Topics in Membranes. 75, 1-23 (2015).
- Murray, D. H., Tamm, L. K., Kiessling, V. Supported double membranes. Journal of Structural Biology. 168 (1), 183-189 (2009).
- Hill, K., et al. Tom40 forms the hydrophilic channel of the mitochondrial import pore for preproteins. Nature. 395 (6701), 516-521 (1998).
- Poynor, M., Eckert, R., Nussberger, S. Dynamics of the preprotein translocation channel of the outer membrane of mitochondria. Biophysical Journal. 95 (3), 1511-1522 (2008).
- Künkele, K. P., et al. The preprotein translocation channel of the outer membrane of mitochondria. Cell. 93 (6), 1009-1019 (1998).
- Ahting, U., et al. The TOM core complex: the general protein import pore of the outer membrane of mitochondria. The Journal of Cell Biology. 147 (5), 959-968 (1999).
- Bausewein, T., et al. Cryo-EM structure of the TOM core complex from Neurospora crassa. Cell. 170 (4), 693-700 (2017).
- Cowan, S. W., et al. Crystal structures explain functional properties of two E. coli porins. Nature. 358 (6389), 727-733 (1992).
- Bieligmeyer, M., et al. Reconstitution of the membrane protein OmpF into biomimetic block copolymer-phospholipid hybrid membranes. Beilstein Journal of Nanotechnology. 7, 881-892 (2016).
- Schindelin, J., et al. Fiji: an open-source platform for biological-image analysis. Nature Methods. 9 (7), 676-682 (2012).
- Vicente, N. B., Zamboni, J. E. D., Adur, J. F., Paravani, E. V., Casco, V. H. Photobleaching correction in fluorescence microscopy images. Journal of Physics: Conference Series. 90 (1), 012068 (2007).
- Tinevez, J. -. Y., et al. TrackMate: An open and extensible platform for single-particle tracking. Methods. 115, 80-90 (2017).
- Ershov, D., et al. TrackMate 7: integrating state-of-the-art segmentation algorithms into tracking pipelines. Nature Methods. 19 (7), 829-832 (2022).
- Thompson, R. E., Larson, D. R., Webb, W. W. Precise nanometer localization analysis for individual fluorescent probes. Biophysical Journal. 82 (5), 2775-2783 (2002).
- Hirsch, M., Wareham, R. J., Martin-Fernandez, M. L., Hobson, M. P., Rolfe, D. J. A stochastic model for electron multiplication charge-coupled devices - From theory to practice. PLoS One. 8 (1), 53671 (2013).
- Bausewein, T., Naveed, H., Liang, J., Nussberger, S. The structure of the TOM core complex in the mitochondrial outer membrane. Biological Chemistry. 401 (6-7), 687-697 (2020).
- Lindsey, H., Petersen, N. O., Chan, S. I. Physicochemical characterization of 1,2-diphytanoyl-sn-glycero-3-phosphocholine in model membrane systems. Biochimica et Biophysica Acta. 555 (1), 147-167 (1979).
- Manafirad, A. Single ion-channel analysis in droplet interface bilayer. Methods in Molecular Biology. 2186, 187-195 (2021).
- Rosholm, K. R., et al. Activation of the mechanosensitive ion channel MscL by mechanical stimulation of supported Droplet-Hydrogel bilayers. Scientific Reports. 7 (1), 45180 (2017).
- Wang, Y., et al. Electrode-free nanopore sensing by DiffusiOptoPhysiology. Science Advances. 5 (9), (2019).
- Ahting, U., et al. the pore-forming component of the protein-conducting TOM channel in the outer membrane of mitochondria. The Journal of Cell Biology. 153 (6), 1151-1160 (2001).
- Romero-Ruiz, M., Mahendran, K. R., Eckert, R., Winterhalter, M., Nussberger, S. Interactions of mitochondrial presequence peptides with the mitochondrial outer membrane preprotein translocase TOM. Biophysical Journal. 99 (3), 774-781 (2010).
- Kuszak, A. J., et al. Evidence of distinct channel conformations and substrate binding affinities for the mitochondrial outer membrane protein translocase pore Tom40. The Journal of Biological Chemistry. 290 (43), 26204-26217 (2015).
- Yanagisawa, M., Iwamoto, M., Kato, A., Yoshikawa, K., Oiki, S. Oriented reconstitution of a membrane protein in a giant unilamellar vesicle: Experimental verification with the potassium channel KcsA. Journal of the American Chemical Society. 133 (30), 11774-11779 (2011).
- Goers, R., et al. Optimized reconstitution of membrane proteins into synthetic membranes. Communications Chemistry. 1, 35 (2018).
- Castell, O. K., Dijkman, P. M., Wiseman, D. N., Goddard, A. D. Single molecule fluorescence for membrane proteins. Methods. 147, 221-228 (2018).
- Thompson, J. R., Cronin, B., Bayley, H., Wallace, M. I. Rapid assembly of a multimeric membrane protein pore. Biophysical Journal. 101 (11), 2679-2683 (2011).
- Senior, M. J. T., et al. Single-molecule tracking of perfringolysin O assembly and membrane insertion uncoupling. The FEBS Journal. , (2022).
- Weatherill, E. E., et al. Fast slow folding of an outer membrane porin. Proceedings of the National Academy of Sciences. 119 (20), 2121487119 (2022).
- Kefauver, J. M., Ward, A. B., Patapoutian, A. Discoveries in structure and physiology of mechanically activated ion channels. Nature. 587 (7835), 567-576 (2020).
- Yan, Z., et al. Drosophila NOMPC is a mechanotransduction channel subunit for gentle-touch sensation. Nature. 493 (7431), 221-225 (2013).
- Wang, Y., et al. The push-to-open mechanism of the tethered mechanosensitive ion channel NompC. eLife. 10, 58388 (2021).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved