
JoVE 비디오를 활용하시려면 도서관을 통한 기관 구독이 필요합니다. 전체 비디오를 보시려면 로그인하거나 무료 트라이얼을 시작하세요.
Method Article
섬모 박동 빈도의 정량화를 위한 1차 인간 비강 상피 세포 모델의 수집, 확장 및 분화
요약
이 프로토콜은 비강 상피 세포 수집, 확장 및 기관형 기도 상피 세포 모델로의 분화와 라이브 셀 이미징 및 맞춤형 스크립트를 통한 섬모 박동 빈도의 정량화를 설명합니다.
초록
섬모 기능 (박동 빈도, 패턴)의 측정은 원발성 섬모 운동 이상증과 같은 호흡기 질환의 진단 도구로 확립되었습니다. 그러나, 이들 기술의 더 넓은 적용은 온도, 습도 및 pH와 같은 환경 인자의 변화에 대한 섬모 기능의 극단적 인 민감성에 의해 제한된다. 낭포 성 섬유증 (CF) 환자의기도에서 점액 축적은 섬모 박동을 방해합니다. 섬모 기능은 CF 막 횡단 전도도 조절기 (CFTR) 채널 활성의 지표로서 1 차기도 세포 모델에서 조사되었습니다. 그러나 동일한 CFTR 돌연변이를 가진 환자의 경우에도 CFTR 조절 약물에 대한 반응으로 섬모 박동 빈도의 상당한 환자 간 변동성이 발견되었습니다. 또한, 기능 장애 CFTR 조절 염화물 분비가 섬모 기능에 미치는 영향은 잘 이해되지 않았습니다. 현재 체외 기도 모델의 샘플 준비, 이미지 획득 및 섬모 박동 주파수 (CBF) 분석을 입증하는 포괄적 인 프로토콜은 없습니다. 환경적으로 통제된 조건에서 수행된 표준화된 배양 조건 및 이미지 획득은 개인 간 및 CFTR 조절 약물에 대한 반응으로 CBF의 일관되고 재현 가능한 정량화를 가능하게 합니다. 이 프로토콜은 세 가지 다른 기도 상피 세포 모델 시스템에서 CBF의 정량화를 설명합니다: 1) 천연 상피 시트, 2) 투과성 지지 삽입물 상에서 이미징된 공기-액체 인터페이스 모델, 및 3) 세포외 기질-포매된 3차원 오가노이드. 후자의 두 가지는 생체 내 폐 생리학을 복제하여 섬모를 뛰고 점액을 생성합니다. 섬모 기능은 환경 제어 챔버에서 고속 비디오 카메라를 사용하여 캡처됩니다. 사용자 정의 스크립트는 CBF 분석에 사용됩니다. CBF 측정을 클리닉으로 번역하는 것은 환자별로 CFTR 조절 약물에 대한 반응을 예측하는 데 중요한 임상 도구로 구상됩니다.
서문
섬모 박동 빈도(CBF) 및 패턴의 측정은 원발성 섬모 운동 이상증(PCD)1과 같은 호흡기 질환의 진단 도구로 확립되었습니다. 낭포 성 섬유증 (CF)에서 CF 막 횡단 전도도 조절기 (CFTR) 염화물 채널의 기능 장애는기도 표면 액체의 탈수 및 점액 섬모 클리어런스 손상2을 유발합니다. 섬모 기능은 CFTR 채널 활성의 지표로서 일차 기도 세포 모델에서 시험관내에서 조사되었다3. 그러나, 동일한 CFTR 돌연변이를 가진 환자의 경우에도, CFTR 조절 약물에 대한 반응으로 CBF에 상당한 환자 대 환자 가변성이 존재한다3. 또한, 기능 장애 CFTR 조절 염화물 분비가 섬모 기능에 미치는 영향은 잘 이해되지 않았습니다. 현재 체 외 기도 모델의 샘플 준비, 이미지 획득 및 CBF 분석을 입증하는 포괄적 인 프로토콜은 없습니다.
비강 점막 칫솔질로부터 분리 된 비강 상피 시트는 PCD 진단4을위한 섬 모체 기능의 측정에 직접 사용된다. 그러나, 수득된 비강 상피 시트의 크기 또는 품질에 대한 제어는 없지만, CBF는 단일 세포 또는 세포 시트 및 파쇄되거나 중단되지 않은 상피 시트 섬모 가장자리에서 측정되는지 여부에 따라달라집니다5. 따라서 비강 점막 칫솔질을 수집하는 동안 세포 손상으로 인한 이차성 운동 이상증은 CBF에 영향을 미칠 수 있습니다. 비강 상피 세포의 1 차 세포 배양 및 공기-액체 계면 (ALI) 또는 3 차원 기저막 매트릭스에서 섬모기도 상피 오가노이드로의 분화는 2 차 운동 이상증 4,6,7,8이없는 섬모를 생성합니다. ALI에서 분화된 기도 상피 세포(이하 ALI 모델이라고 함)는 생체 외 코 점막 칫솔질의 섬모 박동 패턴과 빈도를 복제하고6 환자 특이적 결함을 유지하면서 섬모 미세 구조, 박동 패턴 및 박동 빈도를 분석할 수 있는 중요한 2차 진단 보조제로 간주되었습니다.9 . 그러나 이러한 유사층화되고 점액섬모로 분화된 세포 모델을 만드는 데 사용되는 방법론에는 불일치가 존재합니다. 상이한 배양 확장 또는 분화 프로토콜은 뚜렷한 상피 표현형(섬모 또는 분비)10을 유도하고 CBF11에 상당한 차이를 초래할 수 있습니다. CBF 는 비강 상피 브러싱 4,6,12,13,14,15,16,기도 상피 오가노이드14,17,18 및 ALI 모델 3,4,6,13,19,20 에서 정량화되었으며, 21. 그러나 이러한 프로토콜 중에는 큰 변동성이 있으며 종종 많은 매개 변수가 제어되지 않습니다. 예를 들어, 일부 연구에서, CBF는 ALI 모델의 세포가 투과성 지지체 삽입물 3,19,20,21 상에 남아있는 동안 현장에서 이미징되지만, 다른 것들은 투과성 지지체 삽입물로부터 세포를 긁어내고 이들을 배지 4,6,13에 부유시키는 것을 이미지화한다.
또한, 섬모 기능을 측정하는 기술의 광범위한 적용은 환경 요인의 변화에 대한 섬모 기능의 극단적 인 민감성에 의해 제한됩니다. 온도 22, 습도 23,24 및 pH 25,26과 같은 환경 요인은 섬모 기능에 영향을 미치며 CBF를 정확하게 정량화하기 위해 조절되어야 합니다. 다양한 실험실에서 사용되는 다양한 생리적 매개 변수와 CBF에 미치는 영향은 이전에 검토되었습니다27.
CBF 측정에 대한 다양한 이미징 기술 및 접근법이 문헌에보고되어 있습니다. PCD 진단의 경우 비디오 현미경을 사용하여 섬모 기능28,29를 측정합니다. 최근에, 차동 동적 현미경에 기초한 비디오 분석 알고리즘이 기도 상피 세포 ALI 모델 3,30에서 CBF 및 섬모 협응을 정량화하는 데 사용되었다. 이 방법은 영역을 분할하거나 선택할 필요 없이 빠르고 완전히 자동화된 방식으로 기도 상피 세포에서 섬모 박동의 특성을 분석할 수 있습니다. CBF의 이미징 및 정량화를 위한 다양한 방법은 문헌에서 CBF에 보고된 차이점을 추가할 수 있습니다(보충 파일 1).
배양에서 정량화에 이르는 프로토콜은 기존 방법을 간소화하고, 배양 조건의 표준화 및 엄격한 환경 통제 조건에서 수행되는 이미지 획득을 통해 개인 내 및 개인 간에 CBF의 일관되고 재현 가능한 정량화를 가능하게 합니다.
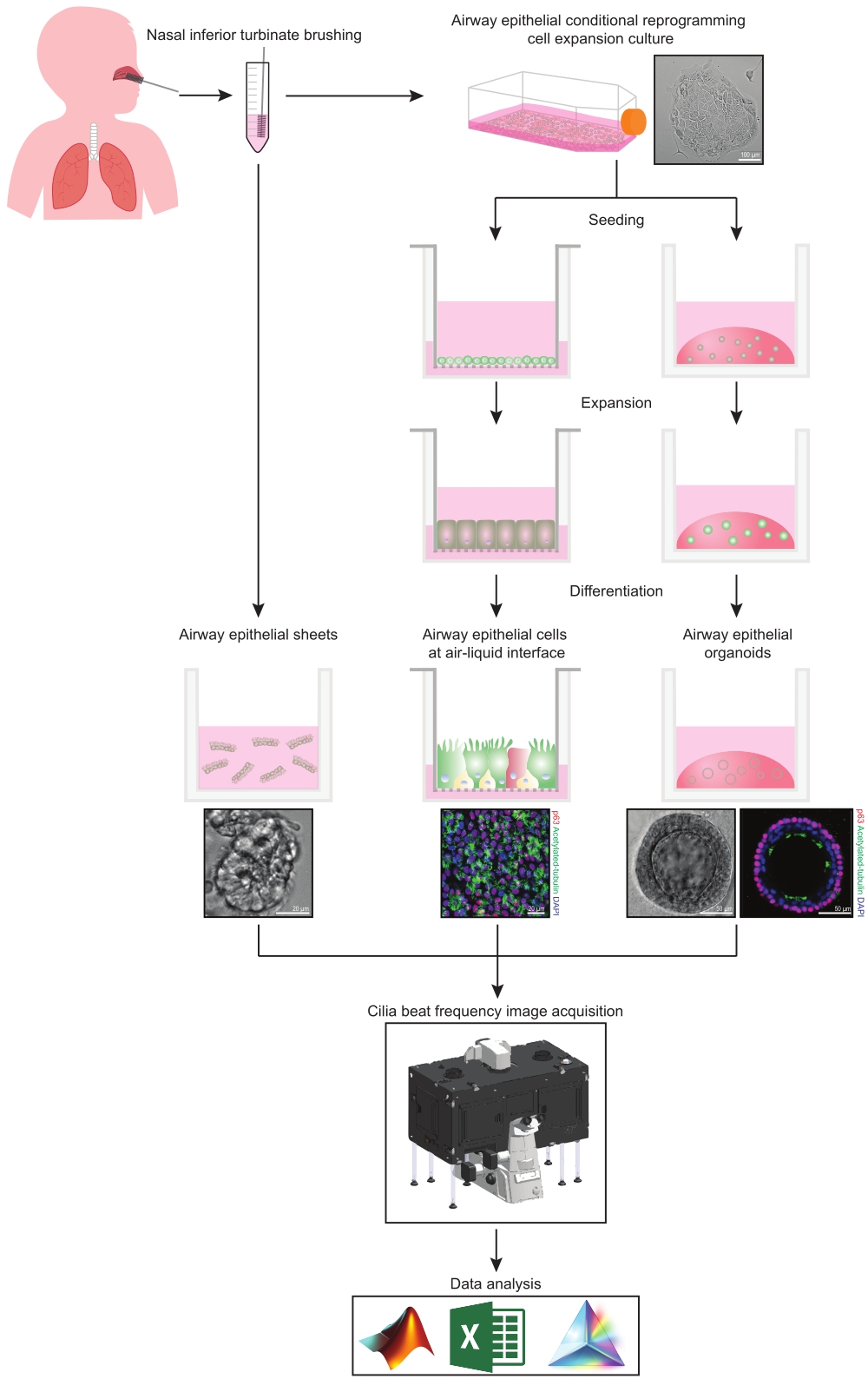
이 프로토콜은 비강 기원의 세 가지 다른 기도 상피 세포 모델 시스템에서 상피 세포 수집, 확장 및 분화 배양 조건, CBF의 정량화에 대한 완전한 설명을 제공합니다: 1) 네이티브 상피 시트, 2) 투과성 지지 삽입물에서 이미징된 ALI 모델 및 3) 세포외 기질(ECM) 내장 3차원 오가노이드(그림 1 ). 비강 하비갑개 칫솔질로부터 수득된 비강 상피 세포는 기관지 칫솔질 수집과 관련된 침습적 절차를 극복하면서 기관지 상피 세포(31)에 대한 효과적인 대용물이기 때문에 기도 상피의 대표자로서 사용된다. 조건부 재프로그래밍 세포(CRC) 방법은 ALI 모델 및 3차원 오가노이드 생성을 위해 일차 기도 상피 세포를 확장하는 데 사용됩니다. 줄기 세포와 유사한 상태로의 기도 상피 세포의 조건부 재프로그래밍은 성장-정지된 섬유아세포 영양세포 시스템 및 Rho-관련 키나아제 (ROCK) 억제제32와의 공동 배양에 의해 유도된다. 중요하게도, CRC 방법은 조직 특이적 분화 잠재력을 유지하면서 기도 상피 세포의 집단 배가를 증가시킵니다(33,34). 모든 기도 상피 세포 모델에서 섬모 기능은 표준화된 이미지 획득 설정을 갖춘 고속 비디오 카메라를 사용하여 온도 제어 챔버에서 캡처됩니다. 맞춤형 스크립트는 CBF의 정량화를 위해 사용됩니다.

그림 1: 워크플로의 개략도. 참가자의 비강 하비갑개를 닦은 후 기도 상피 세포는 두 가지 방법 중 하나로 활용됩니다. 기도 상피 시트가 분리되고 섬모 박동 빈도가 즉시 이미지화되거나 기도 상피 세포가 조건부 재프로그래밍 세포 방법을 통해 확장됩니다. CRC-확장 기도 상피 세포는 공기-액체 계면 또는 기도 상피 오가노이드 배양에서 기도 상피 세포를 확립하기 위해 분화됩니다. 섬모 박동 주파수의 이미징은 가열 및 습도 환경 챔버와 빠른 프레임 속도(>100Hz) 과학 카메라가 있는 라이브 셀 이미징 현미경을 사용하여 획득됩니다. 데이터 분석은 사용자 지정 스크립트를 사용하여 수행됩니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
프로토콜
연구 승인은 Sydney Children's Hospital Network Ethics Review Board (HREC/16/SCHN/120)로부터 받았다. 생체 표본을 수집하기 전에 모든 참가자(또는 참가자의 보호자)로부터 서면 동의를 받았습니다.
1. 기도 상피세포 모델 구축을 위한 준비
- 80% Dulbecco의 변형 독수리 배지와 20% 태아 소 혈청을 결합하여 비강 세포 수집 배지를 준비합니다. 1 μL/mL의 페니실린/스트렙토마이신을 보충하십시오. 4°C에서 최대 3개월 동안 보관하십시오.
- 플라스크 또는 투과성 지지체 삽입물을 1.2.1-1.2.4단계에 따라 필요에 따라 콜라겐 용액으로 코팅합니다. 콜라겐이 코팅된 용기를 장기간 보관하지 마십시오.
- I형 콜라겐 용액(3mg/mL 스톡)을 인산완충식염수(PBS)로 1:100 희석하여 0.03mg/mL의 최종 농도로 만듭니다. 잘 섞는다.
- 준비된 콜라겐 용액의 세포 배양 플라스크(섹션 4)를 160μL/cm2(즉, T25 플라스크당 4mL) 및 투과성 지지체 삽입물(섹션 5)을 455μL/cm2(즉, 6.5mm 삽입물당 150μL)로 코팅합니다.
- 37 ° C에서 2-24 시간 동안 배양하십시오.
- 세포를 파종하기 전에 피펫 또는 진공 흡인기로 콜라겐 용액을 제거합니다. 세포를 파종하기 전에 용기를 씻지 마십시오.
- 표 1에 나열된 구성 요소(32)를 결합하여 조건부 재프로그래밍 셀(CRC) 배지를 준비합니다. 필터는 병 상단 진공 필터 시스템을 사용하여 살균합니다. 4°C에서 최대 2개월 동안 보관하십시오.
- 사용 당일에, 표 1에 나타낸 바와 같이 인간 표피 성장 인자, ROCK 억제제 및 항생제를 첨가한다.
| 구성 요소 | 음량 |
| DMEM, 고 포도당 | 156.7 밀리리터 |
| DMEM / F-12, 헤르페스 | 313.3 밀리리터 |
| 하이드로 코르티손 | 55.6 μL |
| 인슐린 | 1.25 밀리리터 |
| 콜레라 독소 | 21 μL |
| 아데닌 | 1.2 밀리리터 |
| 하이 FBS | 25 밀리리터 |
| 페니실린-스트렙토 마이신 | 5 밀리리터 |
| 인간 표피 성장 인자 | 1 μL/밀리람베르트 |
| 암석 억제제 | 1 μL/밀리람베르트 |
| 곰팡이 지대 | 2 μl/밀리람베르트 |
| 토브라마이신 | 2 μL/mL |
| 세프타지딤 수화물 | 4 μL/밀리리터 |
| 겐타 마이신 용액 | 1 μL/밀리람베르트 |
표 1: 500mL의 조건부 재프로그래밍 세포 배지를 위한 구성 요소
2. 비강 하비갑개 칫솔질 수집
알림: 프로토콜의 이 섹션에는 비강 세포 수집 배지, 세포학 브러시, 조직 및 적절한 개인 보호 장비가 있는 수집 튜브(50mL)가 필요합니다. 상부 호흡기 감염 중 칫솔질을 피하십시오. 출혈의 위험이 적으며 염증이 있으면 증가합니다. 칫솔질의 목적이 생체외 CBF 측정을 위한 기도 상피 시트를 얻는 것이라면 브러싱은 임의의 상부 호흡기 감염 후 최소 6주가 지나야 한다; 이상적으로는 감염 후 10 주 이상35.
- 비강 세포 수집 배지 (섹션 1)를 준비하고 튜브를 얼음 위에 두십시오.
- 참가자에게 불편한 절차를 설명하십시오. 칫솔질하는 동안 콧구멍에 완전한 감각이 느껴지며, 이는 바다/수영장으로 뛰어들고 물이 비강으로 돌진하는 것과 유사하다고 설명합니다. 참가자들에게 절차가 반사로 눈물의 생성을 유도 할 것이라고 조언하십시오.
- 참가자에게 적합한 위치를 평가합니다. 앙와위 자세는 절차 중에 참가자의 머리가 브러시에서 멀어지는 것을 방지하기 때문에 검사 소파를 사용할 수 있는 경우 참가자를 앙와위 자세로 눕힙니다. 또는 참가자를 벽 옆에 앉히고 머리를 뒤로 누를 수 있습니다.
- 비강을 검사하십시오. 중격 편차, 폴립 및 비강에서 브러시의 통과에 영향을 미치고 출혈 위험을 증가시킬 수있는 기타 해부학 적 이상에 유의하십시오.
- 참가자들에게 코를 티슈에 불도록 요청하여 과도한 점액의 코를 청소하십시오.
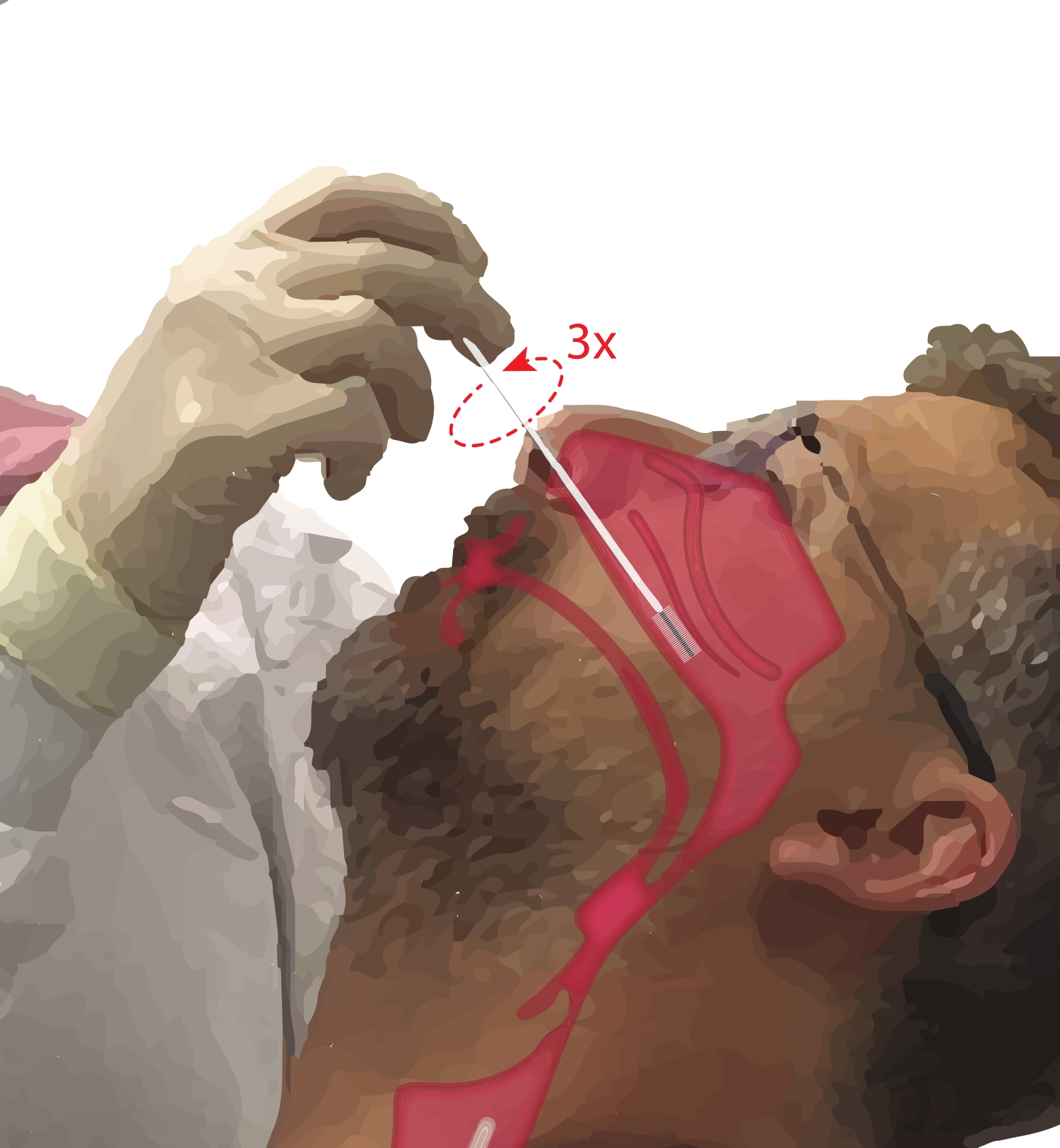
- 참가자에게 입으로 숨을 쉬도록 요청하십시오. 지배적 인 손에 세포학 브러시를 가져 가십시오. 참가자의 턱에 다섯 번째 숫자를 올려 손을 고정하는 동안 세포학 브러시를 참가자의 비강에 삽입합니다(그림 2). 브러시를 참가자의 얼굴에 ~45°로 삽입하여 비강을 통과시킵니다.
- 브러시가 참가자의 얼굴에 수직이 되도록 브러시를 똑바로 회전합니다. 브러시가 하비갑개의 중간에서 뒤쪽 부분이 될 때까지 하비갑개 아래 코의 측면 벽에 대해 부드럽지만 단단히 전진합니다.
알림: 과도한 삽입을 피하십시오. 갑작스런 저항 저하가 느껴지면 비강 인두가 들어간 것이고 절차 주의자가 저항을 다시 느낄 때까지 브러시를 수축시켜야합니다. - 브러시를 360° 최대 3회까지 돌립니다. 삽입 동작의 반대로 브러시를 부드럽게 제거하여 셀이 브러시에서 빠지지 않도록 합니다.
- 브러시를 비강 세포 수집 배지가 있는 준비된 수집 튜브에 넣습니다. 수집 튜브를 얼음 위에 놓습니다.
- 참가자가 동의하거나 많은 수의 세포가 필요한 경우 (예 : 세포 배양을 시작하기 위해) 두 번째 콧 구멍에서 칫솔질을 반복하십시오.
알림: 브러시에 혈액 세포가 보이지 않으면 동일한 콧 구멍을 다시 닦을 수 있지만 동일한 콧 구멍에서 두 번째 칫솔질을하면 출혈 위험이 약간 증가합니다.

그림 2: 비강 상피 세포의 수집. 하부 비갑개의 중간에서 뒤쪽 부분에있는 세포학 브러시의 위치 그림. 이 위치는 콧구멍을 통해 브러시를 삽입하고 브러시를 얼굴에 대해 90° 각도로 회전시킨 다음 브러시를 하비갑개 아래의 비강을 따라 안내하여 도달합니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
3. 기도 상피시트의 제조
참고: 프로토콜의 이 섹션에는 수집 튜브(세포학 브러시 + 비강 세포 수집 배지 1mL)(섹션 2) 및 96웰 평평한 바닥 플레이트가 필요합니다. 기도 상피 시트를 영상화할 목적으로 비비갑개 칫솔질을 수집하는 경우 항생제가 없는 비강 세포 수집 배지 1mL만 사용하십시오. 그렇지 않으면 상피 시트가 이미징을 위해 너무 분산됩니다.
- 세포학 브러시가 포함된 수집 튜브를 부드럽게 소용돌이 치며 브러시에서 기도 상피 시트를 제거합니다.
- P1000 피펫으로 모든 배지와 세포를 수집합니다. 5-6 방울을 96 웰 평평한 바닥 플레이트의 우물에 분배하십시오. 약 7 개의 우물에 대해 반복하십시오.
- 7.1.4단계에 따라 플레이트를 현미경으로 옮기고 섹션 7의 나머지 부분에 따라 섬모 박동 빈도를 이미지화합니다.
- 상피 시트(그림 1)는 섬모 기능이 상피 시트와 단일 부착되지 않은 세포 간에 다르다는 것이 입증되었기 때문에 단일 부착되지 않은 세포가 아닙니다(그림 1).
4. 기도 상피세포 확장 및 유지
- 기도 상피 조건부 재 프로그래밍 세포 확장 배양
참고: 콜라겐 용액 코팅 용기(섹션 1), 조사된 마우스 배아 공급 세포(NIH-3T3), 조건부 재프로그래밍 세포(CRC) 배지(섹션 1), 비강 세포 수집 배지의 세포학 브러시(섹션 2).- 기도 상피 세포와의 공동 배양 전에 최소 2시간 및 72시간 이하의 파종 밀도로 준비된 콜라겐 용액 코팅 배양 용기에 조사된 영양세포를 플레이트화합니다(영양세포 배양 및 조사에 대해서는36 참조).
- 수집 튜브(세포학 브러시) + 비강 세포 수집 배지)의 브러시 세포를 얼음 위의 와류로 옮깁니다. 저속에서 와류 튜브를 10초 켜고, 10초 끄고(그 사이에 얼음 위에 유지) 브러시에서 세포를 제거합니다. 격렬한 소용돌이는 세포 생존력을 감소시킬 수 있습니다. 브러시를 검사하여 점액이 여전히 부착되어 있는지 확인하십시오. 그렇다면 소용돌이를 반복하십시오.
- 얼음 위의 튜브를 생물 안전 캐비닛으로 다시 옮깁니다. 혈청학적 피펫을 사용하여 수집 튜브에서 새 튜브(튜브 B)로 배지를 옮기고 세포학 브러시를 남깁니다. 튜브 B를 300 ×g에서 4 ° C에서 7 분 동안 원 심 분리합니다.
- 원심 분리기에서 튜브 B를 제거하고 상청액을 폐기하십시오. 점액이 보이면 다른 5mL의 비강 세포 수집 배지로 펠릿을 씻고 다시 원심 분리합니다.
- 1mL의 CRC 배지를 추가하여 튜브 B에 세포 펠릿을 재현탁합니다. 5mL 혈청학적 피펫을 사용하여 50mL 튜브(튜브 C) 위에 놓인 세포 체를 원을 그리며 세포를 통과시킵니다.
- 단일 세포 현탁액을 형성하기 위해 여러 번 반복하십시오. 체 바닥에서 잔류 배지를 수집하여 배지와 통합합니다. 세포 체를 폐기하십시오.
- 5mL 혈청학적 피펫을 사용하여 튜브 C에서 1mL의 배지를 취하여 미세 원심분리 튜브로 옮깁니다.
- 이 세포 현탁액 10μL를 취하여 10μL 트리판 블루로 미리 분취된 미세 원심분리 튜브에 추가합니다. 잘 혼합하고 즉시 자동 세포 계수기를 사용하여 세포 수와 생존력을 기록합니다.
- 기도 상피 세포를 조사된 영양세포가 미리 시딩된 T25 플라스크에 시드합니다.
- 기도 상피 세포 유지 및 해리
참고: CRC 매체는 셀에 추가하기 전에 온도 조절이 가능한 실험실 수조 또는 비드 수조 장치에 넣어 37°C로 예열해야 합니다.- 세포 배양 현미경(4× 대물 렌즈)으로 세포를 정기적으로 확인하여 부착, 오염, 형태 및 합류를 확인합니다.
- CRC 미디어는 이틀에 한 번씩 교체합니다. 재프로그래밍된 세포가 관찰되고(그림 1) 오염이 없으면 항생제를 줄이거나 중단합니다.
- 세포가 90% 컨플루언시에 도달하면, 이중 트립신 방법(32 )을 사용하여 세포를 해리시키고 단계 4.1.8에 설명된 바와 같이 세포 계수를 수행한다(세포 해리 및 동결에 대해서는 보충 파일 2 참조).
5. 기도 상피세포의 파종 및 분화 및 분화 및 분화된 ALI 모델의 유지
- 기도 상피 세포를 투과성 지지체 삽입물에 파종
- 콜라겐 용액 코팅 투과성 지지체 삽입물(섹션 1)을CO2 인큐베이터에서 생물안전 캐비닛으로 옮깁니다. 콜라겐 용액을 흡인하고 버립니다. 750 μL 팽창 배지(무항생제)를 투과성 지지체 삽입물의 기초 구획에 추가합니다.
- 해리 된 세포 또는 얼음에서 해동 된 세포를 생물 안전 캐비닛으로 옮깁니다. 150μL의 200,000-250,000 세포를 시드하는 데 필요한 팽창 배지의 부피를 각 투과성 지지체 삽입물의 정점 구획에 추가합니다.
- 거품을 만들지 않도록주의하십시오. 세포가 균질하고 현탁되어 있는지 확인하기 위해 잘 혼합하십시오. 150 μL의 세포 현탁액을 각 투과성 지지체 삽입물의 정점 쪽에 추가합니다.
- 균질한 세포 현탁액을 유지하기 위해 3개의 투과성 지지체 삽입물마다 시딩한 후 세포를 재현탁시킨다.
- 합류 세포 단층이 형성될 때까지 이틀마다(일반적으로 파종 후 4일까지) 배지를 버리고 실온(RT, 15-25°C)으로 가온된 새로운 팽창 배지를 추가합니다.
- 공기-액체 계면에서기도 상피 세포의 분화
- ALI 배지(무항생제)를 RT(15-25°C)로 따뜻하게 합니다.
- 확장 배지를 제거하고 정점 및 기저 구획 모두에서 분화 배지(ALI)로 변경합니다.
- 물에 잠긴 ALI 배지에서 2일 동안 배양한 후 배지를 흡인하고 버립니다.
- 750μL의 ALI 매체를 기저부에 추가하여 공기-액체 인터페이스를 만듭니다.
참고: 배양 1주일 후 단층이 합류하지 않고 구멍이 여전히 관찰되는 경우 세포가 더 이상 공극 영역으로 확장할 수 있는 능력이 없을 수 있으므로 기도 상피 세포를 폐기하는 것이 좋습니다.
- 차별화된 ALI 모델 유지 및 점액 제거
- 완전 분화 될 때까지 2 일마다 정점 및 기저 매체를 교체하십시오 (21-25 일 후 공기-액체 인터페이스 설정).
- 일주일에 한 번 5.3.3-5.3.4 단계에 따라 정점 쪽에서 점액을 씻으십시오.
- PBS를 RT (15-25 ° C)로 따뜻하게합니다.
- 200μL의 PBS를 정점 구획에 추가합니다. CO2 인큐베이터에서 10 분 동안 배양하십시오. 흡인 장치 또는 피펫을 사용하여 PBS를 제거합니다.
6. 3차원 기도 상피 오가노이드
- 기도 상피 오가노이드 배양을위한 준비
- 24웰 플레이트를CO2 인큐베이터에 넣어 밤새 37°C로 예열합니다.
- 제조업체의 지침에 따라 얼음 위에서 ECM(재료 표) 10mL 바이알을 해동합니다. 동결-해동 주기 횟수를 최소화하기 위해 500μL 분취량(1회 사용)을 준비합니다.
참고: 최상의 배양 결과를 위해 단백질 농도가 >10.5mg/mL인 ECM을 사용하는 것이 좋습니다. 농도가 낮 으면 ECM 돔의 분해가 가속화되고 정점을 향한 바깥쪽으로 오가노이드의 발생이 증가합니다. - 기도 오가노이드 키트(재료 표)를 사용하여 제조업체의 지침에 따라 기도 오가노이드 시드 배지(AOSM) 및 분화 배지(AODM)를 준비합니다.
- 표 2에 따라 기도 오가노이드 기저매를 준비합니다.
| 구성 요소 | 음량 |
| 고급 DMEM / F-12 | 500 밀리리터 |
| 헤페스 | 5 밀리리터 |
| 알라닐글루타민 | 5 밀리리터 |
| 페니실린-스트렙토 마이신 | 5 밀리리터 |
표 2: 기도 유기형 기저 매질의 구성 요소
- 섹션 4.2에서 해리 된기도 상피 세포의 수를 사용하여 10,000 세포의 파종 밀도에서 얼마나 많은 웰을 파종 할 수 있는지 계산하십시오 ( 표 3 참조).
- 웰당 1 x 50 μL의 90% ECM 돔(45 μL의 ECM 및 5 μL의 AOSM)을 생성하는 데 필요한 ECM 및 AOSM의 총 부피를 계산합니다.
참고: 웰당 10,000개 세포의 권장 파종 밀도는 계대 1에서 CRC 확장 비강 상피 세포용입니다. 나중에 계대 세포는 동일한 수의 유기체의 형성을 달성하기 위해 더 높은 파종 밀도를 필요로 할 수 있습니다.
| 우물의 수 | 세포 수 | 돔의 수 | 매트리겔 ECM의 권 | AOSM의 권 |
| 1 | 10,000개 셀 | 1 | 45 μL x 1.1 | 5 μL x 1.1 |
| 2 | 20,000개 셀 | 2 | 90 μL x 1.1 | 10 μL x 1.1 |
| 5 | 50,000개 셀 | 5 | 225 μL x 1.1 | 25 μL x 1.1 |
| ......... | ......... 셀 | ......... | μL x 1.1 | μL x 1.1 |
표 3: ECM 돔에서 기도 상피 세포 파종을 위한 계산
- ECM 돔에 기도 상피 세포 파종
알림: ECM은 >10°C의 온도에서 응고되기 시작하므로 ECM을 항상 얼음 위에 놓고 얼음 위에서 ECM과 관련된 모든 단계를 수행하십시오.- 섹션 4.2에서 해리된 기도 상피 세포를 표 3에 따라 계산된 부피 90% ECM으로 재현탁시킨다.
- 피펫을 웰 바닥에 최대한 가깝게 90° 각도(수직)로 잡고 ECM 세포 현탁액 50μL(기포 생성을 방지하기 위해 첫 번째 스톱까지)를 웰 중앙에 분주합니다. 우물 벽을 만지지 마십시오.
- ECM이 고형화될 때까지 플레이트를 37°C에서 20분 동안 인큐베이션한다. ECM이 응고되는 동안 AOSM을 RT (15-25 ° C)로 따뜻하게하여 추가시 ECM 돔의 재액화 및 분해를 방지합니다.
- 웰 벽에 분주하여 예열된 AOSM 500μL를 각 웰에 추가합니다. 미디어를 ECM 돔에 직접 피펫팅하지 마십시오.
- 4-7 일 동안 2 일마다 미디어를 교체하십시오. 매체를 흡입하려면 플레이트를 45° 각도로 기울이고 ECM 돔에서 멀리 떨어진 우물의 하단 가장자리에서 흡입합니다.
- 4-7일 후 각 웰에 500μL의 AODM(15-25°C)을 추가하여 오가노이드 분화를 시작하고 7일 동안 2일마다 배지를 교체합니다.
- 분화 7일째에 기도 상피 오가노이드 재도금
참고: 기도 상피 오가노이드를 재도금하는 것은 ECM 돔의 가장자리가 2주 배양 기간 동안 점차적으로 분해되기 때문에 필요합니다. 돔 가장자리에 있는 기도 상피 오가노이드는 ECM에 완전히 내장되지 않은 경우 손실되거나(매체로 이탈) 정점을 향한 바깥쪽 방향을 가질 수 있습니다. 재도금 단계는 또한 오가노이드를 성공적으로 형성하지 못하는 셀/파편을 제거하여 ECM 돔을 "청소"합니다.- 각 우물에서 미디어를 흡인하십시오. 500μL의 냉기도 유기물 기저 매체 (이후 기저 매체라고 함)를 각 웰에 추가합니다.
- 이 피펫 팁은 오리피스가 가장 크고 피펫팅 중에 오가노이드가 파열될 가능성을 줄여주기 때문에 P1000 피펫을 사용하십시오. 기포가 생기지 않도록 피펫을 350μL로 조정한 다음 피펫을 위아래로 부드럽게 조정하여 각 웰의 ECM 돔을 방해합니다. 모든 ECM/기본 매체를 15mL 원심분리 튜브에 수집합니다.
- 500μL의 차가운 기초 매체로 각 웰을 헹굽니다. 남은 ECM 및 오가노이드가 포함된 기본 배지를 위와 동일한 15mL 원심분리 튜브에 수집합니다.
- 4 °C에서 5분 동안 300 x g 로 원심분리합니다. 원심분리 후 보이는 세 개의 층((1) 상청액, (2) 세포 파편(플러피)을 포함하는 ECM 및 (3) 오가노이드를 포함하는 펠릿 - 상청액과 ECM 층을 버리고 오가노이드 펠릿을 보존합니다.
- 오가노이드 펠릿에 차가운 기저 배지 1mL를 추가하고 피펫을 위아래로 부드럽게 올려 남아 있는 ECM을 분리합니다. 6mL의 차가운 기저 배지를 튜브에 넣고 부드럽게 혼합합니다.
- 300 × g 에서 4 °C에서 5 분 동안 원심 분리합니다. 상청액을 폐기하십시오.
- 과도한 ECM이 여전히 보이면 6.3.5-6.3.6 단계를 반복하여 다른 세척을 수행하십시오.
- 적절한 부피의 90% ECM(AOSM 대신 AODM 사용)으로 오가노이드 펠릿을 재현탁하여 돔의 50μL당 ~30개의 오가노이드를 플레이트합니다.
- 첫 번째 돔을 도금한 후 세포 배양 현미경(4x 대물 렌즈)으로 오가노이드의 밀도를 확인합니다. 밀도가 너무 높으면 90% ECM을 추가하여 원하는 밀도인 ~30개의 오가노이드를 얻습니다.
- 6.2.3-6.2.4 단계에 따라 ECM을 응고시키고 기저 세포, 섬모 세포 및 잔 세포를 포함하는 안쪽을 향한 의사 층상 상피로 둘러싸인 내강 형성으로 성숙에 도달 할 때까지 (분화 21 일 후) 각 웰에 500 μL의 예열 된 AODM으로 이틀마다 세포를 공급합니다.
참고: 여기에 설명된 기도 상피 오가노이드는 말단으로 분화되어 계대하거나 냉동 보존할 수 없습니다.
7. 이미징 섬모 박동 주파수
참고: 프로토콜의 이 섹션에는 가열 및 습도 환경 챔버가 있는 라이브 셀 이미징 현미경, 빠른 프레임 속도(>100Hz) 과학 카메라, 20x 긴 작동 거리 대물렌즈 및 이미징 소프트웨어가 필요합니다(이 프로토콜에 사용되는 권장 장비는 재료 표 참조).
- 현미경 설정
- 현미경 가열 시스템이 켜져 있고 37°C로 평형을 이루는지 확인합니다. 현미경을 켭니다. CO2/공기 가스 혼합기를 통해 가스를 5%CO2로 조정합니다.
- CO2가 통과하는 습도 모듈 병을 정제수로 채우십시오. 스테이지 상단 컨트롤러를 통해 상대 습도를 85%로 설정하여 물이 가열되고 셀에 가습 공기가 공급되도록 합니다. 챔버를 30 분 동안 평형화하십시오.
- 현미경 플레이트 삽입물을 현미경 홀더에 넣습니다.
- 기도 상피 세포 모델을 인큐베이터로부터 현미경으로 옮기고 샘플을 생리학적 온도로 유지하기 위해 37°C로 평형화된 열 블록 또는 열 비드 상에서 진행한다.
- 기도 상피 세포 모델을 포함하는 배양 플레이트를 현미경 플레이트 삽입물에 넣는다. 현미경 환경 챔버를 닫습니다.
- 샘플이 예열된 37°C, 5%CO2-채워진 현미경 챔버에서 30분 동안 평형을 이루도록 합니다.
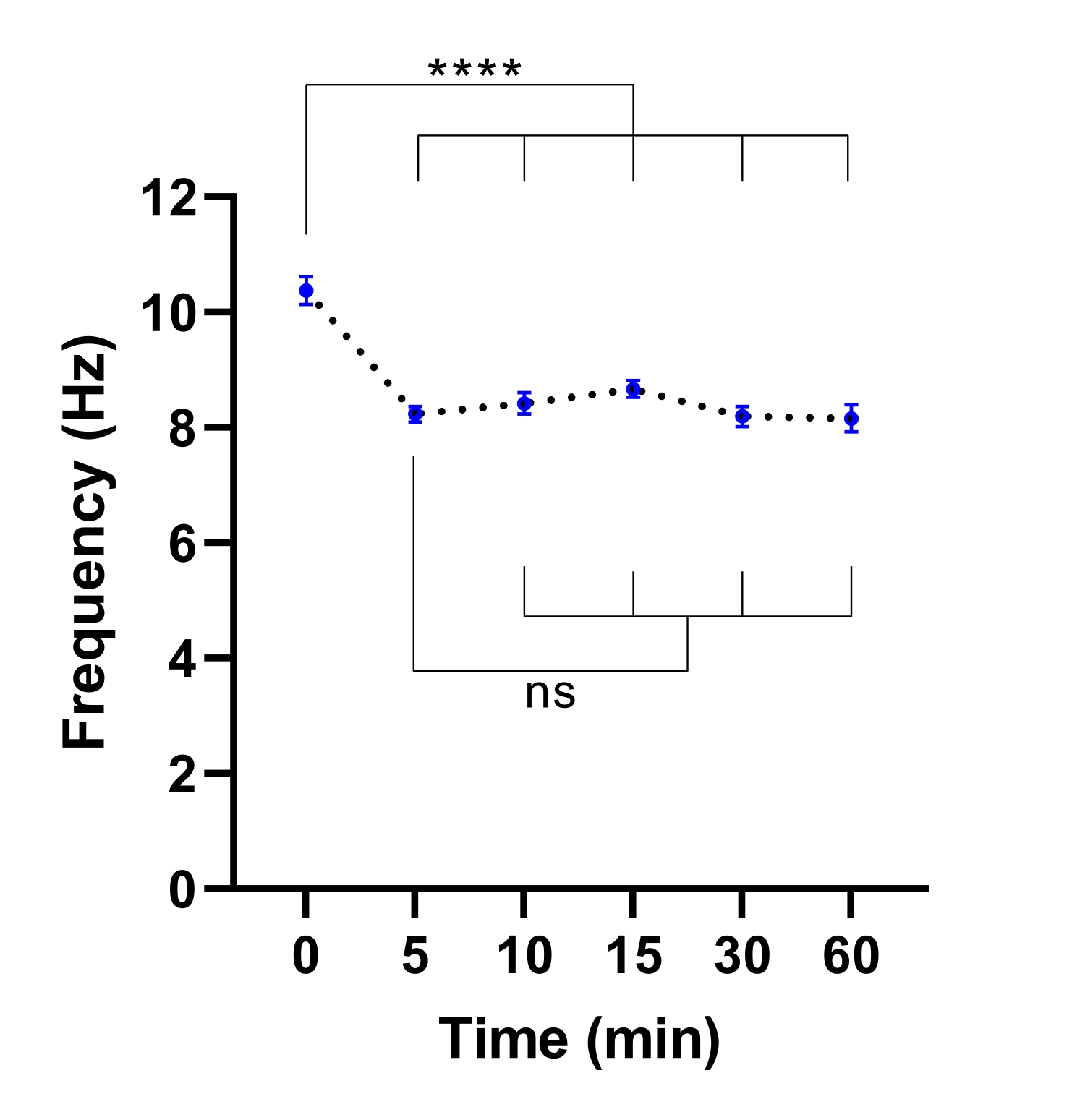
참고: 더 짧은 평형 시간으로 충분할 수 있습니다. 이는 CBF의 안정화에 필요한 시간을 확인하기 위해 실험을 수행하여 결정할 수 있습니다( 도 3 참조).

그림 3: 생세포 이미징 현미경에서 섬모 박동 주파수의 안정화. 공기-액체 계면(ALI 모델)에서 기도 상피 세포의 평균 섬모 박동 빈도(CBF)의 점도 플롯은 환경 챔버가 있는 라이브 셀 이미징 현미경으로 전송한 후입니다. 챔버를 평형화시키고, 챔버 도어를 개방하고 배양 플레이트를 현미경 플레이트 삽입물에 넣기 전에 30분 동안 37°C, 5%CO2 및 85%의 상대 습도에서 유지하였다. 세포 모델을 표시된 간격으로 60분 동안 이미지화하였다. ALI 모델은 CF를 가진 두 명의 참가자로부터 파생되었으며, ALI 모델당 6개의 시야(FOV) 이미지가 획득되었습니다. 각 점(파란색)은 12-36 FOV 이미지에서 평균 CBF를 나타냅니다. 데이터는 평균± SEM으로 표시되며 평균은 점선으로 연결됩니다. 일원 분산 분석(ANOVA)을 사용하여 통계적 차이를 결정하였다. P < 0.0001, ns: 유의성 없음. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
- 평형 기간 동안 컴퓨터에서 수집 소프트웨어를 엽니 다. 20x 긴 작동 거리 대물 렌즈를 선택하십시오.
- 현미경 접안 렌즈에서 세포 모델에 초점을 맞 춥니 다 (~ Z = 8000 μm).
- 투과 광원 전구 필라멘트가 샘플 평면에 초점을 맞추지 않도록 현미경이 Kohler 조명용으로 설정되어 있는지 확인하여 이미징의 아티팩트를 방지합니다. 이를 위해 7.1.10-7.1.13 단계를 수행하십시오.
- 콘덴서 위의 필드 아이리스 다이어프램을 완전히 닫습니다. 필드 아이리스 다이어프램을 천천히 열고 팔각형 모양이 나타날 때까지 콘덴서를 위/아래로 움직입니다.
- 필드 아이리스 다이어프램이 정렬되지 않은 경우(즉, 팔각형이 시야(FOV)의 중심에 있지 않은 경우) 육각 렌치를 사용하여 중앙에 정렬합니다.
- 필드 아이리스 다이어프램이 정렬되면 콘덴서 초점을 조정하여 팔각형의 초점을 선명하게 만듭니다.
- FOV 내에서 더 이상 볼 수 없을 때까지 필드 아이리스 다이어프램을 엽니다.
- 촬영 소프트웨어를 사용하여 L100 을 클릭하여 광 경로를 카메라가 장착된 포트로 전환합니다. 녹색 재생(실행) 버튼을 클릭하여 소프트웨어를 통해 현미경 FOV를 시각화합니다. 섬모에 초점이 맞춰져 있는지 확인하고 필요한 경우 조정하십시오.
- 획득 소프트웨어를 사용하여 다음 설정으로 현미경을 설정하십시오. 필터: 비어 있음; 콘덴서 : 비어 있음; 형식: 범주화 없음; 노출 시간: 0.003 초; 판독 모드: 롤링 셔터; ROI : 512 × 512 픽셀.
알림: 노출 시간은 1/노출 시간이 이 주파수의 두 배 이상이어야 하므로 측정해야 하는 최고 주파수를 기준으로 합니다. 예를 들어, 섬모 박동의 최대 생리적 범위 = 30Hz인 경우 1/노출 시간 = 60이고 노출 시간은 0.016초 ≤이어야 합니다. ROI는 카메라 프레임 속도 사양에 따라 다릅니다. 프레임 속도 >100Hz를 캡처하는 ROI를 선택합니다.
- 이미지 획득
- 메뉴에서 타임랩스 이미지를 획득하려면 획득을 클릭한 다음 빠른 타임랩스를 클릭합니다. 팝업 창에서 저장 위치와 파일 이름을 선택합니다. 1000 프레임을 획득합니다.
- 적용을 클릭합니다. 녹색 재생(실행) 버튼을 클릭하여 현미경 FOV에서 섬모를 미리 보고 필요한 경우 Z 초점을 조정합니다. 지금 실행을 클릭하여 빠른 타임랩스를 캡처합니다.
- 빠른 타임랩스가 캡처되면 녹색 재생(실행) 버튼을 클릭하여 현미경 FOV를 시각화합니다. 현미경 조이스틱을 사용하여 X/Y 축을 따라 다른 FOV로 이동합니다.
- 섬모에 초점을 맞추도록 Z 초점을 조정합니다. 지금 실행을 클릭하여 또 다른 빠른 타임랩스를 캡처합니다.
- 7.2.3-7.2.4단계를 반복합니다. ALI 모델 및 기도 오가노이드의 경우 3x 반복 샘플 각각에서 이미지 6x FOV를 사용합니다. 기도 상피 시트의 경우 참가자 당 최소 4x 복제 이미지를 이미지화합니다.
8. CBF의 데이터 분석 및 정량화
- 데이터 분석 준비
참고: 프로토콜의 이 섹션에는 사용자 지정 분석 스크립트(보충 파일 3), 원시 이미지 파일(섹션 7.2에서 획득), 컴퓨팅 소프트웨어 및 분석 소프트웨어가 필요합니다.- 분석 컴퓨터에 컴퓨팅 소프트웨어(가급적이면 최신 버전)를 설치합니다. 표준 컴퓨팅 소프트웨어 툴박스(elmat, ops, datafun, uitools, datatypes, iofun, iotools, audiovideo)와 이미지 및 신호 처리 툴박스가 설치되어 있는지 확인합니다.
- 사용자 지정 분석 스크립트 'BeatingCiliaBatchOMEfiles_JOVE.m' 및 'LoadRawDataExportFilteredMovies_JOVE.m' 및 '지원 스크립트' 폴더를 컴퓨터의 로컬 드라이브에 복사합니다.
- 컴퓨팅 소프트웨어에서 홈 탭을 클릭합니다. 그런 다음 경로 설정을 클릭합니다(그림 4A-B).
- 팝업 창에서 하위 폴더와 함께 추가를 클릭합니다(그림 4C). 'MATLAB 검색 경로'에서 그림 4D에 표시된 폴더를 선택한 다음 저장 후 닫기를 클릭합니다(그림 4E-F).
- 분석 스크립트가 왼쪽 패널에 나타나는지 확인하여 컴퓨팅 소프트웨어에 연결되어 있는지 확인합니다(그림 4G).
- 섹션 7.2에서 획득한 원시 이미지 파일(개방형 현미경 환경(OME) 형식)을 컴퓨터의 로컬 드라이브로 전송합니다.
참고: Raw 이미지 파일의 예는 https://doi.org/10.6084/m9.figshare.16649878.v1 에서 액세스할 수 있습니다.

그림 4: 데이터 분석을 위한 컴퓨팅 소프트웨어 설정 (ᅡ) 홈 탭을 엽니다. (비) 경로 설정을 선택합니다. (C) 하위 폴더와 함께 추가를 선택합니다. (D) 분석 스크립트가 포함된 폴더를 선택합니다. (E) 저장을 선택합니다. (F) 닫기를 선택합니다. (G) 분석 스크립트가 왼쪽 패널에 나타납니다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
- 단일 픽셀의 강도 스펙트럼의 피크 검출에 의한 CBF의 정량화
- 컴퓨팅 소프트웨어를 엽니다. 'BeatingCiliaBatchOMEfiles_JOVE.m' 분석 스크립트 파일을 클릭합니다(그림 5A).
- 편집기 탭을 클릭한 다음 녹색 재생(실행) 단추를 클릭하여 스크립트를 실행합니다(그림 5B-C). 프롬프트 창에서 분석할 원시 이미지 파일을 선택합니다(그림 5D).
- 7.1.15단계의 노출 시간을 프레임당 획득 시간 프롬프트 창에 입력한 다음 OK(확인 )를 클릭합니다(그림 5E).
- 스크립트가 원시 이미지 파일과 동일한 폴더에 자동으로 저장되는 'AveSpectrum' 파일(보조 파일 4)에서 CBF를 계산하고 출력하는 동안 파일당 ~15분 동안 기다립니다. 진행률 표시줄을 통해 진행률을 시각화합니다(그림 5F).

그림 5: 컴퓨팅 소프트웨어를 사용하여 분석 스크립트 실행 (A) CBF 분석 ( 'BeatingCiliaBatchOMEfiles_JOVE.m') 또는 섬모 박동 영화 ( 'LoadRawDataExportFilteredMovies_JOVE.m')를 만들기 위해 스크립트를 엽니 다. (B) 편집기 탭을 엽니다. (C) 녹색 재생(실행) 버튼을 선택하여 분석 스크립트를 실행합니다. (D) 프롬프트 창에서는 분석 또는 동영상 제작을 위한 파일을 선택해야 합니다. (E) 'BeatingCiliaBatchOMEfiles_JOVE.m'스크립트를 실행하는 동안 파일 읽기 스크립트가 메타 데이터를 제대로 읽지 못하는 경우 프레임 당 수집 시간을 수동으로 입력하라는 메시지가 나타납니다. (F) 계산중인 섬모 박동 빈도를 나타내는 진행률 표시 줄. (G) 'LoadRawDataExportFilteredMovies_JOVE.m' 스크립트를 실행하는 동안 출력할 동영상 유형(mp4 또는 avi), 동영상 프레임 속도(fps), 동영상 데이터에서 움직이지 않는 구성 요소가 제거되었는지 여부('y' 또는 'n'), 프레임 시간(s) 및 동영상으로 내보낸 데이터의 픽셀 크기(미크론)를 수동으로 입력하라는 메시지가 나타납니다. 움직이지 않는 필터링에 'y'를 사용하면 데이터에서 점액이나 기타 방해가 되는 움직이지 않는 레이어가 제거되므로 'y'를 사용하는 것이 좋습니다. (H) 내보낼 동영상을 나타내는 진행률 표시줄. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
- 8.2.1-8.2.2단계의 프로세스를 사용하여 'AveSpectrum' 파일이 포함된 폴더에서 'GetFirstAmplitude.m' 스크립트를 실행합니다. 스크립트가 진폭이 가장 높고 기도 상피 섬모 박동의 생리학적 범위인 ≥3 및 <30Hz 내에 있는 주파수를 포함하는 'FirstAmplitudeStacked.xlsx 파일을 출력할 때까지 기다립니다.
- 'FirstAmplitudeStacked.xlsx 파일에서 주파수 값을 복사하고 과학적 분석 소프트웨어를 사용하여 플롯합니다.
참고: 사용자 지정 분석 스크립트가 CBF를 정량화하는 방법에 대한 설명은 보충 파일 5에 나와 있습니다. 분석된 데이터 세트의 예는 https://doi.org/10.6084/m9.figshare.16649815 에서 액세스할 수 있습니다.
- 섬모 박동 비디오 내보내기
- 컴퓨팅 소프트웨어를 엽니다. 'LoadRawDataExportFilteredMovies_JOVE.m' 스크립트 파일(그림 5A)을 클릭하여 스크립트를 로드합니다.
- 편집기 탭을 클릭한 다음 녹색 재생(실행) 단추를 클릭하여 스크립트를 실행합니다(그림 5C). 프롬프트 창에서 동영상 파일로 내보낼 Raw 이미지 파일을 선택합니다(그림 5D).
- 표 4에 설명된 설정을 '동영상 만들기' 팝업 창에 입력합니다(그림 5G).
- 스크립트가 동영상 파일을 만들고 원시 이미지 파일의 위치로 출력하는 동안 파일당 ~8분 동안 기다립니다. 진행률 표시줄을 통해 진행률을 시각화합니다(그림 5H).
| 동영상 입력 | 묘사 |
| 파일 형식 | 내보낼 파일 형식(mp4 또는 avi)을 입력합니다. |
| 프레임 속도 | 동영상을 내보낼 프레임 속도를 입력합니다. 시계열당 ~1000프레임을 획득한 경우 프레임 속도를 ~30fps로 설정하는 것이 좋습니다. |
| 움직이지 않는 필터링 | 옵션은 'y' 또는 'n'입니다. 기본값은 'y'이고 시간 필터링 스크립트는 푸리에 공간을 사용하여 동영상 데이터에서 움직이지 않는 구성 요소를 제거합니다. 전형적으로, 섬모 또는 움직이지 않는 점액 아래의 임의의 세포 층은 필터링될 수 있는 신호에 제로 주파수 오프셋 성분 또는 시간 불변 성분에 기여할 것이다. |
| 프레임당 획득 시간 | 수집된 데이터의 프레임당 수집 시간입니다. 초 단위로 동영상의 타임 스탬프를 표시하는 데 사용됩니다. |
| 픽셀 크기 | 마이크로미터 단위의 픽셀 크기는 동영상의 비율 막대를 마이크로미터 단위로 표시하는 데 사용됩니다. |
표 4: 동영상 생성을 위한 입력 설정
결과
CBF를 정량화하는 데 있어 이 프로토콜의 효율성을 입증하기 위해, CF를 가진 3명의 참가자와 3명의 건강한 대조군 참가자로부터 유래된 기도 상피 세포 ALI 모델에서 측정된 CBF의 결과가 제시된다. 배양 분화 14일째에 박동섬모가 존재했습니다(그림 6). 배양 분화 14일째부터 21일째까지, 두 코호트 모두에서 통계적으로 유의한(P < 0.0345) CBF 증가가 관찰되었다. 배양 분화 21일차에...
토론
비강 상피 시트에서 CBF의 정량화를 모호하게 할 수있는 여러 요인이 있습니다. 상피 시트는 이 시간 동안 섬모 기능이 가장 안정적이기 때문에 샘플 수집 후 3-9시간 이내에 이미지화되어야 합니다(37). 적혈구와 파편이 적을수록 데이터 수집을 방해하기 때문에 이미징에 가장 적합합니다. 이미징을 위한 ROI를 선택할 때, 이러한 변수가 CBF5에 영향을 미치는 것으?...
공개
저자는 공개 할 것이 없다고 선언합니다.
감사의 말
연구 참가자와 그 가족의 기여에 감사드립니다. 환자 생체 표본의 조직 및 수집에 대한 시드니 아동 병원 (SCH) 랜드 윅 호흡기 부서의 도움에 감사드립니다 - 존 위저 박사, 이본 벨레시스 박사, 리앤 플러시, 아만다 톰슨, 론다 벨에게 특별한 감사를드립니다. 우리는 UNSW 시드니의 Mark Wainwright Analytical Centre 내의 Katharina Gaus Light Microscopy Facility의 Iveta Slapetova와 Renee Wan의 도움을 인정합니다. 이 작업은 NHMRC(National Health and Medical Research Council) Australia(GNT1188987), CF Foundation Australia 및 Sydney Children's Hospital Foundation의 지원을 받습니다. 저자는 Luminesce Alliance - 어린이 건강을위한 혁신의 기여와 지원에 대해 인정하고 싶습니다. Luminesce Alliance - 어린이 건강을위한 혁신은 시드니 어린이 병원 네트워크, 어린이 의학 연구소 및 어린이 암 연구소 간의 비영리 협력 합작 투자입니다. 소아과 연구를 조정하고 통합하기 위해 NSW 정부의 지원으로 설립되었습니다. Luminesce Alliance는 University of Sydney 및 University of New South Wales Sydney와도 제휴하고 있습니다. KMA는 호주 정부 연구 교육 프로그램 장학금의 지원을 받습니다. LKF는 시드니 코브 로타리클럽/시드니 아동 병원 재단과 UNSW 대학교 대학원 장학금의 지원을 받는다.
자료
| Name | Company | Catalog Number | Comments |
| Adenine | Sigma-Aldrich | A2786 | 10 mg/mL |
| Advanced DMEM/F-12 | Thermo Fisher Scientific | 12634-010 | |
| Alanyl-glutamine | Sigma-Aldrich | G8541 | 200 mM |
| Andor Zyla 4.2 sCMOS | Oxford Instruments | Fast frame rate (>100 Hz) scientific camera | |
| Bottle-top vacuum filter system | Sigma-Aldrich | CLS431098 | |
| Ceftazidime hydrate | Sigma-Aldrich | A6987 | 50 mg/mL |
| Cell Culture Microscope | Olympus | CKX53 | |
| CFI S Plan Fluor ELWD 20XC | Nikon Instruments Inc. | MRH08230 | Long working distance objective lens. NA0.45 WD 8.2-6.9 |
| Cholera toxin | Sigma-Aldrich | C8052-1MG | 200 µg/mL |
| Corning Gel Strainer 40 UM | Sigma-Aldrich | CLS431750 | Pore size 40 μm |
| Corning Matrigel Matrix (Phenol red-free) | Corning | 356231 | Extracellular matrix (ECM) |
| Corning bottle-top vacuum filter system | Sigma-Aldrich | CLS431098 | |
| Corning CoolCell LX Cell Freezing Container | Sigma-Aldrich | CLS432002 | |
| Corning Transwell polyester membrane cell culture inserts | Sigma-Aldrich | CLS3470 | Permeable support inserts. 6.5 mm Transwell with 0.4 μm pore polyester membrane insert. |
| Countess Cell Counting Chamber Slides | Thermo Fisher Scientific | C10228 | |
| Countess II Automated Cell Counter | ThermoFisher Scientific | AMQAX1000 | Automated cell counter |
| Cytology brushes | McFarlane Medical | 33009 | |
| DMEM/F12-Ham | Thermo Fisher Scientific | 11330032 | |
| DMEM/F12-Ham | Thermo Fisher Scientific | 11330032 | |
| DMEM-High Glucose | Thermo Fisher Scientific | 11965-092 | |
| Dulbecco′s Phosphate Buffered Saline (PBS) | Sigma-Aldrich | D8537 | |
| Eclipse Ti2-E | Nikon | Live-cell imaging microscope. | |
| Fetal Bovine Serum, certified, heat inactivated, United States | Thermo Fisher Scientific | 10082147 | |
| Fungizone (Amphotericin B) | Thermo Fisher Scientific | 15290018 | 250 µg/mL |
| Gentamicin solution | Sigma-Aldrich | G1397 | 50 mg/mL |
| Graphpad Prism | Graphpad | Scientific analysis software | |
| Greiner Cryo.s vials | Sigma-Aldrich | V3135 | Cryogenic vials |
| HEPES solution | Sigma-Aldrich | H0887 | 1 M |
| HI-FBS | Thermo Fisher Scientific | 10082-147 | |
| Hydrocortisone | Sigma-Aldrich | H0888 | 3.6 mg/mL |
| Incubator NL Ti2 BLACK 2000 | PeCon | Microscope environmental chamber. Allows warm air incubation and local CO2 and O2 gassing | |
| Insulin | Sigma-Aldrich | I2643 | 2 mg/mL |
| Lab Armor 74220 706 Waterless Bead Bath 6L | John Morris Group | 74220 706 | Bead bath |
| Lab Armor Beads | Thermo Fisher Scientific | A1254302 | Thermal beads |
| MATLAB | MathWorks | Computing software | |
| Microsoft Excel | Microscoft | Spreadsheet software | |
| NIH/3T3 | American Type Culture Collection | CRL-1658 | Irradiated NIH-3T3 mouse embryonic feeder cells |
| NIS-Elements AR | Nikon Instruments Inc. | Image acquisition software | |
| Penicillin-Streptomycin | Sigma-Aldrich | P4333 | 10,000 units penicillin and 10 mg streptomycin/mL |
| Dulbecco′s Phosphate Buffered Saline (PBS) | Sigma-Aldrich | D8537 | |
| PneumaCult Airway Organoid Kit | StemCell Technologies | 5060 | Airway Organoid Kit |
| PneumaCult-ALI Medium | StemCell Technologies | 5001 | |
| PneumaCult-Ex Plus Medium | StemCell Technologies | 5040 | |
| PureCol-S | Advanced BioMatrix | 5015 | Type I Collagen solution |
| ReagentPack Subculture Reagents | Lonza | CC-5034 | |
| rhEGF (Epidermal Growth Factor, human) | Sigma-Aldrich | E9644 | 25 µg/mL |
| Y-27632 2HCl (ROCK inhibitor) | Selleckchem | S1049 | 10 mM |
| Tobramycin | Sigma-Aldrich | T4014 | 100 mg/mL |
| Trypan blue solution | Sigma-Aldrich | T8154 | 0.4% |
| UNO Stage Top Incubator | Okolab | Microscope incubator. Allows temperature, humidity and CO2 conditioning |
참고문헌
- Barbato, A., et al. Primary ciliary dyskinesia: a consensus statement on diagnostic and treatment approaches in children. European Respiratory Journal. 34 (6), 1264-1276 (2009).
- Cutting, G. R. Cystic fibrosis genetics: from molecular understanding to clinical application. Nature Reviews Genetics. 16 (1), 45-56 (2015).
- Chioccioli, M., Feriani, L., Kotar, J., Bratcher, P. E., Cicuta, P. Phenotyping ciliary dynamics and coordination in response to CFTR-modulators in Cystic Fibrosis respiratory epithelial cells. Nature Communications. 10 (1), 1763 (2019).
- Hirst, R. A., Rutman, A., Williams, G., O'Callaghan, C. Ciliated air-liquid cultures as an aid to diagnostic testing of primary ciliary dyskinesia. Chest. 138 (6), 1441-1447 (2010).
- Thomas, B., Rutman, A., O'Callaghan, C. Disrupted ciliated epithelium shows slower ciliary beat frequency and increased dyskinesia. European Respiratory Journal. 34 (2), 401-404 (2009).
- Coles, J. L., et al. A revised protocol for culture of airway epithelial cells as a diagnostic tool for primary ciliary dyskinesia. Journal of Clinical Medicine. 9 (11), (2020).
- Pifferi, M., et al. Simplified cell culture method for the diagnosis of atypical primary ciliary dyskinesia. Thorax. 64 (12), 1077-1081 (2009).
- Pifferi, M., et al. Rapid diagnosis of primary ciliary dyskinesia: cell culture and soft computing analysis. European Respiratory Journal. 41 (4), 960-965 (2013).
- Lee, D. D. H., et al. Higher throughput drug screening for rare respiratory diseases: Readthrough therapy in primary ciliary dyskinesia. European Respiratory Journal. 58 (4), 2000455 (2021).
- Saint-Criq, V., et al. Choice of differentiation media significantly impacts cell lineage and response to CFTR modulators in fully differentiated primary cultures of cystic fibrosis human airway epithelial cells. Cells. 9 (9), (2020).
- Awatade, N. T., et al. Significant functional differences in differentiated Conditionally Reprogrammed (CRC)- and Feeder-free Dual SMAD inhibited-expanded human nasal epithelial cells. Journal of Cystic Fibrosis. 20 (2), 364-371 (2021).
- Dabrowski, M., Bukowy-Bieryllo, Z., Jackson, C. L., Zietkiewicz, E. Properties of non-aminoglycoside compounds used to stimulate translational readthrough of PTC mutations in primary ciliary dyskinesia. International Journal of Molecular Sciences. 22 (9), (2021).
- Hirst, R. A., et al. Culture of primary ciliary dyskinesia epithelial cells at air-liquid interface can alter ciliary phenotype but remains a robust and informative diagnostic aid. PloS One. 9 (2), 89675 (2014).
- Marthin, J. K., Stevens, E. M., Larsen, L. A., Christensen, S. T., Nielsen, K. G. Patient-specific three-dimensional explant spheroids derived from human nasal airway epithelium: a simple methodological approach for ex vivo studies of primary ciliary dyskinesia. Cilia. 6, 3 (2017).
- Chilvers, M. A., O'Callaghan, C. Analysis of ciliary beat pattern and beat frequency using digital high speed imaging: comparison with the photomultiplier and photodiode methods. Thorax. 55 (4), 314-317 (2000).
- Chilvers, M. A., Rutman, A., O'Callaghan, C. Functional analysis of cilia and ciliated epithelial ultrastructure in healthy children and young adults. Thorax. 58 (4), 333-338 (2003).
- Castillon, N., et al. Polarized expression of cystic fibrosis transmembrane conductance regulator and associated epithelial proteins during the regeneration of human airway surface epithelium in three-dimensional culture. Laboratory Investigation. 82 (8), 989-998 (2002).
- Jorissen, M., Bessems, A. Normal ciliary beat frequency after ciliogenesis in nasal epithelial cells cultured sequentially as monolayer and in suspension. Acta Oto-Laryngologica. 115 (1), 66-70 (1995).
- Conger, B. T., et al. Comparison of cystic fibrosis transmembrane conductance regulator (CFTR) and ciliary beat frequency activation by the CFTR Modulators Genistein, VRT-532, and UCCF-152 in primary sinonasal epithelial cultures. JAMA Otolaryngology-Head & Neck Surgery. 139 (8), 822-827 (2013).
- Pique, N., De Servi, B. Rhinosectan((R)) spray (containing xyloglucan) on the ciliary function of the nasal respiratory epithelium; results of an in vitro study. Allergy, Asthma & Clinical Immunology. 14, 41 (2018).
- Chen, Q., et al. Host antiviral response suppresses ciliogenesis and motile ciliary functions in the nasal epithelium. Frontiers in Cell and Developmental Biology. 8, 581340 (2020).
- Clary-Meinesz, C. F., Cosson, J., Huitorel, P., Blaive, B. Temperature effect on the ciliary beat frequency of human nasal and tracheal ciliated cells. Biology of the Cell. 76 (3), 335-338 (1992).
- Ballenger, J. J., Orr, M. F. Quantitative measurement of human ciliary activity. Annals of Otology, Rhinology and Laryngology. 72, 31-39 (1963).
- Mercke, U. The influence of varying air humidity on mucociliary activity. Acta Oto-Laryngologica. 79 (1-2), 133-139 (1975).
- Sutto, Z., Conner, G. E., Salathe, M. Regulation of human airway ciliary beat frequency by intracellular pH. Journal of Physiology. 560, 519-532 (2004).
- Salathe, M. Regulation of mammalian ciliary beating. Annual Review of Physiology. 69, 401-422 (2007).
- Kempeneers, C., Seaton, C., Garcia Espinosa, B., Chilvers, M. A. Ciliary functional analysis: Beating a path towards standardization. Pediatric Pulmonology. 54 (10), 1627-1638 (2019).
- Kempeneers, C., Seaton, C., Chilvers, M. A. Variation of ciliary beat pattern in three different beating planes in healthy subjects. Chest. 151 (5), 993-1001 (2017).
- Jackson, C. L., et al. Accuracy of diagnostic testing in primary ciliary dyskinesia. European Respiratory Journal. 47 (3), 837-848 (2016).
- Feriani, L., et al. Assessing the collective dynamics of motile cilia in cultures of human airway cells by multiscale DDM. Biophysical Journal. 113 (1), 109-119 (2017).
- Brewington, J. J., et al. Brushed nasal epithelial cells are a surrogate for bronchial epithelial CFTR studies. JCI Insight. 3 (13), (2018).
- Liu, X., et al. ROCK inhibitor and feeder cells induce the conditional reprogramming of epithelial cells. The American Journal of Pathology. 180 (2), 599-607 (2012).
- Suprynowicz, F. A., et al. Conditionally reprogrammed cells represent a stem-like state of adult epithelial cells. Proceedings of the National Academy of Sciences of the United States of America. 109 (49), 20035-20040 (2012).
- Martinovich, K. M., et al. Conditionally reprogrammed primary airway epithelial cells maintain morphology, lineage and disease specific functional characteristics. Scientific Reports. 7 (1), 17971 (2017).
- Wong, J. Y., Rutman, A., O'Callaghan, C. Recovery of the ciliated epithelium following acute bronchiolitis in infancy. Thorax. 60 (7), 582-587 (2005).
- Gentzsch, M., et al. Pharmacological rescue of conditionally reprogrammed cystic fibrosis bronchial epithelial cells. American Journal of Respiratory Cell and Molecular Biology. 56 (5), 568-574 (2017).
- Sommer, J. U., Gross, S., Hormann, K., Stuck, B. A. Time-dependent changes in nasal ciliary beat frequency. European Archives of Oto-Rhino-Laryngology. 267 (9), 1383-1387 (2010).
- Ratjen, F., et al. Cystic fibrosis. Nature Reviews Disease Primers. 1, 15010 (2015).
- Delmotte, P., Sanderson, M. J. Ciliary beat frequency is maintained at a maximal rate in the small airways of mouse lung slices. American Journal of Respiratory Cell and Molecular Biology. 35 (1), 110-117 (2006).
- Smith, C. M., et al. Cooling of cilia allows functional analysis of the beat pattern for diagnostic testing. Chest. 140 (1), 186-190 (2011).
- Raidt, J., et al. Ciliary beat pattern and frequency in genetic variants of primary ciliary dyskinesia. European Respiratory Journal. 44 (6), 1579-1588 (2014).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기더 많은 기사 탐색
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유