
Method Article
A técnica de coloração diferencial Tempo Juntamente com completa Bilateral Gill denervação para Estudar Ionocytes no Peixe
Neste Artigo
Resumo
This manuscript describes a protocol to track the re-distribution of branchial ionocytes and their innervation using a time differential staining technique coupled with full bilateral gill denervation.
Resumo
Ionocytes branquial (ICs) são as unidades funcionais para regulação iônica em peixes. Em adultos, eles são encontrados na filamentosa e epitélios lamelar das brânquias, onde elas íons de transporte, tais como Na +, Cl - e Ca 2+ através de uma variedade de canais iônicos, bombas e trocadores. O Gill teleost é extrinsecamente inervados pelo facial (VI), glossofaríngeo (IX) e vago (X) nervos. Os nervos IX e X também são a fonte extrínseca de branchial inervação IC. Aqui, duas das técnicas utilizadas para estudar a inervação, proliferação e distribuição de CIs são descritos: a técnica de coloração diferencial de tempo e uma técnica de emalhar desnervação completa bilateral. Resumidamente, peixinho são expostos a um corante específico da mitocôndria vital (por exemplo, MitoTracker Red) que rotula (fluorescência vermelha) CIs pré-existente. Peixes ou foram deixadas recuperar durante 3 - 5 dias, ou imediatamente submetido a uma desnervação brânquia bilateral completa. Depois de 3-5 dias de recuperação, o gmales são colhidas e fixadas para imuno-histoquímica. O tecido é então coradas com um α-5 (anticorpo primário alvos de Na + / K + ATPase que contenha células) em conjunto com um anticorpo secundário que rotula todas (tanto novo e pré-existente) CIs verde. Usando imagem confocal, demonstrou-se que ICs pré-existentes aparecem amarelo (rotulados com tanto um corante específico da mitocôndria viável e α-5) novos ICs e aparecem verde (com apenas α-5). Ambas as técnicas utilizadas em conjunto pode ser aplicado para estudar a inervação, proliferação e distribuição dos componentes no filamento de guelra em que os peixes são expostos aos desafios ambientais.
Introdução
ICs são a unidade funcional para regulação iônica em peixes e são encontradas nas superfícies epiteliais dos filamentos de emalhar e lamelas 4,6-8,10. Embora uma variedade de subtipos foram descritos que possuem características únicas, muitos dos CIs são caracterizados por uma alta densidade de mitocôndrias (assim, eles são também conhecidos como células ricas em mitocôndria) e / ou uma abundância da enzima Na + / K + ATPase (NKA). Tipicamente, estes CIs casa uma variedade de outras bombas, permutadores de iões e canais envolvidos na regulação de iões (por exemplo, Na + / H +, Na + / Cl - o co-transportador, bomba H +) 2,10,11. A redistribuição e proliferação de ICs como um mecanismo compensatório é central para a manutenção da homeostase de íons particularmente durante o estresse iônico (por exemplo, a exposição à água ion-pobre) 4,8,9.
Este estudo descreve um tempo de coloração diferencial techniQue um para identificar ionocytes recém proliferaram (ICs) nas brânquias dos peixes. Esta técnica é acoplado com uma desnervação completa bilateral dos arcos branquiais. Goldfish (Carassius auratus), as espécies utilizadas nestes estudos, é bem adequado para estudar a proliferação de células epiteliais de emalhar, porque eles têm uma notável capacidade de remodelar estruturalmente suas brânquias 2. Gill refere-se a remodelação do crescimento ou de retracção de uma massa celular interlamelar (ILCM) quando o peixe (normalmente mantida entre 15 - 30 ° C) são aclimatados a água fria (<15 ° C) ou hipoxia, respectivamente 3. Estudos anteriores utilizando a técnica de coloração diferencial tempo em peixe têm-se centrado na redistribuição, inervação e proliferação de componentes no brânquia no contexto da brânquia remodelação 4,5. Katoh e Kaneko 1 desenvolveu esta nova técnica para estudar a transformação e substituição de ICs branquial em killifish (heteroclitus Fundulus) transferred da água do mar (SW) para água fresca (FW). Neste estudo, o foco é sobre a proliferação e inervação dos ICs em peixinho aclimatados a 25 ° C.
Usando a técnica de coloração diferencial de tempo, foi demonstrado que, no contexto da remodelação branquial, peixinho manter um número constante de CIs durante a exposição hipóxica e subsequente recuperação normóxico, no entanto, a percentagem de células inervados diminuiu ao longo do período de recuperação de 5 normóxicas. Propôs-se mais de 70 anos atrás, que os mecanismos de captação iônicos em peixes estão sob controle neural 12. O Gill teleost é inervado pelo facial (VII), glossofaríngeo (IX) e vago (X), os nervos também referidos como os "nervos" branchial 13,14. Estudos realizados por Jonz e Nurse (2003) no peixe-zebra (Danio rerio) inervação Gill mostrou que a origem da inervação é extrínseca (corpo celular da fibra nervosa é extrínseca ao Gill), bem como intrínseca (corpo celularfibra nervosa é intrínseco à Gill). Os mesmos autores também demonstraram que ICs branquiais são extrinsecamente inervados 7.
Neste estudo, o tempo de técnica de coloração diferencial acoplado com total denervação Gill bilateral foi utilizado para investigar a proliferação de ICs falta inervação extrínseca em peixinho. Completa desnervação Gill bilateral refere-se a cortar os nervos cranianos IX e X. Essas duas abordagens são viáveis em peixinho porque seu tamanho relativamente grande (30-200 g) simplifica os procedimentos cirúrgicos delicados, e ionocytes são prontamente identificados utilizando técnicas imuno-histoquímica padrão. No presente estudo, CIs foram visualizadas utilizando um corante vital específicos de mitocôndria (por exemplo, MitoTracker Red) ou um anticorpo primário contra o α-subunidade de Na + / K + -ATPase (α-5; Developmental Studies Hybridoma Bank, Universidade de Iowa, Iowa City IA). Este protocolo prevê um meto simplesd de visualizar e analisar a redistribuição e proliferação de ICs na guelra de peixe.
Protocolo
Ambos os protocolos conformados com as diretrizes do Conselho Canadense de Atenção Animal (CCAC) e foram realizadas com a aprovação da Universidade do Comitê Animal Care Ottawa (Protocolo BL-226).
1. Tempo de coloração diferencial Técnica: Mitochondrion-rico Dye Bath
- Preparar uma solução-mãe de MitoTracker Red mM através da dissolução de 50 ug em 94,0 mL de dimetilsulfóxido (DMSO a 100%). Mantenha a solução estoque no escuro a -20 ° C quando não em uso. Evitar ciclos de congelamento / descongelamento.
- Prepare caixas escuras (3-6 caixas) com um volume máximo de 600 ml. Preencha as caixas com 400 ml de água do sistema (água, os peixes são normalmente realizada em) e coloque uma pedra de ar em cada caixa para proporcionar uma fonte de O 2. Obter peixinho (30 - 40 g) e colocá-los nas caixas com 400 ml de água e uma pedra de ar. Após 30 min, adicionar o corante rico em mitocôndria viável para se obter concentrações finais de 0,1 ^ M e 0,01% de DMSO. Banhe os peixes para 4 hr.
- Se estes são os peixes de controle (ou seja, sem a desnervação), ligar o fluxo de água para as caixas, permitir que o corante para lavar e recuperar o peixe para o período de tempo previsto no protocolo. Os peixes são tipicamente recuperado para 3-5 dias.
- Após o período de recuperação prossiga para a Secção 3: Técnica tempo de coloração diferencial: Imunohistoquímica. Se esses peixes devem ser desnervado vá para a Seção 2: Completa Processo denervação Bilateral.
2. completa Bilateral Procedimento denervação
- Obter um par de estudantes tesoura primavera Vannas (curvas), 1 par de fórceps retas padrão standard, 1 par de fórceps curvas padrão standard, 2 pares de No. 5 fórceps, 1 par de afastador de tecidos, pequenas bolas de algodão (1 - 2 mm de diâmetro) e cotonetes (Q dicas ou equivalente).
- Prepare o banho de água anestésico. Em primeiro lugar, dissolve-se 10 g de benzocaína para um volume final de 100 mlde 99% de etanol para preparar uma solução estoque. Para preparar o banho de água anestésico, dissolver 15 ml da solução de estoque benzocaína em 30 L de água do sistema arejada à temperatura requerida. Arejar a água, colocando um ar-pedra conectado a uma bomba de ar ou uma linha central de ar no reservatório de água.
- Coloque o peixe na água do banho anestésico (Figura 1A).
- Uma vez que a respiração cessa, coloque o peixe sobre uma mesa de cirurgia e entubar-lo como mostrado na Figura 1B. Faça isso através da inserção de um tubo em sua cavidade bucal para irrigar as brânquias com solução anestésica aerado. Irrigação das brânquias garante que o peixe é fornecido com uma quantidade suficiente de O 2 e anestésico durante o procedimento de desnervação. Coloque o peixe de modo que a cabeça está inclinada levemente para baixo. Isto permite um melhor acesso ao espaço atrás do quarto arco branquial.
- Levante cuidadosamente o opérculo com a pinça padrão standard retase colocar os afastadores de tecido entre o opérculo e o interior da cabeça. Abra cuidadosamente os afastadores de tecido para manter o opérculo longe da cabeça e manter as brânquias expostas.
- Certifique-se de que a solução anestésica é irrigar as brânquias durante este procedimento. Descanse o afastador lida ao lado da cabeça que dá acesso a todos os quatro arcos de emalhar.
- Coloque a pinça padrão standard curvas entre o quarto arco branquial e parte de trás da cabeça e gentilmente abri-los para criar uma tensão no ligamento prender o quarto arco branquial na cabeça. Com um par de fórceps No. 5 criar uma pequena abertura (2-3 mm), perfurando o epitélio a ligação da extremidade dorsal dos arcos de emalhar para a cavidade do opérculo. Tenha cuidado para não ir muito fundo, porque há o risco de danificar um grande vaso sanguíneo.
- Com uma bola de algodão pequeno realizada com No. 5 fórceps devagar e com cuidado expandir a incisão para expor a IX (glossofaríngeo) e X (vago) nervos. Livrenervos de qualquer tecido conjuntivo usando o No. 5 fórceps, novamente tomando cuidado para não danificar os vasos sanguíneos.
NOTA: O IX branquial e X nervos do peixinho descanso profundo por trás do quarto arco branquial e estão em proximidade com os grandes vasos sanguíneos que alimentam os arcos de emalhar. - Uma vez que os nervos foram identificados Use a tesoura primavera curvas para cortar cuidadosamente os nervos enquanto segura a incisão aberta com as curvas forceps forma padrão. Depois de cortar os nervos, retrair delicadamente as pinças curvas. Não há necessidade de fechar a incisão com suturas porque o epitélio é muito fina e a incisão normalmente fecha por si própria dentro de 24 - 48 h. Remover afastador de tecido.
- Repetir o mesmo procedimento no outro lado da cabeça.
- Alterne a irrigação das brânquias de anestésico a água gasosa fresca para recuperar o peixe da anestesia. Uma vez que os movimentos operculares retomaram mover o peixe em tanques experimentais para se recuperar por pelo menos 24 horas.
- Executar um procedimento de "farsa" em um conjunto separado de peixe. O procedimento de "sham" envolve a perfuração do epitélio atrás do quarto arco branquial, sem cortar os nervos.
3. Tempo de coloração diferencial Técnica: Imunohistoquímica
- Em primeiro lugar, preparar 4% de paraformaldeído (PFA) em 1x tampão fosfato salino (PBS; 4 g PFA em 96 ml de PBS). O PFA não se dissolvem facilmente em PBS à temperatura ambiente. Solução de calor em banho-maria para dissolver PFA. Realize esta em um exaustor. Uma vez PFA está em solução deixe esfriar antes de usar. Armazenar a 4 ° C por até 2 semanas.
- Antes da eutanásia de peixe e extraindo o tecido branquial, coloque 3 - 4 ml de PFA a 4% para um frasco de cintilação para um total de 8 frascos de cintilação (1 frasco) por arco branquial. Além disso, tomar um pequeno barco pesar e preenchê-lo com 1x PBS. Isto irá ser utilizado para lavar o tecido depois de ter sido excisada. Mantenha todas as soluções em gelo.
- Após a exposição de corante rico em mitocôndrias viávelexperimento terminou, eutanásia do peixinho, colocando-o em banho-maria com uma overdose de benzocaína.
- Use uma pinça rombas para levantar o opérculo de um lado da cabeça e tesoura curva para cortar cada extremidade do cesto branquial branquial. Com cuidado, pegue as brânquias pelos rakers usando uma pinça sem corte e levantá-los para fora da cavidade opercular. Lavar imediatamente as brânquias no gelo frio 1x PBS para remover o excesso de benzocaína e sangue.
- Coloque as brânquias excisadas em frascos separados (um frasco para cada arco branquial) cheios de 4% PFA e corrigir O / N a 4 ° C.
- Após a fixação, lavar o excesso de PFA em PBS 1X e colocar o tecido num tubo de bala 2 ml cheia com 1,5 ml de 1% de Triton-X num agitador durante 6 horas à temperatura ambiente ou O / N a 4 ° C. Este passo permeabiliza o tecido. Se toda a guelra é grande demais para colocar em um tubo de 2 ml, em seguida, cortar o tecido em seções pequenas o suficiente para caber o tubo tomando cuidado para não danificar os filamentos.
- Prepare as diluições de anticorpos primários por mixing 4 mL da solução stock de cada anticorpo primário (total de 8 ul) em 992 ul de 1x PBS. Certifique-se de que a NKA (etiquetas células NKA-ricos) e Zn-12 (etiquetas neurônios) anticorpos primários foram levantadas no mesmo host. Isto é importante se os pesquisadores decidir identificar subtipos IC específicas, ao mesmo tempo em que eles vão ter que usar anticorpos primários e secundários criados em uma espécie hospedeira diferentes.
- Remover a solução de Triton-X e lavagem do tecido, sem adicionar um 1: 250 de diluição (dilua em PBS 1x) de um anticorpo monoclonal de NKA (α-5) para detectar as células-NKA ricas e um anticorpo específico neuronal peixe-zebra (zn-12) para detectar as fibras nervosas (anticorpos primários) e incubar num agitador durante 6 horas à temperatura ambiente ou O / N a 4 ° C.
- Lavar o anticorpo primário 3 vezes durante 3 minutos cada uma usando PBS 1x. Para fazer isso, remover a solução de anticorpo primário a partir do tubo de aspiração para fora usando uma pipeta.
- Prepare o anticorpo secundário, a um1: 200 de diluição por mistura de 5 uL de anticorpo secundário estoque em 995 ul de 1x PBS. Aplicar anticorpo secundário (com Alexa Fluor 488) e incuba-se durante 6 h à TA ou O / N a 4 ° C num agitador.
NOTA: MitoTracker Red está animado com a ~ 594 nm de comprimento de onda e fluorescência vermelha. Um anticorpo secundário que está animado com a ~ 488 nm de comprimento de onda e verde fluorescente deve ser usado para rotular NKA e Zn-12 anticorpos primários. - Remover o excesso de anticorpo secundário por lavagem do tecido 3 vezes durante 5 minutos cada (como descrito no passo 3.7).
4. Criação de Imagens
- Após as lavagens, montar o tecido sobre uma lâmina côncava para toda montar imagem confocal de células e fibras nervosas. Para montar o tecido, primeiro colocá-lo em uma gota (200 ul) de 1x PBS numa lâmina de microscópio plana. Isso garante que o tecido não desidratar.
- Separe os hemibranchs emalhar com o micro-tesoura curva. Coloque uma gota de 1x PBS e uma gota de meio de montagem em um slide côncava.
- Coloque as hemibranchs separados no slide côncavo com a borda do filamento voltada para cima e cubra com uma tampa de deslizamento. Umedeça as bordas da lamínula com unha polonês, a fim de evitar que a tampa de deslizamento de mover-se ao redor e deslocar o tecido. Permitir que o tecido se depositem no fundo da corrediça côncava para 10 - 15 minutos antes da imagiologia.
- Para cada arco branquial, selecione seis filamentos de emalhar de forma aleatória para geração de imagens, produzindo seis imagens por arco branquial. Use microscopia confocal convencional para a imagem do tecido, tendo 1 - 3 mm fatias ópticos.
NOTA: Todas as células pré-existentes serão marcados com MitoTracker Red e positivo para NKA aparecerá apenas amarelo. As células que só aparecem em vermelho são ICs pré-existentes que não contêm NKA. Qualquer célula recém-proliferaram só será positivo para NKA e aparece apenas verde. As fibras nervosas também aparecerá verde.
5. Análise de Imagem para Ionocyte Quantificação
- Para each filamento branquial, que foi fotografada, quantificar os ICs e inervação associado percorrendo as seções da pilha Z e contando o número de ICs lamelar e filamentosa presente e se ou não eles são recém-diferenciadas, pré-existente e / ou inervado .
- Quantificar os ICs por filamento ou por área (mm 2) de filamento. Faça isso usando as ferramentas de desenho associados ao software usado para adquirir as imagens para delinear as lamelas do filamento, incluindo a área do filamento em que os ICs foram quantificados. A maioria dos programas de software de imagem confocal tem a opção de calcular a área de uma região delimitada na imagem. Use a opção no software de imagem que permite que você faça isso para adquirir a área.
- Dividir os CIs contados por a área calculada pelo software para obter uma medida de CIs por unidade de área (por exemplo, por 2 mm).
Resultados
A Figura 1 ilustra a mesa de cirurgia criada (Figura 1A), a colocação do peixe durante a cirurgia (Figura 1B) e as três etapas mais importantes para a técnica de coloração de tempo diferencial (Figura 1C). No Passo 1, o peixe é mantido durante 30 min num banho de água bem oxigenada a 25 ° C no escuro. Durante o período de 30 min, o investigador pode preparar o corante alíquota rico em mitocôndria em DMSO que se adiciona à água durante o Passo 2 (Figura 1C). O período de incubação no Passo 2 permite a absorção do corante rico em mitocôndria da água para as células ricas em mitocôndria (isto é, ICs). O peixe pode então submeter ao procedimento completo bilateral desnervação ou um procedimento simulado em que o peixe é anestesiado eo opercula manipulado, mas os nervos permanecem intactos. O criado em Passo 3 representa uma câmara de recuperação fornecida com água corrente durante um peixe que tem either sofrido denervação bilateral completo ou um procedimento de "farsa".
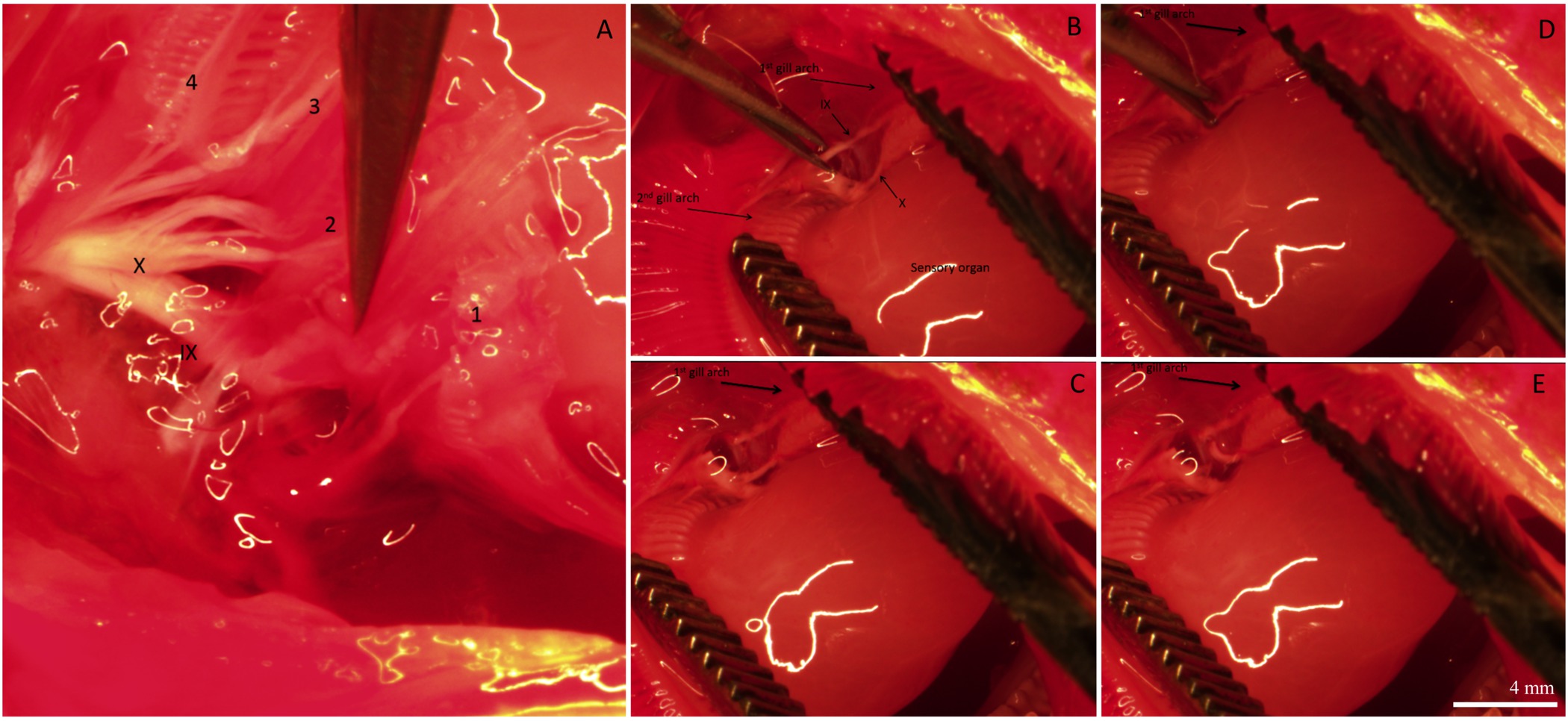
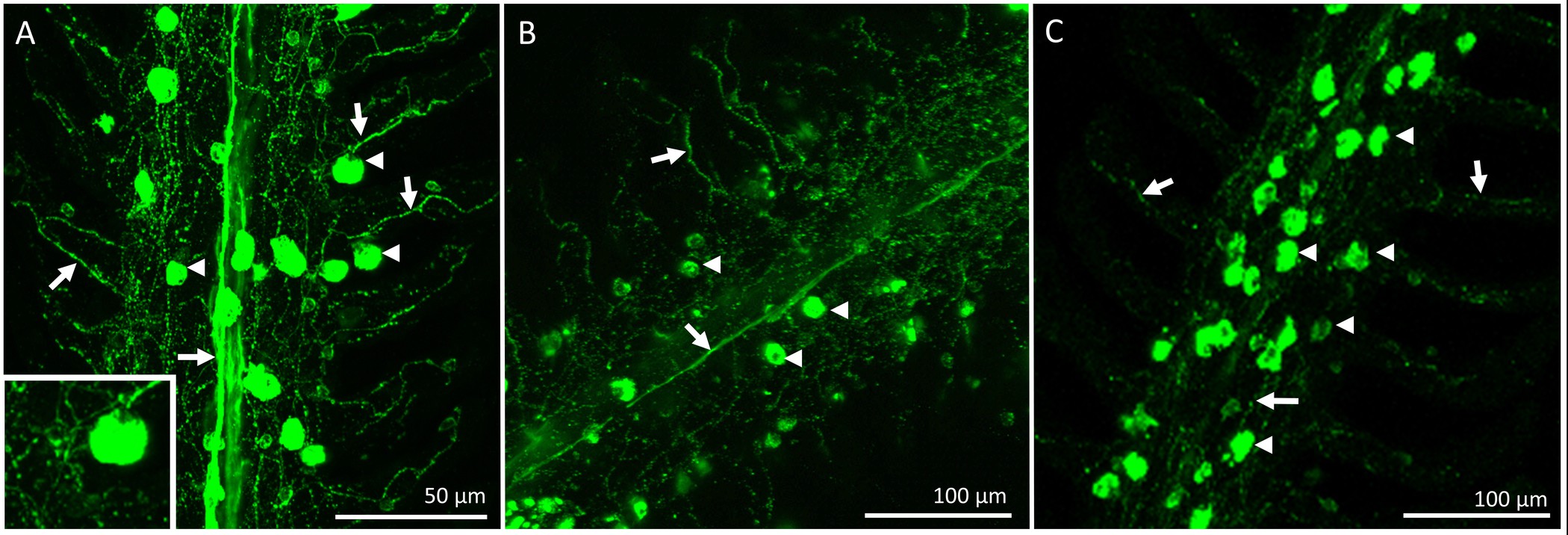
Após o período de recuperação, o peixe foi sacrificado e as brânquias foram excisadas e fixadas para imuno-histoquímica. A distribuição global de inervação e os componentes no filamento de guelra de um peixe que tinham sido submetidos a um procedimento de "sham" está representado na Figura 2. Os CIs estão presentes no epitélio filamentosa, bem como na base das regiões interlamelares. A Figura 2A mostra ICs pré-existentes marcadas com corante rico em mitocôndrias (ou seja, esses ICs existia antes dos procedimentos desnervação / sham foram realizados). A figura 2B mostra fibras nervosas que inervam os ICs pré-existentes e recém-formados (identificado por NKA immunoreactivity) do filamentosa e epitélios lamelar. Por último, a fusão das duas imagens (Figura 2C) revela claramente os ICs pré-existentes (coloração amarela) e os novos ICs (aparecem em verde).A Figura 2D é um gráfico representativo do IC para quantificação do filamento representados na Figura 2A-C. Nesta filamento específico, parece haver um maior número de ICs recém proliferaram por mm2 de CIs pré-existente (N = 1). Para remover a fonte da inervação extrínseca branquial, os nervos cranianos IX e X (inervação extrínseca) foram cortadas. A Figura 3A mostra a região dorsal da cavidade do opérculo depois os filamentos branquiais e os epitélios que cobrem os nervos foram removidos para expor o IX e X nervos cranianos (indicadas com algarismos romanos), que se estendem a partir de dois grandes troncos nervosos para inervar todos os quatro arcos (Figura 3A); os arcos de emalhar são numeradas de 1 a 4. A Figura 3B-E ilustram denervação seletiva do primeiro arco branquial. O primeiro arco branquial é inervado por tanto os nervos cranianos IX e X (Figura 3B), que podem ser removidos sem afectar o resto da inervação para of os arcos de emalhar (Figura 3C-E). Cheio resultados denervação bilaterais em perda gradual da inervação extrínseca aos filamentos de emalhar (Figuras 4A-C). Peixes de controlo apresentam um feixe de nervos óbvio que mede o comprimento do filamento (Figura 4A). Desnervação bilateral completa resultou em alguma perda da inervação extrínseca após 2 dias de recuperação (Figura 4B). Além disso desaparecimento da inervação extrínseca foi observada após 5 dias de recuperação de desnervação (Figura 4C). Qualquer inervação restante para CIs após 5 dias de recuperação foi provavelmente derivada a partir de nervos com corpos celulares dentro do filamento branquial (inervação intrínseca; Figura 4C).

Figura 1. Experimental estabelecido para o procedimento de desnervação e seqüência de passos utilizados notempo técnica de coloração diferencial. (A) a tabela Cirurgia configurado para procedimento de desnervação mostrando os tanques de anestésicos e recuperação. (B) Exemplo da colocação de um anestesiados, peixes entubado. (C) etapas-chave do momento técnica de coloração diferencial. No Passo 1, o peixe é colocado de uma forma controlada, banho de água arejada estático temperatura durante 30 min. O corante rico em mitocôndria é adicionado na etapa 2 até uma concentração final de 0,1 uM e o peixe é permitido tomar banho na solução durante um mínimo de 4 h. O fluxo de água é reiniciado na Etapa 3, altura em que o peixe é submetido a qualquer um denervação bilateral completa do branquial ou uma cirurgia sham. Por favor, clique aqui para ver uma versão maior desta figura.
{kind=link}
 />
/>
Figura 2. Micrografias de luz representativas do tempo técnica de coloração diferencial num peixe (aclimatados a 25 ° C) de 2 dias após a desnervação bilateral completa. (A) A distribuição de ionocytes pré-existentes (ICs; setas). Em um único filamento branquial é revelada por corante coloração rico em mitocôndrias (B) A distribuição dos ICs (novas e pré-existentes) e branquial nervos (setas tracejadas) é revelada por coloração com os anticorpos α-zn-5 e 12, respectivamente. As setas indicam ICs (C) A sobreposição de (A) e (B) distingue ICs pré-existentes. (Coloração amarela; indicadas por setas) de ICs recém proliferaram (aparecem em verde; indicadas pelas setas). Inserção em (C) é uma ampliação de uma pré-existente ionocyte inervados. (D) representativas gráfico de IC quantificação para o filamento mostrado nos painéis (AC). N bar = 1. Escala no painel (C) é de 50 mm e se aplica a todos os painéis."target =" _ /ftp_upload/52548/52548fig2highres.jpg blank "> Clique aqui para ver uma versão maior desta figura.

Figura 3. Imagens representativas que descrevem vários estágios da inervação branquial. View (A) Dorsal da cavidade bucal, mostrando os nervos cranianos IX e X inervam todos os 4 Gill arcos. Arcos Gill são numerados 1-4. Os filamentos de emalhar e do tecido que cobre os nervos foram removidos para melhor visualizar a inervação. (B) O 1º e 2 arcos nd de emalhar são separados para revelar o órgão sensorial e os ramos dos nervos cranianos IX e X que inervam o 1º de emalhar arco. (CE) Sequência de imagens que mostram a denervação seletiva do 1º arco branquial, cortando os ramos dos nervos cranianos IX e X. Para uma re esquemáticaapresentação de emalhar inervação de peixes teleósteos, consulte a Figura 1 em Milsom et al 26 As linhas brancas nas imagens são reflexo da água das luzes de microscópio.; eles não definir qualquer estrutura morfológica. Barra de escala no painel (E) é de 4 mm e aplica-se a todos os painéis. Por favor, clique aqui para ver uma versão maior desta figura.
{kind=link}

Figura 4. micrografias clara que descreve a distribuição e inervação do ionocytes em um único filamento do 1º arco branquial de um peixe dourado aclimatados a 25 ° C. Ionocytes (indicado pela seta cabeças) foram coradas com o α-5 anticorpo e os nervos (indicado bsetas y) foram coradas com o anticorpo zn-12. (A) Gill filamento de um peixe de controlo que mostra um feixe nervoso central, presumivelmente proveniente dos nervos IX e X (inervação extrínseca) com extensiva ramificação lamelar. Alguns dos ionocytes indicados também são inervados (insert). (B) Um filamento branquial dois dias após a denervação bilateral completa. Houve uma redução do feixe nervoso central, enquanto a inervação lamelar apareceu em grande parte intactos. (C) Um filamento branquial 5 dias após a denervação bilateral completa demonstrando que a inervação extrínseca foi praticamente inexistente. A análise qualitativa sugere que a denervação bilateral completa provoca a degradação da inervação extrínseca Gill, mantendo nervos com corpos celulares dentro do filamento branquial (inervação intrínseca) a criação de uma rede de nervos através do filamento e para as lamelas. Por favor,clique aqui para ver uma versão maior desta figura.
{kind=link}
Discussão
O tempo de técnica de coloração diferencial pode ser uma ferramenta útil para compreender a regulação dinâmica de absorção de íons e examinar a redistribuição temporal dos ICs no epitélio branquial. Embora um procedimento simples, existem um número de pontos-chave que são cruciais para o sucesso da técnica de coloração diferencial de tempo. O peixinho deve ser exposto ao corante rico em mitocôndrias para o tempo previsto no protocolo. Exposições mais curtas irá resultar em má absorção do corante pelas células ricas em mitocôndria (ou seja, ionocytes). Durante a fixação, o tecido deve ser brânquia rapidamente excisado e mantidas no escuro para evitar branqueamento foto. O tecido deve ser processado para a imagem latente dentro de 2 semanas de fixação. Durante o procedimento de secção do nervo bilateral garantir que o peixe é bem anestesiado; os nervos e vasos sanguíneos são claramente identificados; e os peixes retoma função opercular completo antes de ser movido para um tanque de recuperação.
"> O ambiente FW apresenta peixe com o duplo desafio de equilibrar as perdas de iões de passivos e ganho de água osmótica 14. O equilíbrio de perdas de íons passiva ocorre através da captação ativa de sal entre os ICs que estão localizadas ao filamentosa e epitélios lamelar onde podem fazer contacto directo com o ambiente externo 2,4,8,9,16. No entanto, a localização dos componentes no brânquia não é estática. Ao longo das últimas três décadas um número de estudos demonstraram que várias espécies de peixes FW, quando confrontados com um desafio iónica e / ou temperatura, redistribuir CIs branquiais do filamento ou base da lamela para as regiões mais afastadas da lamela 1,2,4,5,17-21. Tal redistribuição pode aumentar a espessura das lamelas que pode comprometer a transferência de gás (O 2, CO 2) através do epitélio branquial 22. Os pesquisadores usaram a técnica de coloração diferencial de tempo descrito neste manuscrito para acompanhar a deslocalização e emergence de novos ICs no epitélio branquial, nas condições experimentais diferentes (Figura 1C) 1,4,5.As brânquias e, presumivelmente, os ICs branquial, são inervados pelos IX e X nervos cranianos 7,23-25. Esses nervos transportar tanto eferente e inputs aferentes de e para o Gill, respectivamente. Eles estão localizados no lado dorsal da cavidade bucal por trás do 4º arco branquial. A acessibilidade dos nervos e a facilidade com que o procedimento de desnervação bilateral pode ser realizada é específico da espécie. Em trutas, por exemplo, a anatomia pontiagudo e achatada da cabeça permite que os nervos se encontram num plano atrás do 4º arco branquial sob uma fina camada de tecido. Isso faz com que os nervos visível e de fácil acesso ao pesquisador para realizar o procedimento de desnervação. Em contraste, o peixe dourado tem um focinho mais curto e uma cabeça mais redonda. Os IX e X nervos cranianos de peixinho mentir mais fundo no si dorsalde da cavidade após a 4ª arco branquial ocupando diferentes planos. Esta orientação limita a facilidade de acesso para os nervos e requer uma abordagem mais cuidadosa para identificar e cortar os nervos apropriados. O objectivo do processo é o de remover a desnervação aferente e eferente sensorial entrada e a partir da guelra, respectivamente. A desnervação dos arcos de brânquia também pode ser acoplado com experiências de fluxo de iões utilizando radioisótopos (por exemplo, 22 Na). Estas técnicas podem ser utilizadas em conjunto para estudar a contribuição de entrada nervoso em movimento de iões através do epitélio branquial. Outra limitação com o processo desnervação é a incapacidade de distinguir entre os neurónios sensoriais e motoras, assim quando cortando o feixe de nervos, é possível que ambos os tipos de inervação estão a ser removido. Cortando qualquer neurônios motores podem afetar a guelra e movimento opercular do peixe. Assim, quando a realização de experimentos de denervação de emalhar é importante para monitorar também a ventilação após apeixe tem recuperado do procedimento para garantir que há um movimento de emalhar suficiente para gás e de troca iônica.
Os protocolos descritos neste uso animais adultos manuscrito conservado em um 0:12 luz: ciclo escuro e pelotas de alimentos comerciais alimentados. Estes métodos podem ser modificados de várias maneiras. Em primeiro lugar, o período de recuperação após a exposição corante rico em mitocôndria pode ser ajustado para as exigências do protocolo do pesquisador (por exemplo, 1, 3, 5, ou 14 dias). O mais longo período de recuperação após a exposição corante rico em mitocôndrias no laboratório foi 14 dias 4,5. Não houve uma diminuição significativa na intensidade da fluorescência do corante rico em mitocôndria após 14 dias de recuperação. Em segundo lugar, a utilização de 5 a α-anticorpo primário está limitado a apenas a identificação de células NKA-rico e não distingue entre os diferentes subtipos de CIs branquiais. Felizmente, em peixinho foi estabelecido que a maioria dos ICs são ambos mitochondrion-rico (etiqueta com MitoTracker) e NKA-rich (etiqueta com α-5) células que podem não ser o caso em todas as espécies de peixes 4,27. Experimentos futuros podem se concentrar em seguir a redistribuição temporal do IC subtipos específicos usando anticorpos dirigidos especificamente contra uma variedade de canais, bombas e trocadores (por exemplo, NHE, H + bomba). Estudos anteriores descobriram que a maioria dos ionocytes corados com rico em mitocôndrias corante exposição NKA immunoreactivity 4. Inervação dos CIs pode ser detectada utilizando um anticorpo primário contra o antigénio específico de neurónios derivados de peixe-zebra (zn-12). Neste estudo, α-5 e anticorpos primários foram detectados utilizando o mesmo anticorpo secundário (com Alexa Fluor 488) para ambas 12-zn. Esta limitação é devido a ambos os anticorpos primários ter sido criado num ratinho hospedeiro e é superado pelo fato de que as células-NKA ricas e os neurónios podem ser distinguido morfologicamente mesmo que fluorescem a mesma cor. Pela coloração seqüencialcom diferentes anticorpos secundários (por exemplo, primeiro α-5 com Alexa Fluor 488, em seguida, zn-12 com Alexa Fluor 594) também pode ser usado para eliminar o problema de ter ambos os marcadores fluorescentes da mesma cor. Por fim, o protocolo de secção do nervo bilateral completa pode ser modificado para atingir nervos para arcos de brânquia específicos. Por exemplo, a secção do nervo selectiva pode ser realizada no primeiro arco branquial, separando suavemente os primeiro e segundo arcos de emalhar para expor os nervos que levam ao primeiro arco branquial no final dorsal do carrinho guelras (Figura 2B-E). A técnica de tempo diferencial também pode ser aplicado para estudar a distribuição de ionocytes em peixes-zebra de larvas de peixe como elas se desenvolvem e transições de transporte de iões a partir da pele para o branquial.
Divulgações
The authors declare that they have no competing financial interests.
Agradecimentos
We would like to thank William Fletcher for animal care at the University of Ottawa. The authors would also like to thank Dr. William Milsom for teaching VT the full bilateral denervation technique at the University of British Columbia. A travel grant for this research was provided by the Faculty of Graduate and Postgraduate Studies at the University of Ottawa. This research is also supported by NSERC PGS-D scholarship to VT and Discovery Grants Program to SFP.
Materiais
| Name | Company | Catalog Number | Comments |
| MitoTracker Red | Life Technologies | M-7512 | |
| Dimethyl sulfoxide | Sigma Alrdich | D2650 | |
| α-5 primary antibody | Develompental Hybridoma Bank | a5 | |
| zn12 primary antibody | Develompental Hybridoma Bank | zn12 | |
| Alexa Fluor 488 (anti mouse) | Life Technologies | A-11001 | |
| Benzocaine | Sigma Alrdich | E1501 | 4-aminobenzoic acid ethyl ester, ethyl 4-aminobenzoate |
| Vannas spring scissors | Fine Science Tools | 15000-10 | |
| Standard pattern forceps | Fine Science Tools | 11000-12 | straight |
| Standard pattern forceps | Fine Science Tools | 11001-12 | curved |
| Dumont No. 5 forceps | Fine Science Tools | 11252-30 | |
| Tissue retractor | Fine Science Tools | 17009-07 | |
| Paraformaldehyde | Sigma Alrdich | P6148 | |
| Triton X | Sigma Alrdich | X100 | |
| Vectashield with DAPI | Vector Laboratories | H-1200 |
Referências
- Katoh, F., Kaneko, T. Short-term transformation and long-term replacement of branchial chloride cells in killifish transferred from seawater to freshwater, revealed by morphofunctional observations and a newly established 'time-differential double fluorescent staining' technique. J. Exp. Biol. 206 (22), 4113-4123 (2003).
- Perry, S. F. Relationships between branchial chloride cells and gas transfer in freshwater fish. Comp. Biochem. Physiol. A. Mol. Integr. Physiol. 119 (1), 9-16 (1998).
- Sollid, J., Weber, R. E., Nilsson, G. E. Temperature alters the respiratory surface area of crucian carp Carassius carassius and goldfish Carassius auratus. J. Exp. Biol. 208 (6), 1109-1116 (2005).
- Mitrovic, D., Perry, S. F. The effects of thermally induced gill remodeling on ionocyte distribution and branchial chloride fluxes in goldfish (Carassius auratus). J. Exp. Biol. 212 (6), 843-852 (2009).
- Tzaneva, V., Vadeboncoeur, C., Ting, J., Perry, S. F. Effects of hypoxia-induced gill remodelling on the innervation and distribution of ionocytes in the gill of goldfish, Carassius auratus. J. Comp. Neurol. 522 (1), 118-130 (2014).
- Greco, A. M., Fenwick, J. C., Perry, S. F. The effects of soft-water acclimation on gill structure in the rainbow trout Oncorhynchus mykiss. Cell Tissue Res. 285 (1), 75-82 (1996).
- Jonz, M. G., Nurse, C. A. Epithelial mitochondria-rich cells and associated innervation in adult and developing zebrafish. J. Comp. Neurol. 497 (5), 817-832 (2006).
- Laurent, P., Hebibi, N. Gill morphometry and fish osmoregulation. Can. J. Zool. 67 (12), 3055-3063 (1989).
- Perry, S. F., Laurent, P. Adaptational responses of rainbow trout to lowered external NaCL concentration: contribution of the branchial chloride cell. J. Exp. Biol. 147, 147-168 (1989).
- Evans, D. H., Piermarini, P. M., Choe, K. P. The multifunctional fish gill: dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 85 (1), 97-177 (2005).
- Hwang, P. P., Lee, T. H., Lin, L. Y. Ion regulation in fish gills: recent progress in the cellular and molecular mechanisms. Am. J. Physiol. Regul. Integr. Comp. Physiol. 301 (1), R28-R47 (2011).
- Krogh, A. The Active Uptake of Ions into Cells and Organisms. Proc. Natl. Acad. Sci. U. S. A. 25 (6), 275-277 (1939).
- Nilsson, G. E., Randall, D. J., Hoar, W. S. Innervation and pharmacology of the gills. Fish Physiology. , 185-272 (1984).
- Sundin, L., Nilsson, S. Branchial innervation. J. Exp. Zool. 293 (3), 232-248 (2002).
- Dymowska, A. K., Hwang, P. P., Goss, G. G. Structure and function of ionocytes in the freshwater fish gill). Respir. Physiol. Neurobiol. 184 (3), 282-292 (2012).
- Witters, H., Berckmans, P., Vangenechten, C. Immunolocalization of Na+/K+-ATPase in the gill epithelium of rainbow trout, Oncorhynchus mykiss. Cell Tissue Res. 283 (3), 461-468 (1996).
- Bindon, S., Fenwick, J. C., Perry, S. F. Branchial chloride cell proliferation in the rainbow trout, Onchorhynchus mykiss: implications for gas transfer. Can. J. Zool. 72 (8), 1395-1402 (1994).
- Bradshaw, J. C., Kumai, Y., Perry, S. F. The effects of gill remodeling on transepithelial sodium fluxes and the distribution of presumptive sodium-transporting ionocytes in goldfish (Carassius auratus). J. Comp. Physiol. B. 182 (3), 351-366 (2012).
- Chou, M. Y., et al. Effects of hypothermia on gene expression in zebrafish gills: upregulation in differentiation and function of ionocytes as compensatory responses. J. Exp. Biol. 211 (19), 3077-3084 (2008).
- Kaneko, T., Katoh, F. Functional morphology of chloride cells in killifish Fundulus heteroclitus, a euryhaline teleost with seawater preference. Fisheries Science. 70 (5), 723-733 (2004).
- Ouattara, N., et al. Changes in gill ionocyte morphology and function following transfer from fresh to hypersaline waters in the tilapia Sarotherodon melanotheron. Aquaculture. 290 (1-2), 155-164 (2009).
- Bindon, S., Gilmour, K., Fenwick, J., Perry, S. The effects of branchial chloride cell proliferation on respiratory function in the rainbow trout, Oncorhynchusmykiss. J. Exp. Biol. 197 (1), 47-63 (1994).
- Dunel-Erb, S., Bailly, Y., Laurent, P. Pattern of gill innervation in two teleosts, the perch and the trout. Can. J. Zool. 71, 18-25 (1993).
- Jonz, M. G., Nurse, C. A. New developments on gill innervation: insights from a model vertebrate. J. Exp. Biol. 211 (15), 2371-2378 (2008).
- Jonz, M. G., Zaccone, G. Nervous control of the gills. Acta Histochem. 111 (3), 207-216 (2009).
- Milsom, W. K., Reid, S. G., Rantin, F. T., Sundin, L. Extrabranchial chemoreceptors involved in respiratory reflexes in the neotropical fish, Colossoma macropomum (the tambaqui). J. Exp. Biol. 205 (12), 1765-1774 (2002).
- Hwang, P., Lee, T. New insights into fish ion regulation and mitochondrion-rich cells. Comp Biochem Physiol A Mol Integr Physiol. 148 (3), 479-497 (2007).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados