Method Article
Применение Генетически кодируемые Флуоресцентные окиси азота (NO •) зондов geNOps, в режиме реального времени изображений NO • Сигналы в одиночных камерах
В этой статье
Резюме
This manuscript presents protocols for the application of novel genetically encoded nitric oxide (NO•) probes (geNOps) to monitor single cell NO• fluctuations in real-time using fluorescence microscopy. The Ca2+-triggered NO• formation on the level of individual endothelial cells was visualized by combining geNOps with a chemical Ca2+ sensor.
Аннотация
Оксид азота (NO •) представляет собой небольшой остаток, который опосредует множество важных клеточных функций в организме млекопитающих, бактерий и растений. Несмотря на существование большого количества методов для обнаружения NO • в естественных условиях и в пробирке, мониторинг в режиме реального времени Нет • на уровне одной клетки является весьма сложной задачей. Физиологические или патологические эффекты NO • определяются фактической концентрации и времени выдержки этого радикала. Соответственно, методы, позволяющие выявлять одной ячейки NO • очень желательны. В последнее время мы расширили палитру NO • Индикаторы путем введения одного флуоресцентного белка на основе генетически кодируемых оксида азота (NO) • зонды (geNOps), которые непосредственно реагировать на колебания клеточных NO • и, следовательно, решает эту потребность. Здесь показано использование geNOps для оценки внутриклеточных NO • сигналы в ответ на два различных химических NO • -liberating молекул. Наши результаты ALSO Убедитесь, что свежеприготовленный 3- (2-гидрокси-1-метил-2-nitrosohydrazino) -N-метил-1-пропиламин (ННК-7) имеет гораздо более высокий потенциал, чтобы пробуждать изменение внутриклеточных НЕТ • уровней по сравнению с неорганические NO • нитропруссид донор натрия (SNP). Кроме того, двухцветные изображения живых клеток с использованием зеленых geNOps (G-GENOP) и индикатор химической Са 2+ Fura-2 проводили для визуализации жесткое регулирование Са 2+ -зависимая NO • образование в отдельных эндотелиальных клеток. Эти представительные эксперименты показывают, что geNOps являются подходящими инструментами для расследования генерации в режиме реального времени и деградации одноклеточных NO • сигналы в различных экспериментальных установках.
Введение
Недавно мы разработали новый класс генетически кодируемых флуоресцентных зондов NO •, называют geNOps 1. Эти датчики состоят из просто построены, бактерии-производного, NO • 2 - связывающий домен, который конъюгирован с отчетливым флуоресцентного белка (ФП) вариант 3 (голубой, зеленый или оранжевый FP). НЕТ • привязка к центру 4 негемового железа (II) в пределах geNOps мгновенно уменьшает интенсивность флуоресценции 1. Важно отметить, что geNOps флуоресценция быстро и полностью восстанавливается при внутриклеточных NO • Уровни 1 снижаться. Соответственно, geNOps позволяют в режиме реального времени обработки изображений (суб) клеточных колебаний NO •. Хотя НЕТ • чувствительный механизм geNOps остаются неясными до сих пор, они оказались превосходными NO • репортеры, и, таким образом, имеют потенции, чтобы открыть новую эру полихроматическом, количественного NO • bioimaginг с высоким пространственным и временным разрешением 1, 5. Другие доступные флуоресцентные зонды НЕТ • не основаны на небольших химических соединений, которые должны быть загружены в клетки, и необратимо измененных NO • 6. Дополнительные недостатки НЕТ • чувствительных малых флуорофоров являются их потенциал цитотоксичность и относительно низкая специфичность , которые затрудняют их использовать в надежной, аналитической и убедительным образом 7, 8, 9. Хотя эффективное использование генетически кодируемых флуоресцентных зондов требует эффективных методов переноса генов, FP на основе генетически кодируемые сенсоры появились как незаменимые инструменты , которые произвели революцию в наше понимание внутреннего функционирования клеток 10, 11. До разработки отдельных geNOps FP основе, A Förster резонансной передачи энергии (FRET) не -На NO • датчик, называемый NOA-1 12, был построен. Сато и др. проектировали этот сложный зонд , который состоит из двух субъединиц NO • растворимой , чувствительными к гуанилатциклазы (SGC), оба сопряженными с FRET на основе датчиков отчетности циклического гуанозинмонофосфата (цГМФ) 12 уровней. Поскольку этот датчик реагирует на цГМФ, она лишь косвенно воспринимает внутриклеточных NO • колебания 12. Хотя NOA-1 реагирует на NO • возвышенностей в нано-молярная диапазоне, этот инструмент не часто используется до сих пор, вероятно, из-за ограничений в отношении доступности и практичности этого громоздкого двудольного датчика.
Универсальные функции NO •, что воздействие фундаментальные биологические процессы были хорошо охарактеризованы 13, 14. Многие исследования показали, что NO • concentratioп внутри клеток и субдоменов определяет судьбу клеток в области здравоохранения и болезней , 14, 15, 16. В млекопитающим, НЕТ • в основном генерируется ферментативно в различных типах клеток с помощью семейства 17 хорошо изученным синтазы окиси азота (NOS). До сих пор, три изоформы NOS были описаны 18, 19, 20; они являются Ca 2+ / кальмодулин-зависимой эндотелиальной NOS (Енос или NOS-3) 18 и нейронный NOS (ННО или NOS-1) 19, и Ca 2+ / кальмодулин-независимый конститутивно активный , индуцируемый NOS (иОАС или NOS- 2) 20. Кроме того, существование митохондриальной NOS (mtNOS) также было предложено 21. Тем не менее, mtNOS рассматривается как сплайс вариант нОАС, и поэтому отдельно не классифицирован как изоформы 21. Другой изоформ, за исключением тех , в клетках млекопитающих, является так называемый бактериальный БДУ (bNOS), главным образом, в грам-положительных бактерий 22. Ферментативная производство NO • сильно контролируется и зависит от наличия нескольких кофакторов , таких как никотинамидадениндинуклеотидфосфат (NADPH), флавинадениндинуклеотид (ФАД), тетрагидробиоптерин (BH4), молекулярный кислород и L-аргинин 17. Катионный аминокислота L-аргинин является субстратом , который преобразуется в L-цитруллин на NO • производство 17. В дополнение к высокой регулируемой ферментативной генерации NO •, было постулировано , что радикал может быть уменьшена неферментативно из нитрита бассейнов в митохондриях в условиях гипоксических 23. После того, как НЕТ • продуцируется внутри клетки, он может свободно диффундировать через биомембран 14,исх "> 15. Тем не менее, очень короткий период полураспада этого радикала в основном определяется условиями окружающей среды, а также различные пути и химические реакции , эффективно деградировать NO • Уровни 24. В конце концов, образование, диффузия и деградация NO • зависит по различным экологическим параметрам , которые определяют эффективную концентрацию высоко биологически активной молекулы 24.
Технология geNOps обеспечивает прямое обнаружение (суб) сотовой связи NO • Колебание 1 и поэтому подходит повторно рассмотреть , и вновь открыть для себя механизмы , ответственные за раскачки и разложения клеточных NO • сигналов. Здесь мы приводим простые протоколы и репрезентативные результаты для использования geNOps визуализации экзогенно и эндогенно вызванных сгенерированные NO • профили, на уровне отдельных клеток. Кроме того, технология geNOps может быть адаптирована для арпликация в других модельных системах клеток для изучения сложных моделей образования NO •, диффузии и деградации в ответ на различные клеточные раздражений и стрессов.
протокол
1. Получение химических Буферы и растворы
- Подготовка буфера для хранения , содержащего 135 мМ NaCl, 5 мМ KCl, 2 мМ CaCl 2, 1 мМ MgCl 2, 10 мМ HEPES, 2,6 мМ NaHCO 3, 0,44 мМ KH 2 PO 4, 0,34 мМ Na 2 HPO 4, 10 мМ D- глюкоза, 2 мМ L-глутамина, 1x MEM витамины, 1x MEM аминокислоты, 1% перо Strep и 1% амфотерицин В. Растворить все компоненты в дистиллированной воде и размешать буфере в течение 20 мин с помощью магнитной мешалки при комнатной температуре. Доводят рН до 7,44 с помощью 1 М NaOH. После добавления по каплям раствора NaOH, измерения рН с помощью рН-метра при непрерывном перемешивании.
- Приготовьте физиологический Ca 2+ -содержащие буфер , который состоит из 140 мМ NaCl, 5 мМ KCl, 2 мМ CaCl 2, 1 мМ MgCl 2, 10 мМ D-глюкозы и 10 мМ HEPES. Доводят рН до 7,4 с помощью NaOH, как описано на стадии 1.1.
- Приготовьте Ca 2+ -free буфер , который состоит из одних и тех же ингредиентовкак указано в пункте 1.2. Используйте 1 мМ EGTA вместо 2 мм.
- Солюбилизации фура-2АМ в ДМСО для получения 1 мМ маточного раствора. Алиготе маточного раствора в плотно закрытых флаконах и хранить при температуре -20 ° С в течение до одного месяца. Если заморожен, позволяют маточный раствор уравновешивают при комнатной температуре в течение по меньшей мере 1 ч, защищенном от света месте. Разбавляют исходный раствор Fura-2AM в 1 мл буфера для хранения (этап 1.1), чтобы получить конечную концентрацию 3,3 мкМ.
- Подготовка 1 мл 100 мМ гистамина исходного раствора в дистиллированной воде (рН 7,0). Разбавляют 100 мМ гистамина маточного раствора в 100 мл Ca 2+ -содержащий физиологическом буфере до конечной концентрации 100 мкМ гистамина.
- Солюбилизации Nω-Нитро-L-аргинин (L-NNA) в 100 мл физиологического буфера содержащего кальций, чтобы получить конечную концентрацию 300 мкМ. Хранить решения на C водяной бане 37 ° в течение по крайней мере 1 ч до тех пор, L-NNA полностью не растворится.
- Солюбилизации 10 мг NOC-7 в дистиллированной ватер (рН 7,0) для получения 10 мМ маточного раствора. Алиготе маточного раствора в виде небольших аликвот в плотно закрытых флаконах и хранить непосредственно при -70 ° С. Развести маточного раствора ННК-7 в физиологическом буфере, чтобы получить конечную концентрацию 10 мкМ NOC-7.
Примечание: NOC-7 представляет собой небольшое химическое соединение, которое спонтанно высвобождает NO с коротким периодом полураспада. Всегда подготовить рабочие буферы, которые NOC состоит-7 непосредственно перед каждым экспериментом. - Сделать раствор 1 мМ нитропруссид натрия (SNP) в физиологическом буфере кальция и готовят 10 мкМ разбавление а.
Примечание: Всегда готовьте небольшое количество (~ 10 мл) NO-доноров решений из-за быстрой скорости деградации. - Приготовьте фосфатного буферного раствора (PBS) , состоящий из 137 мМ NaCl, 2,7 мМ KCl, 9,2 мМ Na 2 HPO 4 и 1,5 мМ KH 2 PO 4. Отрегулируйте значение рН до 7,44 с помощью NaOH / HCl.
2. Подготовка сотового
Примечание: в оrder для достижения однородности при выполнении измерения оксида азота (NO) • в одиночных камерах с помощью geNOps, клетки должны быть предварительно инкубировали с железом (II) / витамин С , содержащий буфер перед экспериментами изображений с целью восстановления Fe 2+ в NO • -связывающим домена NO • -probes. В случае EA.hy926 и HEK293, инкубирование в течение 20 мин с раствором бустер железа (II) приводит к полной активации НЕТ • датчиков.
- Семенной 5,5 х 10 5 EA.hy926 или НЕК293 на следующий день или 3,5 х 10 5 клеток на следующий день после следующего дня, на 30-мм покровных стекол микроскопа в лунку 6-луночного планшета. Инкубируйте клетки при 37 ° С в увлажненной среде с 5% CO 2.
Примечание: Подробная информация о аденовирусной инфекции клеток млекопитающих и конструкции вируса было описано Zhang и др. 25. Этот шаг не применяется для клеток НЕК 293, стабильно экспрессирующих G-GENOP. - Возьмем фетальной телячьей сыворотки (FCS) и антибиотиков свободный Eagle модифицированной среде Дульбекко (DMEM), и добавить адено- ассоциированный тип Вирус 5 (AAV5) вектор, несущий ген, кодирующий G-GENOP (MOI: 500 для клеток EA.hy926 дают почти 100 % положительных клеток; MOI: 1 эффективен для клеток НЕК293).
Примечание: Использование аденосателлитные вирусных векторов отнесен к группе риска 2. Уровень биологической безопасности 2 защитной оболочки, как правило, требуется для работы с этим вектором. При необходимости, вирусные инфекции также могут быть выполнены в FCS, содержащие среду. В качестве альтернативы, клетки могут быть трансфицированы с использованием липидной основе несущих 1. - Удалить культуральной среды и промыть клетки с предварительно нагретом (37 ° С) PBS. Добавить 1 мл DMEM / AAV5 среды на каждую лунку в течение 1 часа. Этот шаг не применяется для клеток НЕК 293, стабильно экспрессирующих G-GENOP.
- Добавить 1 мл 20% FCS, содержащей DMEM на каждой лунке до конечной концентрации 10% FCS. Не удаляйте AAV5-содержащий носитель из клеток. джентльменLY качели пластины для гомогенизации среды в лунках. Инкубируйте клетки в течение 48 ч при 37 ° C в увлажненной среде с 5% CO 2. Этот шаг не относится к клеткам НЕК 293, стабильно экспрессирующих G-GENOP.
- Через 48 ч промыть клетки с подогретого PBS. Затем добавляют 2 мл подогретого буфера для хранения (раздел 1.1) в каждую лунку и инкубировать клетки при комнатной комнатной температуре в течение не менее 1 ч, защищенном от светового излучения.
- Замените буфер хранения 1 мл / лунку железа (II) раствором бустера при комнатной температуре (RT). Инкубируйте клетки в течение ровно 20 мин в темноте.
Примечание: Не следует превышать или уменьшить оптимальное время инкубации, так как это может повлиять на отзывчивость NO • зондами. - Промыть клетки один раз буфером для хранения и инкубировать каждую лунку с 2 мл буфера для хранения в течение по меньшей мере 2 ч при комнатной температуре, с тем чтобы позволить клеткам уравновешиваться.
- Заменить буфер хранения с 3,3 мкМ фура-2 утра в буфере для хранения 1 мл в течение 45 мин при комнатной температуре, защищеныот воздействия света.
- Промыть клетки два раза буфером для хранения и инкубировать, еще раз в течение не менее 30 мин, чтобы позволить клеткам уравновешивания.
3. Живая ячейки изображения НЕТ • и Ca 2+ сигналов в отдельных клетках
- Закрепите крышку 30 мм скольжения, покрытую EAhy.926 или HEK293 клетки (со стадии 2.1) в металлической перфузионной камере и поместите его на микроскоп. Подключите наплывом трубку к буферу водохранилищах и оттоку с вакуумным насосом. Обеспечить непрерывный поток и избежать осушение покровным.
- Начало тяжести управляемой перфузию физиологическим буфером кальция (со стадии 1.2), используя полуавтоматическую систему перфузии.
Примечание: Такая система состоит из буферных резервуаров, соответствующей трубки, магнитные клапаны, которые с электронным управлением и вакуумным насосом (смотри рисунок 1В). Скорость потока может находиться в диапазоне от 1 до 3 мл / мин, в зависимости от высоты перфузионных резервуаров. Для получения стабильных местных приме наркотиковКатион, скорость потока всех используемых резервуаров должны быть приблизительно равными. Это должно быть проверено перед экспериментами с изображениями. Учтите, что эндотелиальные клетки отвечают возросшим касательное напряжение, которое может быть вызвано строгой перфузией. - Включите системы формирования изображения и позволяет разогреть всех устройств в течение 30 мин.
- Определение параметров изображения с использованием соответствующего программного обеспечения. Выбор длины волны возбуждения 340 нм и 380 нм для фура-2 изображения и 480 нм для возбуждения G-GENOP. Для того, чтобы свести к минимуму флуоресценции отбеливания увеличить биннинга камеры 4 и уменьшения интенсивности возбуждения и времени экспозиции. Смотрите также шаги 3.6-3.8.
Примечание: настройки и параметры визуализации зависят от используемых устройств, фура-2 эффективность погрузки и уровни экспрессии G-GENOP. - Выберите область изображения, перемещая АБВ стол микроскопа до нескольких флуоресцирующих клеток находятся в фокусе. Затем определите области интереса (трансформирования) с помощью соответствующего программного средства. Draw регионах, охватывающих несколько весь оде флуоресцентные клетки в области формирования изображения вручную. Кроме того, определяют область фона аналогичного размера.
Примечание: После того, как изображения были получены и сохранены трансформирования также могут быть определены заново после процесса формирования изображения для дальнейшего анализа с использованием соответствующего программного обеспечения для анализа изображений (см список материалов). - Начать сбор данных на перевернутом и передовой флуоресцентный микроскоп с моторизованным стадии образца, а также монохроматического источника света. В качестве альтернативы возбуждают при 340 нм и 380 нм для Фура-2АМ и 480 нм для G-GENOP соответственно. Установить соответствующие моменты времени экспозиции, так что для всех каналов, четкий сигнал флуоресценции можно обнаружить в течение долгого времени. Смотрите также шаг 3.4.
ПРИМЕЧАНИЕ: Это зависит от интенсивности возбуждающего света и биннинга камеры. Например, можно использовать 15% интенсивности возбуждающего света, камера биннинга от 4 до 150 мс для 340 нм, 50 мс для 380 нм и 300 мс для 480 нм. Смотрите также шаг 3.4. - Соберите испускаемого света при длине волны 510 нм для фура-2 / ч, 520 нм для G-Генор используя прибор с зарядовой связью (CCD) камера с соответствующим набором фильтров, состоящей из 500 нм возбудителем, в 495 нм дихроичных и 510-520 нм эмиттера. Запись один общий фрейм каждые 3 секунды.
- Запись первых минут (при необходимости до 3 мин) в физиологическом буфере Ca 2+ (1.2) , чтобы получить базовый уровень соответствующих сигналов флуоресценции в течение долгого времени.
- После того, как стабильный базовый уровень флуоресценции наблюдается, перейти на 100 мкМ гистамина или АТФ (1 мкМ или 100 мкМ) , содержащей физиологический Ca 2+ буфера стимулируют клетки в течение 3 мин.
Примечание: EA.hy926 клетки, которые реагируют на агонисты показывают резкое увеличение соотношения фура-2 (при возбуждении флуоресценции при длине волны 340 нм, деленная через флуоресценции возбужденной при 380 нм) и явное уменьшение сигнала флуоресценции G-GENOP. - Вернитесь в физиологическом буфере Ca 2+ без гистамина или АТФ и L-NNA в течение 5 мин для удаления соединений из клеток. Этот шаг не может быть продлен до fluoresИзменения минесценцию восстанавливаются полностью.
- Администрировать 10 мкМ NOC-7 в физиологическом буфере Ca 2+ в течение 2 мин с использованием системы перфузионного. Донор NO сильно влияет на G-GENOP флуоресценции, которые, как правило, уменьшается на> 20% в ответ на 10 мкМ NOC-7. В клетках EA.hy926 эффект ННК-7 примерно в 3 раза сильнее по сравнению с агонистом индуцированной G-GENOP флуоресценция закалочной.
- Промойте NO • -releasing соединение в течение примерно 10 мин с физиологической Ca 2+ буфера и остановить запись , как только базальной флуоресценции восстанавливается.
Анализ 4. Данные
- Экспорт приобрела средней интенсивности флуоресценции данных отдельных клеток в течение долгого времени, чтобы программное обеспечение для анализа данных.
- Вычитание соответствующих фоновых значений и вычислить отношение 340 нм при 380 нм, соответствующих фура-2 сигналов каждой отдельной ячейки с течением времени.
- Вычтите фоновые значения канала G-GENOP, чтобы получить фактическую флuorescence интенсивности NO зонда (F) в течение долгого времени с помощью программного обеспечения расчета.
- Возьмите исходные значения флуоресценции в качестве F 0 (F 0 флуоресценции зонда NO в течение долгого времени без стимуляции). Смотрите также шаг 4.5 и Рисунок 1C.
- Вычислить функцию с течением времени для эффектов флуоресценции отбеливания , используя следующее уравнение: F 0 = F • Начальное ехр (-К • Время) + F плато. F Начальное: максимальная сигнал флуоресценции когда - визуализации запускается; K: константа скорости обесцвечивания флуоресценции с течением времени; F плато: флуоресценция минимальное достигается при отбеливании с течением времени; Смотрите также шаги 4.3- 4.4 и Рисунок 1C.
Примечание: Для приближения, все значения флуоресценции с течением времени до и после стимуляции клеток могут быть использованы. Более подробная информация указаны Bentley и др. 27. - Для нормализации сигнала G-GENOP с течением времени, вычислить 1-F / F0 (шаги 4.3-4.5). Смотрите рисунок 1C и 1D.
Результаты
Визуализация одноклеточные NO • Профили в ответ на скоротечно Applied NO Доноры
Мы использовали клон клеток HEK , которая стабильно экспрессирует G-GENOP (рис 1А), для того , чтобы визуализировать НЕТ • сигналы на одноклеточной уровне в ответ на два различных NO-освободив малых химических соединений, NOC-7 и SNP. В NO • доноры последовательно применяются и удаляются из клеток во время формирования изображения с использованием силы тяжести на основе системы перфузии , что обеспечивается непрерывный поток (рис 1B). Все клетки , экспрессирующие G-GENOP с различной интенсивностью показали явное снижение флуоресценции в ответ на NOC-7 и SNP (рис 1C), что указывает на быстрое NO • накопление внутри клеток при добавлении к NO • доноров. Нормированные сигналы флуоресценции (1-F / F 0) показали , что оба NO • доноры вызывали однородную cellulaг NO • возвышений , что полностью выздоровел после вымывания из -releasing соединений NO • (Рисунок 1D). Тем не менее, 10 мкМ SNP индуцируется только 50% от сотовой связи NO • сигнала (9,63 ± 1,05%, N = 3/38), который был достигнут 10 мкМ NOC-7 (18,10 ± 1,20%, п = 3/38, р < 0,0001). Для достижения равных внутриклеточные уровни NO • как с NO • доноров, требовалась концентрация 1 мМ SNP (рис 1C, 1D).
Далее, мы протестировали способность свежеприготовленной по сравнению с истекшим NOC-7, чтобы поднять внутриклеточных NO • Уровни в НЕК клетках. С этой целью мы подготовили четыре экспериментальных буферов, содержащих 5 мкМ NOC-7. НЕТ • донора либо добавлены свежеприготовленные непосредственно перед измерением, или храниться в резервуарах в течение 1 ч, 2 ч и 3 ч при комнатной температуре перед измерением. Различные буферы были последовательно применены и удаляться пПЗУ клетки , экспрессирующие G-GENOP с помощью перфузионной системы (рис 2B). Такой подход обнародовал стабильность водных NOC-7 решений , которые, как и ожидалось, показал снизилась мощности в течение долгого времени , чтобы поднять внутриклеточные уровни NO • (Фигуры 2А, 2С). Интересно отметить , что НЕТ • сигналы значительно быстрее восстанавливается после удаления истекших буферов, по сравнению с внутриклеточным NO • ответ , что была вызвана свежей NOC-7 (рис 2С), возможно , указывающего слипание нетронутым НЕТ • -liberating молекул на клеточных компонентов ,
Одновременная визуализация Са 2+ и NO • Сигналы в одиночных эндотелиальных клеток
Для изучения Са 2+ не -triggered NO • образование в эндотелиальных клетках, обычно используется эндотелиальной увековечены клеток суррогатным, клеточная линия EA.hy926, была тransiently трансфицируют G-GENOP и нагружали фура-2 / ч (см протокол 2.8). Трансфекция дали примерно 10% G-GENOP положительных эндотелиальных клеток (п = 6, рис 3А), что достаточно для записи Ca 2+ не -evoked NO • производство на уровне отдельных эндотелиальных клеток. Тем не менее, мы достигли почти 100% G-GENOP клетки положительным EA.hy926 с использованием вирусного вектора аденоассоциированный (n = 6; см протокол 2.2). До многоканальных измерений, клетки инкубировали в течение 20 мин при комнатной температуре в буфере для хранения , состоящий L-нна, мощный необратимый ингибитор NOS-26. Контрольные клетки содержали в том же буфере для хранения без L-NNA (см протокола 1.1) (рис 3б). Лечение контрольных клеток гистамина, мощный инозитол 1,4,5-трифосфата (IP 3) -generating агониста, мгновенно повышен цитозольного Ca 2+ уровня с последующим постепенным увеличением внутриклеточного NO • пока агонист не было удалитьD (рис 3C, 3D). Клетки , предварительно обработанные с NOS-ингибиторов показали аналогичные цитозольных сигналы Са 2+, в то время как внутриклеточный NO • уровень остается практически неизменным в ответ на гистамин (рис 3Е). EA.hy926 клетки , экспрессирующие G-GENOP также обрабатывали 1 мкМ и 100 мкМ IP 3 -generating агонистом АТФ для того , чтобы проверить , является ли или нет geNOps подходят для мониторинга за NO • сигналы в ответ на низких , так физиологические и супра-физиологические концентрации агониста (рис 3F). В эндотелиальных клеток линии 1 мкМ АТФ вызывало ясно цитозольного НЕТ • сигнал, который был приблизительно половина сигнала , полученного с помощью 100 мкМ АТФ (рис 3F).

Рисунок 1: Внутриклеточные профили NO в ответ на различные NO-Liberati нг молекул. (A) Широкое поле изображения клеток НЕК , стабильно экспрессирующих цитозольного G-GENOP. Шкала бар = 20 мкм. (Б) Схематическое изображение , основанного на полуавтоматическом перфузионной системы гравитации для контролируемого применения и снятия NOC-7 и SNP. (С) представитель (из 3 независимых экспериментов) ненормированной интенсивности одноклеточного флуоресценции прослеживает в произвольных единицах в зависимости от времени из клеток НЕК , стабильно экспрессирующих цитозольного G-GENOP в ответ на 10 мкМ NOC-7, 10 мкМ СНП и 1 мМ SNP. Black полужирный кривая представляет среднюю кривую 26 одноклеточных следов (светло-серые кривые). Пунктирные черная кривая представляет F 0, который был использован для нормализации. (D) нормировано и перевернутые одиночные следы (1-F / F 0, светло - серые кривые), и средней кривой (черная смелая кривая) с течением времени в ответ на 10 мкМ NOC-7, 10 мкМ SNP и 1 мМ SNP , извлеченные из панель C.эс / ftp_upload / 55486 / 55486fig1large.jpg "целевых =" _blank "> Пожалуйста, нажмите здесь, чтобы посмотреть увеличенную версию этой фигуры.

Рисунок 2: Тест на стабильность ННК-7 с использованием стабильно G-GENOP , выражающие HEK клеток. (А) представитель NO • Кривая концентрации реакция с течением времени стабильно G-GeNOps , выражающих HEK клеток при применении свежих и старых NOC-7 буферных растворов. Все NOC-7, содержащие буферы были первоначально приготовленным с конечной концентрации 5 мкМ, используя один и тот же исходный раствор (50 мМ). Время истечения соответствующих растворов после приготовления до количеств изображений до 1 ч, 2 ч, 3 ч и, как не указано обратное. (Б) Схематическое изображение на основе полуавтомат системы перфузионного тяжести , чтобы последовательно добавлять и удалять NOC-7 , содержащий решения во время съемки. (C) Временные корреляциисотовой связи NO • сигналы в ответ на свежеприготовленную и старые NOC-7 буферов. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
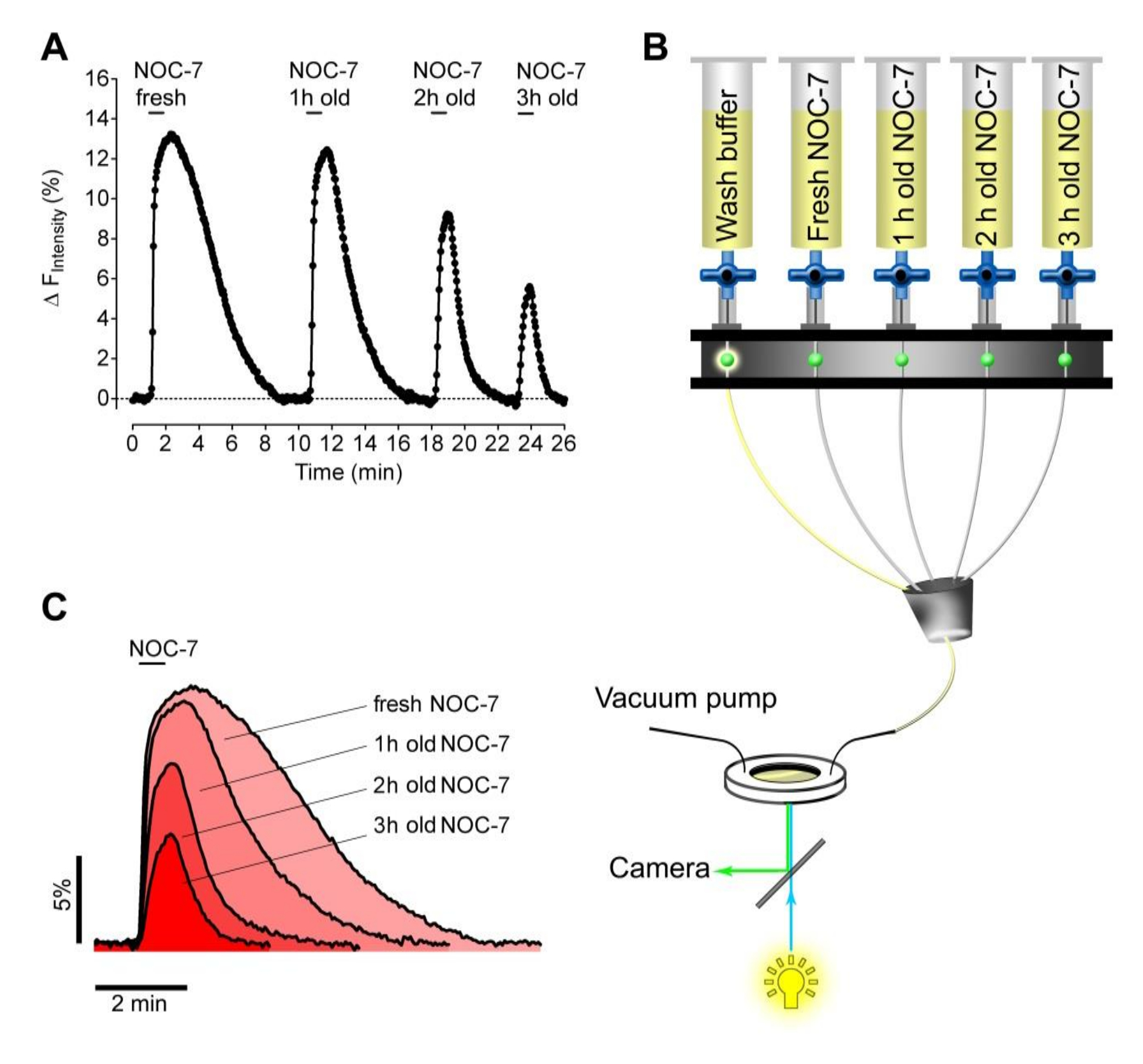{kind=link}

Рисунок 3: Одновременная многоканальная визуализация NO • и Ca 2+ сигналов в одиночных клетках EA.hy926. (A) репрезентативные широким полем флуоресцентного изображения клеток EAhy.926 , экспрессирующих G-GENOP (левая панель), которые загружаются с фура-2 / AM (средней и правой панелей). Шкала бар = 20 мкм. (Б) Схематическое изображение обработки клеток EAhy.926 с 500 мкМ Nω-нитро-L-аргинина (L-ННА) в течение 20 мин в течение шести-луночного планшета перед измерением. (C) представитель Временной ход одновременной записи фура-2 (возбуждение: 340 нм / 380 нм Emissиона: 510 нм) и сигналы G-GENOP (возбуждение: 480 нм; эмиссионные: 520 нм) в ответ на 100 мкМ гистамина. Как указано гистамин удаляли через 3 мин с использованием перфузионной системы. (D) Кривые представляют собой одновременные записи цитозольного Ca 2+ (фура-2 Отношение сигнал, F 340 / F 380 серая сплошная линия) и NO • (нормированные и перевернутый кривая, 1-F / F 0 зеленая сплошная линия) сигналы с течением времени одного фура-2 / я загружен эндотелиальные клетки транзиторно экспрессирующих G-GENOP. Клетки стимулировали 100 мкМ гистамина в течение 3 мин в Са 2+ (2 мМ CaCl 2) , содержащей буфер (n = 8/3). (Е) представитель одновременно записывается Ca 2+ (серая сплошная линия) и NO • сигналы (зеленая сплошная линия) с течением времени из ячейки EA.hy926 , который предварительно обрабатывают с помощью 500 мкМ Nω-Нитро-L-аргинина (L-NNA) до для измерения (п = 12/3). Клетки обрабатывали 100 мкМ гистамина в присутствии 2 мМCa 2+. (F) , усредненные кривые , представляющие цитозольных Нет • сигналы в ответ на 1 мкМ АТФ , за которым следует второй стимуляции клеток с помощью 100 мкМ АТФ. Бары представляют собой средние значения ± SD максимальных сигналов G-GENOP в ответ на 1 мкМ АТФ (белая полоса) и 100 мкМ АТФ (зеленая полоса) из 4-х независимых экспериментов; р <0,001 по сравнению с 1 мкМ АТФ. P-значение было рассчитано с использованием непарный т-тест. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
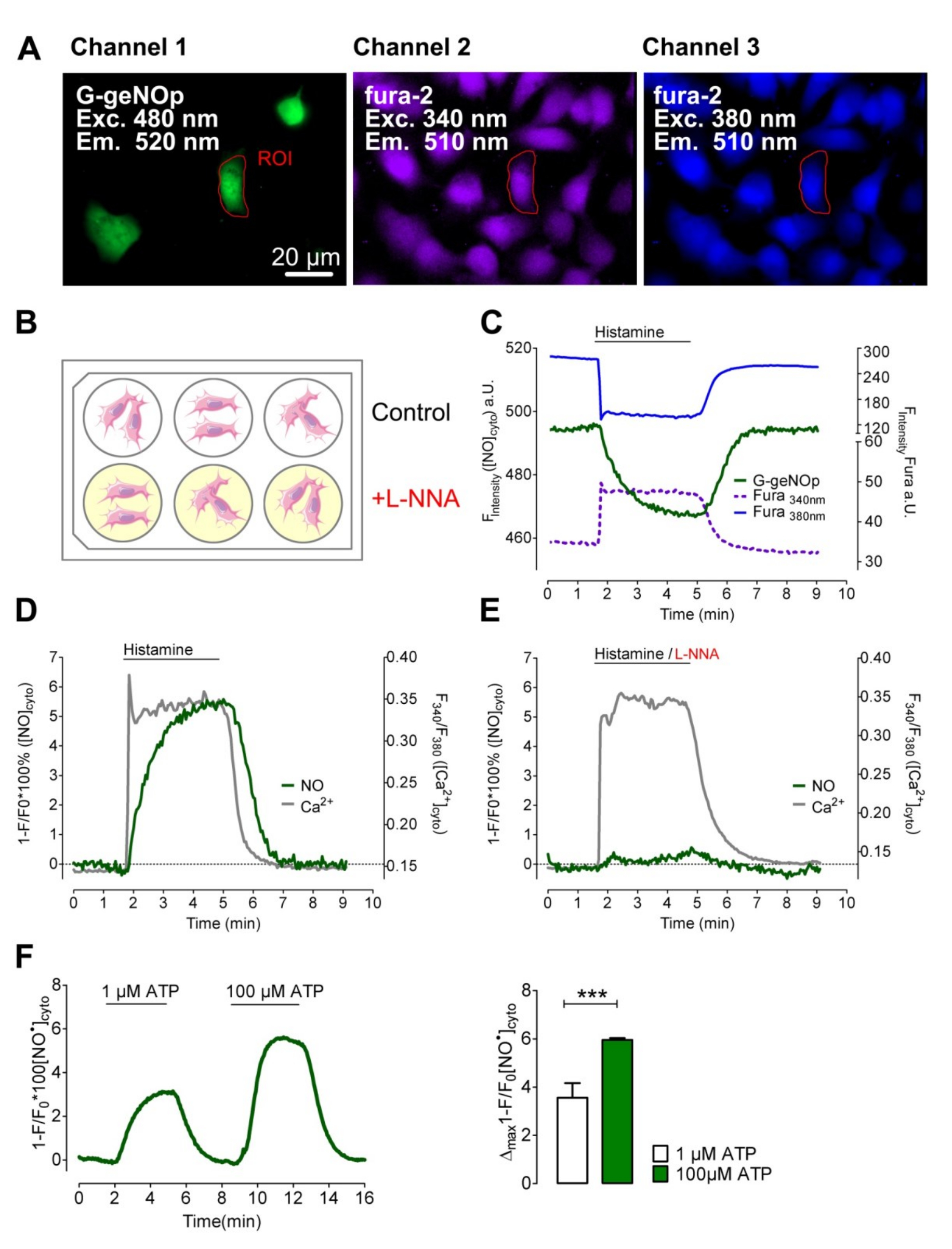{kind=link}
Обсуждение
С момента открытия NO • в качестве важной сигнальной молекулы в биологии 28, конкретное измерение в режиме реального времени радикала в отдельных клетках, тканях и целых животных с высоким разрешением в осуществимый и надежным образом было стремились. Здесь мы сообщаем о применении недавно разработанных генетически кодируемых флуоресцентных NO • зондов (geNOps) , которые позволяют точное жить-ячейки изображения NO • сигналов с использованием широкого поля флуоресцентной микроскопии 1.
Для того, чтобы обойти сложные и инвазивные процедуры трансфекции клона НЕК клеток, которые стабильно экспрессирует зеленый флуоресцентный G-GENOP был использован для количественного определения экзогенного генерироваться одноклеточных Нет • профили. Как НЕК клетки обычно не производят NO • эндогенно 29, этот тип клеток подходит для генерации GENOP на основе датчика клеточной линии, которая может оказаться полезной для многих других применений в условиях совместного культивированияс NO • производить первичные клетки или даже в живых животных 30. Тем не менее, в данном исследовании мы демонстрируем способность различных НЕТ • -liberating соединений, различных концентраций и стабильностей, пробуждать внутриклеточных NO • сигналы с использованием G-GENOP, выражающую модель НЕК клеток. Наши данные показали, что NO • -donor концентрация, качество и способ применения в конечном итоге определить модели внутриклеточных Нет • профили. Такая информация необходима для на месте Фармакокинетические характеристики различных доноров NO •, которые являются признаком ряда заболеваний. Следует отметить, что geNOps было показано , что стабильно реагировать на несколько повторяющихся применений NO • -donor импульсов в течение очень длительного времени 1. Таким образом, эксперименты с использованием NO • -liberating соединения, представленные в настоящем документе, позволяют Полуколичественные выводы относительно различных амплитуд и кинетики соответствующего клеточного NO226; сигналы (фиг.1 и 2).
Несмотря на то, стабильно экспрессирующих клон НЕК клеток , вероятно , происходит из одной клетки, широкая гетерогенность уровней экспрессии G-geNOps наблюдалось (рисунок 1). Это общая особенность стабильных клеточных клонов , как транскрипция (генома) интегрированный интерес гена находится под контролем многих факторов , таких как различных экологических стрессов 31, влияющих на темпы роста 32 клеток, и статус клеточного цикла 33. Отдельные ФП на основе geNOps не являются логометрических датчиков и, следовательно, NO • индуцированную потерю интенсивности флуоресценции возрастает с выражением уровня GENOP 1. Соответственно, нормализация geNOps сигналов имеет важное значение для количественной оценки клеточных NO • сигналы особенно в случае сравнительного анализа. Как показано в нашем недавнем исследовании, строгая линейная корреляция Ьetween базальная интенсивность флуоресценции geNOps и сила НЕТ • индуцированного тушения флуоресценции в широком диапазоне интенсивностей флуоресценции было обнаружено 1. Это важная особенность geNOps для абсолютного количественного определения клеточных NO • сигналов. Как показано на рисунке 1, нормализация G-geNOps сигналов в ответ на NOC-7 и SNP выявлены однородные NO • сигналы в различных НЕК клеток из одной тарелки, указывая , что НЕК клетки не разнообразны в отношении их способности принимать до и ухудшать NO • радикал, который берет свое начало от NO • донора. В отличие от этого, используя geNOps в клетках HeLa продемонстрировали четкие гетерогенность клеточных сигналов нет • между различными клетками в ответ на NOC-7. Эти различия указывают на NO • метаболизма и разложения скоростей типа специфических клеток, которые могут иметь несколько последствий в клеточной физиологии и патологии, а также могут быть обнаружены с помощью GENOPs технологии.
Тем не менее, две важные особенности geNOps должны быть тщательно продуманы для правильного использования датчиков и интерпретации данных: I) geNOps требуют адекватного железа (II) , чтобы полностью ответить на NO • 1 и II) в зависимости от варианта FP, geNOps может быть рН чувствительной 1. Здесь мы опишем протокол, который был найден, чтобы быть пригодным для нетоксичен железа (II) добавок geNOps, которые выражаются в любом НЕК, HeLa или клетки EA.hy926 (см протокол 2.6). В то время как это было показано , что лечение клеток с железом (II) / витамин С не влияет на морфологию клеток, жизнеспособность клеток и метаболическую активность клеток 1, это может быть необходимо оптимизировать этот важный шаг для других типов клеток и тканей. Тем не менее, в некоторых экспериментальных условиях, требование железа (II) загрузка может ограничить применимость geNOps. Следует отметить, что было показано, что аскорбиновая кислота может уменьшить NO • 35 и аскорбат железа (II) комплексы способны продувать NO • 36, 37. Кроме того, избыток железа (II) и аскорбат могут вызывать воспалительные реакции 39 и расцепить ENOS 41. Такие эффекты необходимо учитывать при использовании технологии geNOps. Было показано , что при определенных условиях эксперимента, внутриклеточный рН влияет значительно 34, который имеет потенциал , чтобы влиять на geNOps флуоресценции 1. Следует отметить, что голубые и зеленые geNOps варианты относительно рН чувствительны демонстрируют снижение флуоресценции при подкислении 1. Следовательно, острые изменения (суб) клеточного рН может имитировать ложные сигналы НЕТ • при использовании рН-чувствительного geNOps. Как было предложено в нашей предыдущей работе, параллельное использование NO • Нечувствительность geNOps (geNOps MUT) в качестве отрицательного ControLs рекомендуется рассекать реальный клеточный NO • сигнал от pH изменяется 1. Кроме того изменения клеточного рН могут быть проверены с использованием зондов , рН , такие как SypHer 34.
Кроме того, мы визуализировали эндогенной ферментативной NO • образование в ответ на физиологический Ca 2+ -mobilizing агониста в эндотелиальной клетке суррогатной EA.hy926. Клеточная линия EA.hy926 является часто используемая модель системы последовательно выражать Енос 38. Использование geNOps транзиторно экспрессируется в клетках EA.hy926, подтвердили , что IP - 3 -опосредованной Ca 2+ сигналы вызывают глубокое образование NO • В этом типе клеток, который был почти полностью перекрыт L-NNA. Для того , чтобы временно коррелируют Ca 2+ с NO • сигналы, G-GENOP-экспрессирующие клетки были загружены с УФ-возбудимый химической Ca 2+ -indicator фура-2 / AM. Спектральное разделение Са 2+ связанных и несвязанных фура-2 флуо ценция из сигнала G-GENOP может быть легко достигнуто с коммерчески доступными наборами фильтров 40. IMAGING оба зонда представила , что Ca 2+ -triggered ферментативная NO • формирование происходит гораздо медленнее по сравнению с цитозольного Ca 2+ рост этого типа клеток эндотелия. Подобные кинетика одноклеточных NO • сигналы в эндотелиальных клетках из бычьей легочной артерии при обработке клеток с IP - 3 -generating агонистом брадикинина, а также напряжения сдвига было зарегистрировано с использованием НОА-1, косвенное высоко НЕТ • датчик 12 , чувствительными , Соответственно, эти данные подчеркивают , что Ca 2+ -evoked Енос происхождения NO • образование требует определенного времени начала до полного ферментативная активность не будет достигнута. Хотя кинетика клеточного NO • формирование, распространение и деградации могут быть извлечены из других данных, например , измерения напряжения на основе из NO • индуцированный релаксации сосудовсс = "Xref"> 26, большая польза от флуоресцентных зондов • NO является то , что они непосредственно конвертировать клеточные NO • колебания в видимые сигналы в режиме реального времени. Следовательно, визуализации сотовой связи NO • сигналы с geNOps обеспечивает высокую пространственное и временное разрешение, а также предлагает уникальные возможности в (ре) исследования (суб) сотовой связи NO • гомеостаза. Например, Енос изображений челночные 42 в сочетании с технологией geNOps в одиночных эндотелиальных клетках может быть приемлемым не коррелировать NO • образование с внутриклеточной локализации и транслокации нет • -продуцирующих фермента или других соответствующих белков , таких как кальмодулин и кавеолином 43.
Здесь мы опишем достижимой применение G-GENOP, выражающую НЕК и EA.hy926 клетки визуализировать экзогенно и эндогенно генерируются клеточные NO • сигналы на уровне одной клетки и в режиме реального времени на обычном широком поле флуоресцентной мicroscope. Наши данные свидетельствуют о том, что geNOps подходят конкретно отслеживать (суб) клеточных динамику NO • при различных экспериментальных условиях, используя все виды интересных типов клеток.
Раскрытие информации
EE, MW, RM и WFG, сотрудники медицинского университета Граца, подали заявку на патент Великобритании (патентная заявка номер WO2015EP74877 20151027, приоритет номер GB20140019073 20141027), которые описывают части исследований в этой рукописи. Лицензии , связанные с этим патентом предоставляются Next Generation флуоресцентной томографии (NGFI) GmbH ( http://www.ngfi.eu/ ), спин-офф компании медицинского университета Граца.
Благодарности
The authors acknowledge C.J. Edgell, Pathology Department, University of North Carolina at Chapel Hill, NC, USA for providing the EA.hy926 cells. Author E.E. is supported by Nikon Austria within the Nikon-Center of Excellence, Graz and is a fellow of the Ph.D. program in Molecular Medicine at the Medical University of Graz. The researchers are also supported by the Ph.D. program Metabolic and Cardiovascular Disease (DK-W1226) of the Medical University of Graz. This work was also funded by the FWF project P 28529-B27. Microscopic equipment is part of the Nikon-Center of Excellence, Graz that is supported by the Austrian infrastructure program 2013/2014, Nikon Austria Inc., and BioTechMed, Graz.
Материалы
| Name | Company | Catalog Number | Comments |
| NaCl | Carl Roth, Karlsruhe, Germany | 3957.20 | sodium chloride |
| KCl | Carl Roth, Karlsruhe, Germany | 6781.1 | potassium chloride |
| CaCl2 .2H2O | Carl Roth, Karlsruhe, Germany | T885.1 | calcium chloride dihydrate |
| MgCl2 .6H2O | Carl Roth, Karlsruhe, Germany | 2189.2 | magnesium chloride hexahydrate |
| HEPES | Carl Roth, Karlsruhe, Germany | 9105.3 | 4-(2-Hydroxyethyl)-1-piperazineethanesulfonic acid |
| NaHCO3 | Carl Roth, Karlsruhe, Germany | 8551.1 | sodium hydrogencarbonate |
| KH2PO4 | Merck, Darmstadt, Germany | 104873 | potassium dihydrogen phosphate |
| Na2HPO4 .2H2O | Carl Roth, Karlsruhe, Germany | 4984.1 | sodium hydrogenphosphate dihydrate |
| D(+)-Glucose monohydrate | Carl Roth, Karlsruhe, Germany | 6780.1 | >99.5%; for cell culture, endotoxin free; |
| EGTA | Carl Roth, Karlsruhe, Germany | 3054.2 | ethylene glycol-bis(2-aminoethylether)-N,N,N′,N′-tetraacetic acid; calcium chelating agent |
| NaOH | Carl Roth, Karlsruhe, Germany | 6771.1 | sodium hydroxide |
| HCl | Carl Roth, Karlsruhe, Germany | 4625.1 | hydrochloric acid, fuming, 37% (~10 N) |
| DMSO | Carl Roth, Karlsruhe, Germany | 4720.1 | dimethyl sulfoxide; highly polar, aprotic organic solvent |
| L-Glutamic acid hydrochloride | Sigma Aldrich, Vienna, Austria | G2128 | (S)-2-Aminoglutaric acid |
| DMEM, low glucose | Sigma Aldrich, Vienna, Austria | D5523 | Dulbecco’s Modified Eagle’s Medium - low glucose; with 1000 mg/L glucose and L-glutamine, without sodium bicarbonate, powder, suitable for cell culture |
| MEM Vitamin solution (100X) | Gibco/Thermo Fisher Scientific, Waltham, MA, USA | 11120037 | 100x the vitamins found in the standard Minimum Essential Medium (MEM) |
| MEM Amino acids solution (50X) | Gibco/Thermo Fisher Scientific, Waltham, MA, USA | 11130036 | 50X the essential amino acids (except L-glutamine) found in the standard Minimum Essential Medium (MEM) |
| Penicillin-Streptomycin (10,000 U/ml) | Gibco/Thermo Fisher Scientific, Waltham, MA, USA | 15140122.00 | antibiotics to prevent bacterial contamination of cell cultures |
| Amphotericin B | Gibco/Thermo Fisher Scientific, Waltham, MA, USA | 15290026.00 | Gibco Amphotericin B contains 250 µg of amphotericin B (Fungizone) and 205 µg of sodium deoxycholate; prevents the contamination of cell cultures by yeast and multicellular fungi |
| FCS | Gibco/Thermo Fisher Scientific, Waltham, MA, USA | 10270106 | Fetal Bovine Serum, qualified, E.U.-approved, South America origin |
| PBS, pH 7.4 | Gibco/Thermo Fisher Scientific, Waltham, MA, USA | 10010031.00 | phosphate-buffered saline |
| Fura-2 (AM) | Teflabs, Austin, TX, USA | 102 | fluorescent cytosolic calcium indicators |
| Histamine dihydrochlorid | Sigma Aldrich, Vienna, Austria | H7250 | 2-(4-Imidazolyl)ethylamine dihydrochloride; IP3-generating agonist |
| Nω-Nitro-L-arginine | Sigma Aldrich, Vienna, Austria | N5501 | N5-(Nitroamidino)-L-2,5-diaminopentanoic acid; L-NNA; inhibitor of nitric oxide synthase |
| NOC-7 | Calbiochem/Merck, Darmstadt, Germany | 487952 | 3-(2-Hydroxy-1-methyl-2-nitrosohydrazino)-N-methyl-1-propanamine; nitric oxide (NO) donor short half-life of NO release |
| Sodium nitroprusside dihydrate | Santa Cruz Biotechnology, Texas, USA | sc-203395A | sodium nitroferricyanide(III) dihydrate; nitric oxide releasing compound |
| 30-mm Cover slips | Karl Hecht, Sondheim v. d. Rhön, Germany | 41001130 | glass cover slips, HECHT "Assistent", size 1, round, 30-mm, (VE: 100 pcs.) |
| Iron(II) booster solution | Next generation fluorescent imaging (NGFI), Graz, Austria | n.a. | Iron(II) containing physiological buffer for non toxic iron(II) loading of cells; http://www.ngfi.eu/product/ironii-booster-solution/ |
| G-geNOp plasmid | Next generation fluorescent imaging (NGFI), Graz, Austria | n.a. | plasmid DNA encoding green NO sensitive probe (G-geNOp); http://www.ngfi.eu/product/g-genop/ |
| G-geNOp AAV5 | Next generation fluorescent imaging (NGFI), Graz, Austria | n.a. | adenovirus encoding green NO sensitive probe (G-geNOp); http://www.ngfi.eu/product-category/genops/viral-genops-vectors/g-genop-aav5/ |
| G-geNOp sensor cell line | Next generation fluorescent imaging (NGFI), Graz, Austria | n.a. | human embryonic kidney cell line (HEK293) stably expressing green NO sensitive probe (G-geNOp); http://www.ngfi.eu/product-category/genops/g-genop-sensor-cell-line/ |
| HEK293A cell line | Invitrogen/Thermo Fisher Scientific, Waltham, MA, USA | R70507 | subclone of human embryonic kidney cell line (HEK293) |
| EA.hy926 cell line | American Type Culture Collection (ATCC), Wesel, Germany | CRL-2922 | somatic cell hybrid clone of human umbilical vein cell line with a thioguanine-resistant clone of A549 |
| TILL iMIC | Till Photonics, Graefling, Germany | n.a. | digital microscope |
| Polychrome V monochromator | Till Photonics, Graefling, Germany | n.a. | ultra fast switching |
| AVT Stingray F145B | Allied Vision Technologies, Stadtroda, Germany | n.a. | Versatile CCD camera with Sony ICX285 EXview HAD sensor, IEEE 1394b |
| alpha Plan Fluar 40 | Zeiss, Göttingen, Germany | n.a. | x40 objective |
| dichroic filters | Chroma Technology Corp, Rockingham, Vermont, USA | n.a. | GFP emitter 514/3 nm (515dcxr) |
| ValveBank8 Controller | AutoMate Scientific, Inc., Berkeley, California, USA | 01-08 | programmable perfusion system control unit |
| BVC control | Vacuubrand, Wertheim, Germany | 727200 | Chemistry diaphragm pump ME 1C; vacuum pump for perfusion system |
| ImageJ software | NIH Image | Java image processing program inspired by NIH Image. http://imagej.net/Welcome |
Ссылки
- Eroglu, E., et al. Development of novel FP-based probes for live-cell imaging of nitric oxide dynamics. Nat Commun. 7, 10623 (2016).
- Bush, M., et al. The structural basis for enhancer-dependent assembly and activation of the AAA transcriptional activator NorR. Mo. Microbiol. 95 (1), 17-30 (2015).
- Cranfill, P. J., et al. Quantitative assessment of fluorescent proteins. Nat. Methods. 13 (7), 557-562 (2016).
- D'Autréaux, B., Tucker, N., Spiro, S., Dixon, R., Poole, e. d. .. P. o. o. l. e. ,. R. .. K. .. ,. (. e. d. ). .. ,. Characterization of the Nitric Oxide-Reactive Transcriptional Activator NorR. Globins and Other Nitric Oxide-Reactive Proteins, Part B. 437, 235-251 (2008).
- Strack, R. Sensors and probes: Yes to genetically encoded NO• sensors. Nat Methods. 13 (4), 288 (2016).
- Auten, R. L. Response to 'The use of diaminofluorescein for nitric oxide detection: Conceptual and methodological distinction between NO and nitrosation. Free Radic. Biol. Med. 50 (12), 1812 (2011).
- Sivaraman, G., Anand, T., Chellappa, D. A Fluorescence Switch for the Detection of Nitric Oxide and Histidine and Its Application in Live Cell Imaging. ChemPlusChem. 79 (12), 1761-1766 (2014).
- Ye, X., Rubakhin, S. S., Sweedler, J. V. Detection of nitric oxide in single cells. Analyst. 133 (4), 423-433 (2008).
- Thyagarajan, B., Malli, R., Schmidt, K., Graier, W. F., Groschner, K. Nitric oxide inhibits capacitative Ca2+ entry by suppression of mitochondrial Ca2+ handling. Br J Pharmacol. 137 (6), 821-830 (2002).
- Germond, A., Fujita, H., Ichimura, T., Watanabe, T. M. Design and development of genetically encoded fluorescent sensors to monitor intracellular chemical and physical parameters. Biophy. Rev. 8, 121-138 (2016).
- Malli, R., Eroglu, E., Waldeck-Weiermair, M., Graier, W. F. Filling a GAP-An Optimized Probe for ER Ca2+ Imaging In Vivo. Cell Chem Biol. 23 (6), 641-643 (2016).
- Sato, M., Hida, N., Umezawa, Y. Imaging the nanomolar range of nitric oxide with an amplifier-coupled fluorescent indicator in living cells. Proc Natl Acad Sci USA. 102 (41), 14515-14520 (2005).
- Weidinger, A., Kozlov, A. V. Biological Activities of Reactive Oxygen and Nitrogen Species: Oxidative Stress versus Signal Transduction. Biomolecules. 5 (2), 472-484 (2015).
- Paolo, S. Nitric Oxide in Human Health and Disease. Encyclopedia of life sciences. , (2005).
- Pacher, P., Beckman, J. S., Liaudet, L. Nitric oxide and peroxynitrite in health and disease. Physiol Rev. 87 (1), 315-424 (2007).
- Bonafe, F., Guarnieri, C., Muscari, C. Nitric oxide regulates multiple functions and fate of adult progenitor and stem cells. J Physiol Biochem. 71 (1), 141-153 (2015).
- Forstermann, U., Sessa, W. C. Nitric oxide synthases: regulation and function. Eur. Heart J. 33 (7), 829-837 (2012).
- Dudzinski, D. M., Igarashi, J., Greif, D., Michel, T. The regulation and pharmacology of endothelial nitric oxide synthase. Annu Rev Pharmacol Toxicol. 46, 235-276 (2006).
- Zhou, L., Zhu, D. -. Y. Neuronal nitric oxide synthase: structure, subcellular localization, regulation, and clinical implications. Nitric Oxide. 20 (4), 223-230 (2009).
- Aktan, F. iNOS-mediated nitric oxide production and its regulation. Life Sci. 75 (6), 639-653 (2004).
- Ghafourifar, P., Cadenas, E. Mitochondrial nitric oxide synthase. Trends Pharmacol Sci. 26 (4), 190-195 (2005).
- Crane, B. R., Sudhamsu, J., Patel, B. A. Bacterial nitric oxide synthases. Annu Rev Biochem. 79, 445-470 (2010).
- Lundberg, J. O., Weitzberg, E., Gladwin, M. T. The nitrate-nitrite-nitric oxide pathway in physiology and therapeutics. Nat Rev Drug Discov. 7 (2), 156-167 (2008).
- Kelm, M. Nitric oxide metabolism and breakdown. Biochim Biophys Acta. 1411 (2-3), 273-289 (1999).
- Zhang, Y., et al. Estrogen-related receptors stimulate pyruvate dehydrogenase kinase isoform 4 gene expression. J Biol Chem. 281 (52), 39897-39906 (2006).
- Holzmann, S., Kukovetz, W. R., Windischhofer, W., Paschke, E., Graier, W. F. Pharmacologic differentiation between endothelium-dependent relaxations sensitive and resistant to nitro-L-arginine in coronary arteries. J Cardiovasc Pharmacol. 23 (5), 747-756 (1994).
- Bentley, M., et al. Vesicular calcium regulates coat retention, fusogenicity, and size of pre-Golgi intermediates. Mol Biol Cell. 21 (6), 1033-1046 (2010).
- Ignarro, L. J. Nitric oxide: a unique endogenous signaling molecule in vascular biology. Biosci Rep. 19 (2), 51-71 (1999).
- Upreti, M., Kumar, S., Rath, P. C. Replacement of 198MQMDII203 of mouse IRF-1 by 197IPVEVV202 of human IRF-1 abrogates induction of IFN-β, iNOS, and COX-2 gene expression by IRF-1. Biochem Biophys Res Com. 314 (3), 737-744 (2004).
- Lacin, E., Muller, A., Fernando, M., Kleinfeld, D., Slesinger, P. A. Construction of Cell-based Neurotransmitter Fluorescent Engineered Reporters (CNiFERs) for Optical Detection of Neurotransmitters In Vivo. J Vis Exp. (111), (2016).
- de Nadal, E., Ammerer, G., Posas, F. Controlling gene expression in response to stress. Na. Rev Genet. 12 (12), 833-845 (2011).
- Latchman, D. S. Transcriptional Gene Regulation in Eukaryotes. Encyclopedia of life sciences. , (2005).
- Bertoli, C., Skotheim, J. M., de Bruin, R. A. M. Control of cell cycle transcription during G1 and S phases. Nat Rev Mol Cell Biol. 14 (8), 518-528 (2013).
- Poburko, D., Santo-Domingo, J., Demaurex, N. Dynamic regulation of the mitochondrial proton gradient during cytosolic calcium elevations. J Biol Chem. 286 (13), 11672-11684 (2011).
- Suarez, S. A., et al. Nitric oxide is reduced to HNO by proton-coupled nucleophilic attack by ascorbate, tyrosine, and other alcohols. A new route to HNO in biological media. J Am Chem Soc. 137 (14), 4720-4727 (2015).
- Kuropteva, Z. V., Kudryavtsev, M. E. Ferrous-ascorbate complexes as carriers of nitric oxide. Gen Physiol Biophys. 16 (1), 91-96 (1997).
- Vanin, A. F., Huisman, A., Stroes, E. S., Ruijter-Heijstek, F. C., Rabelink, T. J., van Faassen, E. E. Antioxidant capacity of mononitrosyl-iron-dithiocarbamate complexes: implications for NO trapping. Free Radic Biol Med. 30 (8), 813-824 (2001).
- Lindberg, R. A., Dewhirst, M. W., Buckley, B. J., Hughes, C. S., Whorton, A. R. Ca2+-dependent nitric oxide release in endothelial but not R3230Ac rat mammary adenocarcinoma cells. Am J Physiol. 271 (1), 332-337 (1996).
- Campo, G. M., et al. The SOD mimic MnTM-2-PyP(5+) reduces hyaluronan degradation-induced inflammation in mouse articular chondrocytes stimulated with Fe (II) plus ascorbate. Int J Biochem Cell Biol. 45 (8), 1610-1619 (2013).
- Waldeck-Weiermair, M., et al. Spatiotemporal correlations between cytosolic and mitochondrial Ca2+ signals using a novel red-shifted mitochondrial targeted cameleon. PLOS ONE. 7 (9), 45917 (2012).
- Kuzkaya, N., Weissmann, N., Harrison, D. G., Dikalov, S. Interactions of peroxynitrite with uric acid in the presence of ascorbate and thiols: implications for uncoupling endothelial nitric oxide synthase. Biochem Pharmacol. 70 (3), 343-354 (2005).
- Liu, J., Hughes, T. E., Sessa, W. C. The first 35 amino acids and fatty acylation sites determine the molecular targeting of endothelial nitric oxide synthase into the Golgi region of cells: a green fluorescent protein study. J Cell Biol. 137 (7), 1525-1535 (1997).
- Feron, O. The Endothelial Nitric-oxide Synthase-Caveolin Regulatory Cycle. J Biol Chem. 273 (6), 3125-3128 (1998).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены