
Method Article
Анализ данных SEC-SAXS с помощью деконволюции и рассеяния EFA
* Эти авторы внесли равный вклад
В этой статье
Резюме
Измерения биологических макромолекул SEC-BioSAXS являются стандартным подходом для определения структуры растворов макромолекул и их комплексов. Здесь мы анализируем данные SEC-BioSAXS из двух типов часто встречающихся следов SEC – хроматограмм с полностью разрешенными и частично разрешенными пиками. Мы демонстрируем анализ и деконволюцию с помощью scatter и BioXTAS RAW.
Аннотация
BioSAXS является популярным методом, используемым в молекулярной и структурной биологии для определения структуры раствора, размера и формы частиц, соотношения поверхности к объему и конформации макромолекул и макромолекулярных комплексов. Высококачественный набор данных SAXS для структурного моделирования должен быть из монодисперсных, однородных образцов, и это часто происходит только путем сочетания хроматографии и немедленного измерения SAXS. Чаще всего хроматография с исключением размеров используется для отделяния образцов и исключения загрязняющих веществ и агрегаций из частиц, представляющих интерес, что позволяет проводить измерения SAXS с хорошо решенного хроматографического пика одного вида белка. Тем не менее, в некоторых случаях даже высовывание не является гарантией монодисперсных образцов, либо потому, что несколько компонентов слишком близко друг к другу по размеру, либо изменения формы, вызванные обязательным изменением воспринимаемого времени elution. В этих случаях может быть возможно деконволюция данных SAXS смеси для получения идеализированных кривых SAXS отдельных компонентов. Здесь мы покажем, как это достигается, и практический анализ данных SEC-SAXS выполняется на идеальных и сложных образцах. В частности, мы показываем анализ SEC-SAXS вакцины E9 ДНК полимеразы экзонуклеазы минус мутант.
Введение
Биологические макромолекулы слишком малы, чтобы их можно было увидеть даже с помощью лучших световых микроскопов. Современные методы определения их структур обычно включают кристаллизацию белка или измерения на огромном количестве одинаковых молекул одновременно. Хотя кристаллография предоставляет информацию об атомном уровне, она представляет собой искусственную среду образца, учитывая, что большинство макромолекул не представлены в кристаллической форме в клетке. За последние пару лет криоэлектронная микроскопия доставила аналогичные структуры высокого разрешения крупных макромолекул/ макромолекулярных комплексов, но хотя образцы ближе к физиологическому состоянию, они все еще заморожены, следовательно неподвижны и статичны. Био-малый угол рентгеновского рассеяния (BioSAXS) обеспечивает структурное измерение макромолекулы, в условиях, которые имеют отношение к биологии. Это состояние можно визуализировать как 3-D-форма с низким разрешением, определяемая по нанометровой шкале, и фиксирует все конформациатное пространство макромолекулы в растворе. Эксперименты BioSAXS эффективно оценивают олигомерное состояние, домен и сложные механизмы, а также гибкостьмежду доменами 1,2,3. Метод является точным, в основном неразрушаемым и обычно требует лишь минимума подготовки образца и времени. Однако для наилучшего толкования данных выборки должны быть монодисперсами. Это сложная задача; биологические молекулы часто подвержены загрязнениям, плохой очистке и агрегации, например, от замораживания оттаивания4. Развитие inline хроматографии с последующим немедленным измерением SAXS помогает смягчить эти эффекты. Размер-исключение хроматографии отделяет образцы по размеру, таким образом, исключая большинство загрязняющих веществ и агрегаций5,6,7,8,9,10. Тем не менее, в некоторых случаях даже SEC-SAXS не достаточно, чтобы произвести монодисперсный образец, потому что смесь может состоять из компонентов, которые слишком близко по размеру или их физические свойства или их быстрая динамика приводит к перекрывающихся пиков в УФ-след SEC. В этих случаях этап деконволюции полученных данных SAXS на основе программного обеспечения может привести к идеализированной кривой SAXSотдельного компонента 5,11,12. В качестве примера, в протоколе раздела 2, мы показываем стандартный анализ SEC-SAXS вакцины E9 ДНК полимеразы экзонуклеазы минус мутант (E9 exoминус) в комплексе с ДНК. Vaccinia представляет собой модель организма Poxviridae, семьи, содержащей несколько патогенов, например, вирус оспы человека. Полимераза была показана плотно связываться с ДНК в биохимических подходов, со структурой комплекса недавно решена рентгеновской кристаллографии13.
Большинство синхротронных объектов обеспечат автоматизированный конвейер обработки данных, который будет выполнять нормализацию и интеграцию данных, создавая набор неподписанной рамы. Но подход, описанный в этой рукописи, также может быть использован с лабораторным источником при условии, что SEC-SAXS выполняется. Кроме того, может быть доступна дополнительная автоматизация, которая будет отвергать поврежденные радиацией кадры и выполнять вычитаниебуфера 14. Мы покажем, как проводить первичный анализ данных о предварительно обработанных данных и использовать имеющиеся данные в разделе 2.
В разделе 3 мы показываем, как деконволюция данных SEC-SAXS и эффективно анализировать кривые. Хотя Есть несколько методов деконволюции, таких как гауссийский пик деконволюции, реализованы в США-SOMO15 и Guinier оптимизированный метод максимальной вероятности, реализованный в программном обеспечении DELA16, они обычно требуют модели для пиковойформы 12. Конечный размер отдельных пиков, которые мы расследуем, позволяет использовать развивающийся анализ факторов (EFA), как улучшенную форму разложения особого значения (SVD) для деконволюционных перекрывающихся пиков, не полагаясь на пиковую форму или рассеянныйпрофиль 5,11. Конкретную реализацию SAXS можно найти в BioXTAS RAW17. EFA был впервые использован на данных хроматографии, когда данные 2D диодного массива позволили матрицы быть сформированы из поглощения против времени удержания и длины волныданных 18. Где EFA превосходит в том, что она фокусируется на развивающемся характере особых значений, как они меняются с появлением новых компонентов, с оговоркой, что есть неотъемлемый порядок вприобретении 10. К счастью, данные SEC-SAXS предоставляют все необходимые упорядоченные данные о приобретении в организованных массивах данных 2D, хорошо одалживая себя методу EFA.
В разделе 4 мы продемонстрируем основы независимого от модели анализа SAXS из буферно-фоновой вычитаемой кривой SAXS. Модель-независимый анализ определяет радиус действия частицы (Rg), объем корреляции (Vc), Porod Volume (Vp) и Porod-Debye Exponent (PE). Анализ дает полуколичебную оценку термодинамического состояния частицы с точки зрения компактности или гибкости с помощью без измерения участкаKratky 2,4,19.
Наконец, данные SAXS измеряются во взаимных космических единицах, и мы покажем, как преобразовать данные SAXS в реальное пространство для восстановления парного расстояния, P(r), функции распределения. P(r)-распределение — это набор всех расстояний, найденных внутри частицы, и включает в себя максимальное измерение частицы, dmax. Поскольку это термодинамическое измерение, P(r)-распределение представляет физическое пространство, занимаемое конформациальным пространством частиц. Правильный анализ набора данных SAXS может предоставить информацию о состоянии решения, которая дополняет информацию высокого разрешения из кристаллографии и крио-EM.
протокол
1. Выражение белка, очистка и измерение SEC-SAXS основаны на опубликованном протоколе13
- Кратко следуйте протоколу сбора данных SEC-SAXS (Brennich et al.6).
- Equilibrate SEC-колонка с по крайней мере 2 объемами столбца SEC работает буфер (20 мМ Tris-HCl, рН 7,5, 100 мМ. NaCl).
- Подготовь 50 л образца экзо минус E9 на уровне 8'u201210 мг/мл с 20% мл избыток частичного dsDNA (TCAGGAAGATAACAGCGGTTTAGCC и GGCTAAACCGCTGTTATCTTTTTTTTTTTTTTTTTTTTTT). E9 exoминус связывается с KD 12 ± nM (см. Дополнительные данные).
- Ввись 50 МКЛ этой смеси на SEC-колонку (S200 Increase) в ряд с ячейкой потока для измерений SAXS на уровне 0,3 мл/мин.
- Соберите 1000 кадров на 1 с экспозицией каждый.
ПРИМЕЧАНИЕ: На луче BioSAXS BM29, в Европейском синхротронном комплексе (ESRF) отдельные кадры обрабатываются автоматически и независимо в рамках EDNA14. После сбора данных откройте базу данных ISPyB20 и под вкладкой сбора данных нажмите кнопку Go, чтобы получить доступ к набору данных и результатам автоматического анализа21.
- Скачать данные.
2. Первичный анализ данных
- Откройте java-программу Рассеяние IV (см. Таблица материалов) и выполнить фоновое вычитание данных хроматографии исключения размера (SEC).
- Откройте вкладку SEC. Перетащите и уроните уменьшенныефайлы данных(.dat)в окно "Drop Data ниже". Установите выходной каталог "Out Dir ::", нажав на кнопку выходного Dir с синей маркировкой.
ПРИМЕЧАНИЕ: Если ваши данные были собраны в nm-1, коробка преобразования должна быть проверена (внизу слева от панели) при падении файлов в окно или вкладку вычитания. - Редактировать экспериментальные детали, использовать кнопку Редактировать детали и заполнить как можно больше полей, как это возможно, они включают разделы, на которых источник / луч был использован для сбора данных, параметры сбора и детали выборки. Они будут сохранены с помощью данных и позволят легче заполнить раздел "Параметры сбора данных" в будущих публикациях.
- Введите имя образца в Сохранить как поле. Нажмите на TRACE.
ПРИМЕЧАНИЕ: Это имеет два эффекта. Во-первых, он создаст файл для данных. Это единый текстовый файл, который будет собирать все экспериментальные наблюдения из отдельных файлов .dat файлов. Кроме того, файл «.sec» содержит усредненный набор кадров, который является буферным фоном, все кадры, используемые в усреднении, а также вычитаемые буферные кадры по всему эксперименту SEC-SAXS. Во-вторых, создается сигнальный сюжет, который отображает число кадров по сравнению с соотношением Integral к фону. Это показывает выбранные кадры (серые), которые были усреднились для вычитания буфера. Точки для среднего буфера определяются из всего диапазона данных. Тем не менее, желательно вручную выбрать буферные рамки для усреднения, так как плохо определенный фон может произойти из-за плохо уравновешенных или грязных столбцов или капиллярногозагрязнения 22. - Выберите буферные кадры вручную. Нажмите Clear Buffers затем переизбирайте буферную область с левым щелчком мыши, на кривой трассировки. В идеале это должна быть плоская область перед пустотой, объем колонны SEC составляет около 100 кадров. Нажмите SET BUFFER, а затем обновление, чтобы пересчитать файл.
- Определите область интересов (ROI). На сигнальный сюжет выберите область пика интереса, с левым щелчком мыши.drag.
ПРИМЕЧАНИЕ: Это заполняет три участка в правой панели. Два верхних участка связаны с перекрестием, перемещаясь между ними, второй участок сигнала (вверху справа) показывает только выбранную рентабельность инвестиций, с интенсивностью каждого кадра синим цветом и соответствующей Rg каждого кадра красным цветом и соответствующей тепловой картой ниже, показывая остатки для каждого кадра, окрашенного в соответствии с анализом автоматической корреляции Дурбин-Уотсон. Регионы с высоким сходством окрашены в синий цвет (Дурбин-Уотсон, d No 2), в то время как непохожие кадры будут следовать темные синие до розовых и, наконец, красные в зависимости от тяжести непохожести (d'gt; 2). Нижний участок является вычитаемой кривой I по сравнению с q для центрального выбранного кадра (также обозначаемого вертикальной линией). Клавиши стрелки могут быть использованы для навигации по вычитаемые кадры. Сюжет I против q продемонстрирует качество вычитаемых кадров из эксперимента SEC. - Выберите кадры для слияния. Нажмите на перекрестие в графике тепловой карты, чтобы выбрать подмножество кадров, которые будут использоваться для слияния. Перекрестие определит треугольную область преимущественно сияного, который падает на правую нижнюю сторону перекрестия. Используйте мыши щелчок, чтобы установить эти кадры, как выбрано и выделить кадры в соответствующей области сигнала участка выше. Эти кадры должны идеально выделить область со стабильной Rg.
ПРИМЕЧАНИЕ: По мере необходимости, увеличить на тепловой карте с левым щелчком мыши перетащить и увеличить с левым щелчком салфетки вправо. - При удовлетворении выбранными кадрами щелкните MERGE. Это объединит вычитаемые кадры и представит их во вкладке ANALYSIS.
- Откройте вкладку SEC. Перетащите и уроните уменьшенныефайлы данных(.dat)в окно "Drop Data ниже". Установите выходной каталог "Out Dir ::", нажав на кнопку выходного Dir с синей маркировкой.
3. Деконволюция данных
- Откройте программу деконволюции (например, BioXTAS Raw 2.0.0).
- В программе деконволюции загрузите набор данных под вкладкой Файлы в панели управления,используйте символ foldether, чтобы найти данные или скопировать и вставить местоположение в адресную панель.
ПРИМЕЧАНИЕ: Убедитесь, что папка содержит только необработанные файлы .dat и не обработанные или средние файлы данных. - Выделите все файлы .dat, нажмете кнопку Серии Сюжетов, сюжет интегрированной интенсивности по сравнению с номером кадра будет нарисован в «Сюжете серии».
- В панели управления выберите вкладку серии, а затем нажмите, чтобы выделить кривую. Откройте всплывающее окно LC Analysis с помощью кнопки в основании панели управления. Это окно дает доступ к нескольким вариантам, таким как выбор различных типов молекул (белка или РНК). Это также позволяет пользователю выбрать буферную область для участка. В первом случае нажмите Auto; это должно выбрать подходящий буферный регион.
ПРИМЕЧАНИЕ: Если это не удается, возможно, из-за нестабильной базовой линии, то "Добавить регион" для оптимизации буферной области. Это заполняет поле Буфера с меньшим ящиком, в котором можно вручную добавить номера кадров для использования для буфера. Кроме того, нажмите кнопку "Pick", чтобы дать возможность выбора области на участке. Найдите область, слева нажмите один раз для начала позиции, переместить курсор в следующую позицию и слева нажмите еще раз. Возможно, необходимо добавить несколько буферных мест. Нажмите Набор буфера и кривые будут вычтены и Rg рассчитывается через пик SEC. Если появляется всплывающее окно, нажмите OK. - Чтобы начать анализ эволюционирующим фактором (EFA), нажмите правой кнопкой мыши на выделенный файл в нижней части Панель управления а затем выберите Efa из меню.
- Проверьте, открывается всплывающее окно, которое показывает разложение одного значения (SVD) набора данных. В поле управления проверьте поле Use Frames так, чтобы вся область пика для деконволюта была покрыта участком интенсивности. Участок "Особые значения", в правом верхнем варианте, показывает интенсивность особых значений (отдельных пиков/видов) выше базового уровня.
ПРИМЕЧАНИЕ: Количество точек, присутствующих выше базового уровня, представляет собой количество присутствующих видов рассеяния. С оговоркой, что это относительная величина особого значения плоской области / базовой линии, что вопросы. - Чтобы проверить количество отдельных значений, используйте нижний участок AutoCorrelation. Это показывает правый и левый векторы корреляции. Нажмите далее.
ПРИМЕЧАНИЕ: Они по существу представляют собой профили рассеяния или концентрации вектора в растворе. Где абсолютный размер представляет значение вектора. Значительный компонент будет иметь автокорреляцию вблизи 1 (правило отсечения большого пальца составляет 0,6-u20120.7). RAW услужливо вычисляет это и отображается в #Significant SVs, внизу слева, хотя вы можете изменить это в случае необходимости. При нескольких отдельных значениях (например, 4 евро) может потребоваться взглянуть только на 2 или 3 компонента, изменив диапазон используемых данных. Чем меньше количество компонентов, тем легче будет анализ EFA, но за счет использования меньше данных. В сложной ситуации, когда левые и правые сингулярное векторы, которые должны быть похожи, не совпадают, уменьшают значительное число SV и уменьшают количество кадров, используемых до тех пор, пока левые и правые сингулярное векторы не будут похожи. - Убедитесь, что EFA рассчитывается путем генерации участков в направлениях вперед и назад для каждого вектора. Эти участки показывают, когда компоненты запускаются (форвардный участок) и выходят (назад) профиль решения для выбранных данных SEC-SAXS. RAW пытается определить эти диапазоны; изменить их, используя стрелки рядом с счетчиками, так что каждый круг находится в начале точки перегиба, поднимающейся от или падающей к исходной линии. Нажмите далее.
ПРИМЕЧАНИЕ: Последний этап EFA превращает векторы SVD обратно в кривые рассеяния. Слева от окна в верхней части построены ранее определенные диапазоны. Эти диапазоны являются ограничениями для определения того, где повернуть сингулярные векторы обратно в кривые рассеяния. Правая панель показывает эти соответствующие профили кривой рассеяния, для каждого разделенного пика. Сюжет для концентрации каждого пика, который должен быть репрезентативным для профилей elution и сюжет для среднего ошибка взвешенных чи2. Участок чи2 измеряет набор данных деконволюции к исходному набору данных. В идеале, это будет плоским, однако шипы часто можно увидеть. - Попробуйте уменьшить или устранить шипы, изменив элементы управлениядиапазоном компонентов, сначала определите, какой кадр соответствует шипу (от chi2 участок), а затем, в диапазоне управления, который компонент содержит этот кадр (это может быть больше, чем один), используя стрелки, двигаться вверх или вниз соответствующего диапазона.
ПРИМЕЧАНИЕ: Это должно дать ответ, увеличивая или уменьшая всплеск. Если рама шипа присутствовала в более чем одном компоненте, то может потребоваться небольшое время проб и ошибок между каждым компонентом. - Когда минимальный чи2 был достигнут, выполнить проверку проверки, нажав назад, предыдущее окно, как представляется, позволяют проверить, если изменения, внесенные резко изменили исходные участки EFA. Если они по-прежнему выглядят действительными, нажмите Далее. Нажмите Сохранить данные EFA, чтобы сохранить участки, а затем нажмите Готово; закрыть окно EFA.
ПРИМЕЧАНИЕ: Вторая проверка заключается в том, чтобы нажать от чек-бокс рядом с каждым компонентом диапазона, в свою очередь. Они обеспечивают положительное ограничение концентрации для каждого компонента и выключение будет проверять, если они существенно влияют на набор данных. Если на участке концентрации изменений не наблюдается, то данные действительны.
- Проверьте, открывается всплывающее окно, которое показывает разложение одного значения (SVD) набора данных. В поле управления проверьте поле Use Frames так, чтобы вся область пика для деконволюта была покрыта участком интенсивности. Участок "Особые значения", в правом верхнем варианте, показывает интенсивность особых значений (отдельных пиков/видов) выше базового уровня.
- Вернувшись в окно RAW, нажмите на вкладку Профили в панели управления, чтобы просмотреть кривые и во вкладке Манипулирование панели управления,манипулировать кривыми дальше или сохранить кривые, как файлы No .dat, нажав правой кнопкой мыши на файл и выбрав сохранить выбранный файл (ы) из меню всплывающее. Сохраните файл. Используйте Scatter IV для дальнейшего анализа.
ПРИМЕЧАНИЕ: Дополнительная информация и инструкции по деконволюции и EFA BioXTAS RAW можно найти https://bioxtas-raw.readthedocs.io/en/latest/
4. Определить свойства SAXS
ПРИМЕЧАНИЕ: По крайней мере, в Bioisis.net. Здесь мы показываем основной шаг за шагом подход, подчеркивая наиболее полезные кнопки в Scatter.
- Во вкладке Scatter ANALYSIS нажмите кнопку G для ручного инструмента анализа Guinier справа от каждого файла образца. Сюжет, который открывается, показывает, что в верхней коробке находитсяln'i (q) и q2 в верхней коробке и соответствующие остатки в нижней коробке. Добавить или удалить точки таким образом, что остатки не имеют "улыбка" или "хмуриться" функцию. Выбранные данные в подгонки Guinier не должны превышать максимальный лимит q x Rg 1.3.
- Нажмите кнопку Нормализованный Кратки; участок, который всплывает обеспечивает полуколиченую оценку структурного состояния макромолекулы, нормализуется для массы и концентрации.
ПРИМЕЧАНИЕ: Перекрестие обозначает точку Guinier-Kratky на (√3, 1.1)19. Компактный сферический белок покажет один пик с максимальным значением в точке Гинье-Кратки. Внутренне неупорядоченно или цилиндрический биополимер будет иметь максимум больше, чем перекрестие и не будет уменьшаться. Белок, который имел как сложенные домены, так и длинные удлиненные неструктурированные регионы, может представлять 2000-й максимум через перекрестие, но также покажет очевидную тенденцию к снижению на более высоком q x Rg. - Нажмите на кнопку Vc (Volume-of-correlation), которая воспитывает два участка, общую рассеянную интенсивность и интегрированную область общей рассеянной интенсивности в качестве функции q. Участки используются в качестве быстрой ссылки для проверки качества кривой рассеяния.
ПРИМЕЧАНИЕ: Общая рассеянная интенсивность чувствительна к I(0), и если это не было измерено правильно, то сюжет не будет показывать непрерывную линию. Интегрированный участок области, в идеале, должен показать сигмоидную линию с расширенным плато для каждой кривой SAXS. При несоответствии/вычитании буфера агрегация или интерчастицное вмешательство в выборку наблюдаются при более высоких значениях q-значений. - Нажмите кнопку Гибкость, чтобы начать анализ гибкости. Это откроет окно с четырьмя панелями и ползунок в нижней части. Каждая открытая панель показывает участок, эксплуатируемый отношения между властью и законом, который существует между компактными и удлиненными/гибкимибиополимами 23. Для использования переместите ползунок в нижней части коробки справа налево с нажатием кнопки левой мыши. Продолжайте медленно двигаться влево, пока плато в одном из участков не будет достигнуто.
ПРИМЕЧАНИЕ: Если плато видно на участке Пород-Дебье, то образец компактный по своей природе, который должен соответствовать одной вершине в точке Гинье-Кратки в нормализованной кратки. Если плато достигло первого участка Кратки-Дебье, то образец, скорее всего, удлиненный или гибкий. Если участок SIBYLS сначала плато, то образец, скорее всего, содержит области как компактности и гибкости, частицы со смешанными состояниями. Теория такой гибкости в отношениях с Законом Пород-Дебье изысканно рассматривается в Рэмбо, идр. - Нажмите на том. Определение объема должно быть выполнено сразу же после анализа гибкости сверху. При открываемом после анализа гибкости генерируется всплывающее окно с еще тремя графиками. В нижнем, левом углу участок Porod-Debye помнит, где один оставил ползунок от участка гибкости, показывая площадь плато.
- Чтобы рассчитать объем частицы, переместите стартовые и конечные точки с помощью кнопок стрелки или ввемите в ящики, чтобы синяя линия на участке вписывалась в область плато. Для объективного результата, остатки в правом верхнем Porod-Debye экспонента власти закона подходят, не должны показывать шаблон.
- Нажмите на вкладку P(r). Распределение в реальном пространстве находится в левой панели и кривой рассеяния для образца в правой панели. Цель состоит в том, чтобы создать реальное представление образца из взаимного пространства кривой SAXS. В идеале кривая распределения будет гладкой без волны настоящее время и должны просто осторожно поцеловать x-оси.
ПРИМЕЧАНИЕ: Измеренный q-диапазон прибора не может быть полностью использован из-за плохого сопоставления буфера, агрегации, радиационного повреждения, неоптимальных времен экспозиции и низких концентраций частиц. Этап определения P(r)будет в корне определять используемый диапазон qmin и qmax набора данных SAXS, и именно этот диапазон данных должен использоваться для любого последующего моделирования или установки.- Нажмите правой кнопкой мыши на имя образца, а затем нажмите Найти DMAX, чтобы открыть новое окно. Ограничения для dmax предварительно установлены с предлагаемым qmax (максимальные используемые точки данных), нижними иверхними максимальными ограничениями d и более низким и верхним альфа-баллом. Три модели могут быть выбраны (L1-норма, Legendre и Мур) и использование фона включены. Оставьте их без изменений в первую очередь.
- Нажмите кнопку "Пуск". Композитное распределение создается в левой панели с предлагаемым dmax и альфа-уровнем, написанным под ним. Если это выглядит приемлемым, то закройте окно и вернитесь к вкладке P(r). Взаимный участок пространства будет обрезан, чтобы соответствовать предложенному qmax.
- Выберите модель Moore, нажмите на фон, а затем установите альфа-уровень и dmax к предлагаемым значениям из всплывающего окна. Нажмите кнопку уточнения. Появляется участок перекрестной проверки, показывающий, если какие-либо точки должны были быть отклонены, отмеченные красным цветом. Если есть только несколько точек отклонены и распределение выглядит хорошо, то модель хорошая.
ПРИМЕЧАНИЕ: Участок перекрестной проверки будет выделить области данных, которые несовместимы с определяемой P(r)-распределением. Если отвергнутый регион находится главным образом в регионе с низким уровнем q, то есть в регионе, близком к оси, это, вероятно, предполагает dmax, который является слишком коротким, наличие агрегации или олигомеров более высокого порядка. В нем подчеркивается несоответствие между информацией о более высоком и низком разрешении. Здесь dmax и qmin (увеличение значения старта) должны быть скорректированы с помощью ручного подхода проб и ошибок. Аналогичным образом, если отклоненный регион находится главным образом в регионе с высоким q, это может указывать на проблему с фоновым вычитанием или на то, что сигнал слишком слаб, чтобы быть значимо объясненным определяемым распределением P(r).-. В этом случае q max следуетусечен (уменьшение конца) до тех пор, пока не будут отклонены дополнительные данные. В идеале отклоненные точки должны распределяться случайным образом и составить менее 5% от годных к использованию данных. Правильно определенный qmin,qmax и "dmax"будет производить плавное распределение, где dmax целует x-axis. Тем не менее, не увеличивать это значение настолько, что он полностью удаляет регион Guinier. Эта точка легко найти, проверяя q x l (q) поле (слева от панели над таблицей). Кривая рассеяния заменяется "Всего рассеянного участка интенсивности", на этой кривой все точки до максимального перегиба являются частью области Guinier. После удаления точек попробуйте еще раз увеличить /уменьшить "dmax",а затем уточнить еще раз. Если проблемы сохраняются, особенно когда многие точки отклоняются с самого начала кривой проверки, это убедительно указывает на то, что данные не идеально подходят для структурного моделирования.
- Чтобы распечатать отчет, перейдите к вкладке Анализ, слева нажмите, чтобы выделить образец, а затем право нажмите на имя образца и перейти к Созданию отчета из одного набора данных в меню. Открывается текстовый ящик, позволяющий добавить комментарии. Производится документ PDF, показывающий все цифры и значения, генерируемые.
Результаты
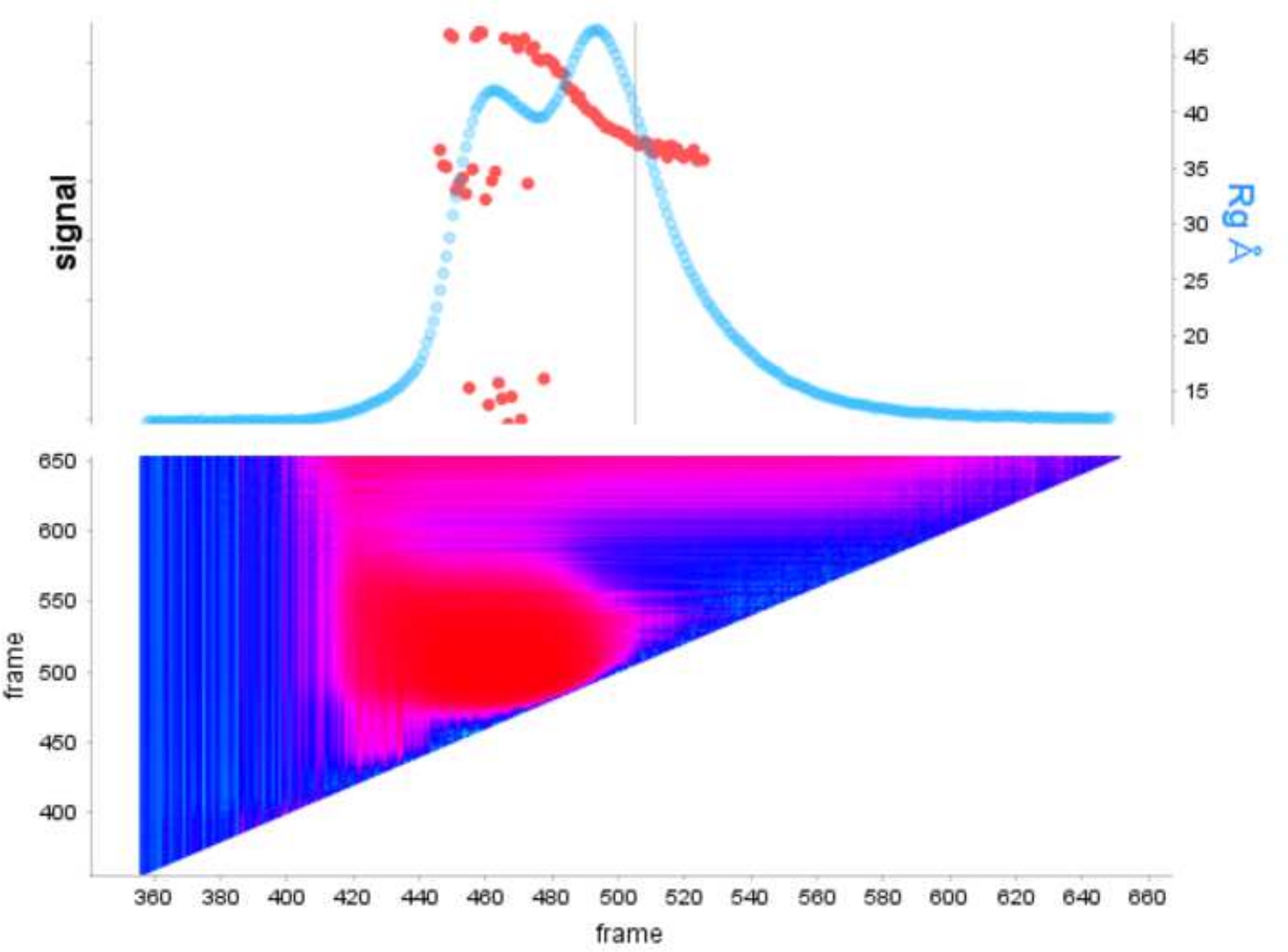
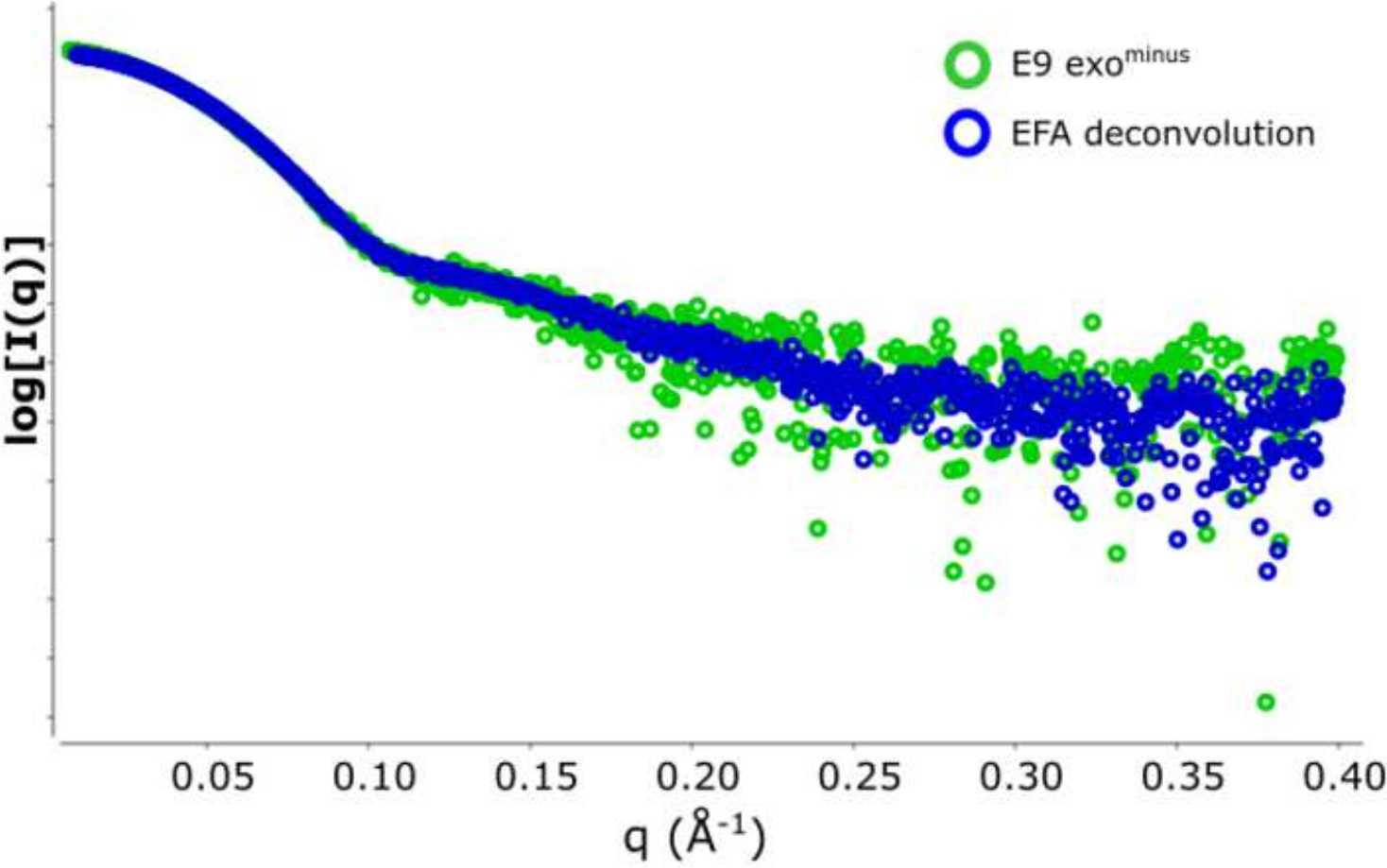
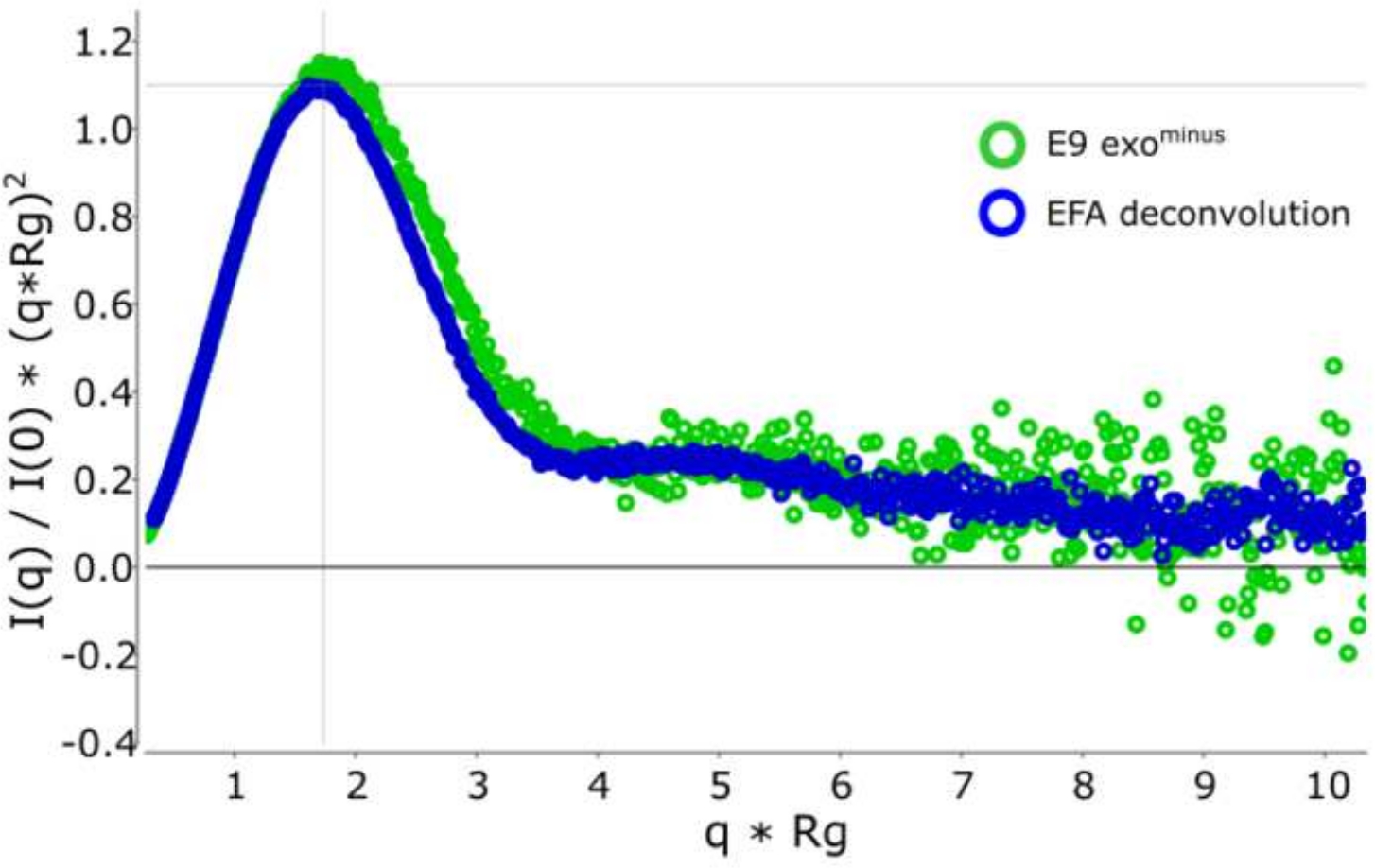
Преимущество использования деконволюции над классическим выборомрамы 13 заключается в том, чтобы снять влияние видов друг на друга, производя монодисперсный сигнал рассеяния. Это также часто сопровождается лучшим соотношением сигнала к шуму. Когда E9 exoминус связан с ДНК и запустить с помощью SEC-SAXS, два пика наблюдаются(рисунок 1). Первый, большой пик (примерно кадры 420-u2012475) является E9 exoминус-DNAкомплекс второй (примерно кадры 475'u2012540), несыхое состояние (см. Дополнительные данные: Рисунок 2). В то время как классический подход к выбору кадров обеспечивает стабильную Rg комплекса в первом пике (см. Дополнительные данные: Рисунок 3), второй пик явно сливается и Rg по сюжету показывает, что второй пик интереса не имеет стабильной Rg, из-за кросс-пик загрязнения. Только 5 кадров могут быть использованы, которые показали полу-стабильный Rg, при вычитании они дали Rg No 36,3(рисунок 2, зеленый). Когда пики были деконволюция с помощью EFA соответствующая кривая для второго пика(рисунок 2, синий) был наложен с оригиналом и показал явное снижение сигнала к шуму, и ниже Rg, 34,1 З был записан. Крацкий участок(рисунок 3) показывает комплекс с деконволюцией пика (синий) является более шаровой. Это подтверждается кривой P(r) (Рисунок 4), который дает dmax 108.5 для деконволюции кривой (синий), в то время как недекололетированная более удлиненная с dmax 120 (зеленый), это, скорее всего, из-за неоднородности, вытекая из несыхватых E9 exoминус.

Рисунок 1: Сигнальный сюжет E9 exoминус в одиночку и с ДНК в комплексе.
Верхняя панель показывает участок интегрального соотношения к фону для каждого кадра запуска SEC-SAXS (светло-голубой). Красные точки показывают Rg на каждом кадре над пиком. Нижняя панель показывает соответствующую тепловую карту, показывающую остатки для каждого кадра, окрашенного в соответствии с анализом автоматической корреляции Дурбина-Уотсона, регионы с высоким сходством окрашены в синий цвет, в то время как непохожие кадры следуют за более темным блюзом и, наконец, красным в зависимости от тяжести непохожести. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 2: Участок интенсивности против вектора рассеяния.
Наложение вычитаемых данных SAXS формирует exoминус E9. В зеленых 5 кадрах (рамка 517-u2012522) усреднены и вычтены из области полуконюшенной Rg и синим цветом репрезентативная кривая рассеяния, полученная из деконволюции EFA пика SEC-SAXS. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 3: Бес измерение Kratky кривой.
Наложение deconvoluted (синий) и не-deconvoluted (зеленый) Kratky кривой, показывающей E9 exoминус является шаровой. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}

Рисунок 4: P (r) кривая.
Наложение deconvoluted (синий) и не-deconvoluted (зеленый) кривые для E9 exoминус. Пожалуйста, нажмите здесь, чтобы просмотреть большую версию этой цифры.
{kind=link}
Дополнительные данные. Пожалуйста, нажмите здесь, чтобы загрузить этот файл
Обсуждение
Перед началом эксперимента SAXS желательно иметь монодисперсную выборку, но в действительности многие коллекции данных не удовлетворяют этому и должны быть улучшены путем объединения измерения с рядной хроматографией-SEC в большинстве случаев. Однако даже нехватка времени между очисткой и монодисперсностью выборки в получении данных не гарантируется. Чаще всего это относится к экспериментам, в которых компоненты слишком близки по размеру или по своим физическим свойствам, чтобы быть разделенными или склонными к быстрой динамике. Здесь мы предоставили протокол, сочетающий разложение одного значения с развивающимся анализом факторов, чтобы удалить влияние экзо минуса DNAbound E9 из его нескончаемой формы, создавая монодисперсный профиль рассеяния, который мы смогли проанализировать с помощью пакета SAXS Scatter IV.
SVD с EFA данных SEC-SAXS являются очень мощными методами, разработанными для деконволюция данных SAXS и улучшения анализа, но они имеют ограничения. Они требуют, чтобы шум или дрейф в буферной базовой линии SEC-SAXS сведены к минимуму. Это может включать дополнительное эквивалентность столбца (лучше использовать более 3 объемов столбца, в зависимости от буфера) перед загрузкой выборки. Однако наиболее важным шагом является выбор количества единичных значений и диапазона используемых данных, поскольку это значительно повлияет на точность деконволюции. Именно по этой причине результаты не должны быть приняты сами по себе, но дальнейшее анализ с использованием таких методов, как аналитическая ультрацентрифугация (AUC) или многоугольный лазер-свет рассеяния (MALLS) для биологической интерпретации.
Scatter IV - это новый пакет программного обеспечения, бесплатный для исследований и промышленного использования с интуитивно понятным пользовательским интерфейсом, который позволяет даже неэкспертам анализировать свои данные. Scatter IV имеет несколько новых функций, которые помогают улучшить анализ данных SEC-SAXS, таких как тепловая карта, связанная с сюжетом сигнала, что позволяет повысить точность выбора кадра. В первичном анализе данных анализ Пика Гинье и график перекрестной проверки, связанный с анализом P(r), предлагают интегрированную способность устранения неполадок в программном обеспечении.
Следует отметить, что многие другие программы могут быть использованы для первичного анализа данных; они содержат те же основные функции, а также регулярно обновляются, такие как BioXTAS RAW17 ATSASпакет 24 и US-SOMO15, чтобы назвать несколько.
Но независимо от того, какой пакет SAXS используется для анализа, основные ограничения являются общими: подготовка выборки, до сбора и анализа. В примере E9 exominus ясно видеть улучшение соотношения сигнала к шуму и снижение rg dmax, связанное с монодисперсной выборкой. Это значительно поможет дальнейшей обработке данных, таких как примерка или моделирование с известными структурами высокого разрешения.
Раскрытие информации
Авторов нечего раскрывать.
Благодарности
Мы признаем финансовую поддержку проекта от французского гранта REPLIPOX ANR-13-BSV8-0014 и научно-исследовательские гранты от Службы Санте-де-Армаес и Делегоации G'an'rale за l'Armement. Мы благодарны ESRF за время пучка SAXS. В этой работе использовались платформы центра Grenoble Instruct-ERIC (ISBG; UMS 3518 CNRS-CEA-UGA-EMBL) в рамках Гренобльского партнерства по структурной биологии (PSB), при поддержке FRISBI (ANR-10-INBS-05-02) и GRAL, финансируемых в рамках университета Гренобль Альпы аспирантуру (Ecoles Universitaires de Recherche) CBH-EUR-GS (ANR-17-EURE-003). IBS признает интеграцию в Междисциплинарный научно-исследовательский институт Гренобля (IRIG, CEA). Мы благодарим Вима.Бурмейстера и Фредерика Исени за финансовуюи научную поддержку, а также благодарим д-ра Джесси Хопкинса из BioCAT в APS за его помощь и за разработку BioXTAS RAW.
Материалы
| Name | Company | Catalog Number | Comments |
| Beamline control software BsXCuBE | ESRF | Pernot et al. (2013), J. Synchrotron Rad. 20, 660-664 | local development |
| BioXTAS Raw 1.2.3. | MacCHESS | http://bioxtas-raw.readthedocs.io/en/latest/index.html | First developed in 2008 by Soren Skou as part of the biological x-ray total analysis system (BioXTAS) project. Since then it has been extensively developed, with recent work being done by Jesse B. Hopkins |
| HPLC program LabSolutions | Shimadzu | n.a. | |
| ISPyB | ESRF | De Maria Antolinos et al. (2015). Acta Cryst. D71, 76-85. | local development |
| NaCl | VWR Chemicals (BDH Prolabo) | 27808.297 | |
| Scatter | Diamond Light Source Ltd | http://www.bioisis.net/tutorial/9 | Supported by SIBYLS beamline (ALS berkeley, Ca) and Bruker Cororation (Karlsruhe, Germany) |
| Superdex 200 Increase 5/150 GL column | GE Healthcare | 28990945 | SEC-SAXS column used |
| Tris base | Euromedex | 26-128-3094-B |
Ссылки
- Pelikan, M., Hura, G., Hammel, M. Structure and flexibility within proteins as identified through small angle X-ray scattering. General Physiology and Biophysics. 28 (2), 174-189 (2009).
- Brosey, C. A., Tainer, J. A. Evolving SAXS versatility: solution X-ray scattering for macromolecular architecture, functional landscapes, and integrative structural biology. Current Opinion in Structural Biology. 58, 197-213 (2019).
- Gräwert, M., Svergun, D. A beginner's guide to solution small-angle X-ray scattering (SAXS). The Biochemist. 42 (1), 36-42 (2020).
- Putnam, C. D., Hammel, M., Hura, G. L., Tainer, J. A. X-ray solution scattering (SAXS) combined with crystallography and computation: defining accurate macromolecular structures, conformations and assemblies in solution. Quarterly reviews of biophysics. 40 (03), 191-285 (2007).
- Meisburger, S. P., et al. Domain Movements upon Activation of Phenylalanine Hydroxylase Characterized by Crystallography and Chromatography-Coupled Small-Angle X-ray Scattering. Journal of the American Chemical Society. 138 (20), 6506-6516 (2016).
- Brennich, M. E., Round, A. R., Hutin, S. Online Size-exclusion and Ion-exchange Chromatography on a SAXS Beamline. Journal of Visualized Experiments. (119), e54861 (2017).
- Watanabe, Y., Inoko, Y. Size-exclusion chromatography combined with small-angle X-ray scattering optics. Journal of Chromatography A. 1216 (44), 7461-7465 (2009).
- Graewert, M. A., et al. Automated Pipeline for Purification, Biophysical and X-Ray Analysis of Biomacromolecular Solutions. Scientific reports. 5, (2015).
- David, G., Pérez, J. Combined sampler robot and high-performance liquid chromatography: a fully automated system for biological small-angle X-ray scattering experiments at the Synchrotron SOLEIL SWING beamline. Journal of applied crystallography. 42 (5), 892-900 (2009).
- Ryan, T. M., et al. An optimized SEC-SAXS system enabling high X-ray dose for rapid SAXS assessment with correlated UV measurements for biomolecular structure analysis. Journal of Applied Crystallography. 51 (1), 97-111 (2018).
- Gampp, H., Maeder, M., Meyer, C. J., Zuberbühler, A. D. Calculation of equilibrium constants from multiwavelength spectroscopic data-III: Model-free analysis of spectrophotometric and ESR titrations. Talanta. 32 (12), 1133-1139 (1985).
- Maeder, M., Neuhold, Y. M. . Practical Data Analysis in Chemistry. , (2007).
- Tarbouriech, N., et al. The vaccinia virus DNA polymerase structure provides insights into the mode of processivity factor binding. Nature Communications. 8 (1), (2017).
- Brennich, M. E., et al. Online data analysis at the ESRF bioSAXS beamline, BM29. Journal of Applied Crystallography. 49 (1), (2016).
- Brookes, E., Rocco, M. Recent advances in the UltraScan SOlution MOdeller (US-SOMO) hydrodynamic and small-angle scattering data analysis and simulation suite. European Biophysics Journal. 47 (7), 855-864 (2018).
- Malaby, A. W., et al. Methods for analysis of size-exclusion chromatography-small-angle X-ray scattering and reconstruction of protein scattering. Journal of Applied Crystallography. 48 (4), 1102-1113 (2015).
- Hopkins, J. B., Gillilan, R. E., Skou, S. BioXTAS RAW: improvements to a free open-source program for small-angle X-ray scattering data reduction and analysis. Journal of Applied Crystallography. 50 (5), 1545-1553 (2017).
- Maeder, M. Evolving factor analysis for the resolution of overlapping chromatographic peaks. Analytical Chemistry. 59 (3), 527-530 (1987).
- Durand, D., et al. NADPH oxidase activator p67phox behaves in solution as a multidomain protein with semi-flexible linkers. Journal of Structural Biology. 169 (1), 45-53 (2010).
- De Maria Antolinos, A., et al. ISPyB for BioSAXS, the gateway to user autonomy in solution scattering experiments. Acta Crystallographica Section D. 71 (1), 76-85 (2015).
- Brennich, M. E., et al. Online data analysis at the ESRF bioSAXS beamline, BM29. Journal of Applied Crystallography. 49 (1), 203-212 (2016).
- Kirby, N., et al. Improved radiation dose efficiency in solution SAXS using a sheath flow sample environment. Acta Crystallographica Section D Structural Biology. 72 (12), 1254-1266 (2016).
- Rambo, R. P., Tainer, J. A. Characterizing flexible and intrinsically unstructured biological macromolecules by SAS using the Porod-Debye law. Biopolymers. 95 (8), 559-571 (2011).
- Franke, D., et al. ATSAS 2.8: a comprehensive data analysis suite for small-angle scattering from macromolecular solutions. Journal of Applied Crystallography. 50 (4), 1212-1225 (2017).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеThis article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены