Method Article
Relating Stomatal Conductance to Leaf Functional Traits
In This Article
Summary
Unraveling how physiology and morphology are linked allows a deeper understanding of mechanistic functioning of plant leaves. We present both a procedure to derive parameters of stomatal regulation from stomatal conductance measurements and correlations with traditional functional leaf traits.
Abstract
Leaf functional traits are important because they reflect physiological functions, such as transpiration and carbon assimilation. In particular, morphological leaf traits have the potential to summarize plants strategies in terms of water use efficiency, growth pattern and nutrient use. The leaf economics spectrum (LES) is a recognized framework in functional plant ecology and reflects a gradient of increasing specific leaf area (SLA), leaf nitrogen, phosphorus and cation content, and decreasing leaf dry matter content (LDMC) and carbon nitrogen ratio (CN). The LES describes different strategies ranging from that of short-lived leaves with high photosynthetic capacity per leaf mass to long-lived leaves with low mass-based carbon assimilation rates. However, traits that are not included in the LES might provide additional information on the species' physiology, such as those related to stomatal control. Protocols are presented for a wide range of leaf functional traits, including traits of the LES, but also traits that are independent of the LES. In particular, a new method is introduced that relates the plants’ regulatory behavior in stomatal conductance to vapor pressure deficit. The resulting parameters of stomatal regulation can then be compared to the LES and other plant functional traits. The results show that functional leaf traits of the LES were also valid predictors for the parameters of stomatal regulation. For example, leaf carbon concentration was positively related to the vapor pressure deficit (vpd) at the point of inflection and the maximum of the conductance-vpd curve. However, traits that are not included in the LES added information in explaining parameters of stomatal control: the vpd at the point of inflection of the conductance-vpd curve was lower for species with higher stomatal density and higher stomatal index. Overall, stomata and vein traits were more powerful predictors for explaining stomatal regulation than traits used in the LES.
Introduction
To advance the functional understanding of plant leaves, many recent studies have attempted to relate morphological, anatomical and chemical leaf traits to physiological responses, such as leaf stomatal conductance (gS) 1-4. In addition, to leaf traits, stomatal conductance is strongly affected by environmental conditions, such as photosynthetically active photon flux density, air temperature and vpd 5. Various ways have been proposed to model gs - vpd curves6-8 which are mainly based on linear regression of gs on vpd 6. In contrast, the model presented in this study regresses the logits of relative stomatal conductance (i.e., the ratio of gS to maximum stomatal conductance gSMAX) on vpd and accounts for the non-linearity by adding vpd as a quadratic regressor term.
Compared to other models, the new model allows for deriving parameters that describe the vpd at which gS is down-regulated under water shortage. Similarly, the vpd is obtained at which gS is maximal. As such physiological parameters can be expected to be tightly linked to carbon assimilation 9,10 a close link between these model parameters and the key leaf traits to nutrient and resource allocation as reflected in the LES should be expected 3,11. In consequence, there should be also a tight relationship between strategies of stomatal regulation with LES traits. Such a relationship is expected in particular for leaf habit (evergreen versus deciduous) as leaf habit is both correlated with the LES and with water use efficiency12,13. Evergreen species tend to grow slower, but are more efficient in environments poor in nutrients 14. Thus, leaf habit should translate into differing stomatal regulation patterns, with a more conservative water use strategy than deciduous species.
Comparing a large set of broad-leaved tree species in a common garden situation, the following hypotheses were tested: 1) Model parameters from gS - vpd models are connected to leaf traits related to the leaf economics spectrum. 2) Evergreen species have lower mean gS and gSMAX values than deciduous species.
Protocol
1. Stomatal Conductance
- Measurements of stomatal conductance
NOTE: The authors made use of a simple type of steady state porometer (Decagon SC1). The porometer’s design has the advantage of small size, intuitive manual operation and high reliability. When measuring stomatal conductance in the field, make sure to minimize the distance between measured individuals for a repeated measurement cycle to be the most efficient possible. - Choose leaves of different species and individuals according to a reproducible pattern (same height, same exposure, same position within the plant, if possible only from the same node, and only from one category; sun or shade leaves, etc.).
- Only measure leaves in healthy, non-damaged and fully developed condition. Mark the leaves on the individuals (e.g., with cable ties or color tape), to ensure that the repeated measurements are done on the same leaf.
NOTE: Measurements of stomatal conductance should be made only on leaf surface whilst avoiding the midrib and strong leaf veins. - Start measurements in the early morning hours before sunrise taking five to ten repeated measurements until stomatal conductance values shows a clear decline at noon.
NOTE: A daily course of measurements will deliver good data for analyzing the relationships between vpd and stomatal conductance.
- Only measure leaves in healthy, non-damaged and fully developed condition. Mark the leaves on the individuals (e.g., with cable ties or color tape), to ensure that the repeated measurements are done on the same leaf.
- Vpd measurements
- With each gS measurement, record temperature and relative humidity preferably with portable loggers to directly measure the conditions at the position of the same leaf. For calculating the vapor pressure deficit use the August-Roche Magnus formula15.

es=saturation water vapor pressure [hPa]
T=temperature [°C]
- Model gS - vpd response
- Now plot species-wise all gS data against vpd, combining all daily courses of individual leaves into one analysis per species. Extract the maximum value observed from the stomatal conductance data by searching for the maximum value. To scale the model for species-wise comparability, divide the observed values through the maximum value observed for that species (gS/gSMAX).
- For each species, regress the logits of gS (gS/gSMAX) to vpd and the quadratic term of vpd using a generalized linear model with a binomial error distribution (a, b and c represent regression parameters):


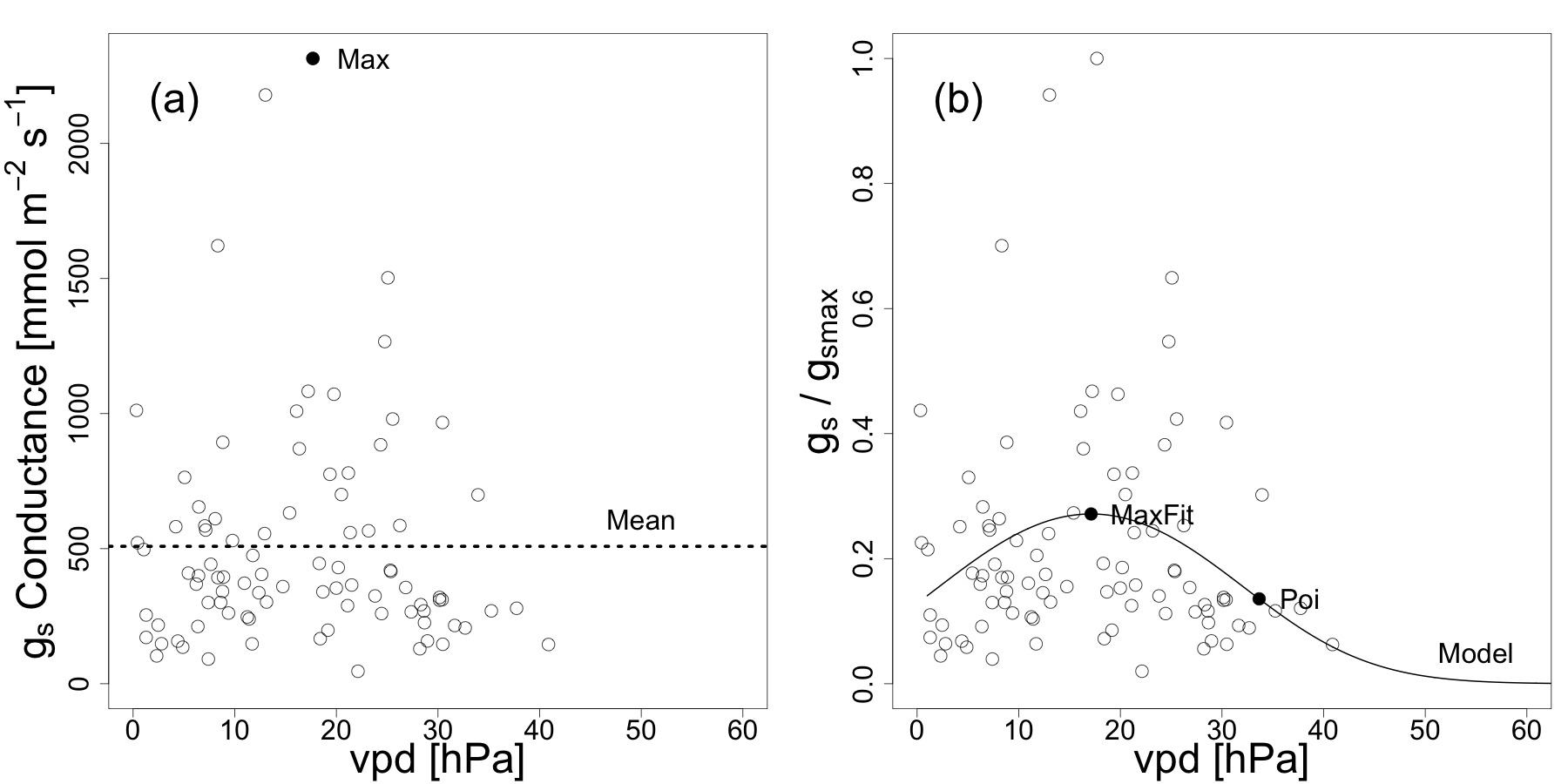
Figure 1. Example of the plotted values and the fitted model for gS - vpd. Stomatal conductance plotted as a function of vapor pressure deficit for the species Liquidambar formosana. Empty dots represent the observed values. (A) Maximum stomatal conductance, the vpd at maximum stomatal conductance and mean stomatal conductance was extracted from the absolute thus non-scaled stomatal conductance (gS) data. (B) Scaled stomatal conductance data (gS/gSMAX) were plotted to extract the relative parameters (shown as filled dots) 20. Re-print with permission from20. Please click here to view a larger version of this figure.
{kind=link}

Figure 2. All fitted models for all species. Model graphs for the stomatal conductance data to vpd regression for all species. Evergreen species are represented by black lines, deciduous species by red lines 20. Re-print with permission from20.
NOTE: Using the logits instead of regressing gS directly to vpd, leads to the point that modeled maximum values do not exceed gSMAX and that gS approaches 0 at high vpd.
- Extract parameter of stomatal regulation for every species
- Calculate absolute modeled gSMAX values (MaxFit in Figure 1B). To do this, calculate the vpd at maximum stomatal conductance from setting the first derivative of 1.4.2 to zero, which gives vpdgsMaxFit = -b/2a. Insert vpdgsMaxFit into the formula of 1.4.2 and raise to the power of e to obtain MaxFit. Calculate the mean of all conductance measurements per species (see Figure 1A).
NOTE: Use statistical software R (http://www.r-project.org). - To calculate relative values, from the scaled model (gS/gSMAX), extract stomatal conductance and vpd values for the following two points: (1) stomatal conductance and vpd at the maximum of the model (MaxFit) and (2) vpd at the second point of inflection of the curve (see Figure 1B). Multiply these values by gSMAX to obtain absolute gS values for these points. See Figure 2 for a complete overlay of the single models of all 39 species analyzed.
- Calculate absolute modeled gSMAX values (MaxFit in Figure 1B). To do this, calculate the vpd at maximum stomatal conductance from setting the first derivative of 1.4.2 to zero, which gives vpdgsMaxFit = -b/2a. Insert vpdgsMaxFit into the formula of 1.4.2 and raise to the power of e to obtain MaxFit. Calculate the mean of all conductance measurements per species (see Figure 1A).
2. Measurements of Stomatal Traits
- Take samples preferably from exactly the same leaves that have been used for the measurements of stomatal conductance. If this is not possible, apply the same selection procedure that was applied to choose the leaves for the stomatal conductance measurements, preferably on the same individuals.
- Apply a thin layer of colorless, quickly drying nail polish (test different makes, some are more suitable than others) to a fresh sample. If the samples cannot be processed immediately, store them in 70% alcohol. After the nail polish has dried out, gently peel the impression of from the leaf and proceed for the microscopic analysis, as with a normal leaf sample.
NOTE: In the case of leaves with high trichome density a preceding step of applying sodium hydroxide or a 1:1 solution of acetic acid and oxygen peroxide can lead to better results. - Optical microscope analytics
- Connect a camera to an optical microscope capable of magnifications between 40X to 400X. After the camera is connected to the microscope, match the pictures taken to the optical magnification and resolution of the picture, e.g., with the help of a scale.
- Employing open source image processing software like ImageJ 16 analyze these pictures.
- Draw a shape with the shape tool from the image analysis tool on the image in an area with no dirt, thumb prints, damaged areas, or large leaf veins. Count stomata in this area and in total of at least 50,000 µm2 per sample.
- Measure stomatal guard cell length and pore length. Calculate the number of stomata per mm2. Calculate stomatal index as the ratio of stomatal density to epidermal cell number per mm2.
3. Assessment of Leaf Vein Traits
NOTE: For the assessment of leaf vein traits, a modified a protocol from Sack & Scoffoni 17 was employed.
- Sample preparation
NOTE: To optimize the visibility of the leaf veins, the leaves are first bleached and then stained with safranine and malachite green.- To bleach the leaves, leave them at least 72 hr in 50% solution of decolorizer (test different makes, again some might work better). Alternatively, use 5% NaOH solution or 10% KOH solution or 25% H2O2 solution.
- Heat the solution up to 30° C or combine the different solutions from step 3.1.1 for better effects. Rinse several times in water afterwards. Adapt the bleaching process for the specific species, depending on their leaf characteristics.
NOTE: Thicker leaves may need longer periods of solution bath and or the more aggressive solutions. Thinner and more tender leaves may be bleached to a satisfactorily degree in less than 72 hr. - To color the leaves place them in 100% ethanol. Color them for 2 – 30 min in 1% safranine solution. To enhance coloring, add an additional treatment with 1% malachite green solution only for several seconds. Adapt the protocol to every species in terms of timing and intensity to achieve optimal results. Rinse several times in water afterwards. If the leaves are to deeply stained, some time in ethanol or decolorizer may help.
- Sample analysis
- Scan the leaves with a backlight scanner at a resolution of ca. 1,200 dpi. Match the scans taken to the resolution of the picture, e.g., with the help of a scale, to ensure that pixel length can be traced back to absolute length measures of the scanned leaf.
- Measure area, circumference, length and width of the leaves. Calculate several indices, e.g., length/width and circumference2/area. Cut a 1 by 1 cm rectangle out of the middle of the picture. Measure diameter of the veins of first and second order (do not include the main middle vein). Measure the length of all veins of first order in this quadrat (vein density).
4. Assessment of Other Leaf Traits
NOTE: Asses typical leaf traits such as specific leaf area (SLA), leaf dry matter content (LDMC), leaf area, element contents, leaf habit, leaf pinnation, leaf compound type, leaf margin type etc. following established protocols 18,19.
- Observational leaf traits
- Assess leaf pinnation, leaf compound type, leaf margin type and the presence of extrafloral nectaries18,19 through observation in the field.
- Analytical leaf traits
- Collect fresh leaf samples, preferably on the same individuals as used for the other measurements, for the determination of specific leaf area (SLA), leaf area and leaf dry matter content (LDMC) 18,19. After 48 hr in the drying oven at 80 °C, measure element contents and ratios, preferably on the same leaves.
Results
Many parameters of stomatal conductance and stomatal regulation were found to be related to morphological, anatomical and chemical leaf traits. In the following, the focus will be on the links to vpd at the point of inflection, which decreased with stomatal density (p=0.04) and stomatal index (p=0.03) and increased with leaf carbon content (p=0.02, See Figure 3). The results show that with decreasing vpd at the point of inflection there was a decrease in stomatal density and stomatal index. In contrast, no parameter of stomatal conductance showed a clear relationship to leaf habit. The high variation within the two groups of leaf habit shows that different regulatory mechanisms exist both within the group of evergreen and deciduous leaf habits.

Figure 3. Result figure for the links between patterns of stomatal conductance regulation and leaf traits. Vapor pressure deficit (vpd) at the point of inflection of the gS - vpd curve (VpdPoi) as a function of (A) stomata density, (B) stomata index, and (C) leaf carbon content 20. Re-print with permission from20. Please click here to view a larger version of this figure.
{kind=link}
Discussion
The parameters of stomatal regulation extracted by the method presented in this paper underline the importance of stomatal traits, such as stomata density and stomata index. These novel relationships demonstrate the potential of linking parameters from physiological models to morphological, anatomical and chemical leaf traits 20. Compared to other methods, the present approach bears the advantage of capturing a unique and unequivocal vpd value at which stomatal conductance is down-regulated to half of the modeled gs maximum.
Of all steps outlined in the protocol the most critical ones are the measurements of stomatal conductance. Due to the multifactorial regulation of stomatal conductance ambient weather conditions have a strong influence on gS. Stomatal conductance measurements at high relative humidity and low light intensity may be unreliable 21-23. With respect to morphological and anatomical traits, the protocol should always be adapted to the target species included in the study. In particular in the vein density analysis, the duration of bleaching and staining of leaves should be varied, depending on leaf structure and toughness. Potential limitations of the method include species, for which measurements of stomatal conductance are impossible or complicated and prone to error owing to extraordinary leaf forms. This may include conifers and grasses with very narrow leaf blades.
Our results partly confirm the first hypothesis of a link between stomatal conductance parameters and leaf traits of the leaf economics spectrum (LES), which corresponds to several other studies. For example, Poorter and Bongers (2006)24 reported a close link between gS and traits represented by the LES, e.g., with gS decreasing with increasing leaf lifespan. Accordingly, Schulze et al. (1994) 1 demonstrated clear links between leaf nitrogen content and gSMAX. Similarly, Juhrbandt et al. (2004) 25 found significant relationships between gSMAX and leaf area and leaf nitrogen content.
Our second hypothesis of clear differences with regard to leaf habit could not be confirmed. The high variation in the measured parameters and traits within evergreen and deciduous leaf habit indicate that leaf habit is not a good descriptor of the LES. Brodribb and Holbrook (2005) 26 discussed that leaf habit and leaf physiological strategies may not be inevitably connected since broad trait variation is common in all types of leaf habit.
The approach may be extended to traits and physiological characteristics of plant organs other than leaves, for example to traits related to xylem hydraulics such as specific xylem hydraulic conductivity and microscopy wood traits 27. Similarly, other types of leaf traits as derived from microscopy such as palisade parenchyma structure and epicuticular wax layer structure could be included 28.
In summary, this study confirmed the close connection between the LES and stomatal regulation. In addition, the method presented here revealed facets of stomatal regulation patterns that are not related to the LES. Especially specific leaf traits such as stomata size, density and index as well as vein length deserve future attention in functional plant studies.
Disclosures
The authors have nothing to disclose.
Acknowledgements
We are indebted to Xuefei Yang, Sabine Both, Lin Chen and Kaitian Wang for coordinating the fieldwork and establishing the BEF-China experiment. We are also grateful to the whole BEF-China research group for their general support. BEF-China is mainly funded by the German Research Foundation (DFG FOR 891/1 and 2) and funding for this particular project was provided by the German Research Foundation to H.B. (DFG BR 1698/9-2). We are also thankful for the travel grants and summer schools financed by the Sino-German Centre for Research Promotion in Beijing (GZ 524, 592, 698, 699 and 785). In addition we would like to thank David Eichenberg, Michael Staab, Katja Grotius, Silvana Tornack, Lin Chen, and Shouren Zhang for their support in the field and in the lab.
Materials
| Name | Company | Catalog Number | Comments |
| SC 1 Porometer | Decagon | NA | Any other porometer is suitable |
| Cable ties to mark leaves | NA | NA | NA |
| Plastic sample bags | NA | NA | NA |
| Paper sample bags | NA | NA | NA |
| Hygrometer | Trotec | NA | Any other is suitable |
| Nail polish | NA | NA | NA |
| Axioskop 2 plus | Zeiss | NA | Any other is suitable |
| Ethanol | NA | NA | NA |
| Bleach | NA | NA | NA |
| 5% NaOH | NA | NA | NA |
| 10% KOH | NA | NA | NA |
| 25% H2O2 | NA | NA | NA |
| Malachite green | NA | NA | NA |
| Safranine | NA | NA | NA |
References
- Schulze, E. -. D., Kelliher, F., Körner, C., Lloyd, J., Leuning, R. Relationships among maximum stomatal conductance, ecosystem surface conductance, carbon assimilation rate, and plant nitrogen nutrition: a global ecology scaling exercise. Annual Review of Ecology and Systematics. 25 (1), 629-662 (1994).
- Chaturvedi, R. K., Raghubanshi, A. S., Singh, J. S. Growth of tree seedlings in a tropical dry forest in relation to soil moisture and leaf traits. Journal of Plant Ecology. 6 (2), 158-170 (2013).
- Santiago, L. S., Kim, S. C. Correlated evolution of leaf shape and physiology in the woody Sonchus. alliance (Asteraceae: Sonchinae) in Macaronesia. International Journal of Plant Sciences. 170 (1), 83-92 (2009).
- Aasamaa, K., Sober, A. Hydraulic conductance and stomatal sensitivity in six deciduous tree species. Biologia Plantarum. 44 (1), 65-73 (2001).
- Waring, R. H., Chen, J., Gao, H. Plant-water relations at multiple scales: integration from observations, modeling and remote sensing. Journal of Plant Ecology-UK. 4, 1-2 (2011).
- Oren, R., Sperry, J. Survey and synthesis of intra- and interspecific variation in stomatal sensitivity to vapour pressure deficit. Plant, Cell and Environment. 22 (12), 1515-1526 (1999).
- Leuning, R. A critical appraisal of a combined stomatal-photosynthesis model for C3 plants. Plant, Cell and Environment. 18 (4), 339-355 (1995).
- Patanè, C. Leaf area index, leaf transpiration and stomatal conductance as affected by soil water deficit and vpd in processing tomato in semi arid mediterranean climate. Journal of Agronomy and Crop Science. 197 (3), 165-176 (2011).
- Jarvis, A. J., Davies, W. J. The coupled response of stomatal conductance to photosynthesis and transpiration. Journal of Experimental Botany. 49 (Special Issue), 399-406 (1998).
- Roelfsema, M. R. G., Hedrich, R. In the light of stomatal opening: new insights into “the Watergate.”. New Phytologist. 167 (3), 665-691 (2005).
- Wong, S. C., Cowan, I. R., Farquhar, G. D. Stomatal conductance correlates with photosynthetic capacity. Nature. 282 (5737), 424-426 (1979).
- Sobrado, M. A. Hydraulic conductance and water potential differences inside leaves of tropical evergreen and deciduous species. Biologia Plantarum. 40 (4), 633-637 (1998).
- Zhang, Y. -. J., Meinzer, F. C., Qi, J. -. H., Goldstein, G., Cao, K. -. F. Midday stomatal conductance is more related to stem rather than leaf water status in subtropical deciduous and evergreen broadleaf trees. Plant, Cell and Environment. 36 (1), 149-158 (2013).
- Aerts, R. The advantages of being evergreen. Trends in Ecology, & Evolution. 10 (10), 402-407 (1995).
- Murray, F. W. On the computation of saturation vapor pressure. Journal of Applied Meteorology. 6 (1), 203-204 (1967).
- Scoffoni, C., Sack, L. Quantifying leaf vein traits. PrometheusWiki. , (2011).
- Cornelissen, J. H. C., Lavorel, S., et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Australian Journal of Botany. 51 (4), 335-380 (2003).
- Perez-Harguindeguy, N., Díaz, S., et al. New handbook for standardised measurement of plant functional traits worldwide. Australian Journal of Botany. 61 (3), 167-234 (2013).
- Kröber, W., Bruelheide, H. Transpiration and stomatal control: A cross-species study of leaf traits in 39 evergreen and deciduous broadleaved subtropical tree species. Trees. 28 (3), 901-914 (2014).
- Larcher, W. . Physiological plant ecology. , 514 (2003).
- Bunce, J. Does transpiration control stomatal responses to water vapour pressure deficit. Plant, Cell and Environment. 20 (1), 131-135 (1997).
- Bunce, J. A. How do leaf hydraulics limit stomatal conductance at high water vapour pressure deficits. Plant, Cell and Environment. 29 (8), 1644-1650 (2006).
- Poorter, L., Bongers, F. Leaf traits are good predictors of plant performance across 53 rain forest species. Ecological Sciences of America. 87 (7), 1733-1743 (2006).
- Juhrbandt, J., Leuschner, C., Hölscher, D. The relationship between maximal stomatal conductance and leaf traits in eight Southeast Asian early successional tree species. Forest Ecology and Management. 202 (1-3), 245-256 (2004).
- Brodribb, T. J., Holbrook, N. M. Leaf physiology does not predict leaf habit; examples from tropical dry forest. Trees. 19 (3), 290-295 (2005).
- Kröber, W., Zhang, S., Ehmig, M., Bruelheide, H. Xylem hydraulic structure and the Leaf Economics Spectrum - A cross-species study of 39 evergreen and deciduous broadleaved subtropical tree species. PLOS ONE. 9 (11), e109211 (2014).
- Kröber, W., Heklau, H., Bruelheide, H. Leaf morphology of 40 evergreen and deciduous broadleaved subtropical tree species and relationships to functional ecophysiological traits. Plant Biology. 17 (2), 373-383 (2015).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved