
Method Article
A Mouse Model of Chronic Liver Fibrosis for the Study of Biliary Atresia
In This Article
Summary
We established a mouse model of chronic liver fibrosis, which provides a suitable animal model for the virus-induced liver fibrosis mechanistic study of biliary atresia (BA) and a platform for future BA treatments.
Abstract
Biliary atresia (BA) is a fatal disease involving obstructive jaundice, and it is the most common indication for liver transplantation in children. Due to the complex etiology and unknown pathogenesis, there are still no effective drug treatments. At present, the classic BA mouse model induced by rhesus rotavirus (RRV) is the most commonly used model for studying the pathogenesis of BA. This model is characterized by growth retardation, jaundice of the skin and mucosa, clay stools, and dark yellow urine. The histopathology shows severe liver inflammation and obstruction of the intrahepatic and extrahepatic bile ducts, which are similar to the symptoms of human BA. However, the livers of end-stage mice in this model lack fibrosis and cannot fully simulate the characteristics of liver fibrosis in clinical BA. The presented study developed a novel BA mouse model of chronic liver fibrosis by injecting 5-10 µg of anti-Ly6G antibody four times, with gaps of 2 days after each injection. The results showed that some of the mice successfully formed chronic BA with typical fibrosis after the time lapse, meaning these mice represent a suitable animal model for the virus-induced liver fibrosis mechanistic study of BA and a platform for developing future BA treatments.
Introduction
Biliary atresia (BA) is a serious hepatobiliary disease that often occurs in infants and young children; specifically, it presents as an obliterative cholangiopathy with neonatal jaundice and pale stools1. Its clinical characteristics are inflammatory destruction of the intrahepatic and extrahepatic bile ducts and progressive fibrosis, which eventually develops into liver failure2. According to statistics, the incidence of BA in Asian countries is higher than in European and American countries, and the incidence of BA in Asian countries is 1/8,000. The etiology of BA includes viral infection, abnormal bile duct development, immune disorders, and genetic variations3. Kasai surgery is the most commonly used method to improve cholestasis in children with BA, but ultimately, this cannot prevent the progression of fibrosis4. The current treatment for BA mainly relies on liver transplantation, which is limited by the lack of liver sources. An in-depth study of the pathogenesis of BA is the most direct means to resolve the challenges of this disease. However, studies on the pathogenesis of BA mainly rely on BA animal models, and it is crucial to select an appropriate animal model.
Most of the histopathological studies of BA have shown that bile duct hyperplasia (BDP), bile thrombosis, and portal vein fibrosis are the most important pathological features of BA, and other pathological features of different grades exist at the same time, such as portal inflammatory cell infiltration and hepatocyte swelling5,6. At present, there are a variety of mouse models that mimic BA, such as the chronic 3,5-die-thoxycarbonyl-1,4-dihydrocollidine (DDC)-fed mouse model with segmental bile duct obstruction and pericholangitis involving the extrahepatic bile ducts7 and the mouse model of liver fibrosis with an intraperitoneal injection of carbon tetrachloride8. The bile duct ligation (BDL) model is characterized by jaundice and rapid portal vein fibrosis9. The alpha-naphthylisothiocyanate (ANIT)-fed mouse model presents with cholangitis confined to the intrahepatic bile duct and hepatocyte injury10. A mouse model with prolonged jaundice is induced by delayed rhesus rotavirus (RRV) inoculation11. Although there are a variety of animal models, especially mouse models, each model has its own limitations. They can only simulate part of the disease characteristics of BA, such as acute inflammation or liver fibrosis in the process of biliary atresia, and there is no model that is highly consistent with the disease process and pathological features of BA.
The classic BA mouse model is induced by RRV, and this model is the most similar model to BA in humans. However, while RRV-induced BA model mice show similar clinical symptoms and pathological characteristics to some of those of extrahepatic biliary atresia, this model lacks liver fibrosis, which greatly limits the in-depth study of the BA mechanisms and the development of new treatments12. Therefore, this study developed a novel BA mouse model of chronic liver fibrosis. Anti-Ly6G antibody was intraperitoneally injected before RRV inoculation on the day of birth. Then, 5-10 µg of anti-Ly6G antibody was injected four times, with gaps of 2 days between each injection. The BA symptoms of mice were improved, the survival time was prolonged, and the mice entered the chronic fibrosis stage. This model not only simulates the acute phase response of BA but also mimics the processes of the liver and progressive fibrosis. Thus, it is a more suitable animal model for the mechanistic study of BA, and it could provide a theoretical basis for developing future treatments for BA.
The Gr-1 molecule is a myeloid-derived cell surface marker originally found to be expressed in neutrophils13. The depletion of anti-Ly6G antibodies reduces circulating neutrophils by more than 90% and alters the response activated by other immune cells. The functions of Gr-1+ cell populations have been reported in various studies using specific scavenging antibodies, with varying effects identified on cytokines and mediated immune protection14. We have studied the function of Gr-1+ cells in RRV-inoculated BA mouse models. However, since the Gr-1 molecule has not been found to be expressed in humans, a similar molecule, CD177, has been studied in BA patients15. Our data prove the significance of Gr-1+ cell populations, particularly in the chronic fibrotic phase of the disease, and provide a suitable animal model to investigate potential BA therapies.
Protocol
This study was approved by the Institutional Animal Care and Use Committee of Guangzhou Yongnuo Biomedical Animal Center (IACUC-AEWC-F2208020), where all the animal experiments were performed.
1. Establishment of the chronic fibrotic BA mouse model
NOTE: All the animals were kept in a specific pathogen-free (SPF) environment in the same room, and the experiments were conducted in a conventional environment. BALB/c mice on day 12.5 of gestation (age: 10-12 weeks; weight 35-40 g) were kept in a room without specific pathogens at 25°C under a 12 h dark/light cycle and were provided with free access to autoclaved food and water. Neonatal mice, within 24 h after birth (average body weight: 1.5-1.6 g), were selected for the mouse BA model.
- Prepare RRV as previously reported16.
NOTE: Due to the poor survival status of the model mice, they need to be separated at 21 days old to avoid them being bitten or even killed by other mice in the same cage. - Divide neonatal mice into three groups: the control group, the RRV group, and the RRV + anti-Ly6G group. Intraperitoneally inject the neonatal mice with 20 µL of RRV (titer: 1.5 x 106 PFU/mL) (RRV group) or saline (control group) within 24 h after birth. To deplete Gr-1+ cells, pretreat each mouse with an intraperitoneal injection of 5 µg of anti-Ly6G antibody 4 h before the RRV injection.
NOTE: The anti-Ly6G antibody solution is stored at 2-8 °C and should not be frozen. Take the antibody out before use and put it at room temperature for 30 min to warm up. - Inject 10 µg of anti-Ly6G antibody into the abdomen of the mouse every 3 days until day 12 after the RRV injection (Figure 1A).
- Check and record the appearance, weight, and survival of all the mice daily.
2. Intraperitoneal injection of the mice
- Remove newborn mice from the cage. Use a 1 mL insulin syringe to inject 20 µL of RRV solution or 50 µL of anti-Ly6G antibody solution. Pinch the neck skin of the young mouse with the index finger and thumb of one hand, and gently hold the hind legs of the mouse with the ring finger and tail finger to expose the abdomen.
- Lift the needle in an upward incline. Insert the needle into the middle thigh of the right hind leg of the mouse at an angle of 15° with the skin. After moving along the subcutaneous path until the needle reaches the right costal edge of the mouse, direct the needle downward into the abdominal cavity. Then, inject the liquid under the liver of the mouse.
NOTE: Newborn mice have their stomach and spleen in the left abdomen. If a needle is inserted on the left side, it is easy to pierce the stomach or cause splenic hemorrhage. - Pull the needle out immediately after the injection, and observe for bleeding or leakage at the injection site. If there is any, wipe it with an autoclaved cotton. Return the mouse to its mother's cage.
NOTE: In order to reduce the influence of fluid leakage during injection on the experimental results, ensure the injection action is gentle, slowly remove the needle, and press the injection site with a cotton swab for 30 s.
3. Collection of sample tissue
- On day 12, anesthetize the mice with 4% isoflurane inhalation, and dissect them under a microscope. Insert a 1 mL insulin syringe into the left ventricle of the heart to collect blood. After blood collection, euthanize the mice by inhalation of 4% isoflurane for 10 min. Centrifuge the blood at 400 × g for 5 min at room temperature, and separate the serum for liver function measurements.
NOTE: Blood collection must be performed while the mice are alive. If the mice die, the blood will be retained in the blood vessels and can not be collected. - Photograph the general appearance of the liver and bile duct. Subsequently, dissect the mouse liver and spleen from the surrounding tissues with scissors and tweezers.
NOTE: The liver and other tissues are collected and reserved at −80°C for RNA and protein extraction or soaked in 10% formalin for the preparation of histological specimens.
4. Fluorescein angiography of the extrahepatic bile duct
- After euthanizing the mouse, entirely expose the liver, gallbladder, and extrahepatic bile ducts with scissors and cotton swabs.
- Observe under a posture microscope, inject 5-10 µL of the fluorescent dye rhodamine 123 (20 mg/mL) into the gallbladder with a 1 mL insulin syringe, and take pictures. This process is the same as that reported in a previously16.
NOTE: Different mice in the same group are used for the sample tissue collection and fluorescence angiography.
5. H&E staining
- Immerse fresh mouse liver tissue in 10% formalin for 24 h.
- After embedding the tissue in paraffin, use a paraffin microtome to cut the paraffin block into sections with a thickness of 4 µm, and place two consecutive sections on the same slide. Experienced personnel are required to standardize the operation to avoid finger cuts17.
- Place the slices into a slicing rack, dewax them in xylene, hydrate them successively in absolute ethanol, 95% ethanol, 80% ethanol, 70% ethanol, and distilled water, and soak in each for 5 min.
- Stain the sections with hematoxylin solution for 5 min, and soak them in 1% hydrochloric acid and 75% alcohol for 5 s. Rinse the sections with clean water, and stain with eosin solution for 1 min.
6. CK19 and F4/80 immunohistochemical staining
- The first three steps are the same as the steps of tissue embedding, sectioning, and dewaxing in the H&E staining section.
- Perform antigen repair with Tris-EDTA buffer (10 mmol/L Tris base, 1 mmol/L EDTA solution, pH 9.0), heat the sections in a microwave oven at 95°C for 10 min, and then remove and cool them naturally to room temperature.
- Place the tissue sections in 3% hydrogen peroxide solution for 10 min to remove endogenous peroxidase.
- Treat the slices with 5% goat serum to block nonspecific binding.
- Add primary rat anti-mouse cytokeratin 19 or rat anti-mouse F4/80 monoclonal antibody to the sections, and incubate overnight at 4 °C.
- Incubate the sections with appropriate secondary antibodies for 30 min at room temperature.
- Use 3,3'-diaminobenzidine (DAB) as a chromogenic agent to observe the chromogenic reaction under the microscope.
- Observe the slices under a 40x microscope to obtain pictures, and analyze them as needed.
7. Sirius Red staining
- Perform the first three steps of tissue embedding, sectioning, and dewaxing as described in the H&E staining section.
- After the tissue sections are counterstained with hematoxylin, cover each tissue with 50 µL of Sirius Red dye solution at room temperature for 1 h.
- Dry the slides naturally at room temperature for 4 h, add a drop of neutral gum to each slide, and use a coverslip to slowly cover the tissue to avoid bubbles. Leave the slides at room temperature for 24 h to solidify the neutral gum.
- Use a polarized contrast light microscopy to observe the details of collagen deposition; select a clear and suitable field of view, and adjust the brightness and white balance of the microscope's visual field. Obtain images, and analyze them as needed under a 40x microscope.
Results
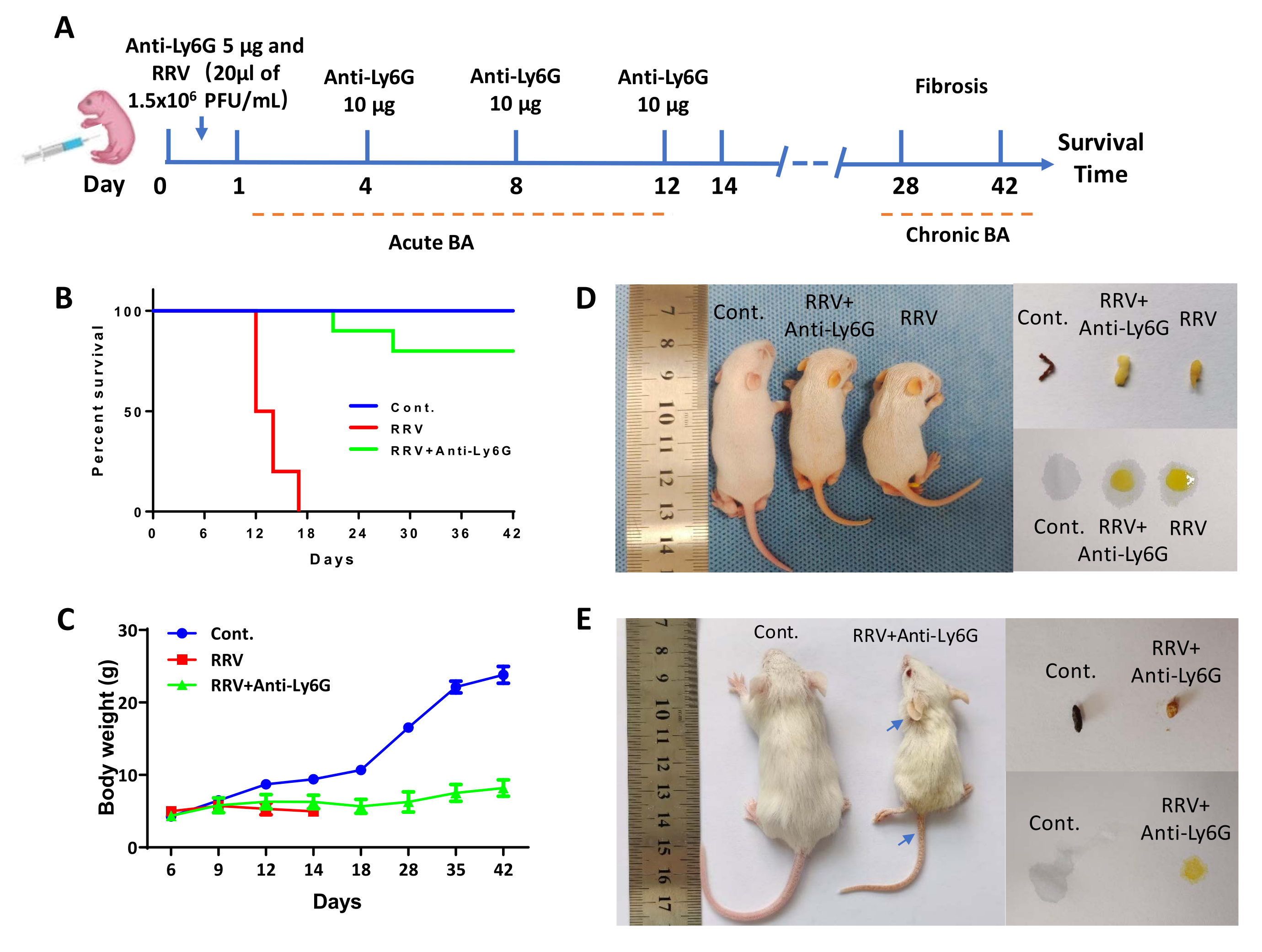
The mice were intraperitoneally injected with 5 µg of anti-Ly6G antibody within 24 h of birth and then intraperitoneally injected with 20 µL of RRV 4 h later. Then, 10 µg of anti-Ly6G antibody was injected every 3 days until day 12 (Figure 1A). The median survival time in the RRV group was 13 days. On the contrary, most of the mice treated with the antibody developed mild jaundice, and no weight loss was observed (Figure 1C). About 20%-30% of the mice had BA syndrome with long-term jaundice and low body weight but survived for more than 42 days. After anti-Ly6G antibody treatment, the number of Gr-1+ cells decreased15, and the mice entered the stage of chronic fibrosis, termed chronic BA. Specimens were collected on day 12 and day 42, and tissue slices stained with Sirius Red showed a gradual increase in liver fibrosis. The chronic BA mice surviving for 42 days were used for detailed analyses. In addition to the small size, the ears, feet, and tail skin showed marked jaundice (blue arrows in Figure 1E). On day 6 after the RRV injection, the feces color of the mice became lighter, and the urine color became darker; this became significantly different from the control group on day 12, when the RRV group mice showed white feces and dark yellow urine. On day 42, the urine and feces of the chronic BA mice also showed obvious features of jaundice (Figure 1D,E). The livers of the BA mice were removed and compared to those of the controls. The livers were smaller (Figure 2B), necrotic lesions were visible to the naked eye (Figure 2A, black triangle), and a segment of bile duct atresia was also observed extrahepatically (Figure 2A, white asterisks).
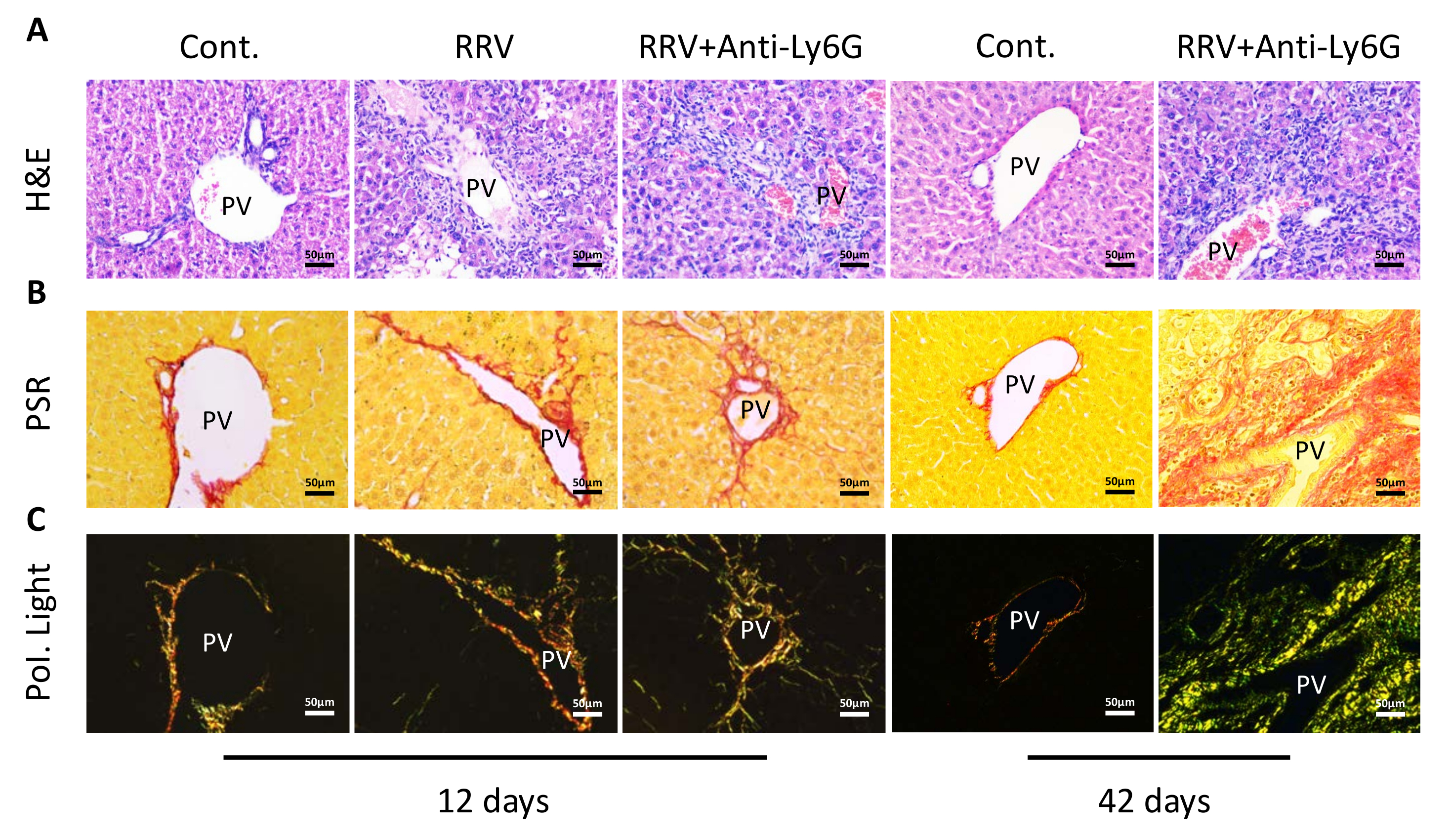
Liver tissue section analysis showed that low-dose anti-Ly6G therapy decreased portal vein inflammation. However, inflammatory cell accumulation was still observed in the liver tissue slices at day 42 (Figure 3A). Sirius Red staining showed a small increase in the deposition of collagen in the portal region on day 12 after the RRV injection. Moreover, no significant changes were observed in collagen expression after treatment with the anti-Ly6g antibody, but the collagen expression in BA tissue samples at day 42 was significantly increased (Figure 3B). When examined under a polarized light microscope, it was observed that collagen fibers had accumulated in the adjacent liver tissue. A substantial deposition of collagen, predominantly green, was seen in the BA tissue specimens at day 42 (Figure 3C), and collagen deposition increased further in mice that survived for 42 days without weight gain. With the reduction of portal inflammation, CK19+ bile duct cells were observed on day 12. On day 42, however, mature bile ducts were hardly detectable, even though an increase in CK19+ bile duct cells was seen (Figure 4A, red arrows indicate bile duct epithelial cells). In the case of extrahepatic bile ducts in mice with chronic BA, serial dissection of the portal region of mice revealed that the extrahepatic bile ducts were blocked and had inflammatory infiltrates of macrophages (Figure 4B, black arrows indicate macrophages).
Compared with RRV alone, treatment with a low dose of anti-Ly6G antibody decreased liver injury in terms of liver enzyme levels on day 12 after RRV inoculation. The alanine aminotransferase and alkaline phosphatase levels in the liver were higher in the chronic BA group than in the normal control group. The most pronounced changes were found in the bilirubin levels, with RRV + anti-Ly6G mice showing reduced TBIL, DBIL, and IBIL levels in acute BA by contrast with RRV alone. In mice with chronic BA, the levels of TBIL, DBIL, and IBIL were increased (Figure 5), indicating that liver function was significantly reduced in the chronic fibrosis stage of BA.

Figure 1: The effects of anti-Ly6G antibody treatment in a mouse model of biliary atresia infected with RRV. (A) Schematic illustration of low doses of antibodiesto induce acute and chronic phase BA in mice with arrows indicating the time of injection of the antibody and RRV. (B) Survival curves of the mice in the normal control (Cont.) group, rhesus rotavirus (RRV) group, and RRV + anti-Ly6G antibody group. (C) Body weight curves of the mice in each group. (D) Representative images of the mice and their feces and urine in each group on day 12. (E) Representative images of the mice and their feces and urine in each group on day 42. The blue arrows indicate the yellow ears and tail. This figure has been reprinted with permission from Zhang et al.15. Please click here to view a larger version of this figure.
{kind=link}

Figure 2: The effects of anti-Ly6G antibody treatment on the liver in a mouse model of biliary atresia infected with RRV. (A) Comparison of liver anatomy and extrahepatic bile duct fluorescence angiography between chronic BA mice (right) and normal mice on day 42. (B) Comparison of the liver size between chronic BA mice and normal mice. The black triangle indicates liver necrosis in mice with chronic BA. The white asterisk indicates extrahepatic biliary atresia. This figure has been reprinted with permission from Zhang et al.15. Please click here to view a larger version of this figure.
{kind=link}

Figure 3: Liver tissue slices of the mice in the normal control (Cont.) group, rhesus rotavirus (RRV) group, and RRV + anti-Ly6G antibody group on day 12 and day 42. (A) Images of liver tissue sections stained with hematoxylin and eosin (H&E). (B) Sirius Red staining showed collagen deposition (PSR). (C) Further observation of the slices using polarized light microscopy (Pol. Light). Abbreviation: PV = portal vein. Scale bar = 50 µm. This figure has been modified from Zhang et al.15. Please click here to view a larger version of this figure.
{kind=link}

Figure 4: Immunohistochemical staining of liver tissue slices from the normal control (Cont.) group, rhesus rotavirus (RRV) group, and RRV + anti-Ly6G antibody group on day 12 and day 42. (A) The expression of bile duct epithelial cells marker (CK19) in different treatment groups. (B) Inflammatory cell infiltration of macrophages was demonstrated in different treatment groups. Abbreviation: PV = portal vein. The red arrow indicates bile duct epithelial cells. The black arrow indicates macrophages. Scale bar = 50 µm. This figure has been modified from Zhang et al.15. Please click here to view a larger version of this figure.
{kind=link}

Figure 5: Comparison of liver function in mice in different treatment groups. Mouse blood was collected after the experiment and tested in the laboratory department of the hospital. Each group contained three to five mice. *P < 0.05, **P < 0.01, ***P < 0.001. Abbreviations: ALT = alanine aminotransferase; AST = aspartate aminotransferase; ALP = alkaline phosphatase; TBIL = total bilirubin; DBIL = direct bilirubin; IBIL = indirect bilirubin. This figure has been modified from Zhang et al.15. Please click here to view a larger version of this figure.
{kind=link}
Discussion
In our study, we used anti-Ly6G antibody to eliminate or decrease Gr-1+ cells in a mouse model of biliary atresia infected with RRV to improve acute BA syndrome, prolong the survival rate, and enable BA mice to enter the chronic phase. Reducing the antibody dose can lead to chronic BA with hepatic fibrosis, which indicates that the number of Gr-1+ cells changes the result of BA in the acute and chronic phases. In our previous study, Gr-1+ cells were reduced after the administration of anti-Ly6G antibody, and the overall survival status of the BA mice was improved15. At the same time, it has been reported that Gr-1+ macrophages19, Gr-1+ neutrophils20, Gr-1+ myeloid cells21, and Gr-1+ granulocytes can enhance fibrosis in some animal models22. However, in this study, Gr-1+ cells in mice were partially depleted with low doses of anti-Ly6G antibody, and collagen deposits remained. As a result, more time may be needed to address the disease more thoroughly.
The application of the anti-Ly6G antibody decreased inflammation, partially preserved the bile ducts, and prevented acute BA-induced death in mice. However, only 20% to 30% of the mice entered the chronic phase of BA; this may be related to the time point and the number of injections of anti-Ly6G antibody. We need to further explore this key point to improve the success rate. In addition, we consider that not only Gr-1+ cells but also other immune cells may play important roles, such as NK cells, T cells, B cells, and macrophages.
As for the protocol, some details need to be noted. (i) In order to avoid the leakage of injected drugs, we use a 1 mL insulin syringe with a 0.33 mm diameter needle, because the needle diameter is small, and the needle hole into the syringe is small, which are conducive to reducing the possibility of drug leakage. (ii) Before the injection, the mouse should be fixed to prevent it from moving, avoid drug leakage, and, thus, further ensure the experimental effect. (iii) During drug injection, we injected the drug into the surface or lower margin of the mouse liver as far as possible so that the drug entered the abdomen and fully contacted the liver to take effect. (iv) The injection is usually made from the upper right thigh of the mouse, because the newborn mouse's stomach and spleen are in the left abdomen, and the stomach is full of milk. If the needle is inserted from the left side, it can easily puncture the stomach, causing milk to flow into the abdomen, or puncture the spleen, causing bleeding.
CD177 is a homolog of Ly6G, and the expression of CD177 in BA patients has been investigated. CD177 can be used as a marker for the early diagnosis of BA in children, indicating that CD177+ cells play a significant role in the course of BA23. By analyzing the RNA sequencing data, our team found that CD177 cells were the dominant population of Gr-1+ cells; meanwhile, Cd177−/− BALB/c mice vaccinated with RRV showed a delayed onset of BA and reduced morbidity and mortality18. Thus, our chronic BA mouse model has obvious advantages for studying the pathogenesis and disease progression of BA; it also provides a suitable animal model for studying potential treatments for BA.
Disclosures
The authors have no conflicts of interest to disclose.
Acknowledgements
This work was supported by grants from the National Natural Science Foundation of China (81974056) to R.Z. and from the National Nature Youth Foundation of China (82101808) and Science and Technology Planning Project of Guangzhou (No. 202102020196) to Z.L.
Materials
| Name | Company | Catalog Number | Comments |
| Balb/c mouse | Guangdong Skarjingda Biotechnology Co., LTD | 20221000112 | |
| rat anti-mouse Ly6G | Bio X Cell | clone 1A8 | West Lebanon, NH |
| rat anti-mouse cytokeratin 19 | Developmental Studies Hybridoma Bank | clone TROMA III | Iowa City, IA |
| rat anti-mouse F4/80 | R&D Systems | MAB5580 | Minneapolis, MN |
| RRV strain | ATCC | MMU 18006 | Manassas, VA |
| Fluorescent stereomicroscope | Olympus | SZX7 | |
| Leica light microscopy | Leica Microsystems | Leica DMI8+DFC7000T | Wetzlar, Germany |
| Hitachi Pre-Analytical Process Automation System | Hitachi | 7600 Clinical Analyzer | Tokyo, Japan |
| Isoflurane anesthetic | RWD | R510-22-10 | |
| Rhodamine 123 | Sigma-Aldrich | 83702 | |
| sirius red dye | Leagene | DC0041 | |
| paraffin microtome | Leica Microsystems | RM2235 | |
| neutral gum | Solarbio | G8590 |
References
- Lakshminarayanan, B., Davenport, M. Biliary atresia: A comprehensive review. Journal of Autoimmunity. 73, 1-9 (2016).
- Bassett, M. D., Murray, K. F. Biliary atresia: Recent progress. Journal of Clinical Gastroenterology. 42 (6), 720-729 (2008).
- Bezerra, J. A., et al. Biliary atresia: Clinical and research challenges for the twenty-first century. Hepatology. 68 (3), 1163-1173 (2018).
- Hartley, J. L., Davenport, M., Kelly, D. A. Biliary atresia. Lancet. 374 (9702), 1704-1713 (2009).
- Lee, W. S., Looi, L. M. Usefulness of a scoring system in the interpretation of histology in neonatal cholestasis. World Journal of Gastroenterology. 15 (42), 5326-5333 (2009).
- Russo, P., et al. Key histopathologic features of liver biopsies that distinguish biliary atresia from other causes of infantile cholestasis and their correlation with outcome: A multicenter study. The American Journal of Surgical Pathology. 40 (12), 1601-1615 (2016).
- Fickert, P., et al. Characterization of animal models for primary sclerosing cholangitis (PSC). Journal of Hepatology. 60 (6), 1290-1303 (2014).
- Sonoda, S., et al. Biliary atresia-specific deciduous pulp stem cells feature biliary deficiency. Stem Cell Research and Therapy. 12 (1), 582 (2021).
- Xiao, Y., et al. Long noncoding RNA H19 contributes to cholangiocyte proliferation and cholestatic liver fibrosis in biliary atresia. Hepatology. 70 (5), 1658-1673 (2019).
- Desmet, V. J., Krstulović, B., Van Damme, B. Histochemical study of rat liver in alpha-naphthyl isothiocyanate (ANIT) induced cholestasis. The American Journal of Pathology. 52 (2), 401-421 (1968).
- Luo, Z., Shivakumar, P., Mourya, R., Gutta, S., Bezerra, J. A. Gene expression signatures associated with survival times of pediatric patients with biliary atresia identify potential therapeutic agents. Gastroenterology. 157 (4), 1138-1152 (2019).
- Mariotti, V., Strazzabosco, M., Fabris, L., Calvisi, D. F. Animal models of biliary injury and altered bile acid metabolism. Biochimica et Biophysica Acta. Molecular Basis of Disease. 1864, 1254-1261 (2018).
- Egan, C. E., Sukhumavasi, W., Bierly, A. L., Denkers, E. Y. Understanding the multiple functions of Gr-1(+) cell subpopulations during microbial infection. Immunologic Research. 40 (1), 35-48 (2008).
- McDermott, A. J., et al. The role of Gr-1(+) cells and tumour necrosis factor-α signalling during Clostridium difficile colitis in mice. Immunology. 144 (4), 704-716 (2015).
- Zhang, R., et al. The role of neonatal Gr-1(+) myeloid cells in a murine model of rhesus-rotavirus-induced biliary atresia. The American Journal of Pathology. 188 (11), 2617-2628 (2018).
- Fu, M., et al. A silver nanoparticle method for ameliorating biliary atresia syndrome in mice. Journal of Visualized Experiments. (140), e58158 (2018).
- Sy, J., Ang, L. C. Microtomy: Cutting formalin-fixed, paraffin-embedded sections. Methods in Molecular Biology. 1897, 269-278 (2019).
- Zhang, R., et al. CD177(+) cells produce neutrophil extracellular traps that promote biliary atresia. Journal of Hepatology. 77 (5), 1299-1310 (2022).
- Engel, D. R., et al. CX3CR1 reduces kidney fibrosis by inhibiting local proliferation of profibrotic macrophages. Journal of Immunology. 194 (4), 1628-1638 (2015).
- Liang, M., et al. A modified murine model of systemic sclerosis: Bleomycin given by pump infusion induced skin and pulmonary inflammation and fibrosis. Laboratory Investigation. 95 (3), 342-350 (2015).
- Chen, Y., et al. Differential effects of sorafenib on liver versus tumor fibrosis mediated by stromal-derived factor 1 alpha/C-X-C receptor type 4 axis and myeloid differentiation antigen-positive myeloid cell infiltration in mice. Hepatology. 59 (4), 1435-1447 (2014).
- Tomasson, M. H., et al. Fatal myeloproliferation, induced in mice by TEL/PDGFbetaR expression, depends on PDGFbetaR tyrosines 579/581. Journal of Clinical Investigation. 105 (4), 423-432 (2000).
- Zhang, R., Huang, J., Shan, J., Chen, Y., Xia, H. Peripheral blood CD177(+) cells as an early diagnostic marker for biliary atresia: A prospective multicentre study in pediatric patients with cholestasis. Journal of Hepatology. 77 (6), 1714-1716 (2022).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved