
Method Article
Kardiyomiyosit DNA Sentezi ve ploidinin Eşzamanlı Değerlendirme: Bir Yöntem kardiyomiyosit Yenilenme ve ciro miktarının belirlenmesi Yrd
Bu Makalede
Özet
Quantification of cardiomyocyte turnover is challenging. The protocol described here makes an important contribution to this challenge by enabling accurate and sensitive quantification of neo-cardiomyocyte nuclei generation and nuclei ploidy.
Özet
kalp, normal yaşlanma sırasında yaralanma aşağıdaki kardiyomiyositlerde kardiyomyosit ciro bu düşük seviyelerde meydana yenilemek için sınırlı bir potansiyele sahip olduğu kabul edilmekle birlikte, bu olayların ölçümü zor kalır. Bu durum sürecin nadir ve çok sayıda hücresel kaynaklar miyokard korunmasına katkıda gerçeğine bölümünde yer almaktadır. Ayrıca, kardiyomiyositlerinin içinde DNA çoğaltılması genellikle polyploid kardiyomiyosit yol açar ve nadiren hücresel bölümü tarafından yeni kardiyomiyositlere yol açar. doğru bu süreçler arasındaki kardiyomiyosit ciro ayrımcılığı ölçmek için esastır. Burada açıklanan protokol çekirdek izolasyon ve sonraki PCM1 immün kullanılarak belirlenen DNA replikasyonu kardiyomyosit çekirdekleri sonucunda ortaya çıkan tüm çekirdeklerin etiketlemek için uzun vadeli nükleozid etiketleme kullanır. Birlikte bu c nükleozid etiketleme doğru ve hassas bir şekilde tanımlanmasına yardımcı olmaktadırçekirdekler nüfus ardiomyocyte. Ayrıca, 4 ', 6-diamidino-2-fenilindol etiketleme ve çekirdekler ploidinin analizi, poliploidleşmeyi sırasında nükleosit dahil olan çekirdeklerin neo-kardiyomiyosit çekirdeklerin ayrım sağlar. Bu yöntem, kardiyomiyosit binucleation için kontrol olamaz da poliploidleşmeyi için gerekli hesaplamaların, neo-kardiyomiyosit çekirdeklerin hızlı ve sağlam bir ölçümü sağlar. Bu yöntem kardiyomyosit rejenerasyonu artırmak için potansiyel terapötikler değerlendirilmesi veya kardiyomiyosit ciro ve ploidleşmede kardiyak hastalığın etkilerini araştıran dahil mansap uygulamaları bir dizi vardır. Bu teknik, aynı zamanda tüm kardiyak hücre tiplerinde nükleosit esas sayılmasına olanak tanıyan ek bir alt immünohistolojik tekniklerle uyumludur.
Giriş
Son yıllarda kalp terminal farklılaşmış, post-mitotik organı 1,2 olduğunu varsayım zorlu delil birikimi olmuştur. Ancak, kardiyomiyosit ciro ve yenilenme ölçümü zor kalır.
Doğru, standart immünohistokimyasal teknikler kullanılarak nadir kardiyomiyosit nesil belirlenmesinde zorluklar iyi 3 bildirilmiştir. Buna ek olarak, kardiyomiyosit nesil hücresel kaynak kardiyomiyosit çoğalması ve kök hücre farklılaşması 4-6 katkıları için kanıt belirsizdir. Bu nedenle, kardiyomiyosit progenitör fenotip bilgisi gerektiren soyu izleme modellerin kullanımı imkansız ve kardiyomiyositlerde dahil olmak üzere tek nüfus, çoğalma ölçümü ise, uygun değildir. Ayrıca, bir kardiyomiyosit karyokinesis olmayan endoreplication potansiyeline sahip (bir poliploid araba sonuçlanandiomyocyte) ya da sitokinez (a binükleer kardiyomiyosit sonuçlanır) 7,8 yokluğunda karyokinesis. Kardiyomiyosit ciro doğru ölçümü bu olaylar ve gerçek neo-kardiyomiyosit nesil arasında ayrım yeteneğine bağlıdır. Kardiyomiyositlerde DNA replikasyonu ve sikline bağımlı kinazlann ekspresyonu sadece gerçek hücre bölünmesini 9,10 göstermektedir, çünkü bu özel sorun çıkarmaktadır.
Uzun vadede yeni yöntemlere göre Bergmann ve ark., 7,11 tarif edildiği gibi, neo-kardiyomiyosit nesil miktar yardımcı olmak için, kurulu bir çekirdek izolasyon tekniği ve kardiyomiyosit çekirdekleri tanımlanması için pericentriolar madde 1 (PCM1) immünolojik etiketleme birleştirdik DNA etiketleme ve ploid analizi. PCM-1 farklılaşmış olmayan bisiklet miyositler nükleer yüzeyinde birikir bir sentrozom proteindir. Daha önceki çalışmalar, karşı antikor olduğunu göstermiştirPCM-1, özellikle kardiyomiyosit çekirdekleri 7,11 etiket ve PCM1 bağımsız grupların sayısına göre kullanılır olmuştur olarak kardiyomiyositlerde 1,12,13 tanımlamak için. Buna ek olarak, PCM1 sentezleme TNT-cre transjenik fare modelinde 14 (Ek Şekil 1) genetik olarak etiketlenmiş kardiyomiyosit çekirdekleri eşleştiren olduğunu göstermiştir.
Burada açıklanan protokol bağımsız bağlı analizi (Şekil 1C, D) için poliploidleşmeyi hücresel kökenli (Şekil 1 A ve B), aynı zamanda nükleozit etiketleme hariç ise bölgesinin fare kalp neo kardiyomiyosit çekirdeklerinin üretiminin doğru ve hassas bir tanımlanmasını sağlar. Bu yöntem, kardiyomiyosit binucleation için kontrol edemez birlikte, kardiyomiyosit devir doğru ölçümü için gerekli olan yeni-kardiyomiyosit çekirdeklerin hızlı ve sağlam bir ölçümü sağlar. Ayrıca,o Kardiyomiyosit nesil dinamikleri potansiyel değişiklikleri değerlendirmek için hızlı bir tarama aracı sağlar.
DNA etiketleme genellikle timidin analoğu gibi, 5-bromo-2'-deoksiüridin (BrdU) içerirken, burada açıklanan protokolü daha hızlı için daha az işlem adımı gerektirir gibi bir 5-etinil-2'-deoksiüridin (edu) bazlı analiz kullanır iş-zaman ve diğer immün protokolleri ve böylece yöntemin potansiyel alt uygulamaları artan ile uyumlu hale immüno saptanması için DNA denatüre gerektirmez.

Şekil 1: Edu ile sürekli darbe bakılmaksızın atalarıdır neo-kardiyomiyositlerde etiketler. (A) edu hücre bölünmesi esnasında kardiyomiyositlerin DNA'ya dahil edilir. kardiyomiyosit popülasyonda çoğalma Resul olacakkardiyomiyositlerinin bir artış ya da değiştirilmesi t ve dolayısıyla verimli DNA sentezi (doku bakım ve onarımı katkıda) 'dir. (B) edu hücre bölünmesi sırasında kalp progenitör hücrelerin DNA içine dahil edilir. Bu kardiyomiyosit soy farklılaşma sırasında hücrede muhafaza edilecektir. Bu kök hücre farklılaşması da kardiyomiyositlerinin sayısında bir artışa neden ve bu nedenle doku bakım ve onarım katkıda olacaktır. (C) Kardiyomiyositler kardiyomiyosit hipertrofisi ve miyokard biçimlenme ile ilişkili artmış kardiyomiyosit ploidi, sonuçlanan "üretken olmayan" DNA replikasyonu geçmesi potansiyeline sahip, ancak kaybedilen kardiyomiyositlerde yerine geçmez. Bu iki homolog kromozomlar (> 2 N), dört veya daha fazla grup ihtiva eden bir tek çekirdek bir kardiyomiyosit sonuçlanan şekilde poliploidleşmeyi bir işlem binucleation farklıdır. Sürekli bir çekirdek p aşağıdakı (D)Ulse, bu protokol kardiyomiyosit ploidinin ve edu dahil hem miktarının izin PCM1 ifade tarafından kardiyomiyosit çekirdeklerinin çekirdekleri izolasyonu ve tanımlanmasını açıklar. PCM1 ifade ve akım sitometri kullanılarak tespit edu kuruluş. Bu rakamın büyük halini görmek için lütfen buraya tıklayınız.
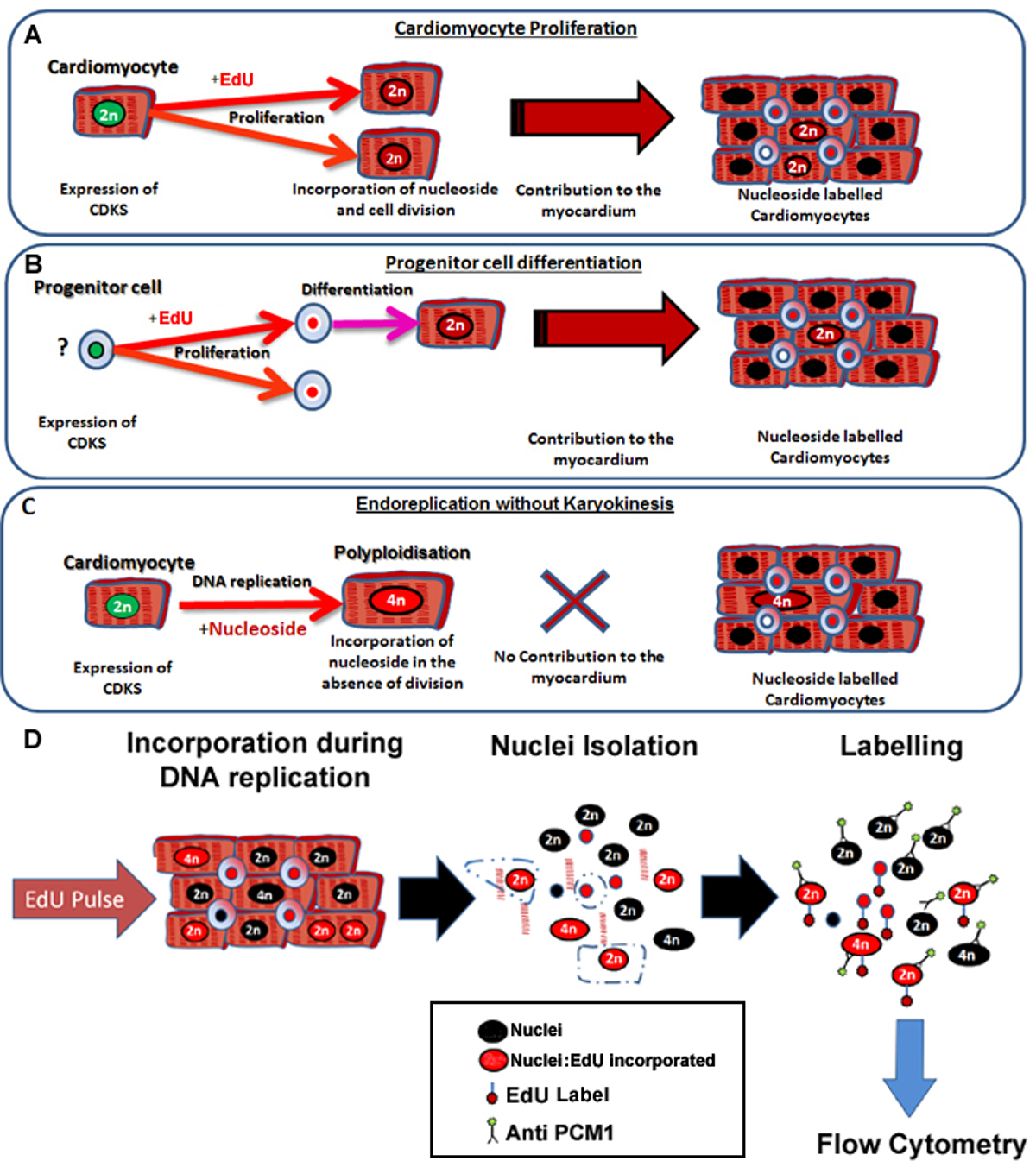{kind=link}
Protokol
Hayvan çalışmaları yetkili ve Newcastle Üniversitesi Etik inceleme kurulu tarafından onaylanmıştır. Tüm hayvan prosedürleri bilimsel amaçlarla kullanılan hayvanların korunmasına ilişkin Yönerge'de 2010 Avrupa Parlamentosu / 63 / EU gelen yönergelere uygun gerçekleştirilir bulundu.
1. Edu İdaresi
- steril tuzlu su çözeltisi içinde çözülür edu (w /% 0.9 h) 12.5 mg / ml'lik bir nihai konsantrasyonda.
- tamamen çözülmesi için 40 ° C ve vorteks çözelti ısıtılır. 6 günlük enjeksiyonlar için izin çalışmada tüm fareleri enjekte 4 ° C'de yeterli edu / Saline saklayın.
Not: Genellikle 6 farelerin her bir deney grubu için gereklidir. Ancak, güç hesapları bağımsız çalışmalar için yapılmalıdır.
- tamamen çözülmesi için 40 ° C ve vorteks çözelti ısıtılır. 6 günlük enjeksiyonlar için izin çalışmada tüm fareleri enjekte 4 ° C'de yeterli edu / Saline saklayın.
- Bireysel fareler tartılır. Her fare edu / tuzlu su çözeltisi içinde 100 ug / g (g başına stok edu solüsyonu 8 ul) yönetmek için gerekli hacmi hesaplayın.
- Uygun volum çizmek25 G iğne ile insülin şırınga içine edu / tuzlu e.
- onaylı Ev Ofis tekniklerini kullanarak fareleri işlerken intraperitoneal (ip) enjeksiyonları gerçekleştirin.
- kafes kapağındaki bir fare yerleştirin. Yavaşça kuyruk tabanı geri çekin ve sıkıca yapışmak fare kavramak.
- kafa tabanına yakın işaret parmağı ve başparmak ile fare tutan ve yere doğru geriye hayvanın kafasını devrilme enjeksiyon için karın Açığa.
- % 70 alkol solüsyonu ile karın silin. Hayvanın orta hat ve sağ alt kadranda bulun. Sağ alt kadran içine hayvana edu / Tuzlu çözüm enjekte.
- Edu / tuzlu su çözeltisi ile fareler enjekte edilir ve günün saatini kaydeder.
- Tekrarlayın 6 gün boyunca, günün aynı anda 1.4 1.2 adımları tekrarlayın.
Not: Bir Edu negatif kontrol akım sitometri analizi için yolluk ayarlamak için gerekli olduğundan, ek bir yaş, cinsiyet enjekte ve deneysel maçEdu olmayan tuz denk hacimde Ed fare. Bu, gerekli kontrolü (hiçbir EDU kontrolü) sağlayacaktır. Her çalışma reaktiflerin gruplar halinde varyasyonlara izin için bu kontrol dahil edilmelidir ve sitometresi kurdu.
2. Hasat Kalpler
- 6. Edu enjeksiyonundan 24 saat sonra, servikal dislokasyon (ya da 1 Home Office onaylı alternatif Çizelgesi yöntemi) kullanılarak fareler kurban.
- Bir yatar pozisyonda kurban hayvan koyun ve cerrahi makas ile diyaframa orta karın cilt kesi yapmak.
- diyaframı kesti ve kalp maruz bilateral kesilmiş vücut boşluğunda uzak sternum tutarak.
- biraz kalbi kaldırın ve göğüs boşluğu dışında kalbini incelemek için çıkış yolunda büyük kan damarları kesilmiş.
- Hemen PBS 4 ° C'ye soğutulmuş 10 ml içeren bağımsız bir 15 ml santrifüj tüpüne her kalp yerleştirin.
- sep kalpleri aktarınarate 10 cm Petri kapları, bir neşter sagital yönde ikiye her kalbi kesilmiş ve yavaşça forseps bir çift ile sıkarak kalbi yıkayın kullanarak. Sonra mümkün olduğunca kan kaldırmak için PBS değiştirin. İki kez bu adımı yineleyin.
- Aynı birey 1.5 ml mikrosantrifüj tüp içine her kalbin iki parçayı yerleştirin.
- 3 veya alternatif mağaza örnekleri adıma -80 ° C adımlarla 2.9 ve 2.10 ayrıntılı olarak devam etmektedir.
- 25 G iğne kullanılarak, kalp işlendiğinde Mikrosantrifüj tüpü kapak küçük bir delik dolayı gazların genişlemesine kapağın açılmasını önlemek için kullanılır.
- dondurma için sıvı azotun, bir kap içinde her bir mikrosantrifüj tüpü ve yer etiketi.
3. Çekirdekler Ayrılma ve kardiyomiyosit Çekirdekler Etiketleme
. 11 Bu çekirdekler ayrılma kardiyomyosit çekirdekleri etiketleme Bergmann ve ark o uyarlanmıştır enjekte tüm örnekler üzerinde bu yordamı gerçekleştirin: NotEdu ve enjekte tuz (hiçbir EDU kontrolü) negatif kontrol ile. Bu adımları gerçekleştirmek için gerekli çözümlerin tarifi için Tablo 1'e bakınız.
- Her bir kalp analiz edilmesi için, 30 dakika inkübe edilir, bir ultra santrifüj tüpüne% 1 BSA / PBS çözeltisi 36 mL yer çözeltisi atılır ve daha sonra kurumaya bırakın.
Not: Bu tüp adım 3.6 kullanılır. - kalpleri kıyma işlemi sırasında defrost sağlayan bir neşter kullanılarak buz ve kıyma üzerinde 10 mm çanak üzerine bireysel dondurulmuş kalpleri yerleştirin.
- 50 ml'lik bir santrifüj tüpünde hücre lisis tamponu, 15 ml inceltilmiş kalpleri yerleştirin ve oda sıcaklığında 25.000 rpm'de 15 saniye için bir sonda homojenleştirici ile homojenize edilmiştir.
- Santrifüj tüpüne Celi Lysis Buffer bir başka 15 ml ilave edilir ve geniş bir açıklık havan tokmağı ile 40 ml'lik bir Dounce transfer. havaneli ile 10 kez gerçekleştirmek ve 50 ml'lik bir santrifüj tüpü içine, bir 100 um ile daha sonra 40 mikron hücre süzgecinden süzülür.
- 10 dk için santrifüjT 700, 4 ° C'de xg ve supernatant çıkarın.
- Sükroz Gradyan çözeltiden 25 ml, nükleer pelet yeniden süspanse edin ve 3.1 hazırlanan bir ultra santrifüj tüpüne taze sukroz gradyanlı çözeltisi 10 ml üzerine çözelti içeren hücre çekirdekleri katman.
- bir salıncak üzerinden boru rotor kullanılarak 4 ° C sıcaklıkta 1 saat boyunca 13.000 x g'de santrifüjleyin.
- Santrifüj sonrasında, süpernatant atılır ve 1.5 ml mikrosantrifüj tüpü ve etiket PCM1 çekirdekler saklama tamponu 1.300 ul çekirdekleri pelletini.
- Yeni bir mikrosantrifüj tüpüne süspansiyon 300 ul çıkarın ve taze çekirdekler depolama tamponu 700 ul ekle. ISO kontrolü gibi bu tüp etiketleyin.
- PCM1 etiketli mikrosantrifüj tüpüne mi 8 ug / mL'lik nihai bir konsantrasyonda anti PCM1 ekleyin ve ISO kontrolü etiketli mikrosantrifüj tüpüne 8 ug / ml bir konsantrasyonda Tavşan IgG izotip kontrol antikoru ilave edin.
- 4 ° C'de bir gece boyunca inkübe edilir. aşağıdaki inkubasyonation, 10 dakika boyunca 700 x g'de santrifüj 700 x g'de 10 dakika boyunca tekrar taze çekirdek saklama tamponu, tekrar süspansiyon ve santrifüj 1 ml yerine, süpernatant atılır. Süpernatantı atın ve taze çekirdekler depolama tamponu 1 ml ile değiştirin.
- 2 ug / ml'lik bir nihai konsantrasyon elde etmek için F (ab ') Keçi anti-tavşan IgG (H + L) antikoru (FITC veya eşdeğer yeşil floresan boya) 2 Fragment 1 ul ekle.
Not: bir F (ab ') kullanımı 2 fragmanı antikorun makrofajlar ve B hücreleri gibi immün hücrelerin spesifik olmayan etiketleme potansiyelini azaltır. - alüminyum folyo sarma borular ışıktan korumak ve 4 ° C'de 1 saat boyunca inkübasyona.
Edu Kuruluş 4. Algılama
- Seyreltin 10x deiyonize su ile edu reaksiyon tamponu 01:10 konsantre.
- Santrifüj PCM-1 ve ISO kontrolü etiketli çekirdek süspansiyonları, 10 dakika boyunca 700 x g hızında, süpernatan atılır ve 1 ml çekirdekleri pelet% 1 BSA / PBS solüsyonu. iki çekirdek pelet yıkayın.
- Son yıkamadan sonra, süpernatantı atmak ve edu fiksatif 100 ul ile değiştirin.
- ışıktan korumak ve 15 dakika oda sıcaklığında inkübe edin, alüminyum folyo ile tüpler sarın.
- 700 xg% 1 BSA / PBS çözeltisi, santrifüj kez 1 ml örneği yıkama ve 15 dakika boyunca oda sıcaklığında 1 x saponin bazlı permeabilizasyon çözeltisi ile inkübe edilir.
- 1x edu etiketleme kokteyl (Tablo 2) hazırlayın. 2 reaksiyonların en az hiçbir EDU kontrolü de gerekecektir.
- şirketinden önce 1x saponin bazlı permeabilizasyon çözeltisi 100 ul çekirdek içeren her bir örnek için 1x edu markalama kokteyl 500 ul ilave edin ve 30 dakika boyunca ışıktan korunarak oda sıcaklığında inkübe edin.
- % 1 BSA / PBS 1 ml 700 xg ve tekrar süspansiyon Santrifüj ve bu adımı iki kez daha tekrarlayın.
- 700 xg'de santrifüj, supernatant atmak ve 4 ile değiştirinDNA boyama çözeltisi 00 ul (Malzeme Tablo).
| Reaktif Adı |
| 1. Hücre Liziz Tamponu |
| 0.32 M sukroz |
| 10 mM Tris-HCI (pH = 8) |
| 5 mM CaCl2 |
| 5 mM magnezyum asetat |
| 2.0 mM EDTA |
| 0.5 mM EGTA |
| 1 mM DTT |
| 2.Sucrose Gradyan Çözelti |
| 2.1 M sukroz |
| 10 mM Tris-HCI (pH = 8) |
| 5 mM magnezyum asetat |
| 1 mM DTT |
| 3. Çekirdekler depolama tamponu (NSB) |
| 0.44 M sukroz |
| 10 mM Tris-HCI (pH = 7.2) |
| 70 mM KCI |
| 10 mM MgCl2 |
| 1.5 mM spermin |
Tablo 1: Çekirdekler izolasyon tampon maddeler protokol bölüm 3 (Çekirdekler ayrışma kardiyomyosit çekirdekleri etiketleme) kullanılan tüm Tamponlar için Reçete..
5. Akım Sitometrisi Analizi
Not: Daha önce 7,11,15 tarif edildiği gibi izole çekirdekler üzerinde sitometri Akışı gerçekleştirin.
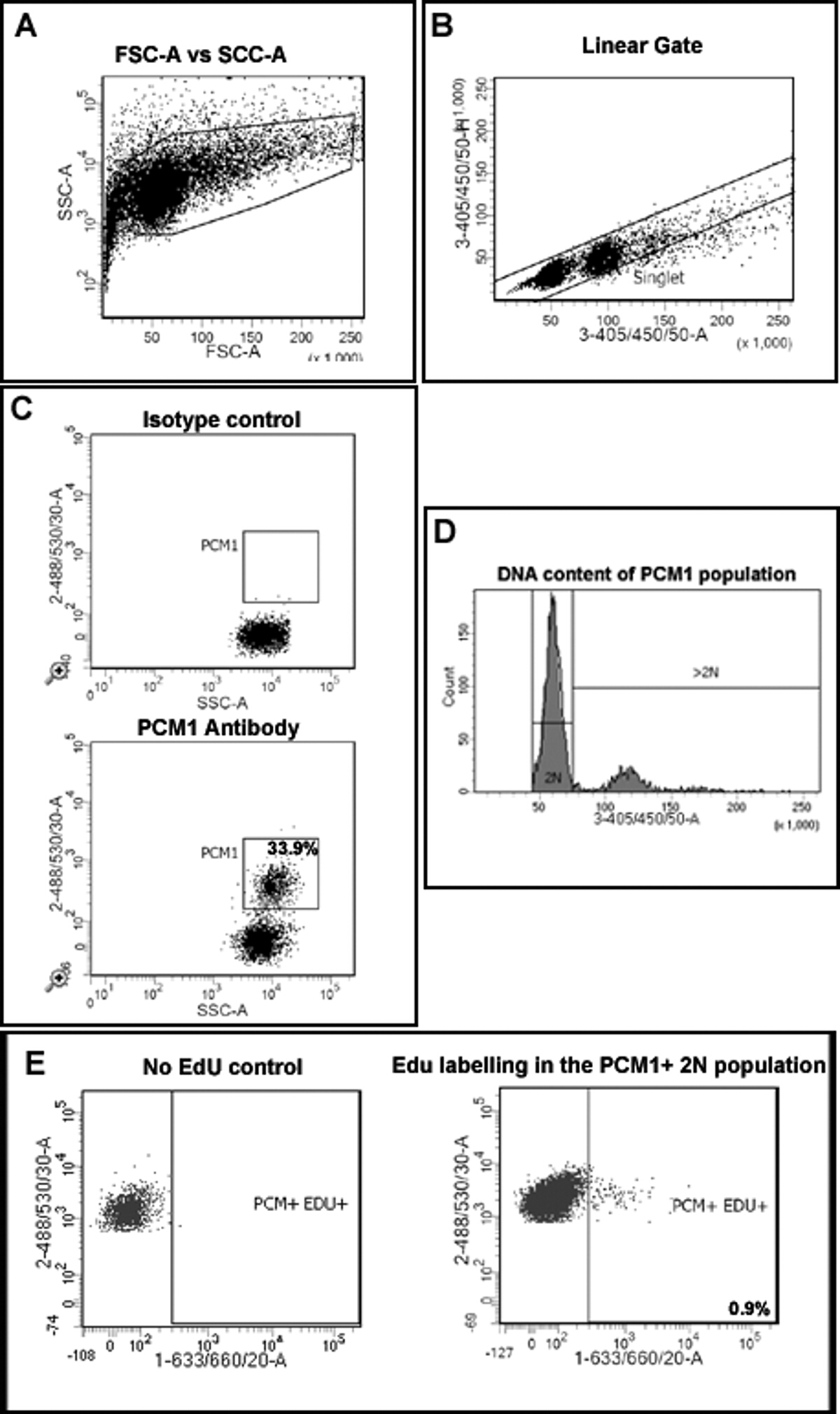
- İlk olarak, enkaz (Şekil 2A) gelen çekirdeklerin ayrımcılık izin İleri Yayılım (FSC) ve Yana Yayılım (SSC) açıklayan bir arsa oluşturun.
- Yüksekliği karşı 4 çizerek agrega (Şekil 2B) dan tek çekirdekleri ayırmak için bir komplo ', 6-diamidino-2-phenylindole) oluşturun (DAPI) alan (3-405 / 450/50-A) (33-405 / 450 / 50-H). tha sağlamakt 3-405 / 450/50 sinyal DNA içeriğinin doğru belirlenmesini sağlamak için doğrusal bir ölçek üzerinde toplanır.
- SSC vs 488/535/30 A açıklayan bir arsa (PCM-1) oluşturma ve tekli popülasyonda PCM-1 ifadesini değerlendirmek için 5.2 oluşturulan tekli kapısından sadece olayları görüntülemek.
- PCM-1 pozitif kapı (Şekil 2C) oluşturmak üzere İzotip kontrol örneği çalıştırın. PCM1 küçük bir miktar nüfus ve yolluk konumunu ifade PCM1 doğrulamak için örnek etiketli çalıştırın.
- 405/450/50-A (DAPI algılamak için) vs SSC-A açıklayan bir arsa oluşturun. Bu arsa içinde, ekran sadece 5.3 oluşturulan kapıları kullanarak, PCM1 ifade çekirdekleri tekli. Sadece 2N çekirdekleri (Şekil 2B) içeren ek bir hiyerarşik kapısı yaratmak için bu komplo kullanın.
- Kardiyomiyosit nüfusu ifade PCM1 EDU etiketleme tanımlanmasını sağlamak için 1-633 / 660/20-A vs 488/535 / 30 A tanımlayan bir arsa oluşturun. tekli, P sadece çekirdekleri görüntülemek Yukarıdaki ayırıp kullanarak CM1 + ve 2N.
- Hiçbir EDU kontrolü çalıştırın kalp örnek edu pozitif kapısı (Şekil 2E) ayarlamak için. Edu + hücreleri için uygun kapı oluşturun.
- Tutanak ve ölçmek tekli, Edu dahil olması PCM-1 ifade 2N çekirdekler.
- toplam tekli, PCM1 + çekirdek bir yüzdesi olarak bu nüfusu hesaplayın. Bu, toplam kardiyomiyosit çekirdekleri bir yüzdesi olarak, 7 günlük edu darbe süresince neo-kardiyomiyosit çekirdek oluşum oranını sağlayacaktır.
Not: DNA bağlanma DAPI stoikiometrik olduğu floresan yoğunluğu DNA miktarı ile orantılıdır. Daha önce tarif edildiği gibi, 7 405/450/50 bir sinyalin yoğunluğuna dayalı olarak ploidi belirler. - Toplam kardiyomiyosit nüfus içinde ploidi değerlendirmek için 5.5 kurulan arsa ve DNA konsantrasyonu 7 dayalı bir yolluk strateji kullanın.
/files/ftp_upload/53979/53979fig2.jpg "/>
Şekil 2:. Kardiyomiyosit Edu katılmasını ve ploidi ölçmek için strateji yolluk (A) Çekirdekler forward scatter (FSC) ve yan dağılım (SSC) dayalı enkaz ayırt edilir. (B) doğrusal kapı oluşturulur ve tekli çekirdek nüfus DAPI etiketleme ve alan sinyali vs 405/450/50 boyuna göre belirlenir. (C), Floresan yolluk kalp dokusunda kardiyomiyosit çekirdekleri (PCM-pozitif 1) ve non-kardiyomiyosit (PCM-1-negatif) çekirdeklerin ayrılmasını sağlar. DAPI sinyalinin (D) Yoğunluk DNA konsantrasyonu ve PCM1 + kardiyomiyosit nüfusun böylece çekirdek ploidi belirlemek için kullanılır. farede> kardiyomiyosit çekirdeklerinin% 80 diploid (2n) 'dir. (E) Floresan yolluk 2N ayrılmasını sağlar 2N Edu (PCM + / EDU + =% 0.9) dahil olması kardiyomiyosit çekirdeklerinin, Edu dahil değil kardiyomiyositlerde (PCM + EDU-). Tüm adımlar protokolde 5.0 detaylandırılmıştır. MDX gösterilmeleri Örnek -. C57 / BL10 arka plan üzerinde / y fareler bu rakamın daha büyük bir versiyonunu görmek için lütfen buraya tıklayınız.
{kind=link}
Sonuçlar
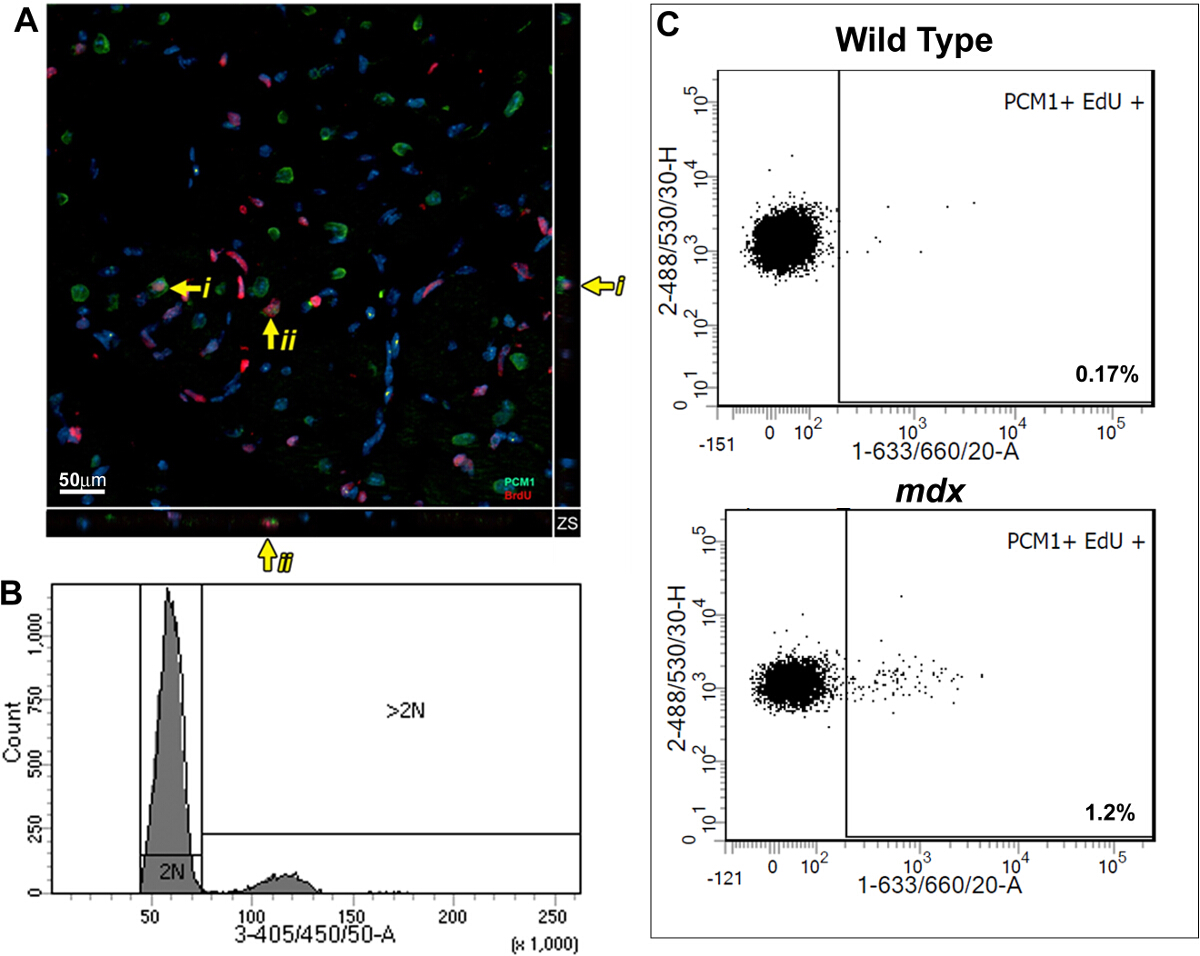
artan poliploidleşmeyi için kontrol ederken bu yöntem nedeniyle kardiyomiyosit çekirdeklerinin bölünme arttı edu kuruluş ölçümü sağlar. Bir BrdU bazlı bir deney kullanılarak, daha önce, memeli kalp yeni kardiyomiyositlerde rejenerasyonu ile, Duchenne kas distrofisi MDX fare modelinde kronik kardiyomiyosit kaybına cevap gösterdi. Biz daha da yayınlanan bulgularını doğrulamak ve protokol yarar göstermek için burada açıklanan yöntemleri kullandık. açıklanan protokol gereği; Yetişkin (12 haftalık) mdx ve kontrol bl10 fareler 7 gün boyunca edu bir günlük enjeksiyonu ile pulse edildi, deney grupları 5,13 arasında statik analiz izin önceden bildirmiştir. Darbeli kalplerin immünohistolojik analizi edu (Şekil 3A) dahil olması kardiyomiyositlerde ifade PCM1 varlığını ortaya koymaktadır. Ancak, veri elde istimalEdu dahil diğer hücre tipleri kardiyomiyositlerde olarak misidentified edilebilir g immünohistolojik yöntemler, yanlış yorumlanabilir. Bunun bir örneği, Ek Şekil 1A ve B'de gösterilmiştir. Ayrıca immünohistolojik yöntemler nükleer bölümünden poliploidleşmeyi ayrım yapmıyor. Bu protokol ayrıntılı sitometri analizi akış nedeniyle poliploidleşmeyi için edu dahil olan hücreleri hariç tutarak, mdx ve kontrol farelerde kardiyomyosit nükleer bölünme hızla miktarının sağlar. Daha önce yayımlanmış verilere 13 benzer şekilde, analiz yaş kıyasla mdx fare kalplerinde neo-kardiyomyosit çekirdekler nesil oranlarında bir artış ortaya denetimleri (Şekil 3B ve C% 0.17 karşı% 1.2) eşleşti.

Şekil 3: Edu Niceleme yabanitipte ve mdx kalplerinde kardiyomiyositlerde etiketli. Mdx kalplerin 40 mikron kalınlığında kesitler alınan Z-yığın projeksiyonlar (A) Görüntü. PCM-1 ifade kardiyomiyosit çekirdeklerinin Edu (kırmızı) yer vermiş (yeşil). Sarı ok Edu kardiyomiyositlerde etiketli gösterir. I ve II ayrı ayrı edu Z düzleminde gösterildiği kardiyomiyositlerde etiketli gösterir. (- C57 / BL10 arka plan üzerinde / y mdx) Çekirdekler DAPI (mavi) (B ve C) sitometri izole çekirdeklerin Flow 12-13 haftalık vahşi tip (C57 / BL10) ve mdx farelerden alınan temsili arsaları gösteren etiketli. (B) Histogram 2N ve> 2N çekirdekleri arasındaki DAPI yoğunluğu ve ayrımcılık gösteren. Bu protokol ayrıntılı olarak analiz edildiğinde 2N kardiyomiyosit çekirdeklerinin popülasyonunda edu birleşmesini gösteren (C) Arsalar. Tüm fareler edu 1'den 1 hafta uygulandıYaş 2 hafta. Bu rakamın büyük halini görmek için lütfen buraya tıklayınız.
{kind=link}
Ek Şekil 1:. PCM-1 kardiyomiyosit çekirdeklerinin için özel bir işaretleyici protokolünü kullanarak çekirdekleri ROSA nT / nG CRE hassas muhabir hattı 16 ile TNT-CRE fareler 14 geçerek üretilen çift transgenik fare izole edildi nitelendirdi. Troponin (TNT) promoterinin kontrolü altında elde edilen, fare cre aktivasyonu çekirdekler lokalize olan GFP geri dönüşümsüz ekspresyonuna yol açar. Çekirdekler GFP ifade kardiyomiyosit çekirdeklerinin nüfusu (488/530/30-A) tanımlamak için yeteneği gösterir İzotip kontrolü (Iso) ile etiketlenmiş. Anti-PCM1 ile çekirdeklerin Etiketleme (tespit ikincil antikor 633-660 / A 20) GFP ifade cardiomyocy ile PCM1 ifade korelasyon göstermektedirte çekirdekler nüfus. TNT promotör aktivitesi ile tanımlanan, PCM1 antikoru ile etiketli kardiyomiyosit çekirdeklerinin bu örnekte 98.8% olarak. Ek Şekil 1 indirmek için tıklayınız.
Ek Şekil 2:. Immünohistolojik tekniklerle kardiyomiyosit yenilenmesini ölçülmesi için Zorluklar (A) 2D görüntü de edu (kırmızı) için etiketlenmiş gibi görünen iki PCM-1 ifade kardiyomiyosit çekirdekleri (yeşil) gösterir. Sarı oklar Edu dahil olması kardiyomiyositlerde gibi görünen gösterir. Bu durum PCM1 ifade ve Edu etiketleme 2-boyutlu görüntüleme kullanılarak görüntülendi belirgin ortak yerelleştirme etmektir. (B) bu görüntünün bir 3 boyutlu projeksiyon Edu hücreleri kardiyomiyositlerde değil, PC örten olmayan kardiyomiyosit hücrelerdir etiketli tanımlarM1 kardiyomiyosit çekirdekleri ifade. Deneylerde kullanılan yaş 12. haftada C57 / BL10 fareler. Ek Şekil 2A indirmek için lütfen buraya tıklayınız. Ek Şekil 2B indirmek için buraya tıklayınız.
| Reaksiyon Bileşenleri | Reaksiyonların sayısını | |
| 2 | 5 | |
| PBS | 875 ul | 2.19 mi |
| bakır koruyucu | 20 ul | 50 ul |
| Floresan boya pikolil azid | 5 ul | 12.5 ul |
| Seyreltilmiş tepkime4.1 hazırlanan tampon | 100 ul | 250 ul |
| Toplam reaksiyon hacmi | 1 mi | 2.5 mi |
Tablo 2: edu kokteyl maddeler. Edu etiketleme kokteyl için reçete protokol bölüm 4 (Algılama edu esas) gerekli.
Tartışmalar
doğru Kardiyomiyosit ciro ölçmek için ve rejenerasyon deneyleri gerçek kardiyomiyosit üretimi ve nonprodüktif DNA bölümü ayırt etmeliyiz. Birçok çalışma, sadece siklin kinesis ve hücre döngüsü belirteçleri ifadesi aracılığıyla kardiyomiyosit çoğalması miktarının, sadece bu nonprodüktif olayları görmezden gelmeye devam. Bu nonprodüktif olaylar için kontrol kinayeli kalırken kardiyomyosit ciro doğru ölçümü sağlar tek bir yöntemi bugüne kadar. Özellikle bu kardiyomiyosit DNA replikasyonu 13 65% ~ kadar katkıda kardiyomiyosit poliploidleşmeyi hesaba güçtür. Bu nedenle Kardiyomiyosit nesil doğru ölçümü biz artan ploidleşmede sonuçlanan DNA replikasyonu hariç ise neo kardiyomiyosit çekirdeklerinin oranlarının güçlü ölçümü sağlar bir protokol geliştirdi yardımcı olmak. Her ne kadar bu protokol can neo-kardiyomiyosit cins arasında değil ayrımcılıkyon kardiyomyosit iki çekirdeklenme, hızla kullanılan ve doğru bir üst limit kardiyomiyosit nesil (ploidleşmede için muhasebe) hesaplamak mümkündür. Bu protokol, bu nedenle hastalık modellerinde kardiyomiyosit üretimi ve poliploidleşmeyi oranlarında potansiyel değişiklikleri değerlendirmek için ya da terapötik potansiyel etkinliğini değerlendirmek için bir tarama aracı sağlar. Neo-kardiyomiyosit çekirdekler üretimi hızındaki değişiklikler bu protokolü kullanılarak tespit edildikten sonra kardiyomiyosit çekirdeklenme sayısının kardiyomiyosit nesil değişiklikler nedeniyle, bu, daha önce tarif edildiği gibi 2,13,17,18, eğer daha sonraki çalışmalarda tespit etmek için kullanılabilir. Bunlar darbe döneminde histolojik miktar kardiyomiyosit çekirdeklenme dinamikleri kullanımını içerir veya mononükleer ve çok çekirdekli kardiyomiyosit popülasyonlarında edu birleşmesini karşılaştırmak için edu darbeli hayvanlardan elde edilen doku kesitlerinde analiz eder.
Nedeniyle kardiyomiyosit ciro bu düşük seviyelerdeprotokol 7 günlük bir süre içinde edu birden enjeksiyonları kullanır. Bu aynı zamanda Kardiyomiyosit nesil tüm potansiyel hücresel kaynakların "peşinde" ve bu süre boyunca kümülatif kardiyomiyosit çekirdeklerinin nesil ölçümü izin verir. Çalışmada bağlı olarak, bu zaman aralığı kardiyomiyosit nesil tahmin düzeylerine uygun şekilde ayarlanabilir. kardiyomiyosit çekirdeklerde edu dahil doğru ölçümü için, PCM-1 reaktivitesini tespit etmek için kullanılan ikincil antikor ile çekirdeklerin herhangi spesifik olmayan etiketleme olup zorunludur. Nedenle, bunun dışında, bir ikincil antikor, bu protokol, kullanılacak önerilen, özellikle protokol bu yönünü optimize etmek için ilave ikincil antikor titrasyon deneyleri gerçekleştirmek için mantıklı olacaktır. Burada açıklanan protokol kardiyomiyosit çekirdekleri tanımlamak için PCM-1 ifadesini kullanır. Bu kurulu bir kardiyomiyosit belirtecidir Ancak, alternatif markörler doğrulamak için kullanılabilirveri; Bu, kısmen kardiyomiyosit çekirdekleri 1. lokalize olarak tanımlanmıştır cTnT'nin için spesifik antikorları içerir. Benzer şekilde, alternatif çekirdek lokalize proteinler kardiyomiyositlerin dışındaki çekirdek popülasyonlarda edu birleşmesini belirlemek ve ölçmek için kullanılabilir. DNA sentezinin akıbeti henüz bilinmiyor ve hücre bölünmesi ya da artmış ploidleşmede ya neden olabilir aktif olarak mitoz geçiren tüm kardiyomiyosit çekirdekler, analize dahil edilmesi önemlidir. PCM1 nedenle mitoz geçiren PCM1 ifade tarafından tespit olmayacaktır kardiyomiyositlerinin hücre döngüsünün M fazında demonte edilir. Buna ek olarak, hücre döngüsünün S fazında Tüm çekirdekler daha sonra analize dahil edilmelidir. Bu 2N ve 4N popülasyonlar arasındaki bir DAPI yoğunluğu da dahil olmak üzere 2N nüfus üzerinde bir DAPI yoğunluğu ile tüm çekirdekleri dışarı yolluk elde edilebilir.
giderek rağmenKalp normal yaşlanma ve sonrası akut yaralanma sırasında kardiyomiyositlerde yerine kapasitesine sahip olduğunu kabul, bu potansiyelin kaynağı ve derecesi tartışmalıdır. Buna ek olarak, kardiyomiyosit devir farklı oranları 1,7,20-22 bildirilmiştir. Bu, doğru tanımlanması ve neo-kardiyomiyosit Üretimi 19 miktarının zorluklara kısmen olabilir. Çalışmaların büyük çoğunluğu sadece histolojik analizinin kullanımı kardiyomyosit devir ve yenileme 2,4,23,24 miktarının sarkomerinde proteinler de dahil olmak üzere sitoplazmik proteinlerin ekspresyonu ile kardiyomiyositlerin tanımlanması dayanmıştır Bugüne kadar. Bu yöntemlerin kullanılması çoğalması markerlerinin ifadesi tespit etmek için, ya da burada gösterildiği gibi, timidin analogları dahil kolayca kardiyomiyositlerde ve diğer kardiyak hücre tiplerinin hatalı tanımlama neden olabilir. 3D konfokal görüntüleme kullanımı inci bu sorunları hafifletmek için yardımcı olabilirESE yöntemler pahalı ve zaman alıcıdır. İlginç bir şekilde, burada açıklanan protokol, neo-kardiyomiyosit çekirdekler üretimi haftada% 0.17 oranında oluşur göstermektedir. Bu kadar 0.13% 5 haftalık ciro oranlarını gösteren bazlı çalışmalar sitometri diğer akışı ile tutarlıdır. Önceki çalışmalarda 2,5,25,26 gibi bu verilere dayalı yıllık cirosu oranları tahmin etmek cazip olmasına rağmen ciro oranları bir hayvanın 13 yaşam süresi boyunca dinamik olarak, bu uygunsuz.
Bu yöntem kardiyomyosit rejenerasyonu artırmak için potansiyel terapötikler değerlendirilmesi veya kardiyomiyosit ciro kalp hastalığı etkilerini kardiyomyosit poliploidleşmeyi oranları araştıran dahil olmak üzere potansiyel uygulamaları bir dizi vardır.
Açıklamalar
The authors have nothing to disclose.
Teşekkürler
This work was supported by the British Heart Foundation, project grant PG/13/69/30454.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 0.32 M sucrose | Sigma | 84100 | |
| 10 mM Tris-HCl (pH = 8) | Sigma | T3253 | |
| 5 mM CaCl2 | Sigma | c5086 | |
| 5 mM magnesium acetate | sigma | M-5661 | |
| 2.0 mM EDTA | Sigma | E5134 | |
| 0.5 mM EGTA | Sigma | 63779 | |
| 1 mM DTT | Sigma | D0632 | |
| 70 mM KCl | Sigma | P9541 | |
| 10 mM MgCl2 | Sigma | M8266 | |
| 1.5 mM spermine | Sigma | 85590 | |
| Isotype rabbit IgG- ChIP Grade | abcam | abc7415 | |
| Rabbit anti-PCM-1 antibody | Sigma | HPA023374 | |
| Alexa Fluor 488 F(ab')2 Fragment of Goat Anti-Rabbit IgG (H+L) Antibody | Life technologies | A-11070 | |
| cell strainers 70 μm and 100 μm | Fisher scientific | 11597522, 11517532 | |
| Glass dounce (40 ml) and pestle large clearance | Sigma | D9188-1SET | |
| EdU (5-ethynyl-2’-deoxyuridine) | Life technologies | A10044 | |
| Click-iT Plus EdU Alexa Fluor 647 Flow Cytometry Assay Kit | Life technologies | C10634 | This kit inlcudes reagents required for section, EdU reaction buffer, EdU fixative, saponin-based permeabilization solution and the reagents required for the EdU labelling cocktail. |
| CyStain DNA 2 step kit, | Sysmex Partec | 05 5005 | This kit inlcudes reagents required for DAPI labelling (DNA staining solution) |
| Probe homogeniser, e.g., TissueRuptor | Qiagen | 9001273 | |
| TissueRuptor Disposable Probes | Qiagen | 990890 | |
| ultracentrifuge | Sorvall | ||
| Facscanto II | BD Biosciences | ||
| Ultracentrifuge Tube, Thinwall, Polypropylene. 38.5 ml, 25 x 89 mm | Beckman Coulter | 326823 | |
| Bovine serum albumin | Sigma | A2153 |
Referanslar
- Bergmann, O., et al. Evidence for cardiomyocyte renewal in humans. Science. 324, 98-102 (2009).
- Senyo, S. E., et al. Mammalian heart renewal by pre-existing cardiomyocytes. Nature. 493, 433-436 (2013).
- Soonpaa, M. H., Rubart, M., Field, L. J. Challenges measuring cardiomyocyte renewal. Biochim Biophys Acta. 1833, 799-803 (2013).
- Loffredo, F. S., Steinhauser, M. L., Gannon, J., Lee, R. T. Bone marrow-derived cell therapy stimulates endogenous cardiomyocyte progenitors and promotes cardiac repair. Cell Stem Cell. 8, 389-398 (2011).
- Malliaras, K., et al. Cardiomyocyte proliferation and progenitor cell recruitment underlie therapeutic regeneration after myocardial infarction in the adult mouse heart. EMBO Mol Med. 5, 191-209 (2013).
- Hsieh, P. C., et al. Evidence from a genetic fate-mapping study that stem cells refresh adult mammalian cardiomyocytes after injury. Nat Med. 13, 970-974 (2007).
- Bergmann, O., et al. Identification of cardiomyocyte nuclei and assessment of ploidy for the analysis of cell turnover. Exp Cell Res. 317, 188-194 (2011).
- Herget, G. W., Neuburger, M., Plagwitz, R., Adler, C. P. DNA content, ploidy level and number of nuclei in the human heart after myocardial infarction. Cardiovasc Res. 36, 45-51 (1997).
- Scholzen, T., Gerdes, J. The Ki-67 protein: from the known and the unknown. J Cell Physiol. 182, 311-322 (2000).
- Carmena, M., Earnshaw, W. C. The cellular geography of aurora kinases. Nat Rev Mol Cell Biol. 4, 842-854 (2003).
- Bergmann, O., Jovinge, S. Isolation of Cardiomyocyte Nuclei from Post-mortem Tissue. J. Vis. Exp. , e4205 (2012).
- Gilsbach, R., et al. Dynamic DNA methylation orchestrates cardiomyocyte development, maturation and disease. Nat Commun. 5, 5288 (2014).
- Richardson, G., Laval, S., Owens, W. A. Cardiomyocyte regeneration in the mdx mouse model of non-ischemic cardiomyopathy. Stem Cells Dev. , (2015).
- Jiao, K., et al. An essential role of Bmp4 in the atrioventricular septation of the mouse heart. Genes Dev. 17, 2362-2367 (2003).
- Bergmann, O., et al. Cardiomyocyte renewal in humans. Circ Res. 110, e17-e18 (2012).
- Prigge, J. R., et al. Nuclear double-fluorescent reporter for in vivo and ex vivo analyses of biological transitions in mouse nuclei. Mamm Genome. , (2013).
- Naqvi, N., et al. A proliferative burst during preadolescence establishes the final cardiomyocyte number. Cell. 157, 795-807 (2014).
- Liu, Z., Yue, S., Chen, X., Kubin, T., Braun, T. Regulation of cardiomyocyte polyploidy and multinucleation by CyclinG1. Circ Res. 106, 1498-1506 (2010).
- Ang, K. L., et al. Limitations of conventional approaches to identify myocyte nuclei in histologic sections of the heart. Am J Physiol Cell Physiol. 298, C1603-C1609 (2010).
- Kajstura, J., et al. Myocyte turnover in the aging human heart. Circ Res. 107, 1374-1386 (2010).
- Kajstura, J., et al. Cardiomyogenesis in the adult human heart. Circ Res. 107, 305-315 (2010).
- Walsh, S., Ponten, A., Fleischmann, B. K., Jovinge, S. Cardiomyocyte cell cycle control and growth estimation in vivo--an analysis based on cardiomyocyte nuclei. Cardiovasc Res. 86, 365-373 (2010).
- Anversa, P., Leri, A., Kajstura, J. Cardiac regeneration. J Am Coll Cardiol. 47, 1769-1776 (2006).
- Gonzalez-Valdes, I., et al. Bmi1 limits dilated cardiomyopathy and heart failure by inhibiting cardiac senescence. Nat Commun. 6, 6473 (2015).
- Kimura, W., et al. Hypoxia fate mapping identifies cycling cardiomyocytes in the adult heart. Nature. 523, 226-230 (2015).
- Soonpaa, M. H., Field, L. J. Assessment of cardiomyocyte DNA synthesis in normal and injured adult mouse hearts. Am J Physiol. 272, H220-H226 (1997).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiDaha Fazla Makale Keşfet
This article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır