
Method Article
Одновременная оценка кардиомиоцитов синтеза ДНК и плоидности: метод для оказания помощи квантификации кардиомиоцитов регенерации и оборота
В этой статье
Резюме
Quantification of cardiomyocyte turnover is challenging. The protocol described here makes an important contribution to this challenge by enabling accurate and sensitive quantification of neo-cardiomyocyte nuclei generation and nuclei ploidy.
Аннотация
Хотя принято считать, что сердце имеет ограниченный потенциал для регенерации кардиомиоцитов после травмы и что низкий уровень оборота кардиомиоцитов происходят во время нормального старения, количественная оценка этих событий остается сложной. Это происходит частично из-за редкости процесса и тот факт, что многочисленные клеточные источники способствуют инфарктом обслуживания. Кроме того, дублирование ДНК в кардиомиоцитах часто приводит к полиплоидных кардиомиоцитов и лишь в редких случаях приводит к новым кардиомиоцитов путем клеточного деления. Для того, чтобы точно определить их кардиомиоцитов дискриминации оборот между этими процессами имеет важное значение. Протокол, описанный здесь, использует долгосрочный нуклеозида маркировку для того, чтобы обозначить все ядра, которые возникли в результате репликации ДНК ядер и кардиомиоцитов, идентифицированных с использованием ядер изоляции и последующему PCM1 иммунноокрашивания. Вместе это позволяет точную и чувствительную идентификацию нуклеозида мечения Cardiomyocyte ядер населения. Кроме того, 4 ', 6-диамидино-2-фенилиндол маркировка и анализ ядер плоидности, позволяет дискриминацию нео-кардиомиоцитов ядер из ядер, которые включили нуклеозида в течение полиплоидизацией. Несмотря на то, что этот способ не может контролировать для кардиомиоцитов binucleation, она позволяет быстрое и надежное определение количества нео-кардиомиоцитов ядер в то время как учет полиплоидизацией. Этот метод имеет ряд последующих применений, включая оценку потенциальных терапевтических средств для улучшения регенерации кардиомиоцитов или изучения влияния болезни сердца на обороте кардиомиоцитов и плоидности. Этот метод также совместим с дополнительными ниже по течению иммуногистологических методов, позволяющих количественно оценить нуклеозид-инкорпорации во всех типах клеток сердца.
Введение
В последние годы наблюдается накопление доказательств оспорить предположение , что сердце является терминально дифференцированными, постмитотическими 1,2 органа. Тем не менее, количественное определение оборота кардиомиоцитов и регенерации остается сложной.
Трудности в точной идентификации редкого поколения кардиомиоцитов с использованием стандартных иммуногистохимических методов хорошо сообщили 3. Кроме того, сотовый источник кардиомиоцитов поколения остается неопределенной с доказательствами для вкладов пролиферации кардиомиоцитов, а также путем дифференцировки стволовых клеток 4-6. Таким образом, использование модели линии дифференцировки трассировки, которые требуют знания фенотипом кардиомиоцитов прародителей невозможно и количественное определение пролиферации в одной популяции, в том числе кардиомиоцитов, является несоответствующим. Кроме того, кардиомиоциты имеет потенциал для эндорепликации без кариокинеза (в результате чего автомобиль полиплоидногоdiomyocyte) или кариокинез при отсутствии цитокинеза ( что приводит к двуядерных кардиомиоцитов) 7,8. Точная количественная оценка оборота кардиомиоциты зависит от способности различать между этими событиями и истинным нео-кардиомиоциты поколения. Это создает уникальные осложнения , поскольку репликация ДНК и экспрессия циклин-зависимых киназ в кардиомиоциты не только демонстрируют истинное деление клеток 9,10.
Для оказания помощи в количественной оценке нео-кардиомиоцитов поколения, мы объединили устоявшуюся ядер технику изоляции и иммунологическую маркировки pericentriolar материала 1 (PCM1) для идентификации ядер кардиомиоцитов , как описано Бергман и др. 7,11 с новыми методами долгосрочной перспективе мечении ДНК и анализ плоидности. PCM-1 представляет собой центросома белок, который накапливается на поверхности ядра дифференцированных, без велосипедного миоцитов. Предыдущие исследования показали, что антитела противPCM-1 специально маркировать ядер кардиомиоцитов 7,11 и как таковой PCM1 был использован ряд независимых групп для выявления кардиомиоциты 1,12,13. Кроме того, мы показали , что экспрессия PCM1 карты генетически меченых ядер кардиомиоцитов в TnT-Cre трансгенной модели мыши 14 (Дополнительный рисунок 1).
Протокол , описанный здесь , обеспечивает точное и чувствительное идентификацию нео поколения кардиомиоцитов ядер в сердце мышей, независимо от клеточных происхождения (Фигура 1А и В), одновременно исключая нуклеозида маркировки результате полиплоидизации из анализа (рис 1C и D). Несмотря на то, что этот способ не может контролировать для кардиомиоцитов binucleation, она позволяет быстрое и надежное определение количества нео-кардиомиоцитов ядер, которая требуется для точного количественного оборота кардиомиоцитов. Более того,она обеспечивает быстрый инструмент скрининга для оценки потенциальных изменений в динамике поколения кардиомиоцитов.
В то время как маркировка ДНК, как правило, включает в себя 5-бром-2'-дезоксиуридина (BrdU) в качестве аналога тимидина, протокол, описанный здесь, использует 5-этинил-2'-дезоксиуридин (ОБР) на основе анализа, поскольку она требует меньшего количества стадий обработки для более быстрого сквозными ставить и не требует денатурации ДНК для иммунологического обнаружения, что делает его совместимым с другими иммунным окрашиванием протоколами и, таким образом, увеличивая потенциальные применения ниже по потоку от способа.

Рисунок 1: Непрерывное пульсирующее с EDU этикетки нео-кардиомиоциты независимо от их предшественников. (А) Edu включена в ДНК кардиомиоцитов во время деления клетки. Распространение в популяции кардиомиоцитов будет Ресулт в увеличении или замене кардиомиоцитов и поэтому продуктивный синтез ДНК (способствует поддержанию тканей и ремонт). (В) Edu включена в ДНК сердечных клеток - предшественников во время деления клетки. Это будет сохранено в ячейке во время дифференцировки в кардиомиоциты линии. Эта дифференцировка стволовых клеток также приведет к увеличению количества кардиомиоцитов и, следовательно, способствует поддержанию и восстановлению тканей. (C) кардиомиоциты имеют потенциал , чтобы пройти "непродуктивный" репликацию ДНК , что приводит к увеличению кардиомиоцитов плоидности, что связано с кардиомиоцитов гипертрофии и ремоделирования миокарда, но не заменяет потерянные кардиомиоциты. Процесс полиплоидизацией отличается от binucleation как это приводит к кардиомиоцитов с одного ядра, который содержит четыре или более групп из двух гомологичных хромосом (> 2N). (D) После непрерывного ядра рulse, этот протокол описывает ядер выделения и идентификации ядер кардиомиоцитов экспрессии PCM1, чтобы количественно оценить как кардиомиоциты плоидности и Эду регистрации. Выражение PCM1 и Эду включение обнаружено с помощью проточной цитометрии. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
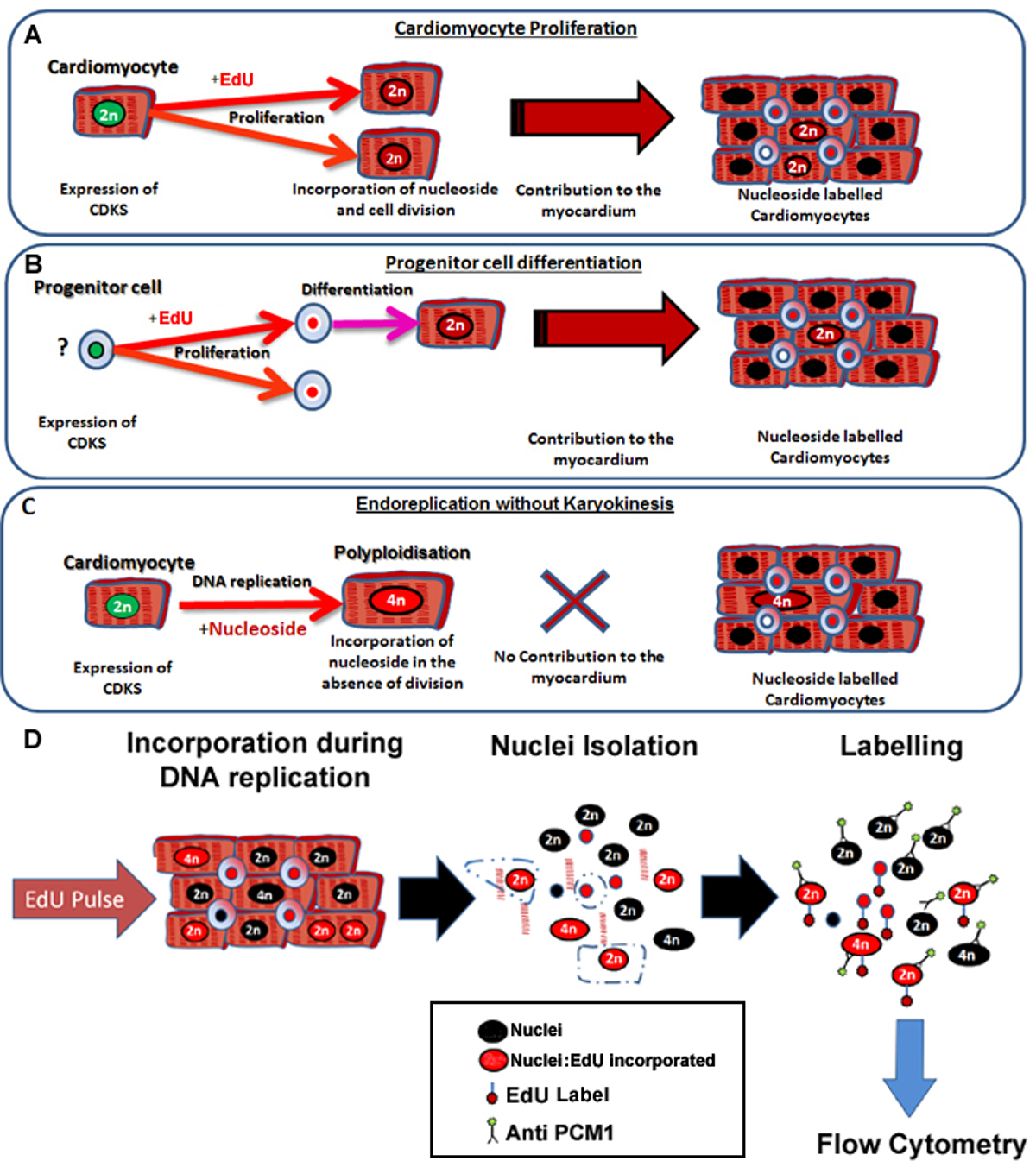{kind=link}
протокол
Работа для животных была утверждена и одобрена наблюдательным советом Университета Ньюкасла по этике. Все процедуры на животных были проведены в соответствии с руководящими принципами из Директивы 2010/63 / ЕС Европейского парламента по вопросу о защите животных, используемых для научных целей.
1. Администрация EDU
- Растворить в ОБР стерильным физиологическим раствором (0,9% вес / об) при конечной концентрации 12,5 мг / мл.
- Раствор нагревают до 40 ° С и вихрем для того, чтобы полностью раствориться. Хранить достаточное количество ОБР / физиологическом растворе при 4 ° С, чтобы впрыснуть всех мышей в исследовании, что позволяет в течение 6 ежедневных инъекций.
Примечание: Как правило более 6 мышей требуются для каждой экспериментальной группы. Однако расчеты мощности должны быть выполнены для независимых исследований.
- Раствор нагревают до 40 ° С и вихрем для того, чтобы полностью раствориться. Хранить достаточное количество ОБР / физиологическом растворе при 4 ° С, чтобы впрыснуть всех мышей в исследовании, что позволяет в течение 6 ежедневных инъекций.
- Взвешивание отдельных мышей. Рассчитать необходимый объем для администрирования 100 мкг / г EDU / солевого раствора к каждой мыши (8 мкл акций Edu раствора на грамм).
- Проведем соответствующий Volumе / Edu физиологический раствор в шприц инсулина с иглой 25 G.
- Выполните внутрибрюшинно (IP) инъекции при обработке мышей с использованием утвержденных методов домашнего офиса.
- Поместите мышь на крышке клетки. Осторожно потяните на основание хвоста и крепко возьмитесь мышь за шкирку.
- Выставляют живота для инъекций, держа мышь с указательным и большим пальцем у основания головы и наклоняя голову животного назад по направлению к полу.
- Протрите живота с 70% -ным раствором спирта. Найдите среднюю линию животного и нижний правый квадрант. Вводят раствор EDU / физиологического раствора животного в нижний правый квадрант.
- Вводят мышей с EDU / солевом растворе и фиксируют время суток.
- Повторите шаги от 1,2 до 1,4, в то же время суток, в течение 6 дней подряд.
Примечание: В качестве отрицательного контроля Эду требуется установить стробирования для анализа проточной цитометрии, вводят дополнительный возраст, пол и экспериментально совпадаютред мышь с эквивалентным объемом физиологического раствора без Эду. Это обеспечит необходимый контроль (без управления EDU). Этот контроль должен быть включен для каждого исследования, чтобы учесть различия в партиях реагентов и цитометра настройки.
2. Сбор сердца
- Через 24 ч после 6 - й инъекции Edu, пожертвуйте мышей с помощью цервикальной дислокации (или альтернативного графика , утвержденного 1 Home Office метод).
- Поместите жертвенного животного в положении лежа на спине и сделать разрез кожи от середины живота к диафрагме с хирургическими ножницами.
- Вырезать диафрагму, и удерживая грудину от полости тела, срезанные на двусторонней основе, чтобы обнажить сердце.
- Слегка приподнимите сердце и перерезать основные кровеносные сосуды тракта оттока рассекать сердце из грудной полости.
- Сразу же место каждое сердце в физическому лицу 15 мл центрифужную пробирку, содержащую 10 мл PBS, охлаждали до 4 ° С.
- Передача сердца SEPлярном 10 см чашки Петри, используя скальпель вырезать каждое сердце в двух в сагиттальной ориентации и моют сердце, осторожно сжимая с парой щипцов. Затем замените PBS, чтобы удалить как можно больше крови, насколько это возможно. Повторите этот шаг дважды.
- Поместите обе части каждого сердца в той же индивидуальной 1,5 мл микроцентрифужных трубки.
- Переходите к шагу 3 или альтернативно хранить образцы -80 ° C, как описано в пунктах 2.9 и 2.10.
- С помощью 25 G иглы, сделать небольшое отверстие в крышке микроцентрифужных трубки, чтобы предотвратить крышку от отверстия за счет расширения газов, когда сердце обрабатывается.
- Добавьте каждый микроцентрифужных трубки и поместить в емкость с жидким азотом для замораживания.
3. Ядра диссоциация и кардиомиоцитов Ядра Этикетировочное
Примечание: Это ядра маркировки диссоциации и кардиомиоцитов ядер приспособлена от такового Бергманн и др 11 Выполните эту процедуру на всех образцах инжектированных.с Эду и солевым раствором закачиваемой (отсутствие управления EDU) отрицательного контроля. Приведены в таблице 1 для рецепта решений , необходимых для выполнения этих шагов.
- Для каждого сердца, чтобы проанализировать, поместите 36 мл 1% BSA / PBS раствора в ультрацентрифужную пробирку, инкубировать в течение 30 мин, вылить, а затем дайте высохнуть на воздухе.
Примечание: Эта трубка используется на этапе 3.6. - Поместите отдельные замерзшие сердца на 10 мм блюдо на льду и пропустить через мясорубку, используя скальпель, позволяющий сердца размораживать в процессе мясорубки.
- Поместите фарш сердца в 15 мл буфера для лизиса клеток в центрифужные пробирки объемом 50 мл и гомогенизируют с помощью гомогенизатора зонда в течение 15 секунд при 25000 оборотов в минуту при комнатной температуре.
- Добавить еще 15 мл буфера для лизиса клеток в пробирку центрифуги и переносят в 40 мл гомогенизатора Даунса с большим зазором пестиком. Выполните 10 ударов с пестиком и процеживают через 100 мкм затем мкм клеточный фильтр 40 в центрифужную пробирку на 50 мл.
- Центрифуга в течение 10 мин медленнот 700 мкг при 4 ° C и удалить супернатант.
- Ресуспендируют ядерный осадок в 25 мл раствора в градиенте плотности сахарозы и слой Ядра клеток раствора на вершине 10 мл свежего раствора градиенте сахарозы в ультрацентрифуге трубки, полученного в разделе 3.1.
- Центрифуга при 13000 мкг в течение 1 ч при 4 ° С, используя качаться роторной трубы.
- После центрифугирования, отбросить супернатант и ресуспендируют ядер гранул в 1300 мкл ядер буфера для хранения в 1,5 мл трубки микроцентрифужных и этикетки PCM1.
- Удалить 300 мкл суспензии в новую пробирку микроцентрифужных и добавляют к 700 мкл свежего буфера для хранения ядер. Добавьте эту трубку в качестве контроля ISO.
- Добавить анти-PCM1 при конечной концентрации 8 мкг / мл в микроцентрифужных пробирку PCM1 и добавить кролика IgG-антитела контрольного изотипа в концентрации 8 мкг / мл в микроцентрифужных пробирку управления ISO.
- Инкубируют в течение ночи при температуре 4 ° С. После incubвания, центрифуге при 700 мкг в течение 10 мин, отбросить супернатант, заменить 1 мл свежего буфера для хранения ядра, ресуспендируют и центрифуга снова в течение 10 мин при 700 х г. Удалите супернатант и заменить 1 мл свежего буфера для хранения ядра.
- Добавить 1 мкл F (аb ') 2-фрагмент козьих антител против кроличьего IgG (H + L) антитела (FITC или эквивалентного зеленого флуоресцентного красителя), чтобы достичь конечной концентрации 2 мкг / мл.
Примечание: Использование F (аb ') 2-фрагмент антитела, уменьшает вероятность возникновения неспецифического мечения клеток иммунной системы, включая макрофаги и В-клетки. - Wrap трубы в алюминиевой фольгой для защиты от света и инкубировать в течение 1 часа при 4 ° С.
4. Обнаружение Edu инкорпорации
- Развести 10x концентрируют Edu реакционного буфера в соотношении 1:10 деионизированной водой.
- Центрифуга PCM-1 и управления меченых ядер суспензий ISO при 700 мкг в течение 10 мин, отбросить супернатант и ресуспендируют осадок ядер в 1 мл1% БСА / раствор PBS. Вымойте ядер гранул дважды.
- После последней промывки, отбросить супернатант и заменить 100 мкл Edu закрепителя.
- Обертка трубы в алюминиевой фольгой для защиты от света и инкубировать при комнатной температуре в течение 15 мин.
- Промыть образцы дважды 1 мл 1% БСА / раствор PBS, центрифугируют при 700 х г и инкубировать с 1x сапонина на основе раствора пермеабилизации при комнатной температуре в течение 15 мин.
- Приготовьте 1х EDU маркировки коктейль (таблица 2). Минимум 2 реакций необходимо будет не включать никакого контроля Edu.
- Добавить 500 мкл 1x EDU маркировки коктейль непосредственно к каждому образцу, уже содержащей ядра в 100 мкл 1х сапонина раствора на основе пермеабилизации и инкубировать при комнатной температуре в защищенном от света месте в течение 30 мин.
- Центрифуга при 700 мкг и ресуспендируют в 1 мл 1% BSA / PBS и повторить этот шаг еще два раза.
- Центрифуга при 700 мкг, отбросить супернатант и заменить 400 мкл раствора ДНК окрашивания (см Таблицу материалов).
| Название реагента |
| 1. буфера для лизиса клеток |
| 0,32 М сахарозы |
| 10 мМ Трис-HCl (рН = 8) |
| 5 мМ CaCl 2 |
| 5 мМ ацетат магния |
| 2,0 мМ ЭДТА |
| 0,5 мМ EGTA |
| 1 мМ ДТТ |
| 2.Sucrose Градиент Solution |
| 2,1 М сахарозы |
| 10 мМ Трис-HCl (рН = 8) |
| 5 мМ ацетат магния |
| 1 мМ ДТТ |
| 3. Ядра буфер хранения (NSB) |
| 0,44 М сахарозы |
| 10 мМ Трис-HCl (рН = 7,2) |
| 70 мМ KCl |
| 10 мМ MgCl 2 |
| 1,5 мМ спермина |
Таблица 1: Ядра буферные изоляция ингредиенты Рецепт для всех буферов , используемых в разделе протокола 3 (ядер и диссоциации кардиомиоцитов ядер маркировки)..
5. Поток Cytometric Анализ
Примечание: Выполните проточной цитометрии на изолированных ядрах , как описано ранее 7,11,15.
- Во- первых, создать сюжет , описывающий рассеяния вперед (FSC) и боковое рассеивание (SSC) , чтобы позволить дискриминацию ядер от мусора (Рисунок 2А).
- Создание сюжет дискриминировать одиночных ядер из агрегатов (Фигура 2В), откладывая 4 ', 6-диамидино-2-фенилиндол) (DAPI) область (3-405 / 450/50-А) по сравнению с высотой (33-405 / 450 / 50-Н). Обеспечить ТНАт-А 50 сигнал 3-405 / 450 / собрана на линейном масштабе, чтобы обеспечить правильное определение содержания ДНК.
- Создать график, описывающий 488/535/30-A (PCM-1) против SSC и отображать только события из синглетного ворот, созданного в 5.2, чтобы оценить PCM-1 выражение в популяции синглетного.
- Выполнить контрольный образец изотипа для того , чтобы установить ИКМ-1 положительные ворота (фиг.2с). Выполнить небольшое количество PCM1 меченый образец для проверки PCM1 выражения населения и литниковой позицию.
- Создать сюжет, описывающий SSC-A против 405/450/50-А (для обнаружения DAPI). На этом графике, отображение только синглетному PCM1, выражающих ядра, используя ворота, созданные в 5.3. Используйте этот участок , чтобы создать дополнительный иерархический ворота , который содержит только 2н ядра (рис 2D).
- Создать график, описывающий 488/535 / 30-A против 1-633 / 660/20-A, чтобы идентифицировать Edu маркировке PCM1, выражающей кардиомиоцитов населения. Показывать только ядра, которые являются синглет, P СМ1 + и 2н с использованием вышеуказанного стробирования.
- Запуск образца сердце от полного отсутствия контроля Эду установить Edu положительные ворота (рис 2E). Создать соответствующие ворота для Edu + клеток.
- Запись и количественного синглет, PCM-1, выражающий 2N ядер, которые включены EDU.
- Вычислить эту популяцию в процентах от общего синглета, PCM1 + ядер. Это обеспечит скорость нео-кардиомиоцитов ядер поколения, в течение 7-дневного периода ОБР импульса, как процент от общего количества ядер кардиомиоцитов.
Примечание: В качестве DAPI связывания с молекулой ДНК является стехиометрическим интенсивность флуоресценции пропорциональна количеству ДНК. Определение плоидности на основе интенсивности / 450/50-А сигнала 405, как было описано выше 7. - Для оценки плоидности в пределах общей популяции кардиомиоцитов использовать участок , установленный в 5.5 и стратегию , основанную на стробирования концентрации ДНК 7.
/files/ftp_upload/53979/53979fig2.jpg "/>
Рис . 2: стробирования стратегию количественной оценки кардиомиоцитов Edu включения и плоидности (A) Ядра подвергаются дискриминации от мусора на основе рассеяния вперед (FSC) и бокового рассеяния (SSC). (Б) Линейное ворота создается и синглетный популяция ядер определяется на основе на маркировке DAPI и 405/450/50 высоты против сигнала площади. (С) Флуоресцентные стробирования позволяет разделение ядер кардиомиоцитов (ПКМ-1-положительных) и некардиомиоцитов (PCM-1-отрицательных) ядер от сердечной ткани. (D) Интенсивность сигнала DAPI используется для определения концентрации ДНК и , таким образом , ядер плоидности кардиомиоцитов популяции PCM1 +. У мышей> 80% ядер кардиомиоцитов диплоидны (2n). (E) Люминесцентная стробирования позволяет разделить 2н, ядер кардиомиоцитов , которые включали Эду (PCM + / + EDU = 0,9%) от 2N, кардиомиоциты , которые не включены EDU (PCM + обученным), Все этапы подробно описаны в протоколе 5.0. Пример показан от MDX -. / У мышей на фоне C57 / BL10 Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
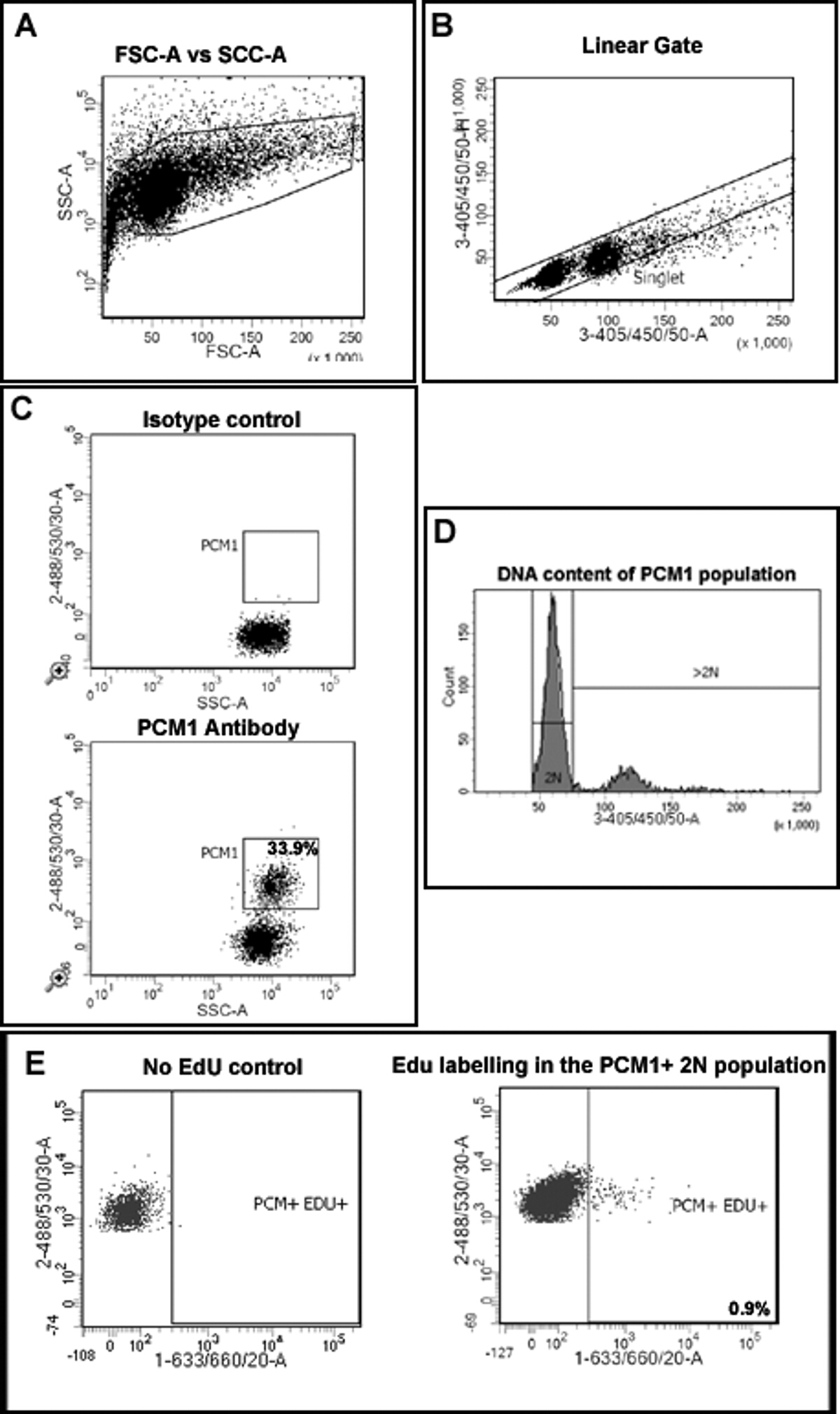{kind=link}
Результаты
Этот метод позволяет количественно оценить повышенной Edu включения из-за кардиомиоцитов ядер деления контролируя при этом для увеличения полиплоидизации. С помощью анализа , основанного BrdU, ранее мы показали , что сердце млекопитающего реагирует на хроническую потерю кардиомиоцитов, в мышиной модели MDX мышечной дистрофии Дюшенна, с регенерацией новых кардиомиоцитов. Мы использовали методы, описанные здесь, для дальнейшей проверки наших опубликованных результатов и продемонстрировать полезность протокола. В соответствии с протоколом, описанным; взрослые (12 недель) MDX и контрольных мышей BL10 импульсно одной ежедневной инъекции ОБР в течение 7 дней, уже сообщалось ранее , чтобы позволить статически анализ между экспериментальными группами 5,13. Иммуногистохимического анализа импульсных сердца демонстрирует наличие PCM1 , выражающую кардиомиоцитов , которые включены EDU (рис 3А). Тем не менее, полученные данные Усинг иммуногистологические методы могут быть неправильно истолкованы, как и другие типы клеток, которые включены EDU может быть ошибочно, как кардиомиоциты. Примером этого демонстрируется в дополнительном рисунке 1А и В. Кроме иммуногистологические методы не различают полиплоидизацию от ядерного деления. Проточной цитометрии анализа подробно в данном протоколе позволяет быстро количественно оценить кардиомиоцитов ядерного деления в MDX и контрольных мышей, в то время как за исключением клеток , которые были включены EDU вследствие полиплоидизации. Как и ранее опубликованных данных 13, анализ показал увеличение темпов нео-кардиомиоциты ядер поколения в сердцах MDX мышей по сравнению с возрастом контрольной группы (1,2% против 0,17%; Рисунок 3B и C).

Рисунок 3: Количественное Edu маркированы кардиомиоцитов в дикого типа и MDX сердца. (A) Изображение проекций Z-стека , взятых из 40 мкм толщиной секций MDX сердца. PCM-1, выражающие ядер кардиомиоцитов (зеленый), который включил Эду (красный). Желтая стрелка указывает Edu помечены кардиомиоциты. я и II указывает на индивидуальный Edu помечены кардиомиоциты, показанные на Z-плоскости. Ядра помечены DAPI (синий) (B и C) проточной цитометрии изолированных ядер показывает репрезентативные участки от 12-13 недель старого дикого типа (C57 / BL10) и MDX мышей (MDX - / у на C57 / BL10 фоне). (B) Гистограмма показывает интенсивность DAPI и дискриминации между 2N и> ядрами 2N. (C) Участки , показывающие включение Edu в ядрах населения 2н кардиомиоцитов при анализе , как описано в данном протоколе. Для всех мышей Эду вводили в течение 1 недели с 12 - недельного возраста. Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
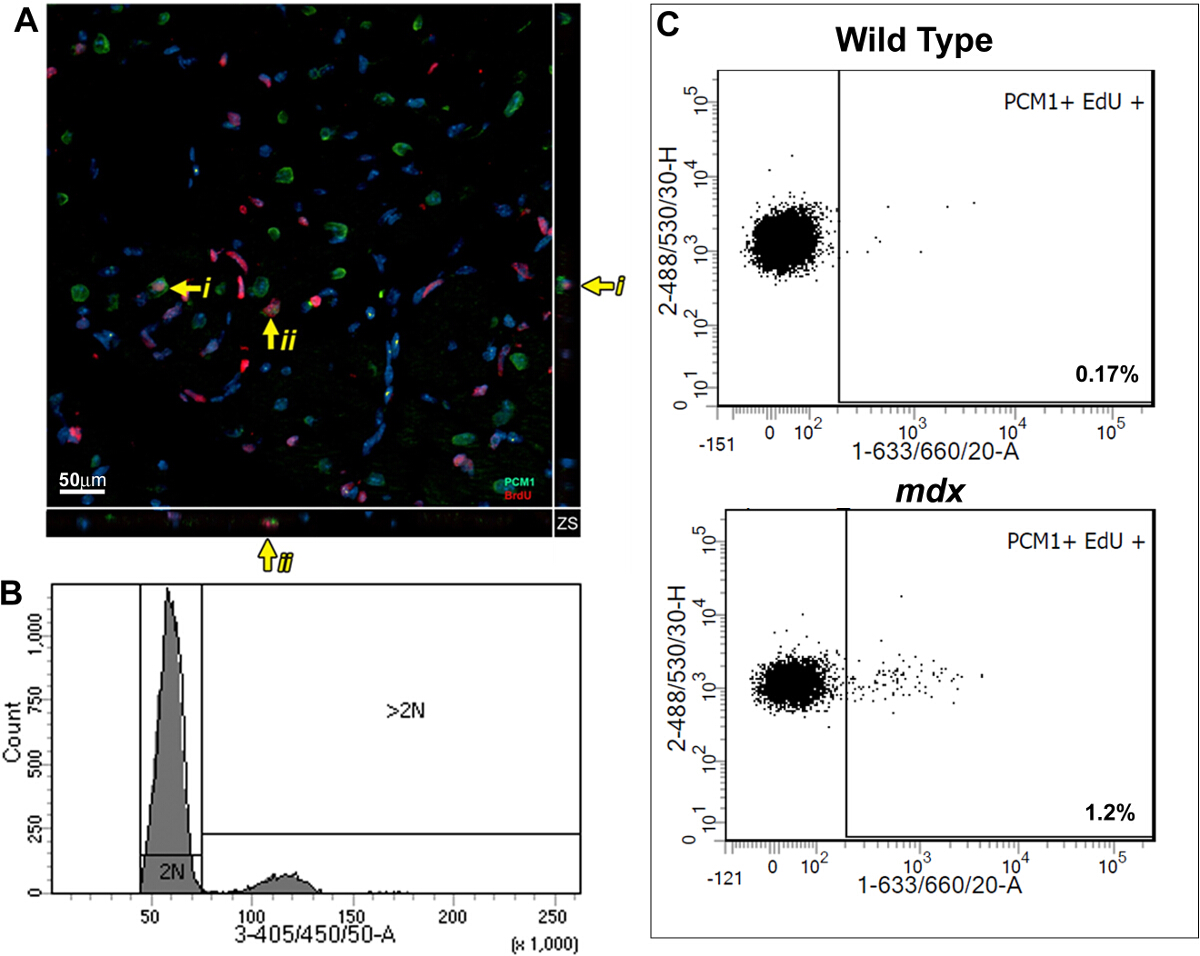{kind=link}
Дополнительное Рисунок 1:. ИКМ-1 является специфическим маркером для ядер кардиомиоцитов использованием протокола , описанного ядра были выделены из двойной трансгенной мыши , полученного путем скрещивания ТНТ-Cre мышей 14 с РОСА нТл / Ng CRE чувствительного репортерного линии 16. В результате активации мыши Cre под контролем тропонина Т (TNT) промотора приводит к необратимому экспрессии GFP с локализуется в ядрах. Ядра помечены с контролем изотипа (МОС) демонстрирует способность идентифицировать GFP выражения кардиомиоцитов ядер населения (488/530/30-А). Маркировку ядер с анти-PCM1 (вторичным антителом, обнаруживаемых 633-660 / 20-А) демонстрирует корреляцию экспрессии PCM1 с GFP, выражающей cardiomyocyт.е ядер населения. В этом примере 98,8% ядер кардиомиоцитов, определенных промоторной активности TNT, помечены PCM1 антитела. Пожалуйста , нажмите здесь , чтобы загрузить Дополнительный рисунок 1.
Дополнительный Рис . 2: Проблемы количественной оценки обновление кардиомиоцитов иммуногистологических методов (A) 2D изображения показаны два PCM-1 , выражающие ядер кардиомиоцитов (зеленый) , которые по всей видимости, также помечены для Эду (красный). Желтые стрелки указывают на то, что, как представляется, кардиомиоциты, которые включены EDU. Это связано с очевидной совместной локализации экспрессии PCM1 и маркировки EDU, когда визуализируется с помощью 2-х изображений объемную. (B) 3-мерной проекции этого изображения идентифицирует Edu меченых клеток не являются кардиомиоциты , но не являются кардиомиоцитов клетки , лежащие поверх ПКM1 выражения ядер кардиомиоцитов. C57 / BL10 мышей в возрасте 12 недель , используемых для экспериментов. Пожалуйста , нажмите здесь , чтобы загрузить Дополнительный рисунок 2A. Пожалуйста , нажмите здесь , чтобы загрузить Дополнительный рисунок 2B.
| Реакционные компоненты | Количество реакций | |
| 2 | 5 | |
| PBS | 875 мкл | 2,19 мл |
| Медь защищающее | 20 мкл | 50 мкл |
| Флуоресцентный краситель пиколил азид | 5 мкл | 12.5 мкл |
| Разведенные реакциябуфера, приготовленного в 4.1 | 100 мкл | 250 мкл |
| Общий объем реакционной смеси | 1 мл | 2,5 мл |
Таблица 2: Эду коктейль ингредиентов. Рецепт для EDU маркировки коктейля требуется в разделе протокола 4 (Обнаружение Edu включения).
Обсуждение
Для того, чтобы точно определить их оборот кардиомиоцитов и анализы регенерации должны различать истинного поколения кардиомиоцитов и непроизводственного деления ДНК. Многие исследования продолжают просто игнорировать эти непродуктивные события, количественной оценки пролиферации кардиомиоцитов исключительно с помощью экспрессии циклин кинезиса и маркеров клеточного цикла. На сегодняшний день единственный метод, который позволяет точную количественную оценку оборота кардиомиоцитов, контролируя при этом для этих непроизводительных событий остается аллюзивная. В частности , по- прежнему трудно объяснить кардиомиоцитов полиплоидизации что способствует до ~ 65% репликации ДНК кардиомиоцитов 13. Поэтому, чтобы помочь в точной количественной генерации кардиомиоцитов мы разработали протокол, позволяющий надежную количественную оценку темпов нео ядер кардиомиоцитов, исключая репликацию ДНК, что приводит к увеличению плоидности. Хотя этот протокол не может дискриминации между нео-кардиомиоцитов родовции и кардиомиоцитов би-нуклеации, она может быть быстро использована и точно рассчитать верхний предел (с учетом плоидности) генерации кардиомиоцитов. Таким образом, этот протокол обеспечивает инструмент скрининга для оценки потенциальных изменений в темпах генерации кардиомиоцитов и полиплоидизации в моделях болезни или для оценки потенциальной эффективности терапии. После того, как изменения в скорости нео-кардиомиоцитов ядер поколения идентифицируются с использованием этого протокола последующие исследования могут быть использованы , чтобы установить , является ли это из - за изменений в генерации кардиомиоцитов числа кардиомиоцитов нуклеации, как было описано ранее 2,13,17,18. К ним относятся использование гистологических динамики нуклеации Количественное кардиомиоцитов в течение периода импульса или анализа срезов тканей, полученных из ОБР импульсными животных, чтобы сравнить ОБР включение в Мононуклеированные и многоядерные популяции кардиомиоцитов.
Из-за низкого уровня оборота кардиомиоцитов этойПротокол использует множественные инъекции Edu в течение 7 дней. Это также позволяет "чеканка" всех потенциальных клеточных источников генерации кардиомиоцитов и позволяет количественно оценить кумулятивный поколения кардиомиоциты ядер за этот период времени. В зависимости от исследования, этот срок может быть скорректирована с учетом прогнозируемых уровней генерации кардиомиоцитов. Для точного количественного определения ОБР включения в ядрах кардиомиоцитов, крайне важно, чтобы не существует неспецифическое мечение ядер с вторичным антителом, используемого для обнаружения ИКМ-1 реакционной способностью. Поэтому было бы целесообразно провести дополнительные эксперименты вторичные антитела титрования с целью оптимизации этого аспекта протокола, особенно, если вторичное антитело кроме того, что предложено в этот протокол будет использоваться. Протокол, описанный здесь использует PCM-1 выражение для идентификации ядер кардиомиоцитов. В то время как это установлено маркера кардиомиоцитов, альтернативные маркеры могут быть использованы для проверкиданные; они включают антитела , специфичные для сердечного тропонина Т , который был идентифицирован как частично локализован в кардиомиоцитов ядрах 1. Аналогичным образом, альтернативные ядерные белки локализованы, могут быть использованы для идентификации и количественного определения ОБР включения в других, чем у кардиомиоцитов ядер популяций. Важно, чтобы все ядер кардиомиоцитов, которые активно подвергающиеся митоз исключены из анализа, поскольку судьба этого синтеза ДНК неизвестна и может привести или к делению клеток или повышенной плоидности. PCM1 разбирается во время фазы М клеточного цикла, поэтому кардиомиоцитов подвергается митоза не будут определены выражением PCM1. Кроме того, все ядра в s- фазе клеточного цикла, должны быть исключены из последующего анализа. Это может быть достигнуто путем стробирования из всех ядер с интенсивностью DAPI выше населения 2н в том числе и с интенсивностью DAPI между 2н и 4N населения.
Несмотря на то, что все больше и большеПринято считать, что сердце обладает способностью заменить кардиомиоцитов во время нормального старения и после острой травмы, источник и степень этого потенциала остается спорным. Кроме того, несопоставимые показатели оборота кардиомиоцитов было зарегистрировано 1,7,20-22. Это может быть отчасти из - за трудностей , связанных с точной идентификации и количественной оценки нео-кардиомиоциты поколение 19. На сегодняшний день большинство исследований основывались только на использовании гистологического анализа и идентификации кардиомиоцитов посредством экспрессии цитоплазматических белков, в том числе белки саркомера, для количественной оценки оборота кардиомиоцитов и обновления 2,4,23,24. Использование этих методов для определения экспрессии маркеров пролиферации, или, как показано здесь, введение аналогов тимидина может легко привести к неправильной идентификации других типов клеток сердца как кардиомиоциты. Хотя использование 3D-визуализации конфокальной может помочь облегчить эти проблемы йESE методы являются дорогостоящими и отнимает много времени. Интересно отметить, что протокол, описанный здесь, показывает, нео-кардиомиоцитов ядра генерации происходит со скоростью 0,17% в неделю. Это согласуется с другими проточной цитометрии исследований на основе демонстрирующих еженедельные темпы оборота до 0,13% 5. Хотя заманчиво экстраполировать годовые темпы оборота на основе этих данных, как и в предыдущих исследованиях 2,5,25,26, это неуместно , поскольку темпы оборота являются динамическими во время жизни животного 13.
Этот метод имеет ряд потенциальных применений, включая оценку потенциальных терапевтических средств для улучшения регенерации кардиомиоцитов или изучения влияния болезни сердца на обороте кардиомиоцитов и ставки кардиомиоцитов полиплоидизации.
Раскрытие информации
The authors have nothing to disclose.
Благодарности
This work was supported by the British Heart Foundation, project grant PG/13/69/30454.
Материалы
| Name | Company | Catalog Number | Comments |
| 0.32 M sucrose | Sigma | 84100 | |
| 10 mM Tris-HCl (pH = 8) | Sigma | T3253 | |
| 5 mM CaCl2 | Sigma | c5086 | |
| 5 mM magnesium acetate | sigma | M-5661 | |
| 2.0 mM EDTA | Sigma | E5134 | |
| 0.5 mM EGTA | Sigma | 63779 | |
| 1 mM DTT | Sigma | D0632 | |
| 70 mM KCl | Sigma | P9541 | |
| 10 mM MgCl2 | Sigma | M8266 | |
| 1.5 mM spermine | Sigma | 85590 | |
| Isotype rabbit IgG- ChIP Grade | abcam | abc7415 | |
| Rabbit anti-PCM-1 antibody | Sigma | HPA023374 | |
| Alexa Fluor 488 F(ab')2 Fragment of Goat Anti-Rabbit IgG (H+L) Antibody | Life technologies | A-11070 | |
| cell strainers 70 μm and 100 μm | Fisher scientific | 11597522, 11517532 | |
| Glass dounce (40 ml) and pestle large clearance | Sigma | D9188-1SET | |
| EdU (5-ethynyl-2’-deoxyuridine) | Life technologies | A10044 | |
| Click-iT Plus EdU Alexa Fluor 647 Flow Cytometry Assay Kit | Life technologies | C10634 | This kit inlcudes reagents required for section, EdU reaction buffer, EdU fixative, saponin-based permeabilization solution and the reagents required for the EdU labelling cocktail. |
| CyStain DNA 2 step kit, | Sysmex Partec | 05 5005 | This kit inlcudes reagents required for DAPI labelling (DNA staining solution) |
| Probe homogeniser, e.g., TissueRuptor | Qiagen | 9001273 | |
| TissueRuptor Disposable Probes | Qiagen | 990890 | |
| ultracentrifuge | Sorvall | ||
| Facscanto II | BD Biosciences | ||
| Ultracentrifuge Tube, Thinwall, Polypropylene. 38.5 ml, 25 x 89 mm | Beckman Coulter | 326823 | |
| Bovine serum albumin | Sigma | A2153 |
Ссылки
- Bergmann, O., et al. Evidence for cardiomyocyte renewal in humans. Science. 324, 98-102 (2009).
- Senyo, S. E., et al. Mammalian heart renewal by pre-existing cardiomyocytes. Nature. 493, 433-436 (2013).
- Soonpaa, M. H., Rubart, M., Field, L. J. Challenges measuring cardiomyocyte renewal. Biochim Biophys Acta. 1833, 799-803 (2013).
- Loffredo, F. S., Steinhauser, M. L., Gannon, J., Lee, R. T. Bone marrow-derived cell therapy stimulates endogenous cardiomyocyte progenitors and promotes cardiac repair. Cell Stem Cell. 8, 389-398 (2011).
- Malliaras, K., et al. Cardiomyocyte proliferation and progenitor cell recruitment underlie therapeutic regeneration after myocardial infarction in the adult mouse heart. EMBO Mol Med. 5, 191-209 (2013).
- Hsieh, P. C., et al. Evidence from a genetic fate-mapping study that stem cells refresh adult mammalian cardiomyocytes after injury. Nat Med. 13, 970-974 (2007).
- Bergmann, O., et al. Identification of cardiomyocyte nuclei and assessment of ploidy for the analysis of cell turnover. Exp Cell Res. 317, 188-194 (2011).
- Herget, G. W., Neuburger, M., Plagwitz, R., Adler, C. P. DNA content, ploidy level and number of nuclei in the human heart after myocardial infarction. Cardiovasc Res. 36, 45-51 (1997).
- Scholzen, T., Gerdes, J. The Ki-67 protein: from the known and the unknown. J Cell Physiol. 182, 311-322 (2000).
- Carmena, M., Earnshaw, W. C. The cellular geography of aurora kinases. Nat Rev Mol Cell Biol. 4, 842-854 (2003).
- Bergmann, O., Jovinge, S. Isolation of Cardiomyocyte Nuclei from Post-mortem Tissue. J. Vis. Exp. , e4205 (2012).
- Gilsbach, R., et al. Dynamic DNA methylation orchestrates cardiomyocyte development, maturation and disease. Nat Commun. 5, 5288 (2014).
- Richardson, G., Laval, S., Owens, W. A. Cardiomyocyte regeneration in the mdx mouse model of non-ischemic cardiomyopathy. Stem Cells Dev. , (2015).
- Jiao, K., et al. An essential role of Bmp4 in the atrioventricular septation of the mouse heart. Genes Dev. 17, 2362-2367 (2003).
- Bergmann, O., et al. Cardiomyocyte renewal in humans. Circ Res. 110, e17-e18 (2012).
- Prigge, J. R., et al. Nuclear double-fluorescent reporter for in vivo and ex vivo analyses of biological transitions in mouse nuclei. Mamm Genome. , (2013).
- Naqvi, N., et al. A proliferative burst during preadolescence establishes the final cardiomyocyte number. Cell. 157, 795-807 (2014).
- Liu, Z., Yue, S., Chen, X., Kubin, T., Braun, T. Regulation of cardiomyocyte polyploidy and multinucleation by CyclinG1. Circ Res. 106, 1498-1506 (2010).
- Ang, K. L., et al. Limitations of conventional approaches to identify myocyte nuclei in histologic sections of the heart. Am J Physiol Cell Physiol. 298, C1603-C1609 (2010).
- Kajstura, J., et al. Myocyte turnover in the aging human heart. Circ Res. 107, 1374-1386 (2010).
- Kajstura, J., et al. Cardiomyogenesis in the adult human heart. Circ Res. 107, 305-315 (2010).
- Walsh, S., Ponten, A., Fleischmann, B. K., Jovinge, S. Cardiomyocyte cell cycle control and growth estimation in vivo--an analysis based on cardiomyocyte nuclei. Cardiovasc Res. 86, 365-373 (2010).
- Anversa, P., Leri, A., Kajstura, J. Cardiac regeneration. J Am Coll Cardiol. 47, 1769-1776 (2006).
- Gonzalez-Valdes, I., et al. Bmi1 limits dilated cardiomyopathy and heart failure by inhibiting cardiac senescence. Nat Commun. 6, 6473 (2015).
- Kimura, W., et al. Hypoxia fate mapping identifies cycling cardiomyocytes in the adult heart. Nature. 523, 226-230 (2015).
- Soonpaa, M. H., Field, L. J. Assessment of cardiomyocyte DNA synthesis in normal and injured adult mouse hearts. Am J Physiol. 272, H220-H226 (1997).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены