
Bu içeriği görüntülemek için JoVE aboneliği gereklidir. Oturum açın veya ücretsiz deneme sürümünü başlatın.
Method Article
Hedeflenen
Bu Makalede
Özet
A strategy for generating mutations in histone genes at their endogenous location in Saccharomyces cerevisiae is presented.
Özet
We describe a PCR- and homologous recombination-based system for generating targeted mutations in histone genes in budding yeast cells. The resulting mutant alleles reside at their endogenous genomic sites and no exogenous DNA sequences are left in the genome following the procedure. Since in haploid yeast cells each of the four core histone proteins is encoded by two non-allelic genes with highly homologous open reading frames (ORFs), targeting mutagenesis specifically to one of two genes encoding a particular histone protein can be problematic. The strategy we describe here bypasses this problem by utilizing sequences outside, rather than within, the ORF of the target genes for the homologous recombination step. Another feature of this system is that the regions of DNA driving the homologous recombination steps can be made to be very extensive, thus increasing the likelihood of successful integration events. These features make this strategy particularly well-suited for histone gene mutagenesis, but can also be adapted for mutagenesis of other genes in the yeast genome.
Giriş
Dört çekirdekli histon proteinleri H2A, H2B, H3, H4 ve ökaryotik kromozomların sıkıştırma, organizasyon ve fonksiyonunda önemli rol oynarlar. Bu histonlar her iki set sonuçta nükleozom 1 oluşumu ile sonuçlanır, histon oktameri, kendi etrafında DNA ~ 147 baz çifti sarma yönlendiren bir molekül makara oluşturur. Nükleozomlar bu tür kromozom üzerinde gen transkripsiyonu düzenlenmesi ve ökromatin oluşumu ve heterokromatin olarak kromozom tabanlı süreçleri, çeşitli aktif katılımcılar vardır ve gibi, son birkaç on yıl boyunca yoğun bir araştırmanın odak noktası olmuştur. Bu mekanizmalar histon kalıntılarının sonrası modifikasyon, ATP-bağımlı nükleozom remodeling ve ATP-bağımsız nükleozom yeniden düzenlenmesi dahil - bir takım mekanizmalar hangi nükleozom belirli işlemlerin uygulanmasını kolaylaştırabilir yollarla manipüle edilebilir tarif edilmiştirve montaj / demontaj 2, 3.
Tomurcuklanan maya Saccharomyces cerevisiae ökaryotlarda histon işlevinin anlaşılması için özellikle güçlü bir model organizmadır. Bu büyük ölçüde etki ökaryotlarının boyunca histon proteinlerinin evrimsel korunması yüksek derecede ve genetik ve biyokimyasal deneysel çeşitli maya amenability isnat edilebilir 4 yaklaşır. maya Ters genetik yaklaşımlar yaygın kromatin biyolojinin çeşitli yönlerine ilişkin belirli histon mutasyonların etkilerini incelemek için kullanılır olmuştur. deneyler bu tür için anormal hücre içi (nedeniyle hücrelerde plazmidlerin sayıları değişen) histon proteinleri seviyeleri ve yol açabilir bağımsız plazmidlerden ifade mutant histonlar kendi ana genomik lokusları eksprese edildiği hücrelerin kullanımı genellikle tercih edilir kromatin tr eş zamanlı değişikliksonuçta sonuçların yorumunu karıştırabilir vironments.
Burada, genomu içinde kalan eksojen DNA sekansları olmadan arzu edilen mutasyon (lar) ın üretiminde bir klonlama aşaması ve sonuç gerektirmez kendi ana genomik konumlarda histon genlerinin hedeflenen mutagenezi için izin veren bir PCR bazlı teknik açıklar. Bu teknik, maya etkili homolog rekombinasyon sistemi yararlanır ve diğer gruplar tarafından geliştirilen diğer benzer teknikler ile ortak birçok özelliğe sahiptir - özellikle Delitto Perfetto, alana özgü genomik (SSG) mutajenez ve klonlama içermeyen PCR bazlı alel yedek yöntemleri 5, 6, 7. Bununla birlikte, tarif tekniği özellikle histon gen mutagenezi için çok uygundur hale getiren bir yönü vardır. haploid maya hücreleri, dört çekirdek histon her iki olmayan a göre kodlanmıştırllelic ve yüksek ölçüde homolog genlerin: Örneğin, histon H3 HHT1 ve HHT2 genleri tarafından kodlanan ve iki genin açık okuma çerçeveleri (ORF'ler), sırayla% 90 üzerinde özdeştir. homoloji Bu yüksek derecede spesifik mutagenez için iki histon-kodlayan genlerin biri hedef için tasarlanmış deneyler karmaşık hale getirebilir. Yukarıda bahsedilen yöntemler genellikle homolog rekombinasyon sürücü hedef genin ORF içinde en azından bazı dizilerin kullanılmasını gerektirir ise, burada tarif tekniği için (daha az sekans benzerliği paylaşan) histon gen ORF eteklenen dizileri kullanır rekombinasyon adımı ve böylece arzu edilen mahaline mutagenez başarılı hedefleme olasılığını arttırır. Ayrıca, rekombinasyonu sürücü homolog bölgeleri daha verimli hedeflenen homolog rekombinasyon katkıda çok geniş olabilir.
Protokol
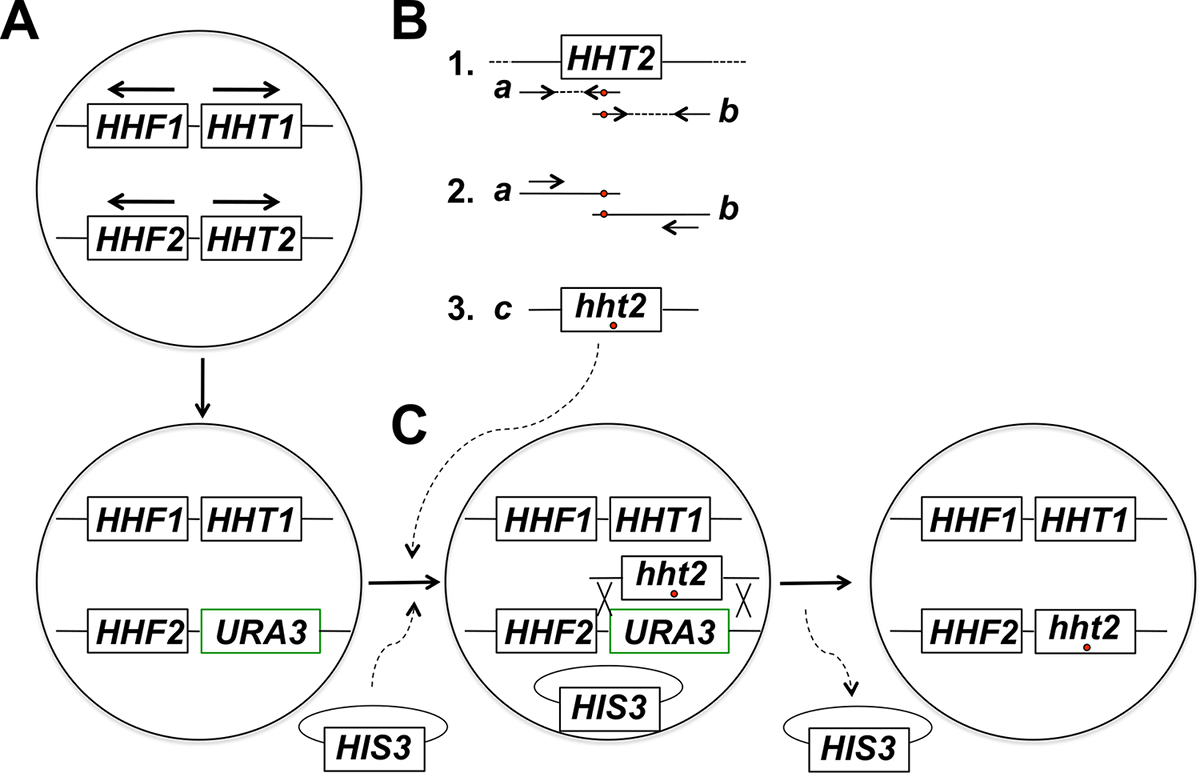
Not: in situ histon geni mutagenez hedef için deneysel strateji (Şekil 1 'de özetlenmektedir) çeşitli adımlar içerir. Bu adımlar şunlardır: URA3 geni hedef histon geninin (1) değiştirilmesi, (2) üretimi ve arzu edilen mutasyon (lar) yataklık primerler kullanılarak, hedef histon geninin iki kısmen üst üste parçalarının karşılık gelen PCR ürünlerinin saflaştırılması (3 ) kısmen örtüşen parçalarının PCR erimesi işaretleyici ile plazmit üzerinde (4) tam boyut PCR ürünlerinin ve omurga plasmid eş-dönüşüm ve seçim, (5) Resim 5-FOA dayanıklı, entegrasyon için tam boyut PCR ürünleri elde etmek için transformantlar, (6) 5-FOA-dirençli koloni saflaştırılması ve omurga plasmid kaybı, ve (7) Moleküler mutant alelinin uygun entegrasyonu için tahlil analiz eder.

Şekil 1: Budding Maya Histon Genlerin yerinde Mutajenesisin Hedeflenen Stratejisi Genel Bakış. Bu örnekte, hedeflenen gen HHT2, ancak herhangi bir diğer temel histon geni de bu strateji kullanılarak mutajenize edilebilir. Şekilde görüldüğü gibi (A) 'Haploid maya hücreleri düzenlenmiş iki histon H3 kodlayan genler (HHT1 ve HHT2) ve iki histon H4 kodlayan genler (HHF1 ve HHF2) liman (HHT1 ve HHF1 genleri kromozom II HHT2 yer almaktadır ve HHF2 genleri kromozom XIV bulunmaktadır - her bir durumda, oklar), transkripsiyon yönünde işaret etmektedir. Yöntemin birinci aşamasında, HHT2 geni ORF, bir hht2Δ :: URA3 suşu sebebiyet veren, URA3 geni ile değiştirilmiştir. Bölüm 1 (B), genomik bir DNA örneğinden HHT2 geninin doğal tipte bir kopyalama cins için iki PCR reaksiyonu için şablon olarak kullanılırgeninin iki kısmen üst üste parçaları te. Birinci reaksiyon için ters primer genomuna yerleştirilebilir istenen mutasyonu (ler) e karşılık gelir (kırmızı bir daire ile gösterilir), bir veya daha fazla uyumsuz olan nükleotidler içermektedir. İkinci reaksiyon için ileri primer (aynı zamanda kırmızı daire ile gösterilen) bir ters tamamlayıcı konfigürasyonda eşdeğer uyumsuzluk vardır. Iki PCR bölümü 1 'de üretilen ürün (ürün A ve B) daha sonra, füzyon PCR için şablon şekilde ürün A ve B tavlama Bu tam boy PCR oluşmasına neden Bölüm 2 de gösterilen iki primer kullanılarak olarak kullanılan istenen mutasyon (ler) barındıran ürünleri (bölüm 3 ürün c). (C) hht2Δ :: URA3 soyu daha sonra eş-dönüştürülmüş yoksun ortam (tam boy PCR ürünlerinin ve omurga plasmid (a HIS3- bu örnekte plazmidi işaretli) ile ve hücreler, plasmid varlığı için seçilmektedir hBu örnekte istidine). Transformantlar daha sonra 5-FOA direnci için elenir - dirençli hücrelerin gösterildiği gibi, URA3 geni PCR ürünü ve eksizyon üyelik için homolog bir rekombinasyon olayı geçirmiş için adaydırlar. mitotik hücre bölünmesi sureti ile omurga plazmitinin kaybetmesine istenen son histon mutant yol açar. Çoğunlukla kendiliğinden URA3 mutasyonlar ulaşan hücreleri tanımlayan 5-FOA plakalar doğrudan seçim ile karşılaştırıldığında doğru entegrasyon olaylarının belirlenmesi çok daha yüksek bir frekansta 5-FOA direnci sonuçları ve akabinde omurga plazmitinin bu seçimi bulduk. (Bu rakam referans 14 modifiye edilmiştir). Bu rakamın büyük halini görmek için lütfen buraya tıklayınız.
{kind=link}
URA3 ile Hedef Histon Gene 1. YedekGen
- URA3 geni 8, 9 hedef histon geninin ORF yerine standart PCR dolayımlı tek-aşamalı bir gen bozulması gerçekleştirin.
NOT: Bu mutasyon, böylece URA3 mahalinde 8 içine PCR ürününün entegrasyonu kaçınarak, tüm endojen URA3 ORF kaldırır gibi ura3Δ0 taşıyan maya hücrelerinin kullanılması tavsiye edilir. S. cerevisiae'de fonksiyonel ama S. Cerevisiae URA3 geni ile sadece kısmi dizi benzerliğine sahip olan Alternatif olarak, K lactis URA3 geni bir URA3 arka histon değiştirme üretilmesi için etkin olarak kullanılabilir. türü de transformasyon deneyinde (Bu protokolün adım 4) omurga plazmidin seçilmesine izin verecek en az bir bileşik için oksotropik olmalıdır. Bu adım, bir hedef histon geneΔ halinde :: URA3 gerekli değildirsuşu zaten mevcuttur.
2. Üretim ve istenen mutasyonu barındıran primerler kullanılarak hedef Histon geninin iki kısmen üst üste fragmanlarına karşılık gelen PCR ürünlerinin saflaştırılması (lar)
- Hedef histon geninin iki kısmen üst üste parçalarının karşılık gelen PCR ürünleri.
- şu şekilde iki PCR reaksiyonları hazırlanması:
- Isı ayarlı 1 ul DNA şablonu, 5 μl10 uM ileri primer, 5 μl10 uM ters primer, 0.5 ul (1.25 U): Aşağıdaki reaksiyon ayarlamak gen (Şekil 1B ürünü a), ilk yarısında tekabül eden PCR ürünleri üretmek için DNA polimeraz, 10 ul 5x DNA polimeraz tamponu, 5 ul dNTP karışımı (her biri 2 mm) ve 23.5 ul dH 2 O
Not: Şablon DNA, hedef histon geni için bir gerilme vahşi tip türetilen genomik DNA olabilir standart pro kullanılarak izole10 prosedürlerinin. DNA konsantrasyonu ve genomik preparasyonlar safsızlıkların seviyesi farklılaşmaların hesaba katılması için, bu seyreltilmemiş DNA veya genomik preparatlar değişik seyreltiler kullanılarak reaksiyonları optimize etmek için tavsiye edilir (örneğin, 1:10 ve 1: 100). ileri primer, hedef genin üst akışında bir bölgeye tavlama gerekir. Ters primer (örnekler için Şekil 1B-1 ve Temsilcisi Sonuçlar bölümüne bakınız) uzunluğunda ~ olması, ORF içinde 40 nükleotidleri tavlama ve bunun ortasında bir yerde istenen mutasyonu (ler) içermelidir. yüksek doğrulukta bir DNA polimeraz kullanılması PCR ürünlerinin sentezi sırasında istenmeyen mutasyonlar oranlarını azaltmak için tavsiye edilir. - 2.1.1.1 fakat farklı primerler ile belirtildiği gibi gen (Şekil 1B ürünü b), ikinci yarısına tekabül eden PCR ürünleri üretmek için, bir tepki ayarlayın.
NOT: ileri primer uzunluğunda ~ olması, ORF içinde 40 nükleotidleri tavlama, ve bir yerde onun ortasında istenen mutasyonu (ler) içermelidir. Bu primer mutasyon (lar) ının 2.1.1.1 ters primer mutasyon (lar) ın ters komplemanı olduğuna dikkat edin. Ters primer, hedef genin alt bölgeye tavlanması gerekmektedir (örnekler için Şekil 1B-1 ve Örnek Sonuçlar bölümüne bakınız).
- Isı ayarlı 1 ul DNA şablonu, 5 μl10 uM ileri primer, 5 μl10 uM ters primer, 0.5 ul (1.25 U): Aşağıdaki reaksiyon ayarlamak gen (Şekil 1B ürünü a), ilk yarısında tekabül eden PCR ürünleri üretmek için DNA polimeraz, 10 ul 5x DNA polimeraz tamponu, 5 ul dNTP karışımı (her biri 2 mm) ve 23.5 ul dH 2 O
- Aşağıdaki ayarlarla bir PCR reaksiyonları yerleştirin: 94 ˚C 30 saniye; Aşağıdaki ayarlardan 30 döngü: 98 ˚C 10 sn, 60 ° 5 sn, 72 ˚C 1.5 dakika; ve 72 ° C'de 10 dakika.
PCR parametrelerinin optimizasyonu spesifik primer setleri için gerekli olabilir ve histon geni hedef Not:.
- şu şekilde iki PCR reaksiyonları hazırlanması:
- 89 mM Tris baz, 89 mM borik asit, 2.5 mM EDTA (TBE) tampon maddesi içinde bir% 0.9 düşük erime noktalı agaroz jeli üzerinde PCR reaksiyonu ürünü 50 ul - 20 çalıştırın.
- PCR pr içeren Cut agaroz jel bölümlerive temiz bir neşter veya jilet kullanarak jelden oducts 1.5 ml mikrosantrifüj tüpüne her aktarın. Kullanıma hazır olana kadar -20 ° C'de, PCR ürünlerini içeren agaroz bölümleri saklayın.
İki Parçalı Çakışan Fragments 3. Fusion PCR Entegrasyon Tam Boyut PCR Ürünleri Elde
- PCR reaksiyonları için şablon hazırlanması
- 5 dakika boyunca 65 ° C'de bir ısıtma bloğu kümesindeki mikrosantrifüj tüpleri yerleştirerek adım 2.3 agaroz jel bölümleri eriyik (ya da tam olarak eriyene kadar). Vorteks tüpler her 1-2 dk eritme sürecini kolaylaştırmak için.
- Her bir numuneden erimiş agaroz bir miktar aktarın (örneğin 50 ul, her biri 100 ul toplam) tek bir mikrosantrfuj tübüne ve vorteks ile karıştırın. Füzyon PCR reaksiyonlarında şablon olarak kullanın. Kullanıma hazır olana kadar -20 ° C'de tüp yerleştirin.
- Tam boy PCR ürününün büyük miktarda yükseltmek (ürün c) Şekil 1B
- 2 ul hedef DNA, 10 ul 10 uM ileri primer, 10 ul 10 uM ters primeri, 1 ul (2.5 U) termostabil DNA polimeraz, 20 ul 5x DNA polimeraz tamponu, 10 ul altı PCR reaksiyonları, aşağıdaki bileşenlerin her ayarlama dNTP karışımı (2 mM her biri) ve 47 ul dH 2 O
Not: reaksiyonların sayısı, PCR etkinliğine bağlı değiştirilebilir. Erimiş vorteks ıle karıştırılmıştır, ve PCR reaksiyon karışımına Son eklenen kadar model DNA (3.1.2) 65 ° C'ye ısıtılmalıdır. Eklendikten sonra çözümü pipetleme ve birkaç kez yavaşça ama iyice karıştırın. Farklı örneklerde DNA konsantrasyonunun farklılıkları hesaba katmak, ilk sulandırılmamış şablon veya şablonun farklı seyreltiler birini kullanarak tepkilerini optimize etmek için tavsiye edilir (örneğin, 1:10 ve 1: 100). hastalıklı olarak hedef genin iki kısmen üst üste binen fragmanlara tavlama Kullanılan iki primerŞekil 1B-2 ustrated ve son PCR ürünleri, homolog rekombinasyon adım (örnekler için Örnek Sonuçlar bölümüne bakınız) izlerler, URA3 ORF kuşatan bölgelere iki tarafında homolog en az 40 baz çiftine sahip olacaktır şekilde tasarlanabilir. yüksek doğrulukta bir DNA polimeraz kullanılması PCR ürünlerinin sentezi sırasında istenmeyen mutasyonlar oranlarını azaltmak için tavsiye edilir. - Aşağıdaki ayarlarla bir PCR tüpleri yerleştirin: 94 ˚C 30 saniye; Aşağıdaki ayarlardan 30 döngü: 98 ˚C 10 sn, 50 ˚C 15 sn, 72 ˚C 1.5 dakika; ve 72 ° C'de 10 dakika.
Not: PCR parametrelerinin optimizasyonu özel primer kümeleri ve hedef histon geni için gerekli olabilir.
- 2 ul hedef DNA, 10 ul 10 uM ileri primer, 10 ul 10 uM ters primeri, 1 ul (2.5 U) termostabil DNA polimeraz, 20 ul 5x DNA polimeraz tamponu, 10 ul altı PCR reaksiyonları, aşağıdaki bileşenlerin her ayarlama dNTP karışımı (2 mM her biri) ve 47 ul dH 2 O
Plazmid üzerinde Marker için 4. Tam Boyut PCR Ürünleri ve Omurga Plazmid Co-dönüşümü, ve Seçim
- PCR ürünlerinin konsantre
- s havuzix PCR reaksiyonları tek bir mikrosantrifüj tüp içine adım 3.2.2 den (600 ul toplam) ve vorteks karıştırın.
- mikrosantrifüj tüplerinde üç 200 ul kısma örnek bölün. 3M sodyum asetat 20 ul (pH 5.2) ve% 100 etanol içinde 550 ul ekleyerek, her bir tüp içinde DNA hızlandırabilir. İyice çözelti birleştirilir ve en az 15 dakika boyunca buz üzerine yerleştirin. 10 dakika boyunca ~ 14,000 x g'de santrifüj ile DNA'nın toplanması 200,% 70 etanol ul, ve kuru hava ile pelet yıkayın.
- DH 2 O 25 ul her bir DNA pelletini, ve (75 ul toplam) tek bir tüp içine havuz.
- Maya co-dönüşüm
- Maya ekstraktı Peptonlu Dekstroz (YPD) 11 orta sıvı bölüm 1'de oluşturulan gerginlik gecede kültür 10 ml hazırlayın.
- Sonraki sabah, doymuş bir gece boyunca kültür 8 ml YPD sıvı ortam 400 ml inoküle ve çalkalanarak inkübe4 30 ° C'de - 5 saat hücrelerin büyüme logaritmik fazı girmesine izin vermek için.
- 10 dakika boyunca ~ 3.220 x g santrifüj ile hücreler toplama sıvı ortam atmak ve 10 mM Tris-HCl (pH 8.0), 1 mM EDTA, 1 hacim hücreleri tekrar süspansiyon 0.1 M lityum asetat çözeltisi (TE / LiAc) .
- 10 dakika boyunca ~ 3,220 x g'de santrifüj ile hücreler toplamak ve TE / LiAc atın.
- 1 ml TE / LiAc hücrelerinin yeniden süspanse.
- mikrosantrifüj tüpü içinde aşağıdaki reaksiyon kokteyli kurun: hücre 800 ul aşama 4.2.5 arasında, kaynatılmış, 10 mg / ml som balığı spermi DNA'sı 40 uL, omurga plasmid DNA 12.5 ug toplam ve konsantre PCR ürünü 75 ul adım 4.1.3 den.
Not: somon sperm DNA 5 dakika için kaynatılır ve reaksiyonunda kullanılmadan önce en az 5 dakika için buz üzerine yerleştirilmiş olmalıdır. ilave omurga plasmid DNA toplam hacmi en az (~ 80 ul ya da daha az) tutulmalıdır. Bir omurga plasm bir örnek için Temsilcisi Sonuçlar bölümüne bakınİD. - eşit sekiz mikrosantrifüj tüpler (- 8 Tüpler 1) içine kokteyl iyice tüp ve kısım karıştırın.
- Aşağıdaki iki kontrol dönüşüm reaksiyon tüpleri ayarlayın:
- Boru 9 (Resim PCR ürünü kontrol): Aşama 4.2.5 den 100 ul hücre, kaynatılmış, 10 mg / ml som balığı spermi DNA'sı 5 ul;, 1.56 ug toplam (5 dakika için kaynatılır adım 4.2.6 nota bakınız) omurga plasmid DNA ve hiçbir PCR ürününün ilave edildi.
- Tüp 10 (Resim DNA kontrol): Aşama 4.2.5, kaynatılmış, 10 mg / ml somon spermi DNA'sı (aşama 4.2.6 Not) 5 uL hücreler 100 ul DNA eklenmiş omurga plasmid ve hiç PCR ürünü ilave edildi.
- yukarı pipetleme ve birkaç kez yavaşça ama iyice her iki tüpü karıştırın.
- 30 dakika boyunca 30 ° C'de on inkübe edin.
- Her bir tüpe, TE / LiAc içinde% 40 polietilen glikol, 1.2 ml (PEG 3350) ekleyin. Çözelti homojen hale gelene kadar iyice P-1000 pipeti kullanarak karıştırın.
- 30 on tüpler inkübe30 dakika ° C. Yavaşça aşağı pipetleme çözeltisi karışımı ve ardından 15 dakika boyunca 42 ° C'de inkübe edin.
- 30 sn için ~ 14,000 xg bir mikrosantrifüj tüpleri iplik hücreleri toplamak. Sıvı atın ve steril dH 2 O 1 ml hücreleri tekrar süspansiyon
- 30 sn için ~ 14,000 xg bir mikrosantrifüj tüpleri iplik hücreleri toplamak. Sıvı atın ve steril dH 2 O 500 ul hücreleri tekrar süspansiyon
- Havuz tüpleri 1-8 birlikte (toplam 4 ml hacim) ve aşağı yukarı pipetleme ile iyice karıştırın.
- Levha omurgası plazmidin seçilmesine yirmi tam az bırakma ortamı plakaları 11 (levhalar 1-20) her biri için yukarıdaki karışım 200 ul.
- Levha boru 9'daki karışıma 200 ul ve kendi seçimi plaka üzerinde tüp 10, her karışımın 200 ul (plakaları, sırasıyla 21 ve 22).
- 5 gün 3 - 30 ° C 'de 22 inkübe edinplazmid transformantlar seçin.
- inkübasyon 5 gün - 3 sonra dönüşüm plakaları kontrol edin. Yaklaşık 5.000 koloni plakalar 1-21 (bir örnek için Temsilcisi Sonuçlar bakınız) görünür olmalı ve hiçbir kolonileri 22 mevcut olmalıdır.
5-FOA dirençli transformantlar 5. Ekran
- Plakalardan hücreleri aktarın 1-20 (ve dönüşüm, kontrol olarak plaka 21) 5-fluoroorotic asit (5-FOA) plakaları 11 ile çoğaltma kaplama entegrasyonu sonucu URA3 geninin kaybı taranması amacıyla 12 istenilen yerde PCR ürünleri.
- plaka kapağını çıkarın ve steril bir kadife koloniler içeren plaka basın. kadife plaka basarak 5-FOA plakasına kadife hücreleri aktarın. 2 gün boyunca 30 ° C'de inkübe edin.
- 2 günlük inkübasyondan sonra, dikkatli bir şekilde gr 5-FOA plakaları kontrolowth.
NOT: Bir aday entegrasyon olay 5-FOA plaka üzerinde küçük bir asimetrik "ezilmiş" koloni tarafından temsil edilecek - tersine, 5-FOA plakaları üzerinde büyüyen küçük papillalar kolonilerin büyüme sırasında ortaya çıkan spontan URA3 mutasyonların olası temsilcisi dönüşüm plakaları ve istenen uyum olayı (bazı örneklerde bu noktada daha hazırlanması için Örnek Sonuçlar bölümünde ve için bakınız Şekil 3) 'i temsil etmek ve böylece olası değildir.
5-FOA dayanıklı Kolonileri ve Omurga Plazmid Kaybının 6. saflaştırılması
- Steril kürdan kullanarak, YPD plakalar üzerine tek koloniler için adım 5.2 ve çizgi açıklanan 5-FOA plakaları aday koloniler seçin. 30 ° C 'de 3 gün - 2 inkübe edin.
- Kuluçka, kopya ardından - plaka taze YPD plaka her YPD arıtma plaka, bir bırakma plakası kaybı kontrol etmek için urasil eksikURA3 geni, ve bir ikinci çıkış elemanı omurga plazmitinin olup olmadığını belirlemek için izlemek için. 30 ° C'de 2 gün 1 - inkübe edin.
- Inkübasyondan sonra, YPD plaka üzerinde büyüyen ama ya bırakma plaka üzerinde büyüyen değil her aday örnek bir koloni tanımlamak (örneğin bir koloni rekombinasyon olayı ile URA3 genini kaybetmiş beklenen ve mitoz sırasında omurga plazmid kaybolur hücre bölünmesi). Taze YPD plakalar üzerinde bu tür koloniler Restreak. Bu koloniler entegrasyon adayları ve 7. adımda daha da analiz edilecektir.
Mutant alelinin Uygun Bütünleşme tahlile 7. Moleküler Analiz
- Standart prosedürler 10 kullanılarak aday örneklerinden genomik DNA izole edin.
- Hedef sitesini kapsayan genomik bölge yükseltin.
- 0.5 ul DNA şablonu, 5 ul: her bir örnek için, aşağıdaki PCR reaksiyon ayarlama10 uM ileri primer, 5 ul 10 uM ters primer, 0.5 ul (2.5 birim) Taq DNA polimeraz, 5 ul 10x Taq DNA polimeraz tamponu, 5 ul dNTP karışımı (her biri 2 mm) ve 29 ul DH 2 O
Not: Şablon DNA, aday örneklerinden türetilmiş genomik DNA'dır. Bir orijinal histon geneΔ türetilen genomik DNA :: şablon olarak URA3 suşu kullanarak, şablon olarak yabani tip bir histon soyundan başka bir kullanılarak genomik DNA: iki kontrol reaksiyonları dahil edilmesi önerilmektedir. DNA konsantrasyonu ve genomik preparasyonlar safsızlıkların seviyesi farklılaşmaların hesaba katılması için, bu seyreltilmemiş DNA veya genomik preparatlar değişik seyreltiler kullanılarak reaksiyonları optimize etmek için tavsiye edilir (örneğin, 1:10 ve 1: 100). Emin olmak için önemli olduğu varsayımsal entegre PCR ürünü kapsadığı bölgenin dışından DNA dizilerine, bu primerler, tavlama - bu şekilde, bu r PCR ürünlerinin boyutunureaksiyonlarda doğru genomik konumda ürünlerin entegrasyonu için bir tanı aracı olarak kullanılabilir (bir örnek için Temsilcisi Sonuçlar bakınız). - aşağıdaki ayarlarla bir PCR reaksiyonları yerleştirin: 94 ° C 3 dakika; Aşağıdaki ayarlardan 30 döngü: 94 ° C'de 45 saniye, 50 ° C, 45 saniye, 72 ° C'de 2 dakika; ve 72 ° C'de 10 dakika.
Not: PCR parametrelerinin optimizasyonu özel primer kümeleri ve hedef histon geni için gerekli olabilir.
- 0.5 ul DNA şablonu, 5 ul: her bir örnek için, aşağıdaki PCR reaksiyon ayarlama10 uM ileri primer, 5 ul 10 uM ters primer, 0.5 ul (2.5 birim) Taq DNA polimeraz, 5 ul 10x Taq DNA polimeraz tamponu, 5 ul dNTP karışımı (her biri 2 mm) ve 29 ul DH 2 O
- PCR ürünlerinin işlenmesi
- % 0.8 TBE agaroz jeli üzerinde her bir reaksiyonun 20 ul çalıştırın.
- URA3 geni başarılı varsayımsal mutasyona uğramış histon geni tarafından değiştirilmiştir olup olmadığını belirlemek için bir referans olarak, DNA standartları kullanılarak PCR ürünlerinin boyutunu değerlendirmek (örnek için Örnek sonuçlar).
NOT: Bazı durumlarda, arzu edilen mutasyon (lar) histon genler girmiş ya oluşturabilir ya da oturup bir kısıtlama yoke. Bu durumda ise, doğru entegrasyon gösterge büyüklükte PCR ürünlerinde arzu edilen mutasyonun varlığı jel elektroforez analizi ile takip karşılık gelen sınırlama enzimi ile sindirme ürünleri tabi tutulması ile tespit edilebilir (örnek için Örnek Sonuçlar bakınız) . - DNA dizilimi doğru entegrasyon gösterge boyutu Konusu PCR ürünleri istenilen mutasyon (lar) varlığını doğrulamak için ve hiçbir ek mutasyonlar genomu içine girmiştir emin olmak için.
Sonuçlar
In situ mutagenez stratejisi hedef arasında temsili bir örnek olarak, bir glutamik asit (mutan H3-R53E) bir arjininden pozisyonda 53 bir ikame barındıran bir histon H3 mutan proteini ifade eden bir hht2 alel üretilmesini tarif eder.
Biz HHT2 bütün ORF URA3 geni (protokol aşamasını 1 e bakınız) ile ikame edildiği bir soy elde. Bu soy, yAAD156 da hücrelerin histidin içi...
Tartışmalar
Haploid S. cerevisiae hücreleri dört temel histon proteinlerinin her biri için kod özel mutagenez için iki genin biri hedeflemek isteyen araştırmacılar için bir meydan okuma temsil edebilir olmayan iki alel genler arasındaki dizi homoloji yüksek düzeyde. Daha önce sık kullanan, Delitto Perfetto, alana özgü genomik (SSG) mutajenez ve klonlama içermeyen PCR bazlı alel değiştirme yöntemleri 5, 6, 7...
Açıklamalar
The authors declare that they have no competing financial interests.
Teşekkürler
We thank Reine Protacio for helpful comments during the preparation of this manuscript. We express our gratitude to the National Science Foundation (grants nos. 1243680 and 1613754) and the Hendrix College Odyssey Program for funding support.
Malzemeler
| Name | Company | Catalog Number | Comments |
| 1 kb DNA Ladder (DNA standards) | New England BioLabs | N3232L | |
| Agarose | Sigma | A5093-100G | |
| Boric Acid | Sigma | B0394-500G | |
| dNTP mix (10 mM each) | ThermoFisher Scientific | R0192 | |
| EDTA solution (0.5 M, pH 8.0) | AmericanBio | AB00502-01000 | |
| Ethanol (200 Proof) | Fisher Scientific | 16-100-824 | |
| Ethylenediaminetetraacetic acid disodium salt dihydrate (EDTA) | Sigma | E4884-500G | |
| Lithium acetate dihydrate | Sigma | L6883-250G | |
| MyCycler Thermal Cycler | BioRad | 170-9703 | |
| Poly(ethylene glycol) (PEG) | Sigma | P3640-1KG | |
| PrimeSTAR HS DNA Polymerase (high fidelity DNA polymerase) and 5x buffer | Fisher Scientific | 50-443-960 | |
| Salmon sperm DNA solution | ThermoFisher Scientific | 15632-011 | |
| Sigma 7-9 (Tris base, powder form) | Sigma | T1378-1KG | |
| Sodium acetate trihydrate | Sigma | 236500-500G | |
| Supra Sieve GPG Agarose (low metling temperature agarose) | AmericanBio | AB00985-00100 | |
| Taq Polymerase and 10x Buffer | New England BioLabs | M0273X | |
| Toothpicks | Fisher Scientific | S67859 | |
| Tris-HCl (1 M, pH 8.0) | AmericanBio | AB14043-01000 | |
| a-D(+)-Glucose | Fisher Scientific | AC170080025 | for yeast media |
| Agar | Fisher Scientific | DF0140-01-0 | for yeast media |
| Peptone | Fisher Scientific | DF0118-07-2 | for YPD medium |
| Yeast Extract | Fisher Scientific | DF0127-17-9 | for YPD medium |
| 4-aminobenzoic acid | Sigma | A9878-100G | for complete minimal dropout medium |
| Adenine | Sigma | A8626-100G | for complete minimal dropout medium |
| Glycine hydrochloride | Sigma | G2879-100G | for complete minimal dropout medium |
| L-Alanine | Sigma | A7627-100G | for complete minimal dropout medium |
| L-Arginine monohydrochloride | Sigma | A5131-100G | for complete minimal dropout medium |
| L-Asparagine monohydrate | Sigma | A8381-100G | for complete minimal dropout medium |
| L-Aspartic acid sodium salt monohydrate | Sigma | A6683-100G | for complete minimal dropout medium |
| L-Cysteine hydrochloride monohydrate | Sigma | C7880-100G | for complete minimal dropout medium |
| L-Glutamic acid hydrochloride | Sigma | G2128-100G | for complete minimal dropout medium |
| L-Glutamine | Sigma | G3126-100G | for complete minimal dropout medium |
| L-Histidine monohydrochloride monohydrate | Sigma | H8125-100G | for complete minimal dropout medium |
| L-Isoleucine | Sigma | I2752-100G | for complete minimal dropout medium |
| L-Leucine | Sigma | L8000-100G | for complete minimal dropout medium |
| L-Lysine monohydrochloride | Sigma | L5626-100G | for complete minimal dropout medium |
| L-Methionine | Sigma | M9625-100G | for complete minimal dropout medium |
| L-Phenylalanine | Sigma | P2126-100G | for complete minimal dropout medium |
| L-Proline | Sigma | P0380-100G | for complete minimal dropout medium |
| L-Serine | Sigma | S4500-100G | for complete minimal dropout medium |
| L-Threonine | Sigma | T8625-100G | for complete minimal dropout medium |
| L-Tryptophan | Sigma | T0254-100G | for complete minimal dropout medium |
| L-Tyrosine | Sigma | T3754-100G | for complete minimal dropout medium |
| L-Valine | Sigma | V0500-100G | for complete minimal dropout medium |
| myo-Inositol | Sigma | I5125-100G | for complete minimal dropout medium |
| Uracil | Sigma | U0750-100G | for complete minimal dropout medium |
| Ammonium Sulfate | Fisher Scientific | A702-500 | for complete minimal dropout medium |
| Yeast Nitrogen Base | Fisher Scientific | DF0919-07-3 | for complete minimal dropout medium |
| 5-Fluoroorotic acid (5-FOA) | AmericanBio | AB04067-00005 | for 5-FOA medium |
Referanslar
- Luger, K., Mader, A. W., Richmond, R. K., Sargent, D. F., Richmond, T. J. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature. 389 (6648), 251-260 (1997).
- Campos, E. I., Reinberg, D. Histones: annotating chromatin. Annu Rev Genet. 43, 559-599 (2009).
- Rando, O. J., Winston, F. Chromatin and transcription in yeast. Genetics. 190 (2), 351-387 (2012).
- Duina, A. A., Miller, M. E., Keeney, J. B. Budding yeast for budding geneticists: a primer on the Saccharomyces cerevisiae model system. Genetics. 197 (1), 33-48 (2014).
- Storici, F., Resnick, M. A. Delitto perfetto targeted mutagenesis in yeast with oligonucleotides. Genet Eng (N Y). 25, 189-207 (2003).
- Gray, M., Kupiec, M., Honigberg, S. M. Site-specific genomic (SSG) and random domain-localized (RDL) mutagenesis in yeast. BMC Biotechnol. 4, 7 (2004).
- Erdeniz, N., Mortensen, U. H., Rothstein, R. Cloning-free PCR-based allele replacement methods. Genome Res. 7 (12), 1174-1183 (1997).
- Brachmann, C. B., et al. Designer deletion strains derived from Saccharomyces cerevisiae S288C: a useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast. 14 (2), 115-132 (1998).
- Lundblad, V., Hartzog, G., Moqtaderi, Z. Manipulation of cloned yeast DNA. Curr Protoc Mol Biol. Chapter 13, (2001).
- Hoffman, C. S. Preparation of yeast DNA. Curr Protoc Mol Biol. Chapter 13, (2001).
- Treco, D. A., Lundblad, V. Preparation of yeast media. Curr Protoc Mol Biol. Chapter 13, (2001).
- Lederberg, J., Lederberg, E. M. Replica plating and indirect selection of bacterial mutants. J Bacteriol. 63 (3), 399-406 (1952).
- Sikorski, R. S., Hieter, P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics. 122 (1), 19-27 (1989).
- Johnson, P., et al. A systematic mutational analysis of a histone H3 residue in budding yeast provides insights into chromatin dynamics. G3 (Bethesda). 5 (5), 741-749 (2015).
- DiCarlo, J. E., et al. Genome engineering in Saccharomyces cerevisiae using CRISPR-Cas systems. Nucleic Acids Res. 41 (7), 4336-4343 (2013).
- Cross, S. L., Smith, M. M. Comparison of the structure and cell cycle expression of mRNAs encoded by two histone H3-H4 loci in Saccharomyces cerevisiae. Mol Cell Biol. 8 (2), 945-954 (1988).
- Libuda, D. E., Winston, F. Amplification of histone genes by circular chromosome formation in Saccharomyces cerevisiae. Nature. 443 (7114), 1003-1007 (2006).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiDaha Fazla Makale Keşfet
This article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır