
Для просмотра этого контента требуется подписка на Jove Войдите в систему или начните бесплатную пробную версию.
Method Article
целевое
В этой статье
Резюме
A strategy for generating mutations in histone genes at their endogenous location in Saccharomyces cerevisiae is presented.
Аннотация
We describe a PCR- and homologous recombination-based system for generating targeted mutations in histone genes in budding yeast cells. The resulting mutant alleles reside at their endogenous genomic sites and no exogenous DNA sequences are left in the genome following the procedure. Since in haploid yeast cells each of the four core histone proteins is encoded by two non-allelic genes with highly homologous open reading frames (ORFs), targeting mutagenesis specifically to one of two genes encoding a particular histone protein can be problematic. The strategy we describe here bypasses this problem by utilizing sequences outside, rather than within, the ORF of the target genes for the homologous recombination step. Another feature of this system is that the regions of DNA driving the homologous recombination steps can be made to be very extensive, thus increasing the likelihood of successful integration events. These features make this strategy particularly well-suited for histone gene mutagenesis, but can also be adapted for mutagenesis of other genes in the yeast genome.
Введение
Четыре основных белков гистонов H2A, H2B, H3, H4 и играют центральную роль в уплотнению, организации и функции эукариотических хромосом. Два комплекта каждого из этих гистонов образуют гистонов октамером, молекулярный катушку , которая направляет оборачивание ~ 147 пар оснований ДНК вокруг себя, в конечном счете , приводит к образованию нуклеосом 1. Нуклеосомы являются активными участниками различных процессов хромосом на основе, например, регуляции транскрипции генов и формирования эухроматином и гетерохроматина через хромосом, и как таковые были в центре внимания интенсивных исследований в течение последних нескольких десятилетий. Ряд механизмов, были описаны, с помощью которых нуклеосомы можно манипулировать такими способами, которые могут облегчить выполнение определенных процессов - эти механизмы включают в посттрансляционной модификации гистонов остатков, АТФ-зависимой нуклеосом ремоделирование и АТФ-независимую реорганизацию нуклеосоми монтаж / демонтаж 2, 3.
Многообещающий дрожжей Saccharomyces CEREVISIAE является особенно мощным модельным организмом для понимания функции гистонов в эукариот. Это может быть в значительной степени связано с высокой степенью эволюционного сохранения гистоновых белков во всей области Eukarya и аменабельности дрожжей до разнообразных генетических и биохимических экспериментальных подходов 4. Реверс-генетические подходы у дрожжей широко используется для изучения влияния специфических мутаций гистонов по различным аспектам биологии хроматина. Для этих типов экспериментов часто предпочтительно использовать клетки, в которых мутантный гистоны, экспрессируемые из их родного геномного локуса, как экспрессия из автономных плазмид может привести к ненормальной внутриклеточных уровней белков гистонов (из-за разного количества плазмид в клетках) и сопутствующее изменение хроматина анvironments, которые могут в конечном счете, запутать интерпретацию результатов.
Здесь мы опишем метод ПЦР на основе, которая позволяет направленного мутагенеза генов гистонов на их родных геномных местах, которые не требуют стадии клонирования и приводит к генерации желаемой мутации (ов) без оставшихся последовательностей экзогенной ДНК в геноме. Этот метод использует преимущества эффективной системы гомологичной рекомбинации у дрожжей и имеет ряд общих черт с другими подобными методами , разработанными другими группами - в первую очередь на Delitto Perfetto, сайт-специфической геномной (SSG) мутагенеза и клонированию свободной ПЦР на основе аллель методы замены 5, 6, 7. Тем не менее, этот метод описан имеет аспект, который делает его особенно хорошо подходит для мутагенеза генов гистонов. В гаплоидных клетках дрожжей, каждый из четырех основных гистонов кодируется двумя не-Аllelic и высоко гомологичных генов: например, гистона Н3 , кодируемые генами HHT1 и HHT2, и открытые рамки считывания (ORFs) двух генов более 90% идентична последовательности в определенной последовательности. Такая высокая степень гомологии может осложнить эксперименты, направленные на специально направлены одной из двух гистонов генов, кодирующих для мутагенеза. Принимая во внимание вышеупомянутые методы часто требуют использования, по крайней мере некоторых последовательностей в пределах ORF гена-мишени для привода гомологичной рекомбинации, методика описана здесь использует последовательности, фланкирующие ORFs генов гистонов (в которых намного меньше гомологии последовательностей) для стадия рекомбинации, таким образом, увеличивая вероятность успешного нацеливания мутагенеза до нужного локуса. Кроме того, гомологичные регионы, что диск рекомбинации может быть очень обширным, дополнительно способствуя эффективной целевой гомологичной рекомбинации.
протокол
Примечание: Экспериментальная стратегия направлена на месте гена гистона мутагенеза включает в себя несколько этапов (показаны на рисунке 1). Эти шаги включают в себя: (1) замена гена мишени гистона с геном URA3, (2) Производство и очистка продуктов ПЦР , соответствующих двум частично перекрывающихся фрагментов гена гистона - мишени с использованием праймеров , несущих желаемую мутацию (и), (3 ) Fusion ПЦР из двух частично перекрывающихся фрагментов, чтобы получить полный размер PCR продуктов для интеграции, (4) совместное преобразование полного размера продуктов ПЦР и магистральную плазмидой, и выделение для маркера на плазмиде, (5) экран для 5-FOA-резистентный трансформанты, (6) Очистка 5-FOA-устойчивых колоний и потере основной цепи плазмиды, и (7) Молекулярные анализы для анализа для правильной интеграции мутантного аллеля.

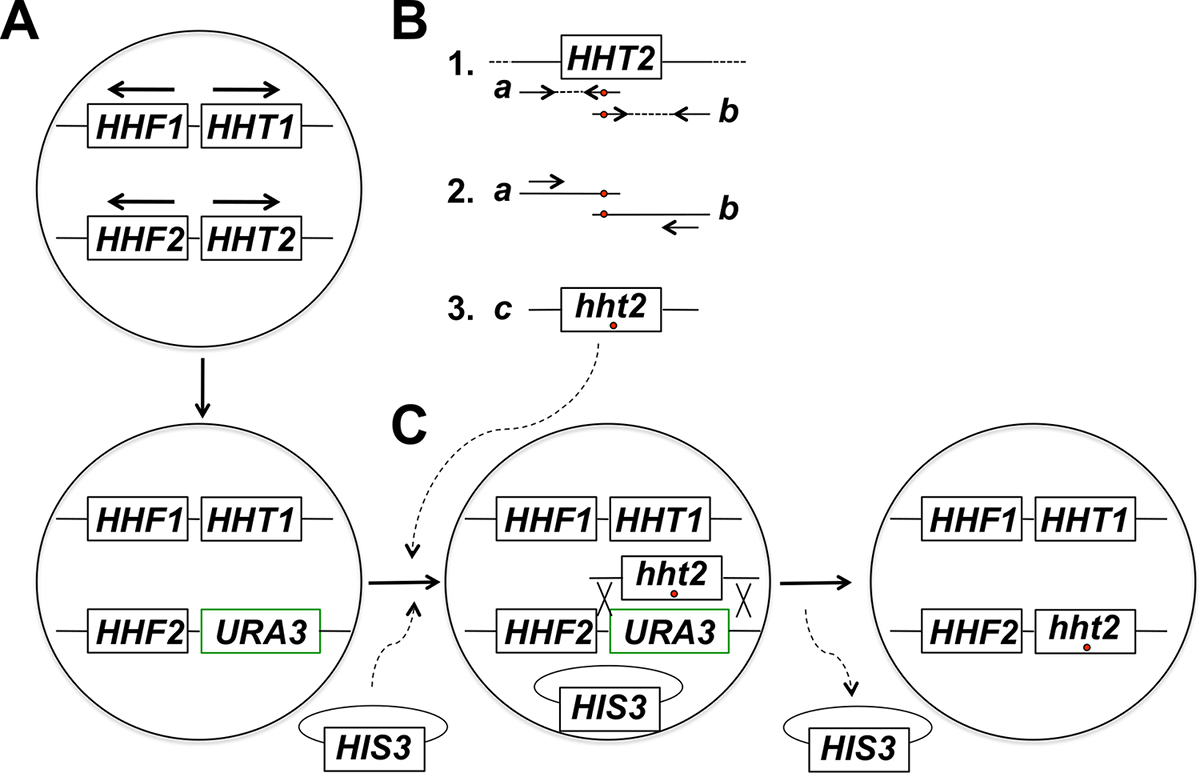
Рисунок 1: Обзор Стратегии Targeted на месте мутагенеза гистонов генов у почкующихся дрожжей. В этом примере целевой ген HHT2, но и любой другой ген корового гистона также может быть мутагенезу с помощью этой стратегии. (A) гаплоидных клетки дрожжей питают два гистона H3-кодирующие гены (HHT1 и HHT2) и два гистонов генов H4-кодировщика (HHF1 и HHF2) , расположенные , как показано на рисунке (гены HHT1 и HHF1 расположены на хромосоме II и HHT2 и гены HHF2 расположены на хромосоме XIV - в каждом случае, стрелки указывают направление транскрипции). На первом этапе процедуры ORF гена HHT2 заменен геном URA3, что приводит к деформации hht2Δ :: URA3. (В) В части 1 дикого типа копию гена HHT2 из геномной ДНК образца используют в качестве матрицы для двух ПЦР - реакций к родамт.е два частично перекрывающихся фрагментов гена. Обратный праймер для первой реакции включает в себя один или более несовпадающих нуклеотида (помеченные красным кружком), которые соответствуют желаемой мутации (ы), которые будут введены в геном. Прямой праймер для второй реакции имеет эквивалентное несоответствие в обратной дополнительной конфигурации (также обозначенной красным кружком). Два ПЦР - продукты , образующиеся в части 1 (продукции А и Б), затем используются в качестве шаблонов для слитого ПЦР с использованием двух праймеров , которые отжигаются к продуктам а и Ь в моде , показанные в части 2. Это приводит к генерации полноразмерного ПЦР продукты (продукт C в части 3) , несущие требуемую мутацию (и). (C) hht2Δ :: URA3 штамм затем совместно трансформировали полноразмерных продуктов ПЦР и позвоночника плазмиды (а HIS3- отмеченные плазмиды в данном примере), и клетки отбирают на присутствие плазмиды (на средах , не имеющих часistidine в данном примере). Трансформанты затем подвергают скринингу на 5-FOA сопротивления - резистентные клетки являются кандидатами для претерпев гомологичной рекомбинацию , ведущей к интеграции продукта ПЦР и иссечение гена URA3, как показано на рисунке. Последующейпотерей магистральную плазмиды митотического деления клеток приводит к конечной требуемой гистона мутантного штамма. Мы обнаружили , что выбор магистральную плазмиды с последующим скринингом для сопротивления приводит к 5-FOA , в гораздо более высокой частоте выявления правильных событий интеграции , по сравнению с прямого выбора на 5-FOA , скребков, которые в основном определяет клетки, которые приобрели спонтанные URA3 мутации. (Эта цифра была изменена со ссылкой 14). Пожалуйста , нажмите здесь , чтобы посмотреть увеличенную версию этой фигуры.
{kind=link}
1. Замена целевого гена гистона с URA3Ген
- Выполните стандартную ПЦР-опосредованного нарушения гена один шаг , заменяющего ORF гена гистона - мишени с геном URA3 8, 9.
Примечание: Использование дрожжевых клеток , несущих ura3Δ0 рекомендуется как эта мутация удаляет весь эндогенный URA3 ORF, избегая таким образом интеграции продукта ПЦР в URA3 локусе 8. В качестве альтернативы, ген URA3 Lactis К. могут быть эффективно использованы для генерации замены гистона в любом ura3 фоне , как это функциональный в S.cerevisiae , но имеет лишь частичную гомологию последовательности с геном URA3 S.cerevisiae , . Штамм должен также быть ауксотрофная, по крайней мере, одно соединение, которое позволит для выбора основной цепи плазмиды в эксперименте трансформации (этап 4 настоящего Протокола). Этот шаг не является необходимым , если цель гистонов geneΔ :: URA3Штамм уже доступен.
2. Формирование и очистка продуктов ПЦР, соответствующие двум частично перекрывающихся фрагментов из целевых Гистона гена с использованием праймеров, несущих желаемую мутацию (и)
- Создание продуктов PCR , соответствующие двум частично перекрывающихся фрагментов гена мишени гистонов.
- Приготовьте две реакции ПЦР следующим образом:
- Для создания продуктов ПЦР , соответствующие первой половине гена (продукт А на рисунке 1В), установить следующие реакции: 1 мкл матричной ДНК, 5 μl10 мкМ вперед праймера, 5 μl10 мкМ обратного праймера, 0,5 мкл (1,25 U) термостабильный ДНК - полимеразы, 10 мкл 5х буфера ДНК - полимеразы, 5 мкл дНТФ смесь (2 мМ каждого), и 23,5 мкл дН 2 O.
Примечание: ДНК-матрицы могут быть геномной ДНК, полученный из штамма дикого типа для мишени данного гена гистона изолированы с использованием стандартных продуры 10. Для учета различий в концентрации ДНК и уровень примесей в различных геномных препаратов, рекомендуется , чтобы оптимизировать реакции с использованием либо неразбавленный ДНК или различных разведений геномных препаратов (например, 1:10 и 1: 100). Прямой праймер должен отжечь в области выше по течению от гена-мишени. Обратный праймер должен отжигать в ORF, быть ~ 40 нуклеотидов в длину, и содержит требуемую мутацию () где - то в середине его (см Фигура 1В-1 и репрезентативные результаты раздел примеров). Использование ДНК-полимеразы высокой точности рекомендуется для того, чтобы снизить частоту нежелательных мутаций в процессе синтеза продуктов ПЦР. - Для генерирования ПЦР - продуктов , соответствующих второй половине гена (продукт Б на рисунке 1В), установить реакцию , как указано в 2.1.1.1 , но с различными праймерами.
Примечание: Передняя ПОИМер должна отжечь в ORF, будет ~ 40 нуклеотидов в длину, и содержат требуемую мутацию (ы) где-то в середине. Следует отметить, что мутации (ов) в этом примере является обратным комплементом мутации (ов) в обратного праймера на этапе 2.1.1.1. Обратный праймер должен отжечь в области ниже по течению гена - мишени (см Фигура 1В-1 и репрезентативные результаты раздел примеров).
- Для создания продуктов ПЦР , соответствующие первой половине гена (продукт А на рисунке 1В), установить следующие реакции: 1 мкл матричной ДНК, 5 μl10 мкМ вперед праймера, 5 μl10 мкМ обратного праймера, 0,5 мкл (1,25 U) термостабильный ДНК - полимеразы, 10 мкл 5х буфера ДНК - полимеразы, 5 мкл дНТФ смесь (2 мМ каждого), и 23,5 мкл дН 2 O.
- Поместите реакции в амплификатор со следующими параметрами: 94 ° С 30 сек; 30 циклов следующих параметров: 98 ° С 10 сек, 60 ° C 5 сек, 72 ° С 1,5 мин; и 72 ° С 10 мин.
Примечание: Оптимизация параметров ПЦР может потребоваться для определенных наборов праймеров и целевого гена гистона.
- Приготовьте две реакции ПЦР следующим образом:
- Выполнить 20 - 50 мкл материала от ПЦР-реакций, на 0,9% с низкой температурой плавления агарозном геле в 89 мМ Трис-основание, 89 мМ борной кислоты, 2,5 мМ ЭДТА (КЭ) буфера.
- Вырезать агарозном секции геля, содержащего пр ПЦРoducts из геля, используя чистую скальпель или лезвие бритвы и передавать друг к 1,5 мл трубки микроцентрифужных. Храните агарозном разделы, содержащие продукты ПЦР при температуре от -20 ° С до готовности к использованию.
3. Fusion ПЦР из двух частично перекрывающихся фрагментов, чтобы получить полный Размер продуктов ПЦР для интеграции
- Подготовьте шаблон для ПЦР - реакций
- Melt агарозном секции геля из шага 2.3 путем размещения микроцентрифужных труб в блок набора тепла при 65 ° С в течение 5 мин (или до полного расплавления). Вихревые трубки каждые 1 - 2 мин, чтобы облегчить процесс плавления.
- Передача определенное количество расплавленную агарозу из каждого образца (например, 50 мкл каждой, в общей сложности на 100 мкл) в одну пробирку микроцентрифужных и перемешать встряхиванием. Используйте это в качестве матрицы в ПЦР-реакций термоядерного синтеза. Поместите трубку при -20 ° С до готовности к использованию.
- Amplify большое количество полный размер ПЦР - продукта (продукт сна фиг.1В)
- Установить шесть реакции ПЦР, каждый из которых имеет следующие компоненты: 2 мкл матрицы ДНК, 10 мкл 10 мкМ прямого праймера, 10 мкл 10 мкМ обратного праймера, 1 мкл (2,5 U) термостабильной ДНК-полимеразы, 20 мкл 5х буфера ДНК полимеразы, 10 мкл дНТФ смесь (2 мМ каждого), и 47 мкл дН 2 O.
Примечание: Число реакций могут быть изменены в зависимости от эффективности ПЦР. ДНК-матрицы (см раздел 3.1.2) должен быть нагрет до 65 ° С, пока не расплавится, смешивали при помощи вортекса и добавили в прошлом к реакционной смеси для ПЦР. После добавления, аккуратно перемешать, но тщательно пипеткой решение вверх и вниз несколько раз. Для учета различий в концентрации ДНК в различных образцах, рекомендуется сначала оптимизировать реакции с использованием либо неразбавленный шаблона или различных разведений шаблона (например, 1:10 и 1: 100). Два праймера использовали следует прокалить на двух частично перекрывающихся фрагментов гена-мишени, как больнойustrated на рисунке 1В-2 и должны быть сконструированы таким образом, чтобы конечные продукты ПЦР будут иметь по крайней мере 40 пар оснований , по обе стороны , гомологичных областей , фланкирующих URA3 ORF , который будет управлять стадии гомологичной рекомбинации (см репрезентативные результаты раздел примеров). Использование ДНК-полимеразы высокой точности рекомендуется для того, чтобы снизить частоту нежелательных мутаций в процессе синтеза продуктов ПЦР. - Поместите пробирки в амплификатор со следующими параметрами: 94 ° С 30 сек; 30 циклов следующих параметров: 98 ° С 10 сек, 50 ° С 15 сек, 72 ° С 1,5 мин; и 72 ° С 10 мин.
Примечание: Оптимизация параметров ПЦР может потребоваться для определенных наборов праймеров и целевой ген гистона.
- Установить шесть реакции ПЦР, каждый из которых имеет следующие компоненты: 2 мкл матрицы ДНК, 10 мкл 10 мкМ прямого праймера, 10 мкл 10 мкМ обратного праймера, 1 мкл (2,5 U) термостабильной ДНК-полимеразы, 20 мкл 5х буфера ДНК полимеразы, 10 мкл дНТФ смесь (2 мМ каждого), и 47 мкл дН 2 O.
4. Совместное преобразование Full Size продуктов ПЦР и Backbone плазмиды, и выбор для маркера на плазмиды
- Концентрация продуктов ПЦР
- Объединяют SРеакции ПЦР IX (600 мкл всего), начиная с шага 3.2.2 в одну пробирку микроцентрифужных и перемешать встряхиванием.
- Разделите образец на три 200 мкл аликвоты в микроцентрифужных пробирках. Осадить ДНК в каждую пробирку, добавляя 20 мкл 3 М ацетата натрия (рН 5,2) и 550 мкл 100% -ного этанола. Смешайте раствор тщательно и место на льду в течение не менее 15 мин. Сбор ДНК центрифугированием при ~ 14000 мкг в течение 10 мин, промыть осадок 200 мкл 70% этанола, и воздух сухой.
- Ресуспендируют каждый ДНК гранул в 25 мкл дН 2 O, и объединить в одну трубу (в общей сложности 75 мкл).
- Дрожжи котрансформации
- Приготовьте 10 мл ночной культуры штамма генерируемой в разделе 1 , в дрожжевой экстракт пептон в Декстроза (YPD) жидкая среда 11.
- На следующее утро, инокуляции 400 мл YPD жидкой среды с 8 мл насыщенного ночной культуры и инкубировать при встряхиваниипри 30 ° С в течение 4 - 5 ч, чтобы клетки ввести логарифмической фазе роста.
- Собирают клетки центрифугированием при ~ 3,220 мкг в течение 10 мин, отбрасывать жидкую среду, и клетки вновь суспендируют в 1 объеме 10 мМ Трис-HCl (рН 8,0), 1 мМ ЭДТА, 0,1 М раствор ацетата лития (ТЭ / LiAc) ,
- Собирают клетки центрифугированием при ~ 3,220 мкг в течение 10 мин, и выбросить TE / LiAc.
- Ресуспендируют клеток в 1 мл ТЕ / LiAc.
- Установить следующие реакции коктейлем в микроцентрифужных трубки: 800 мкл клеток из стадии 4.2.5, 40 мкл кипяченой ДНК спермы / мл лосося 10 мг, в общей сложности 12,5 мкг плазмидной ДНК позвоночника, и 75 мкл концентрированного продукта ПЦР со стадии 4.1.3.
Примечание: ДНК спермы лосося должна быть кипятили в течение 5 мин и помещали на лед в течение не менее 5 мин перед использованием в реакции. Общий объем магистральная плазмидной ДНК добавленной должна быть сведена к минимуму (~ 80 мкл или меньше). См репрезентативные результаты раздел для примера позвоночника плазмыЯ бы. - Смешайте коктейль трубку тщательно и равномерно аликвоты в восьми микропробирок (трубки 1 - 8).
- Настройте следующие две трубки реакции преобразования управления:
- Труба 9 (без контроля ПЦР-продукт): 100 мкл клеток от этапа 4.2.5, 5 мкл вареного ДНК / мл спермы лосося 10 мг (кипятили в течение 5 мин; смотри стадию 4.2.6 Примечание), в общей сложности 1,56 мкг магистральная плазмидной ДНК и ПЦР-продукт не добавляют.
- Трубка 10 (не контрольную ДНК): 100 мкл клеток от этапа 4.2.5 5 мкл кипяченого ДНК спермы лосося 10 мг / мл (этап 4.2.6 Примечание), без добавления ДНК позвоночник плазмида, и никакой продукт ПЦР не было добавлено.
- Смешайте обе трубки мягко, но тщательно пипетированием вверх и вниз несколько раз.
- Выдержите десять труб при температуре 30 ° С в течение 30 мин.
- В каждую пробирку добавляют 1,2 мл 40% полиэтиленгликоля (ПЭГ 3350) в TE / LiAc. Тщательно перемешать с помощью P-1000 пипеткой до тех пор, пока раствор не станет однородным.
- Выдержите десять труб на 30° С в течение 30 мин. Аккуратно перемешать раствор с помощью пипетки вверх и вниз, а затем инкубировать пробирки при температуре 42 ° С в течение 15 мин.
- Сбор клеток путем прядения пробирки в микроцентрифуге при ~ 14000 мкг в течение 30 сек. Выбросите жидкость и клетки вновь суспендируют в 1 мл стерильной дН 2 O.
- Сбор клеток путем прядения пробирки в микроцентрифуге при ~ 14000 мкг в течение 30 сек. Выбросите жидкость и клетки вновь суспендируют в 500 мкл стерильной дН 2 O.
- Бассейн трубки 1 - 8 вместе (общий объем 4 мл) и тщательно перемешать с помощью пипетки вверх и вниз.
- Пластина 200 мкл вышеуказанной смеси на каждой из двадцати полных минимальных выбывания средних пластин 11 (плиты 1-20) для выбора основной цепи плазмиды.
- Пластина 200 мкл смеси из трубки 9 и 200 мкл смеси из трубки 10 каждый сам по собственному выбору пластины (пластины 21 и 22, соответственно).
- Инкубируют 22 пластин при 30 ° С в течение 3 - 5 дней довыберите для плазмиды трансформантов.
- Проверьте преобразования пластин через 3 - 5 дней инкубации. Приблизительно 5000 колоний должны быть видны на пластинах 1-21 (см Представитель Результаты для примера) и колонии не должны присутствовать на пластине 22.
5. Экран для 5-FOA устойчивых трансформантов
- Передача клетки из пластин 1 - 20 (и трансформация пластины 21 в качестве контроля) до 5-fluoroorotic кислоты (5-FOA) пластины 11 с репликой покрытием 12 для того , чтобы показать на экране для потери гена URA3 в результате интеграции из ПЦР-продукты в нужном месте.
- Снимите крышку пластины и нажмите пластину, содержащую колонии на стерильный бархата. Передача клетки из вельвета к пластине 5-FOA, нажав на пластину бархата. Инкубируйте пластин при 30 ° С в течение 2-х дней.
- После 2-дневной инкубации, внимательно осмотрите пластины 5-FOA для грowth.
Примечание: Событие кандидат интеграции будет представлен небольшой асимметричной "раздавленный" колонии на пластине 5-FOA - наоборот, маленькие сосочки , растущие на 5-FOA пластин, вероятно , представитель спонтанных URA3 мутаций , которые возникли в процессе роста колоний на преобразования пластины, и, таким образом , вряд ли будет представлять желаемое событие интеграции (смотри рисунок 3 в разделе репрезентативные результаты для дальнейшей разработки по данному вопросу , а также для некоторых примеров).
6. Очистка 5-FOA устойчивых колоний и потеря Backbone плазмиды
- Использование стерильных зубочистки, выбрать кандидатов колонии из пластин 5-FOA, описанных в пункте 5.2 и полоса для отдельных колоний на YPD пластин. Инкубировать в течение 2 - 3 дней при 30 ° C.
- После инкубации реплику - пластины каждая YPD очистки пластины на свежую YPD пластину, отсев пластины без урацила для проверки потеригена URA3, и второй раскрывающихся из пластины следить за наличием или отсутствием магистральную плазмиды. Инкубировать в течение 1 - 2 дней при 30 ° C.
- После инкубации идентифицировать колонию от каждого кандидата образца , который растет на плите YPD , но не растет на любом выбывания из пластины (например , колония , как ожидается, потеряли ген URA3 через событие рекомбинации и потерял магистральную плазмиды во время митотического деление клеток). Restreak такие колонии на свежих YPD пластин. Эти колонии являются кандидатами интеграции и будут проанализированы далее в шаге 7.
7. Молекулярные анализы для анализа на надлежащей интеграции мутантного аллеля
- Изолировать геномной ДНК из проб - кандидатов с использованием стандартных процедур 10.
- Amplify геномную область , охватывающую целевой сайт.
- Настройка следующей реакции ПЦР для каждого образца: 0,5 мкл матричной ДНК, 5 мкл10 мкМ прямого праймера, 5 мкл 10 мкМ обратного праймера, 0,5 мкл (2,5 единицы) Taq - ДНК - полимеразы, 5 мкл 10х Taq - ДНК - полимеразы буфера, 5 мкл дНТФ смесь (2 мМ каждого), и 29 мкл дН 2 O.
Примечание: матричную ДНК является геномная ДНК, полученной из образцов-кандидатов. Рекомендуется также включать в себя две реакции управления: один с использованием геномной ДНК , полученной из исходных гистонов geneΔ :: штамм URA3 в качестве матрицы и другой использованием геномной ДНК из штамма гистона дикого типа в качестве матрицы. Для учета различий в концентрации ДНК и уровень примесей в различных геномных препаратов, рекомендуется , чтобы оптимизировать реакции с использованием либо неразбавленный ДНК или различных разведений геномных препаратов (например, 1:10 и 1: 100). Важно , чтобы убедиться , что эти праймеры отжигают с ДНК - последовательностей за пределами области , охваченной предполагаемо интегрированного продукта ПЦР - таким образом, размер продуктов ПЦР в этих гeactions может быть использован в качестве диагностического инструмента для интеграции продуктов в правильном месте генома (см Представитель Результаты для примера). - Поместите реакции в амплификатор со следующими параметрами: 94 ° C 3 мин; 30 циклов следующих параметров: 94 ° C 45 сек, 50 ° C 45 сек, 72 ° С 2 мин; и 72 ° С 10 мин.
Примечание: Оптимизация параметров ПЦР может потребоваться для определенных наборов праймеров и целевой ген гистона.
- Настройка следующей реакции ПЦР для каждого образца: 0,5 мкл матричной ДНК, 5 мкл10 мкМ прямого праймера, 5 мкл 10 мкМ обратного праймера, 0,5 мкл (2,5 единицы) Taq - ДНК - полимеразы, 5 мкл 10х Taq - ДНК - полимеразы буфера, 5 мкл дНТФ смесь (2 мМ каждого), и 29 мкл дН 2 O.
- Обработка продуктов ПЦР
- Выполните по 20 мкл из каждой реакции на 0,8% агарозном геле КЭ.
- Оценка размера продуктов ПЦР с использованием ДНК - стандартами в качестве эталона , чтобы определить , является ли ген URA3 , был успешно заменен предполагаемо мутантного гена гистона (см Представитель результаты примера).
Примечание: В некоторых случаях желательно мутации (ы) введены в гены гистонов либо создать или разрушить ограничение сидетье. Если это так, то наличие желаемой мутации в ПЦР-продукты размера, указывающей правильной интеграции может быть оценена путем воздействия на продукты к перевариванию с соответствующим ферментом рестрикции с последующим анализом гель-электрофореза (см Представитель результаты примера) , - Предметные ПЦР-продукты размера, указывающей правильной интеграции ДНК-секвенированию, чтобы подтвердить наличие желаемой мутации (ов), а также, чтобы гарантировать, что никакие дополнительные мутации не были введены в геном.
Результаты
Описана генерация hht2 аллеля , выражающей мутантный белок гистона H3 укрывают замещение в положении 53 из аргинином к глутаминовой кислоты (Н3-R53E мутант) в качестве типичного примера целевой стратегии Situ мутагенеза.
Мы создали ш...
Обсуждение
Высокий уровень гомологии последовательностей между двумя неаллельных генов , которые кодируют для каждого из четырех основных белков гистонов в клетках гаплоидных S.cerevisiae , может представлять собой проблему для исследователей , которые хотят конкретно адресовано одному из двух ...
Раскрытие информации
The authors declare that they have no competing financial interests.
Благодарности
We thank Reine Protacio for helpful comments during the preparation of this manuscript. We express our gratitude to the National Science Foundation (grants nos. 1243680 and 1613754) and the Hendrix College Odyssey Program for funding support.
Материалы
| Name | Company | Catalog Number | Comments |
| 1 kb DNA Ladder (DNA standards) | New England BioLabs | N3232L | |
| Agarose | Sigma | A5093-100G | |
| Boric Acid | Sigma | B0394-500G | |
| dNTP mix (10 mM each) | ThermoFisher Scientific | R0192 | |
| EDTA solution (0.5 M, pH 8.0) | AmericanBio | AB00502-01000 | |
| Ethanol (200 Proof) | Fisher Scientific | 16-100-824 | |
| Ethylenediaminetetraacetic acid disodium salt dihydrate (EDTA) | Sigma | E4884-500G | |
| Lithium acetate dihydrate | Sigma | L6883-250G | |
| MyCycler Thermal Cycler | BioRad | 170-9703 | |
| Poly(ethylene glycol) (PEG) | Sigma | P3640-1KG | |
| PrimeSTAR HS DNA Polymerase (high fidelity DNA polymerase) and 5x buffer | Fisher Scientific | 50-443-960 | |
| Salmon sperm DNA solution | ThermoFisher Scientific | 15632-011 | |
| Sigma 7-9 (Tris base, powder form) | Sigma | T1378-1KG | |
| Sodium acetate trihydrate | Sigma | 236500-500G | |
| Supra Sieve GPG Agarose (low metling temperature agarose) | AmericanBio | AB00985-00100 | |
| Taq Polymerase and 10x Buffer | New England BioLabs | M0273X | |
| Toothpicks | Fisher Scientific | S67859 | |
| Tris-HCl (1 M, pH 8.0) | AmericanBio | AB14043-01000 | |
| a-D(+)-Glucose | Fisher Scientific | AC170080025 | for yeast media |
| Agar | Fisher Scientific | DF0140-01-0 | for yeast media |
| Peptone | Fisher Scientific | DF0118-07-2 | for YPD medium |
| Yeast Extract | Fisher Scientific | DF0127-17-9 | for YPD medium |
| 4-aminobenzoic acid | Sigma | A9878-100G | for complete minimal dropout medium |
| Adenine | Sigma | A8626-100G | for complete minimal dropout medium |
| Glycine hydrochloride | Sigma | G2879-100G | for complete minimal dropout medium |
| L-Alanine | Sigma | A7627-100G | for complete minimal dropout medium |
| L-Arginine monohydrochloride | Sigma | A5131-100G | for complete minimal dropout medium |
| L-Asparagine monohydrate | Sigma | A8381-100G | for complete minimal dropout medium |
| L-Aspartic acid sodium salt monohydrate | Sigma | A6683-100G | for complete minimal dropout medium |
| L-Cysteine hydrochloride monohydrate | Sigma | C7880-100G | for complete minimal dropout medium |
| L-Glutamic acid hydrochloride | Sigma | G2128-100G | for complete minimal dropout medium |
| L-Glutamine | Sigma | G3126-100G | for complete minimal dropout medium |
| L-Histidine monohydrochloride monohydrate | Sigma | H8125-100G | for complete minimal dropout medium |
| L-Isoleucine | Sigma | I2752-100G | for complete minimal dropout medium |
| L-Leucine | Sigma | L8000-100G | for complete minimal dropout medium |
| L-Lysine monohydrochloride | Sigma | L5626-100G | for complete minimal dropout medium |
| L-Methionine | Sigma | M9625-100G | for complete minimal dropout medium |
| L-Phenylalanine | Sigma | P2126-100G | for complete minimal dropout medium |
| L-Proline | Sigma | P0380-100G | for complete minimal dropout medium |
| L-Serine | Sigma | S4500-100G | for complete minimal dropout medium |
| L-Threonine | Sigma | T8625-100G | for complete minimal dropout medium |
| L-Tryptophan | Sigma | T0254-100G | for complete minimal dropout medium |
| L-Tyrosine | Sigma | T3754-100G | for complete minimal dropout medium |
| L-Valine | Sigma | V0500-100G | for complete minimal dropout medium |
| myo-Inositol | Sigma | I5125-100G | for complete minimal dropout medium |
| Uracil | Sigma | U0750-100G | for complete minimal dropout medium |
| Ammonium Sulfate | Fisher Scientific | A702-500 | for complete minimal dropout medium |
| Yeast Nitrogen Base | Fisher Scientific | DF0919-07-3 | for complete minimal dropout medium |
| 5-Fluoroorotic acid (5-FOA) | AmericanBio | AB04067-00005 | for 5-FOA medium |
Ссылки
- Luger, K., Mader, A. W., Richmond, R. K., Sargent, D. F., Richmond, T. J. Crystal structure of the nucleosome core particle at 2.8 A resolution. Nature. 389 (6648), 251-260 (1997).
- Campos, E. I., Reinberg, D. Histones: annotating chromatin. Annu Rev Genet. 43, 559-599 (2009).
- Rando, O. J., Winston, F. Chromatin and transcription in yeast. Genetics. 190 (2), 351-387 (2012).
- Duina, A. A., Miller, M. E., Keeney, J. B. Budding yeast for budding geneticists: a primer on the Saccharomyces cerevisiae model system. Genetics. 197 (1), 33-48 (2014).
- Storici, F., Resnick, M. A. Delitto perfetto targeted mutagenesis in yeast with oligonucleotides. Genet Eng (N Y). 25, 189-207 (2003).
- Gray, M., Kupiec, M., Honigberg, S. M. Site-specific genomic (SSG) and random domain-localized (RDL) mutagenesis in yeast. BMC Biotechnol. 4, 7 (2004).
- Erdeniz, N., Mortensen, U. H., Rothstein, R. Cloning-free PCR-based allele replacement methods. Genome Res. 7 (12), 1174-1183 (1997).
- Brachmann, C. B., et al. Designer deletion strains derived from Saccharomyces cerevisiae S288C: a useful set of strains and plasmids for PCR-mediated gene disruption and other applications. Yeast. 14 (2), 115-132 (1998).
- Lundblad, V., Hartzog, G., Moqtaderi, Z. Manipulation of cloned yeast DNA. Curr Protoc Mol Biol. Chapter 13, (2001).
- Hoffman, C. S. Preparation of yeast DNA. Curr Protoc Mol Biol. Chapter 13, (2001).
- Treco, D. A., Lundblad, V. Preparation of yeast media. Curr Protoc Mol Biol. Chapter 13, (2001).
- Lederberg, J., Lederberg, E. M. Replica plating and indirect selection of bacterial mutants. J Bacteriol. 63 (3), 399-406 (1952).
- Sikorski, R. S., Hieter, P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae. Genetics. 122 (1), 19-27 (1989).
- Johnson, P., et al. A systematic mutational analysis of a histone H3 residue in budding yeast provides insights into chromatin dynamics. G3 (Bethesda). 5 (5), 741-749 (2015).
- DiCarlo, J. E., et al. Genome engineering in Saccharomyces cerevisiae using CRISPR-Cas systems. Nucleic Acids Res. 41 (7), 4336-4343 (2013).
- Cross, S. L., Smith, M. M. Comparison of the structure and cell cycle expression of mRNAs encoded by two histone H3-H4 loci in Saccharomyces cerevisiae. Mol Cell Biol. 8 (2), 945-954 (1988).
- Libuda, D. E., Winston, F. Amplification of histone genes by circular chromosome formation in Saccharomyces cerevisiae. Nature. 443 (7114), 1003-1007 (2006).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены