
Method Article
الترتيب الزمني للبيانات التعبير الحيوي من التعبير المكاني مفصل خرائط
In This Article
Summary
The segmentation clock drives oscillatory gene expression across the pre-somitic mesoderm (PSM). Dynamic Notch activity is key to this process. We use imaging and computational analyses to extract temporal dynamics from spatial expression data to demonstrate that Delta ligand and Notch receptor expression oscillate in the vertebrate PSM.
Abstract
During somitogenesis, pairs of epithelial somites form in a progressive manner, budding off from the anterior end of the pre-somitic mesoderm (PSM) with a strict species-specific periodicity. The periodicity of the process is regulated by a molecular oscillator, known as the "segmentation clock," acting in the PSM cells. This clock drives the oscillatory patterns of gene expression across the PSM in a posterior-anterior direction. These so-called clock genes are key components of three signaling pathways: Wnt, Notch, and fibroblast growth factor (FGF). In addition, Notch signaling is essential for synchronizing intracellular oscillations in neighboring cells. We recently gained insight into how this may be mechanistically regulated. Upon ligand activation, the Notch receptor is cleaved, releasing the intracellular domain (NICD), which moves to the nucleus and regulates gene expression. NICD is highly labile, and its phosphorylation-dependent turnover acts to restrict Notch signaling. The profile of NICD production (and degradation) in the PSM is known to be oscillatory and to resemble that of a clock gene. We recently reported that both the Notch receptor and the Delta ligand, which mediate intercellular coupling, themselves exhibit dynamic expression at both the mRNA and protein levels. In this article, we describe the sensitive detection methods and detailed image analysis tools that we used, in combination with the computational modeling that we designed, to extract and overlay expression data from distinct points in the expression cycle. This allowed us to construct a spatio-temporal picture of the dynamic expression profile for the receptor, the ligand, and the Notch target clock genes throughout an oscillation cycle. Here, we describe the protocols used to generate and culture the PSM explants, as well as the procedure to stain for the mRNA or protein. We also explain how the confocal images were subsequently analyzed and temporally ordered computationally to generate ordered sequences of clock expression snapshots, hereafter defined as "kymographs," for the visualization of the spatiotemporal expression of Delta-like1 (Dll1) and Notch1 throughout the PSM.
Introduction
القطع البدنية النمطية هي أول القطاعات التي تشكلت في التطويل محور الجسم في تطوير أنواع الفقاريات وهي السلائف في العمود الفقري، الأضلاع، والأدمة الأنسجة، وكذلك العضلات والخلايا البطانية. أثناء تكون الجسيدات، تشكل القطع البدنية النمطية الظهارية من الأديم المتوسط unsegmented presomitic (PSM) (إعادة النظر في المرجع 1). وينظم هذه العملية من خلال "ساعة تجزئة"، والذي يتكون من شبكة من الجينات متذبذبة والبروتينات، ومعظمهم ينتمون إلى مسار الشق الإشارات. وتتكون الساعة تجزئة مختلف الحلقات ردود الفعل السلبية، والتي تمكن من إنتاج نابض النشاط الشق داخل خلية واحدة 2 (إعادة النظر في المراجع 3-6). في حين أن طريقة الخلايا من التذبذب يتميز كذلك، فإنه لا يزال غير معروف إلى حد كبير كيف يتم تنسيق هذه التذبذبات في جميع أنحاء الأنسجة PSM. وقد تبين في الآونة الأخيرة، سواء من خلال الدراسات التجريبية والنظرية، أن هذه التذبذبات ضروريهل لعملية تكون الجسيدات وأن مسار الشق يلعب دورا حاسما في عملية على حد سواء تجزئة والتعبير الجيني متذبذبة 7 و 8. ومع ذلك، فقد أفيد على نطاق واسع أن الشق مستقبلات 1 (Notch1) ودلتا تشبه يجند (دلل) -1 ديك التدرجات ثابتة في PSM 9 و 10 و 11.
ونحن افترضنا أن التذبذبات التي تعتمد على الشق من ساعة PSM تجزئة تعتمد على تفعيل الدوري لمستقبلات الرئيسي الشق مسار ويجند، Notch1 وDLL1، على التوالي، عبر PSM الماوس. وكانت نتائج الدراسات السابقة التي سجلت التدرج منقاري الذيلية ثابت من هذه البروتينات الواجبة، نتوقع، لعدم وجود حساسية في التقنيات المناعية. لذا كانوا قادر على الكشف عن التقلبات على مستوى منخفض من DLL1 وNotch1 في PSM الذيلية.
لديناوضعت البريد وسيلة لدراسة عن كثب هذه العوامل، والجمع بين البيانات التجريبية مع النماذج الرياضية للتنبؤ آلية التي يتم من خلالها تنسيق التذبذبات من البروتينات من المكونات على مدار الساعة عبر PSM 12.
ويتمثل الهدف العام من هذه الطريقة هو كشف وتحديد مستوى منخفض، والتعبير البروتين الحيوي في إدارة المشتريات والتوريدات ولتعيين ملامح التعبير من البروتينات التي تهم وفقا للتعبير عن الجينات على مدار الساعة المعروفة، جماعات متطرفة مجنونة (Lfng). منذ دورة واحدة على مدار الساعة تجزئة في جنين الفأر يأخذ 2 ساعة لاستكمال، هناك حاجة عينات مختلفة لبناء الشخصية الزمانية المكانية كاملة من DLL1 وNotch1 تعبير البروتين أثناء واحدة التذبذب Lfng في إدارة المشتريات والتوريدات. وبذلك نكون قد وضعت هذا البروتوكول للسماح للكشف عن الإنتاجية العالية للتعبير البروتين على مستوى منخفض في كامل جبل، إإكسبلنتس PSM المقابل. ومع ذلك، يمكن أيضا أن تكون هذه التقنية مفيدة للدراسات روتهدف قبعة لوصف ديناميات البروتين على مستوى منخفض داخل أي نوع من الأنسجة الجنينية التي يمكن تقسيمها إلى نصفين المقابل.
Protocol
تم إجراء جميع التجارب في إطار مشروع رقم الترخيص 6004219 في الالتزام الصارم الحيوانات (الإجراءات العلمية) لعام 1986 والقوانين في المملكة المتحدة وزارة الداخلية من الممارسة لاستخدام الحيوانات في إجراءات علمية.
1. PSM يزدرع تشريح
- الحصول على الأنسجة الذيل من الأجنة التي تنتجها التزاوج توقيت البرية من نوع (CD1) الفئران (13). لفترة وجيزة، في يوم الجنينية (E) 10.5، الموت ببطء الماوس المانحة الحامل في غرفة غاز ثاني أكسيد الكربون. حصاد قرن الرحم ووضعه في حل 1X العقيمة الفوسفات مخزنة المالحة (PBS) وفقا للإجراءات الترخيص وزارة الداخلية أو القواعد المحلية ما يعادلها. نقل قرن الرحم إلى صحن زراعة الأنسجة التي تحتوي على الطازجة، برنامج تلفزيوني العقيمة. تنفيذ جميع الخطوات اللاحقة تشريح في هذا الحل.
- تحت مجهر تشريحي، وقطع غشاء عضلي سميك من قرن الرحم باستخدام مقص المنحنية واستخراج كل الجنين بعناية باستخدام ملقط غرامة. اعتن بنفسكللتأكد من أن الأنسجة ذيل غير تالف في هذه العملية. باستخدام مقص منحني وملقط غرامة، بعيدا تشريح الكيس الذي يحيط بالجنين من كل جنين، مع الحرص على عدم الإضرار الجنين.
- استخدام إما إبرة جراحية أو مقص منحني لحصاد الأنسجة ذيل كل الجنين عن طريق خفض الخلفي الجنين للبراعم الأطراف الخلفية.
- توازن الأنسجة الذيل الجانب أسفل بطني باستخدام كل من الملقط والإبرة. إنشاء أزواج من إإكسبلنتس PSM من كل ذيل الجنينية التي كتبها تشريح الأنسجة الذيل إلى نصفين على طول خط الوسط. إجراء حركة التأرجح لطيف مع إبرة. تأكد من أن أنبوب، الحبل الظهري، والأنسجة العصبية PSM تقسم بالتساوي بين إإكسبلنتس اثنين.
- ماصة كل يزدرع PSM المقابل على الجانب السفلي من 35 ملم ثقافة البلاستيك طبق غطاء في حجم صغير من قبل تحسنت (37 درجة مئوية) مستنبت (DMEM-F12 + 0.1٪، الجلوتامين بديلا تستكمل مع 10٪ مصل العجل الجنين 10 نانومتر الإنسان Bfgf، و 1٪ البنسلين / الستربتومايسين). <لى> ضع الطبق على الجزء العلوي من الغطاء وبسرعة عكس ذلك بحيث يتم تعليق الأنسجة PSM من الغطاء في "معلق قطرة" من المتوسطة. ثقافة إإكسبلنتس PSM في غرفة ترطيب عند 37 درجة مئوية لمدة 1-2 ساعة.
- نقل أزواج من إإكسبلنتس PSM إلى الآبار الفردية من 24 جيدا نسيج لوحة الثقافة. احتضان في 4٪ امتصاص العرق في برنامج تلفزيوني لمدة 1 ساعة في درجة حرارة الغرفة (RT) أو 4 درجات مئوية خلال الليل (O / N). تنبيه: لامتصاص العرق هي سامة، ويجب اتخاذ تدابير السلامة المناسبة عند التعامل مع هذا الحل.
ملاحظة: إجراء جميع الغسيل وحضانة الخطوات اللاحقة في 24-جيدا لوحة زراعة الأنسجة. - غسل الآبار العينة في برنامج تلفزيوني في RT على منصة هزاز، وذلك باستخدام غرامة ماصة باستير البلاستيك لتبادل الحل برنامج تلفزيوني على عينات لبرنامج تلفزيوني جديد 3-4 مرات. معالجة واحدة يزدرع PSM من كل زوج باستخدام المناعية (الخطوة 2) والآخر باستخدام الفلورية في الموقع التهجين لجين ساعة المعروفة (شارعالجيش الشعبي 3).
2. المناعية من PSM إإكسبلنتس
- يغسل بعضكم يزدرع PSM من كل زوج الجنينية إنشاؤه في الخطوة 1 في 2٪ تريتون X-100 في برنامج تلفزيوني لمدة 1 ساعة على RT على منصة هزاز، ثم شطف عينات لفترة وجيزة في برنامج تلفزيوني. استبدال برنامج تلفزيوني على عينات مع عرقلة الحل (2٪ زلال المصل البقري (BSA) و 10٪ مصل الماعز العادي (خ ع) في برنامج تلفزيوني + 0.1 توين 20٪)، واحتضان O / N عند 4 درجات مئوية على منصة هزاز.
ملاحظة: يجب إجراء جميع يغسل اللاحقة والخطوات حضانة في هذا القسم في RT على منصة هزاز، ما لم ينص على خلاف ذلك. حلول غسل يمكن تغييرها بسهولة باستخدام البلاستيك أو الزجاج ماصة باستير يميل غرامة. - تمييع المطلوب الابتدائية الأجسام المضادة / الأجسام المضادة في المخزن العمل (0.1٪ BSA، 0.3٪ خ ع، و 0.2٪ تريتون X-100 في برنامج تلفزيوني). في هذا المثال، يخفف من DLL1 وNotch1 الأجسام المضادة 01:25 العازلة في العمل.
سوف تكون هناك حاجة الأمثل لتحديد عامل التخفيف المناسبة المطلوبة في ثي ملاحظة:الصورة الخطوة إذا تم استخدام الأجسام المضادة بديلة. - احتضان إإكسبلنتس في حل الأجسام المضادة ل3-5 أيام في 4 درجات مئوية على منصة هزاز. تأكد من تضمين بعض العينات مع العازلة العمل التي لا تحتوي على الأجسام المضادة الأولية لتكون بمثابة الضوابط الأجسام المضادة الثانوية.
- استرداد حل الأجسام المضادة الأولية في أنبوب التخزين 1.5 مل باستخدام ماصة وتخزينها في 4 درجات مئوية.
ملاحظة: المستردة الأجسام المضادة الأولية يمكن استخدامها عدة مرات، اعتمادا على الأجسام المضادة المستخدمة. - أداء 2 يغسل من العينات ل5-10 دقيقة لكل منهما في برنامج تلفزيوني، تليها 3 غسلات لمدة 10 دقيقة لكل منهما في 2٪ تريتون X-100 في برنامج تلفزيوني في RT على منصة هزاز.
- تمييع fluorescently المسمى الثانوية الضد / الأجسام المضادة (حاتمة مطابقة لالأجسام المضادة الأولية / الأجسام المضادة المستخدمة) في المخزن العمل. اختياريا، إضافة 20 ميكروغرام / مل هويشت 33342 إلى هذا الحل لمباين نوى.
قد تكون هناك حاجة الأمثل لتحديد عامل التخفيف المناسبة المطلوبة في هذه الخطوة: ملاحظة. في هذا البريدxample، وهو عامل التخفيف من 1: تم استخدام 400 عادة. - أجهزة الطرد المركزي في حل الضد الثانوية لمدة 10 دقيقة في 16 x ج لمنع تشكيل المجاميع الضد. إضافة 250-500 ميكرولتر من محلول الأجسام المضادة الثانوية إلى كل عينة بشكل جيد، مع الحرص على عدم استخدام ميكرولتر القليلة الماضية من الحل، والتي قد تحتوي على مجاميع الأجسام المضادة.
- تغطية لوحة عينة بورق القصدير لتقليل التعرض للضوء واحتضان العينات في حل الضد الثانوية لمدة 3 - 5 ايام في 4 درجات مئوية في الظلام.
- قبل أخذ عينات تصاعد، وغسل العينات مرتين لمدة 10 دقيقة في كل من 0.1٪ توين 20 في برنامج تلفزيوني (PBST) ومرة واحدة لمدة 5 دقائق في برنامج تلفزيوني في RT على منصة هزاز (راجع الخطوة 4).
3. نيون في الموقع التهجين (FISH) لPSM إإكسبلنتس
- إذا المخزنة في سفينة بديلة، ونقل إإكسبلنتس PSM المقابل المتبقية إلى الآبار الفردية من 24 جيدا نسيج لوحة الثقافة.
- غسل العينات ل10 دقيقة في 50٪ من الإيثانول في PBST، ومن ثم أداء 2 يغسل لمدة 10 دقيقة في كل من الإيثانول بنسبة 100٪ على منصة هزاز في RT ليذوى الأنسجة.
ملاحظة: يجب إجراء جميع يغسل اللاحقة والخطوات حضانة في هذا القسم في RT على منصة هزاز، ما لم ينص على خلاف ذلك. - ترطيب الأنسجة عن طريق الغسيل لمدة 10 دقيقة في 50٪ من الإيثانول في PBST، تليها غسل مرتين لمدة 5 دقائق في كل من PBST.
ملاحظة: خطوات 3.2 و 3.3 خطوات تثبيت اللازمة لهذا البروتوكول والتي لا يمكن حذفها. - احتضان هذه العينات مع 10 ميكروغرام / مل بروتين كاف في 0.1٪ توين 20 في برنامج تلفزيوني (PBST) لمدة 5 دقائق من دون الانفعالات. بسرعة إزالة بروتين كاف وشطف عينات لفترة وجيزة مع PBST قبل بعد إصلاح الأنسجة لمدة 30 دقيقة في 4٪ الفورمالديهايد + 0.1٪ غلوتارالدهيد في PBST. تنبيه: كل من الفورمالديهايد وغلوتارالدهيد سامة، ويجب اتخاذ تدابير السلامة المناسبة عند التعامل مع هذه الحلول.
ملاحظة: غسل التاليةوالخطوات حضانة شاركت فيها 50٪ و 100٪ يمزج التهجين (الخطوات 3،6-3،9) يجب أن يتم تنفيذ دون الانفعالات. - بعد غسل العينات مرتين لمدة 10 دقيقة لكل منهما في PBST، وغسل العينات مرة واحدة في مزيج التهجين 50٪ (مناسبة للتحقيقات intronic: 50٪ الفورماميد، 5X المالحة الصوديوم سترات (SSC)، و 5 ملي EDTA، 50 ميكروغرام / مل الحمض الريبي النووي النقال، أعد 0.2٪ توين 20، 0.1٪ SDS، و 100 ميكروغرام / مل الهيبارين) في PBST في RT. احتضان هذه العينات في هذا الحل لمدة 10 دقيقة عند 65 درجة مئوية دون التحريض.
- شطف عينات مرتين مع ما قبل تحسنت (65 ° C) مزيج التهجين قبل احتضان العينات في مزيج التهجين ل≥ 2 ساعة (تصل إلى 48 ساعة) عند 65 درجة مئوية (مرات حضانة أطول تحسين الناتج إشارة إلى الضجيج النقيض) . إزالة مزيج التهجين من الخطوة السابقة، واستبدالها ،25-،5 مل من قبل حرارة (65 درجة مئوية) مزيج التهجين تحتوي على digoxigenin (DIG) -labeled مكافحة الشعور RNA التحقيق ضد عنصر مدار الساعة تجزئة معروفة.
ليسE: على سبيل المثال، تم استخدام جماعات متطرفة مجنونة intronic (Lfng (ط)) التحقيق في تركيز 20 ميكرولتر / مل للكشف عن الوليد Lfng مرنا. التخفيف المستخدمة في هذه الخطوة هو تعتمد على التحقيق، وسوف تتطلب التحسين. - ختم لوحة باستخدام شريط لاصق لمنع التبخر واحتضان العينات في حل التحقيق لمدة ليلتين عند 65 درجة مئوية.
- باستخدام البلاستيك ماصة باستير يميل غرامة، واستعادة المسبار لإعادة استخدامها وتخزينها عند 20 درجة مئوية. شطف عينات مرتين مع ما قبل تحسنت (65 ° C) مزيج ما بعد التهجين (50٪ الفورماميد، 0.2٪ توين-20، و1X SSC) قبل غسل العينات مرتين أخريين لمدة 20 دقيقة لكل منهما عند 65 درجة مئوية في مرحلة ما قبل ارتفعت درجة حرارة المزيج بعد التهجين.
- غسل العينات لمدة 15 دقيقة عند 65 درجة مئوية في مزيج التهجين 50٪ قبل تحسنت في 0.1٪ توين 20 في محلول ملحي مخزنة تريس (TBST). شطف عينات مرتين مع TBST قبل غسل لمدة 30 دقيقة في RT في TBST على منصة هزاز.
- قبل احتضان إإكسبلنتس في المفكرهحل cking وب (TBST + 2٪ حجب كاشف العازلة (ببر) + 20٪ المعالج حراريا مصل الماعز) لمدة لا تقل عن 2 ساعة. استبدال هذا الحل مع عرقلة الحل الطازجة التي تحتوي على نسبة 1: 200 التخفيف من الفجل البيروكسيداز (HRP) -conjugated الأجسام المضادة لمكافحة digoxigenin. احتضان العينات O / N عند 4 درجات مئوية.
- بعد الحضانة الضد، شطف العينات 3 مرات مع TBST في RT ونقلها إلى الآبار الفردية من 24 جيدا نسيج لوحة الثقافة الجديدة. غسل إإكسبلنتس مع TBST 3 مرات لمدة 1 ساعة لكل منهما.
- في هذه المرحلة، ونقل العينات إلى 0.5 مل أنابيب التخزين أو الآبار الفردية ل48-جيدا لوحة زراعة الأنسجة للحد من حجم المطلوب من Tyramide تضخيم الإشارات (TSA) الكواشف كشف في الخطوات التالية.
- احتضان العينات في المخزن التضخيم TSA (انظر قائمة الكواشف) في RT لمدة 1 دقيقة دون التحريض تستخدم صغير كحجم وقت ممكن، وضمان أن العينات مغمورة تماما في الحل.
- إضافة TSA كاشف (انظرقائمة الكواشف) إلى المخزن المؤقت التضخيم عينة في التخفيف من 01:50. بسرعة مزيج الحل حتى يتم توزيع الكاشف TSA بالتساوي، وتغطية لوحة أو الأنابيب في احباط القصدير، واحتضان عينات ل60-90 دقيقة في الظلام.
- إزالة الحل التضخيم TSA وغسل العينات في TBST 3 مرات لمدة 5 دقائق لكل منهما. نقل إإكسبلنتس مرة أخرى إلى 24-جيدا لوحة زراعة الأنسجة لزيادة حجم غسل واحتضان العينات في 1٪ بيروكسيد الهيدروجين في TBST لمدة 1 ساعة. غسل العينات مع TBST 3 مرات لمدة 5 دقائق لكل منهما، ثم مرتين لمدة 5 دقائق مع كل PBST قبل أخذ عينات تركيب (راجع الخطوة 4).
4. إعداد نموذج للتصوير
- إعداد واحدة مشحونة شريحة التصاق الزجاج لكل زوج يزدرع بإضافة 0.12 ملم الفواصل التصوير سميكة، مما يمنع عينات من سحقهم من خلال إضافة ساترة. إزالة بطانة لاصقة من سطح واحد من فاصل ووضعه الجانب اللاصق لأسفل على شريحة زجاجية، الضغط وirmly لاغلاق هل إلى الشريحة.
ملاحظة: للحصول على الخطوات المتبقية، تسعى للحفاظ على عينات في ضوء انخفاض أو في الظلام لتجنب photobleaching من. ماصة أزواج يزدرع على شريحة مستعدة باستخدام الزجاج ماصة باستير داخل مركز للمباعدة، وضمان أن الجانب تشريح لليزدرع يواجه الشريحة. ترتيب أزواج المقابل من الجانب إإكسبلنتس إلى جنب. - إزالة أكبر قدر من السائل ممكن من الشريحة باستخدام ماصة باستير الزجاج والفتيل من أي الرطوبة المتبقية المحيطة العينات باستخدام قطعة من المناديل الورقية من الوبر منخفض مطوية.
- السماح للعينات لتنضم إلى شريحة ل45 - 60 ثانية، حتى تبدأ الأنسجة لتظهر لزجة وشفافة. خلال هذا الوقت، وإزالة بطانة ما تبقى لاصقة من فاصل باستخدام ملقط. لا تسمح عينات لتجف.
- إضافة قطرة كبير من مرس المزدوجة وظيفة وحل المقاصة (0.5٪ ف phenylenediamine و 20 ملي تريس، ودرجة الحموضة 8.8، في 90٪ الجلسرين) للعينات داخل المركزمن الفاصل. ملاحظة: هذا الحل يتحول اللون البني / أسود عندما يسمح للأكسدة.
- وضع بعناية ساترة دائرية (رقم 1.5) عبر العينات، وضمان أن مرس توزع بالتساوي وأن جميع حواف ساترة اجراء اتصالات مع الفاصل. ضع الشريحة انزلق غطاء رأسا على عقب على بعض المناديل الورقية من الوبر منخفضة.
- اضغط لأسفل بشدة لضمان أن تكون ساترة تلتزم تماما هل وأن تتم إزالة أي مرس الزائد. كرر حتى لا البقع أكثر مرس ورقة.
- نظيفة وتسمية الشريحة (ق) بشكل مناسب، وتخزينها في الظلام حتى التصوير، على المدى القصير عند درجة حرارة -20 درجة مئوية أو طويل الأجل في -80 درجة مئوية. بعد إزالة الشرائح من التخزين، والسماح لهم لذوبان الجليد بشكل كامل قبل التصوير.
- صورة العينات التي شنت باستخدام مجهر متحد البؤر مع اكتساب القرميد وهدف تضخم عالية. صورة أزواج يزدرع باستخدام الهدف الغمر النفط 40X في زي فترات 4 ميكرون باستخدام 488 نانومتر، 568 نانومتر وبين 647 نانومتر لى الليزرمتنوعه لإثارة fluorophores الأخضر والأحمر، وبعيدة الحمراء، على التوالي، يعملون على البروتين وكشف مرنا في هذه الدراسة 12.
ومخيط تجانب صور ما بعد الاستحواذ لتشكيل صورة واحدة للتحليل: ملاحظة.
5. بعد اكتساب تحليل الصور
- استخدام صورة برامج التحليل لتحديد المنطقة ذات الاهتمام داخل PSM من كل عينة تجريبية.
- لقياس مستويات التعبير، خلفية طرح وصور العتبة إلى مستوى عينة التحكم لم الأولية قبل الكمي لاحق. تعريف الأصل، محور، وطول وحدة لكل عينة.
- حساب كثافة مضان كدالة للموقف على طول محور تطبيع rostro-الذيلية لكل من العينات M 12. بعد تطبيع المؤامرات كثافة، وضع الجانب ملامح كثافة إلى جنب، والحصول على كثافة مصفوفة و (ط، ي) التي تصف كثافة في ط موقف المكاني في ي عينة عشر والعشرين.
6. الترتيب الزمني للعينات
- لاستنتاج ترتيب الزمني للعنصر مدار الساعة المعروفة، وتحديد مصفوفة شدته. ثم، وإعادة ترتيب أعمدة مصفوفة كثافة وذلك للحصول على نمط الدوري مؤقتا. للقيام بذلك، وتحديد وظيفة

حيث A (و ي. ك) تمثل وظيفة الارتباط الذاتي من ي عمود F و A T عشر هي وظيفة الارتباط الذاتي الهدف، الذي تم اختياره لفرض الدورية الزمنية للنمط، التي قدمها
- استخدام متروبوليس، هاستينغز (أو خوارزمية التقليل آخر) 12 لتحديد ترتيب العينات التي تقلل من وظيفة ز. وبالتالي، تحديد ترتيبعينات M الذي يزيد من وتيرة الزمنية عنصر مدار الساعة معروفة.
- عن طريق ترتيب الزمني يستدل من العينات M، بناء على أمر مخطاط التموج لنمط التعبير في قناة شاركت 12.
النتائج
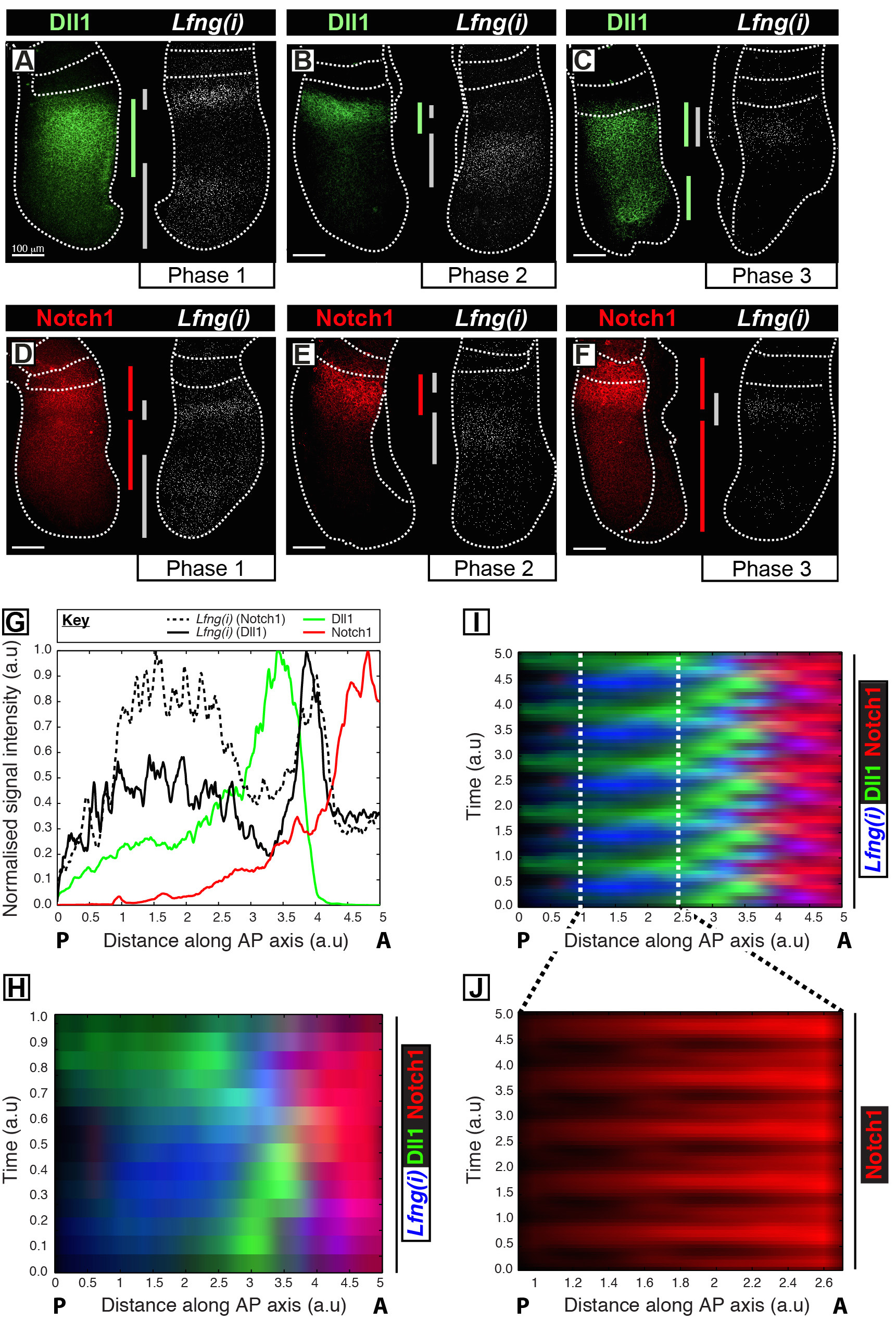
يسمح هذا البروتوكول تصور الشخصية الزمانية المكانية من البروتين من الفائدة إلى جانب النسخ الجيني على مدار الساعة في PSM الماوس 12. على سبيل المثال، أظهرت DLL1 (الشكل 1A-C) وNotch1 (1D الشكل-F) تعبير البروتين إلى التأرجح من التزامن مع النسخ الوليد للتنظيم الشق الساعة تجزئة الجين Lfng. الكمي لDLL1، Notch1، وLfng (ط) كثافة إشارة فيما يتعلق انتيرو-الخلفي (ا ف ب) محور PSM (الشكل 1G) يكشف واضحة ديناميات التعبير متذبذبة لهذه الأهداف (الشكل 1H-J). البيانات الشخصية الزمانية المكانية من DLL1 وNotch1 تعبير البروتين في جميع أنحاء دورة على مدار الساعة هي تصور واضح وكميا باستخدام هذا البروتوكول من خلال تحليل الصور بعد الحيازة عالية الدقة بيانات الصورة الأنسجة الثابتة.
together.within الصفحات = "1"> 
الشكل 1: التصور المكانية والزمانية والكمي لDLL1 وNotch1 البروتين التعبير حيوية. (AF) أزواج من إإكسبلنتس من ستة E10.5 الأجنة (AF) تبين التوزيع المكاني للبروتين DLL1 (AC) أو البروتين Notch1 (DF) في نصف إلى جانب الكشف عن Lfng قبل مرنا (Lfng (ط)) في نصف المقابل من كل زوج المقابلة. يتم ترتيب الألواح وفقا لالمرحلة 1 (A و D)، المرحلة 2 (B و E)، والمرحلة 3 (C و F) من دورة على مدار الساعة تجزئة، على النحو الذي تحدده الشخصية المكاني للLfng (ط) التعبير. مدى مجالات التعبير عن DLL1 (الأخضر)، Notch1 (الحمراء)، وLfng (ط) (الرمادي) على طول المحور انتيرو الخلفي من PSM يكون بالتابعين رسمتها الحانات مرمزة. الخطوط المنقطة وتفصل بين مواقف الجسيدة الأكثر شكلت مؤخرا (ق)، والحواف الخارجية للإدارة المشتريات والتوريدات، والأنسجة العصبية المجاورة (C) و (E). أشرطة النطاق (أسفل اليسار من كل لوحة، AF) تمثل 100 ميكرون. (G) مثال شدة المؤامرة التي تصور الاختلاف المحوري في كثافة إشارة عبر PSM. يتم رسم البيانات من اثنين من أزواج يزدرع تظهر Lfng قبل مرنا (خط المجزأة الأسود) في يزدرع واحد مقارنة مع البروتين Notch1 (الحمراء) في يزدرع المقابل (جنين 1)، وكذلك Lfng قبل مرنا (خط الصلبة السوداء) في يزدرع آخر مقارنة مع البروتين DLL1 (الأخضر) في يزدرع المقابل (جنين 2). يتم رسم قياس كثافة إشارة (المحور الصادي) ضد الموقف المحوري (محور س. الأمامي PSM [A] إلى اليمين والخلفي للPSM [ف] إلى اليسار). (H) ومخطاط التموج تبين التوزيع المكاني للDLL1، Notch1، وLfng (ط) عبرالعديد من PSMS. كل صف من مخطاط التموج يمثل كثافة إشارة من يزدرع PSM الفردية. يتم ترتيب الصفوف في التسلسل الزمني وفقا لتوزيع الزمانية المكانية من Lfng قبل مرنا (I) والتوزيع الزماني المكاني من DLL1، Notch1، وLfng (ط) من خلال ويحاكيه التمديد الدوري لالبيانات الواردة في التذبذبات متعددة على مدار الساعة (H) ، وتسليط الضوء على طبيعة متذبذبة من DLL1 وNotch1 ديناميات التعبير. ومن أبرز تعبير البروتين (J) نابض Notch1 في PSM الذيلية التي كتبها التكبير من المنطقة ترسيمها في مخطاط التموج الظاهري هو مبين في (I). تعديل من المرجع 12. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
 الشكل 2: الكمي لديناميات المكانية والزمانية من DLL1 وNotch1 البروتين التعبير. (أ) مؤامرة سبيل المثال كثافة يصور الاختلاف المحوري في كثافة إشارة عبر PSM. تآمر البيانات من اثنين من أزواج يزدرع تظهر Lfng قبل مرنا (خط المجزأة الأسود) في يزدرع واحد مقارنة مع البروتين Notch1 (الحمراء) في النصف يزدرع المقابل، وكذلك Lfng قبل مرنا (خط الصلبة السوداء) في نصف يزدرع من الذيل الثاني مقارنة مع البروتين DLL1 (الأخضر) في النصف يزدرع المقابل من الذيل الثاني. يتم رسم شدة قياس (المحور الصادي) ضد الموقف المحوري (محور س. منقاري [A] إلى الحق والذيلية [ف] إلى اليسار). وتشير (BH) Kymographs التوزيع المكاني للNotch1، DLL1، حوال، وLfng (ط) عبر العديد من PSMS. (B و C) حوال (ب) وDLL1 (C) في التعبير أقسام إدارة المشتريات والتوريدات. (D و E) Lfng (ط) (D) وDLL1 (<قوي> E) في شطري يزدرع المقابل. (F و G) Lfng (ط) (F) وNotch1 (G) في شطري يزدرع المقابل. من المرجعي 12. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
الشكل 2: الكمي لديناميات المكانية والزمانية من DLL1 وNotch1 البروتين التعبير. (أ) مؤامرة سبيل المثال كثافة يصور الاختلاف المحوري في كثافة إشارة عبر PSM. تآمر البيانات من اثنين من أزواج يزدرع تظهر Lfng قبل مرنا (خط المجزأة الأسود) في يزدرع واحد مقارنة مع البروتين Notch1 (الحمراء) في النصف يزدرع المقابل، وكذلك Lfng قبل مرنا (خط الصلبة السوداء) في نصف يزدرع من الذيل الثاني مقارنة مع البروتين DLL1 (الأخضر) في النصف يزدرع المقابل من الذيل الثاني. يتم رسم شدة قياس (المحور الصادي) ضد الموقف المحوري (محور س. منقاري [A] إلى الحق والذيلية [ف] إلى اليسار). وتشير (BH) Kymographs التوزيع المكاني للNotch1، DLL1، حوال، وLfng (ط) عبر العديد من PSMS. (B و C) حوال (ب) وDLL1 (C) في التعبير أقسام إدارة المشتريات والتوريدات. (D و E) Lfng (ط) (D) وDLL1 (<قوي> E) في شطري يزدرع المقابل. (F و G) Lfng (ط) (F) وNotch1 (G) في شطري يزدرع المقابل. من المرجعي 12. الرجاء انقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
Discussion
خطوات حاسمة في إطار بروتوكول
يصف هذا البروتوكول طريقة حساسة لإجراء تحليل كمي لمستوى منخفض تعبير البروتين ومتذبذبة ديناميكية في إإكسبلنتس E10.5 الماوس PSM. ويتبع بروتوكول قوية لكلا المناعية والفلورية في الموقع التهجين (FISH) بواسطة عالية الدقة كله يشن التصوير متحد البؤر، ومن ثم عن طريق تحليل الصور وتجزئة الزمنية للkymographs لتوليد خريطة الزمانية المكانية للتعبير البروتين في جميع أنحاء PSM. وارتفاع نسبة الإشارة إلى الضوضاء في البروتين ومرنا كشف أمر ضروري لضمان نجاح هذه التقنية. يجب الحرص على تبادل تماما عن حلول فعالة خلال خطوات غسل والحفاظ على درجة حرارة 65 ° C يغسل في المراحل ذات الصلة من الخطوة 3. هو الأكثر فائدة لتأخذ من الوقت لمصدر الأجسام المضادة فعالة وتحقيقات RNA ضد أهداف الفائدة ولاختبار هذه الكواشف تماما على عينات جبل بأكمله قبل البدء في هذا البروتوكول.
التعديلات واستكشاف الأخطاء وإصلاحها
تنشأ القضايا الرئيسية التي يمكن مواجهتها عند تنفيذ هذا البروتوكول من ضعف قوة الكشف عن إشارة والجودة. هذا يعتمد إلى حد كبير على فعالية الأجسام المضادة أو تحقيقات RNA المستخدمة لالمناعية أو FISH الخطوات في البروتوكول، على التوالي. قد تتطلب عدد من الخطوات المختلفة الأمثل قبل أن يتحقق الكشف عن إشارة كافية. أحد الأسباب الشائعة لضعف إشارة الكشف هو تثبيت غير لائق، لا بد من أن أي PFA الطازجة أو PFA تخزينها في 4 درجة مئوية لمدة لا تزيد عن أسبوع واحد يستخدم لتحديد العينات. قد تتطلب طول التثبيت أيضا الأمثل، اعتمادا على الأجسام المضادة أو RNA التحقيق المستخدمة. عن الأجسام المضادة، وينصح اتباع تعليمات الشركة الصانعة حيثما كان ذلك ممكنا، في حين أن تحقيقات الجيش الملكي النيبالي، ونحن ننصح استشارة الكتابات المنشورة.
الصورة = "jove_content"> في هذه الدراسة، استخدمنا التحقيق RNA الذي يكشف على وجه التحديد ما قبل مرنا من الجينات على مدار الساعة Lfng. بسبب النقص النسبي في وفرة، والكشف عن Lfng قبل مرنا يتطلب فترة طويلة من الحضانة مع لجنة التحقيق في مزيج التهجين التي تحتوي على 5X سيترات الصوديوم المالحة (SSC) للكشف عن إشارة جيدة. قد تنطبق نفس هذه الشروط لتحقيقات الأخرى التي كشف من mRNAs أعرب ضعيفة، ولكن في تجربتنا، والكشف عن أهداف مرنا أكثر استقرارا قد يتطلب أقصر خطوة التحقيق التهجين وانخفاض تركيزات SSC في مزيج التهجين (على سبيل المثال، 1.3x SSC). لكل المناعية والأسماك، يجب أولا أن يكون الأمثل البروتوكول على الأجنة كلها، وتركيز الأمثل من الأجسام المضادة أو التحقيق يجب أن تحدد تجريبيا.
القيود المفروضة على تقنية
وكما ذكر أعلاه، فإن نجاح هذه التقنية يعتمد بشكل كبير على نوعية من البروتين وكشف مرنا. Wوقد أوجز ه عدة اقتراحات بشأن الكيفية التي يمكن بها تحسين البروتين وكشف مرنا، ولكن في حالة عدم وجود كشف ذات جودة عالية إشارة فلوري، لا توجد وسيلة التجربة يمكن المضي قدما. ويقتصر عدد من الأهداف البروتين التي يمكن تحليلها في كل عينة الأنسجة بقرار الطيفي للالمجهر متحد البؤر والحواتم من الأجسام المضادة المستخدمة. في هذه الدراسة، كنا قادرين على استخدام ما يصل إلى ثلاثة الحواتم للكشف عن البروتين جنبا إلى جنب مع وصمة عار الحمض النووي في كل عينة 12. هذا البروتوكول يسمح فقط للكشف عن الهدف مرنا واحد، على الرغم من أن الطرق البديلة الحالية يمكن أن تستخدم لزيادة هذه النسبة إلى ما يصل إلى ثلاثة أهداف (14).
أهمية تقنية مع الاحترام لالحالية / الطرق البديلة
الطريقة الموصوفة هنا يوفر تقنية حساسة للكشف عن تقلبات البروتين على مستوى منخفض في إإكسبلنتس PSM-جبل بأكمله. القياس الكمي لهذه الديناميات هوممكن عن طريق أداء FISH لجين على مدار الساعة المعروفة في إإكسبلنتس المقابل المقابلة. يتم إنشاء مكتبة kymographs التي يمكن تنظيمها خلال دورة على مدار الساعة تجزئة واحدة، وتسليط الضوء على ديناميات التعبير الزمانية المكانية من هدف المصالح داخل هذا الإطار الزمني. والفرق الرئيسي في هذه التقنية مقارنة مع الآخرين هو استخدام الأتمتة الحسابي لأجل زمنيا مجموعات كبيرة من البيانات، التي تسمح للديناميات التعبير الزمانية المكانية من مكونات ساعة جديدة ليتم تحليلها بطريقة غير متحيزة. على سبيل المثال، هذه التقنية توفر نظرة ثاقبة كيف DLL1 وNotch1 البروتينات والتذبذبات هم شارك في تنظيمها عبر PSM بأكمله. وقد اعتمدت أساليب بديلة في هذا السياق أيضا على المناعية، لكنها لم يكشف عن تقلبات صغيرة في DLL1 وNotch1 مستويات البروتين في PSM الذيلية التي كانت واضحة باستخدام هذا الأسلوب. بدلا من ذلك، وذكرت أنها الانحدار مستمر من التعبير الذي هو الأقوى في المنطقة منقاري 9 ، 10، 11. وهذا يمكن أن يكون راجعا إلى حقيقة أن هذا البروتوكول لديه الأجسام المضادة الأولية فترة أطول الحضانة (3-5 أيام، في مقابل ليلة وضحاها)، والتي قد تكون مطلوبة للكشف عن مستويات منخفضة من البروتين. وذلك لأن مستويات DLL1 والتعبير Notch1 مرتفعة نسبيا في PSM منقاري، وهذا قد أثرت على الكتاب لصورة ان عينات في أقل من التعرض تحديد من سيكون ضروريا للكشف عن بروتين تعبير الذيلية. ينشأ واحد مزيد من التناقض المحتمل من استخدام الأنسجة غير المثبتة في الدراسة التي تشابمان وآخرون. ، الذي التعبير مؤقتة من DLL1 وNotch1 في PSM الذيلية قد تم أقل جيدا الحفاظ 9.
التطبيقات المستقبلية أو الاتجاهات بعد اتقان تقنية
مرة واحدة وقد أتقن هذا البروتوكول، وتحليل التعبير عالية الإنتاجية لا يمكن أن يؤديها أي بروتين من الفائدة في إدارة المشتريات والتوريدات.إإكسبلنتس PSM الناتجة عن عدة ليترات الماوس يمكن معالجتها في وقت واحد لتوليد عدد العينات عالية الضروري لتحليلها. على الرغم من أننا قد استخدمت فقط البرية من نوع الأجنة في هذه الدراسات، فمن الممكن لإجراء هذا التحليل باستخدام أجنة المعدلة وراثيا من أجل تقييم أهمية العوامل واحد أو أكثر على ديناميكية تعبير البروتين. ما وراء PSM، هذا البروتوكول يمكن تكييفها لغيرها من النظم التي تتكون من نصفين المقابل، ويمكن استخدامها للكشف عن حساسية على مستوى منخفض تعبير البروتين ومتذبذبة ديناميات. ومن الأمثلة التي يمكن تكييفها هذا البروتوكول هو دراسة تعبير البروتين الحيوي في الأنبوب العصبي الماوس، منذ نصفين المقابل يمكن توليدها وتربيتها، ولقد ثبت النشاط الشق على حد سواء في الحاضر والمهم بالنسبة الزخرفة 15. ونحن نشجع مجموعات أخرى للتكيف مع هذا البروتوكول إلى أنظمة أخرى وتقديم التغذية الراجعة للتحسين في المستقبل.
Disclosures
The authors have nothing to disclose.
Acknowledgements
وأيد هذا العمل من قبل دراسية لجنة نهر الميكونج إلى RAB، وهو دراسية لجنة نهر الميكونج إلى CSLB، ومنحة مشروع WT إلى JKD (WT089357MA). وأيد العمل أيضا على جائزة الثقة الاستراتيجية الترحيب (097945 / Z / 11 / Z). نشكر الدكتور E. Kremmer للهدية نوع من الأجسام المضادة DLL1 والدكتور سين Pourquie للتحقيق Lfng الحمض النووي الريبي.
Materials
| Name | Company | Catalog Number | Comments |
| DMEM-F12 | Gibco (ThermoFisher Scientific) | 11320033 | |
| GlutaMAX™-1 (100x) | Gibco (ThermoFisher Scientific) | 35050 | |
| Fetal Bovine Serum, qualified, E.U.-approved, South America origin | Gibco (ThermoFisher Scientific) | 10270106 | |
| Recombinant Human FGF-basic (154 a.a.) | Peprotech | 100-18B | |
| Penicillin/Streptomycin | Gibco (ThermoFisher Scientific) | 15140122 | |
| anti-mouse monoclonal Notch1 antibody^ | BD Pharmingen | 552466 | |
| anti-rat polyclonal Dll1 antibody^* | N/A | N/A | |
| Lfng intronic anti-sense RNA probe^* | N/A | N/A | |
| 16% paraformaldehyde | Pierce (ThermoFischer Scientific) | PI28908 | |
| Proteinase K, recombinant, PCR grade | Roche (Sigma-Aldrich) | 31158 | |
| Phosphate buffered saline (PBS), pH 7.4 | Made in house | N/A | |
| Triton-X 100 | Sigma-Aldrich | T8787 | |
| Bovine serum albumin (BSA) | Sigma-Aldrich | 5470 | |
| Normal goat serum (NGS) (heat-treated) | Gibco (ThermoFisher Scientific) | 16210072 | |
| Hoechst 33342 | ThermoFischer Scientific | H3570 | |
| Tween-20 | Sigma-Aldrich | P9416 | |
| Ethanol | Sigma-Aldrich | 46139 | |
| Glutaraldehyde | Sigma-Aldrich | 340855 | |
| Formamide | Sigma-Aldrich | F9037 | |
| Saline-sodium citrate (SSC) | Sigma-Aldrich | 93017 | |
| EDTA | Sigma-Aldrich | 798681 | |
| tRNA | Roche (Sigma-Aldrich) | 101095 | |
| Heparin | Sigma-Aldrich | H3149 | |
| Tris-buffered saline (TBS) | Made in house | N/A | |
| Blocking Buffer Reagent | Roche (Sigma-Aldrich) | 11096176001 | |
| anti-DIG horseradish peroxidase (HRP) conjugated antibody | Roche (Sigma-Aldrich) | 11207733910 | |
| Tyramide signal amplification (TSA) kit | Perkin Elmer | NEL744001KT | |
| *The Dll1 antibody and RNA probe used in this study are not commercially available. Please see acknowledgements for sources. | |||
| ^Antibodies/RNA probes should be sourced which are applicable to the research interests of the reader. | |||
References
- Oates, A. C., Morelli, L. G., Ares, S. Patterning embryos with oscillations: structure, function and dynamics of the vertebrate segmentation clock. Development. 139 (4), 625-639 (2012).
- Krol, A. J., Roellig, D., et al. Evolutionary plasticity of segmentation clock networks. Development. 138 (13), 2783-2792 (2011).
- Dequéant, M. -. L., Ahnert, S., et al. Comparison of Pattern Detection Methods in Microarray Time Series of the Segmentation Clock. PLoS ONE. 3 (8), 2856 (2008).
- Bailey, C., Dale, K. . Somitogenesis in Vertebrate Development. , 1-15 (2015).
- Maroto, M., Bone, R. A., Somitogenesis Dale, J. K. Somitogenesis. Development. 139 (14), 2453-2456 (2012).
- Kageyama, R., Masamizu, Y., Niwa, Y. Oscillator mechanism of notch pathway in the segmentation clock. Developmental Dynamics. 236 (6), 1403-1409 (2007).
- Ferjentsik, Z., Hayashi, S., et al. Notch Is a Critical Component of the Mouse Somitogenesis Oscillator and Is Essential for the Formation of the Somites. PLoS Genetics. 5 (9), 1000662 (2009).
- Wiedermann, G., Bone, R. A., Silva, J. C., Bjorklund, M., Murray, P. J., Dale, J. K. A balance of positive and negative regulators determines the pace of the segmentation clock. eLife. 4, 05842 (2015).
- Chapman, G., Sparrow, D. B., Kremmer, E., Dunwoodie, S. L. Notch inhibition by the ligand DELTA-LIKE 3 defines the mechanism of abnormal vertebral segmentation in spondylocostal dysostosis. Human Molecular Genetics. 20 (5), 905-916 (2011).
- Sparrow, D. B., Chapman, G., et al. A Mechanism for Gene-Environment Interaction in the Etiology of Congenital Scoliosis. Cell. 149 (2), 295-306 (2012).
- Okubo, Y., Sugawara, T., Abe-Koduka, N., Kanno, J., Kimura, A., Saga, Y. Lfng regulates the synchronized oscillation of the mouse segmentation clock via trans-repression of Notch signalling. Nature communications. 3, 1141 (2012).
- Bone, R. A., Bailey, C. S. L., et al. Spatiotemporal oscillations of Notch1, Dll1 and NICD are coordinated across the mouse PSM. Development. 141 (24), 4806-4816 (2014).
- Shea, K., Geijsen, N. Dissection of 6.5 dpc mouse embryos. Journal of Visualized Experiments. (2), 160 (2007).
- Denkers, N., García-Villalba, P., Rodesch, C. K., Nielson, K. R., Mauch, T. J. FISHing for chick genes: Triple-label whole-mount fluorescence in situ hybridization detects simultaneous and overlapping gene expression in avian embryos. Developmental Dynamics. 229 (3), 651-657 (2004).
- Stasiulewicz, M., Gray, S. D., et al. A conserved role for Notch signaling in priming the cellular response to Shh through ciliary localisation of the key Shh transducer Smo. Development. 142 (13), 2291-2303 (2015).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved