Method Article
تشريح العضلات الطائرة الصباغية لنهج الomics
In This Article
Summary
تعتبر عضلات الرحلات الجوية نموذجا قويا لدراسة التنظيم العابر ، والربط البديل ، والأيض ، والبيولوجيا الميكانيكية. نقدم بروتوكولا لتشريح العضلات الطائرة الفلورية المسمية من العرائس الحية لتوليد عينات عاليه التخصيب مثاليه للبروميات والتسلسل العميق. هذه العينات يمكن ان تقدم رؤى ميكانيكيه هامه في جوانب مختلفه من تنميه العضلات.
Abstract
تعتبر عضلات الرحلات الجوية نموذجا قويا لدراسة العمليات المتنوعة مثل التنظيم المتحول ، والربط البديل ، والأيض ، والبيولوجيا الميكانيكية ، والتي تؤثر جميعها علي تنميه العضلات والميوفيبريلوجينيسيس. ويمكن لبيانات الomics ، مثل تلك المتولدة عن قياس الطيف الكتلي أو التسلسل العميق ، ان توفر رؤى ميكانيكيه هامه في هذه العمليات البيولوجية. النسبة لمثل هذه النهج ، من المفيد تحليل العينات الخاصة بالانسجه لزيادة الانتقائية والخصوصية لبصمات الأصابع. هنا نقدم بروتوكول لتشريح الفلورسنت المسمي العضلات الطيران من عذارى الحية لتوليد عينات العضلات المخصب للغاية للتطبيقات اوميكس. نحن أولا وصف كيفيه تشريح عضلات الطيران في مراحل الخوادر في وقت مبكر (< 48 ح بعد تشكيل puparium [منتديات البيئة الاتحادية]) ، عندما تكون العضلات التي يمكن التعرف عليها من قبل البروتين الفلوري الأخضر (gfp) وضع العلامات. ثم نقوم بوصف كيفيه تشريح العضلات من الدمى المتاخره (> 48 ح المنتدى) أو البالغين ، عندما يتم تمييز العضلات تحت المجهر تشريح. سيجعل البروتوكول المصاحب للفيديو هذه الأجزاء الصعبة من الناحية الفنية متاحه علي نطاق أوسع لمجتمعات البحوث العضلية والبحثية. للتطبيقات RNA ، ونحن فحص كميه ونوعيه RNA التي يمكن ان تكون معزولة في نقاط زمنيه مختلفه ومع نهج مختلفه. ونعرض كذلك ان Bruno1 (Bru1) ضروري للتحول الزمني في ميوسين سلسله ثقيله (Mhc) الربط ، مما يدل علي ان العضلات تشريح يمكن استخدامها ل mrna-Seq ، الطيف الكتلي ، وعكس النسخ العكسي البلمره سلسله التفاعل (RT-PCR) التطبيقات. هذا البروتوكول تشريح سوف تساعد علي تعزيز التحليلات اوميكس الانسجه المحددة ويمكن تطبيقها عموما لدراسة الجوانب البيولوجية متعددة من التولد.
Introduction
توفر تقنيات اوميكس الحديثة رؤى هامه في تنميه العضلات واليات الكامنة وراء اضطرابات العضلات البشرية. علي سبيل المثال ، كشف تحليل بيانات النسخ المدمجة مع التحقق الوراثي والبيوكيميائي في النماذج الحيوانية ان فقدان عامل الربط RBM20 يسبب اعتلال عضله القلب المتوسعة بسبب تنظيمها لشبكه مستهدفه من أكثر من 30 ساركوميري الجينات المرتبطة سابقا بامراض القلب ، بما في ذلك التيتين1،2،3.
في المثال الثاني ، أظهرت الدراسات التي أجريت من ثقافة الخلية ، والنماذج الحيوانية ، والمرضي البشريين ان ضمور الخلايا العضلية يسببه اضطراب في تنظيم الجيش النيبالي الريبي بسبب تنحيه المسك الأعمى (mbnl) والحامل لCELF14،5. تساعد الديناميكيات المتعددة الانظمه والزمانيه بين MBNL و CELF1 (التي تسمي أيضا CUGBP1 أو برونو-Like 2) علي شرح أنماط الربط الجنينية المستمرة في مرضي الضمور العضلي. بالاضافه إلى ذلك ، فان الشبكة الكبيرة من الأهداف المقننة تساعد علي شرح الطبيعة المعقدة للمرض4،6،7،8. غالبيه هذه الدراسات تستخدم نهج اوميكس في الكائنات الحية النموذجية الوراثية لفهم أليات الكامنة وراء مرض العضلات البشرية. وعلاوة علي ذلك ، فانها تسلط الضوء علي اهميه أول فهم الزمنيه والانسجه من نوع التعبير الجيني محدده ، وتعديل البروتين ، وأنماط الأيض في العضلات السليمة لفهم التغييرات في العضلات المريضة أو الشيخوخة.
دروفيا ميلانوجاستر هو كائن آخر النموذج الجيني الراسخة. هيكل ساركوميري وكذلك مكونات ساركوميري الفردية يتم الحفاظ عليها بشكل كبير من الذباب إلى الفقاريات4،9،10، وعضلات الطيران غير المباشرة (ifms) أصبحت نموذجا قويا للدراسة جوانب متعددة من تنميه العضلات11,12. أولا, عضلات الطيران لييفي وظيفيا ومتميزة شكليا من عضلات الجسم أنبوبي11,13, السماح بالتحقيق في العضلات من نوع أليات التنموية المحددة. وقد تم تحديد عوامل النسخ بما في ذلك Spalt الرئيسية (سالم)14، اكسترادينتيكلي (exd) ، والصدرية (hth)15 والمنظمين مصير لييفي. بالاضافه إلى ذلك, المصب من سالم, CELF1 homolog Bruno1 (Bru1, أريت) يوجه برنامج الربط لييفي محدده16,17.
ثانيا ، ifms هي نموذج مهم لفهم عمليه التولد نفسها ، من الانصهار استدامه والتعلق myotube إلى ميوفيبريلوجينيسيس ونضوج ساركوميري9،18،19. ثالثا ، تسمح الوراثة الوراثية بالتحقيق في المساهمات التي تقدمها البروتينات الفردية ، ومجالات البروتين ، والبروتينات البروتينية إلى تشكيل ساركوميري ، والوظيفة ، والخصائص الفيزيائية الحيوية20،21،22 ،23. أخيرا, وقد وضعت نماذج ifm لدراسة اضطرابات العضلات البشرية متعددة, مثل ضمور عضلي, ميوفيبريلار ميات, اضطرابات العضلات التنكسية, الاكتيباثيس, الخ.24,25,26 ،27، وقدمت رؤى هامه في أليات المرض والعلاجات المحتملة28،29،30. وهكذا ، دروفيبيلا هو نموذج مفيد لمعالجه العديد من الاسئله المفتوحة في مجال التولد ، بما في ذلك أليات من نوع العضلات النسخ المحددة ، والربط ، وتنظيم الكروماتين ، فضلا عن دور الأيض في تنميه العضلات. تطبيق تقنيات الاوميكس الحديثة ، وخاصه بالاقتران مع مجموعه واسعه من الاختبارات البيولوجية الوراثية والبيوكيميائية والخلوية المتاحة في دروفيا، لديه القدرة علي تعزيز فهم العضلات بشكل كبير التنمية ، والشيخوخة ، والمرض.
Ifms هي أكبر العضلات في ذبابه, تمتد ما يقرب من 1 ملم عبر طول الصدر في البالغين31,32. ومع ذلك ، فان هذا الحجم الصغير يولد التحدي المتمثل في الحصول علي عينه كافيه لتطبيق تقنيات الاوميكس في دروفيا بطريقه محدده من نوع الانسجه. وعلاوة علي ذلك ، ifms هي جزء من العضلات الكبار التي تتشكل خلال مراحل الخوادر. الصمامات المنصهرة لتشكيل myotubes ، والتي تعلق علي الأوتار حوالي 24 ساعة بعد تشكيل puparium (منتدى المحيط الهادي) والخضوع لخطوه الضغط اللازمة لبدء ميوفيبريلوجينيسيس حول 30 ح المنتدى الاتحادي العام (الشكل 1ا-د)18،33، 34.
ألياف العضلية ثم تنمو لتمتد علي طول الصدر ، مع myoالفايلز تمر مرحله النمو الاوليه التي تركز علي ساركوميري بالاضافه إلى حوالي 48 h منتديات التنمية الاتحادية ، ومن ثم الانتقال إلى مرحله النضوج ، التي ساركوميري تنمو في الطول والعرض وهي تشكيلها لإنشاء امتداد التنشيط بواسطة 72 h المنتدى (الشكل 1ا-د)32،35. بداية نضوج ألياف يتم التحكم فيها جزئيا علي الأقل من قبل سالم و E2F32،36،37، ومتعددة ifm المحددة البروتين ساركومير التي يتم التحكم الربط من قبل Bru1 وأدرجت خلال هذا مرحله16،17. الذباب الناضجة eclose من 90-100 ح منتدى الدعوة. وهذا يعني ان لدراسة تنميه العضلات, ifm يجب ان تكون معزولة مع كميه كافيه, الجودة, والنقاء من النقاط الزمنيه الخوادر متعددة لتسهيل التحليل باستخدام نهج اوميكس.
وقد تم نشر العديد من البروتوكولات لتشريح IFM. في حين ان هذه البروتوكولات تعمل بشكل جيد للتطبيقات المقصودة ، لا شيء مثالي للنهج اوميكس. البروتوكولات التي تحافظ علي المورفولوجية ifm للمناعة من الخوادر والكبار ifm19، عزل ألياف ifm للتقييم الميكانيكي31، أو الاستفادة من التشريح المجهري لل ifm من الحلوى38 هي متخصصة جدا والوقت و العمالة المكثفة للحصول علي معقول كميات كافيه من الانسجه ifm للتطبيقات اوميكس. وقد وضعت بروتوكولات أخرى لتشريح السريعل38البالغ ifm علي وجه التحديد،39، التالي لا تنطبق علي مراحل الخوادر ، واستخدام المخازن المؤقتة التي ليست مثاليه أو قد تكون غير متوافقة مع ، علي سبيل المثال ، العزل RNA. التالي ، هناك حاجه لتطوير نهج جديده لعزل الخوادر ifm للتطبيقات الكيمياء الحيوية أو اوميكس.
هنا نقدم بروتوكولا لتشريح ifm خلال مراحل الخوادر التي تم استخدامها بنجاح للتحليل mrna-Seq من 16 ح المنتدى من خلال مراحل الكبار16,32. يستخدم البروتوكول تسميه البروتين الفلوري الأخضر (GFP) للتعرف علي IFMs في جميع مراحل النمو العفن والبالغ ، مما يسمح بالتشريح الحي تحت مجهر تشريح الفلورسنت. النهج هو اقل كثافة العمالة ، مع إنتاجيه اعلي من بروتوكولات تشريح IFM القائمة. وهذا يسمح العزلة السريعة والتبريد من العينات ، وتوليد ما يكفي من المواد بعد عده جولات من تشريح للنهج اوميكس ، فضلا عن النسخ العكسي القياسية تفاعل البلمره سلسله (RT-PCR) أو الغربية التنقيط.
نحن نقدم البروتوكول في جزاين ، مما يدل علي كيفيه تشريح بسرعة ifms قبل 48 h منتديات الدعوة (خلال التحول في وقت مبكر ، عندما ifms المرفقات هي أكثر هشاشة) وبعد 48 h منتديات الدعوة (عندما خطه الجسم الخوادر و ifms المرفقات محدده جيدا). ونحن نثبت اننا يمكن عزل الحمض الريبي النيبالي عاليه الجودة من تشريح IFMs في جميع النقاط الزمنيه وتقديم البيانات علي نهج مختلفه لعزل الجيش النيبالي الريبي والنسخ العكسي. وأخيرا ، فاننا نظهر تطبيق بروتوكول تشريح إلى mRNA-Seq ، الطيف الكتلي ، و RT-PCR باستخدام CELF1 homolog Bruno1 كمثال. نحن نظهر سوء التعبير عن البروتين ساركوميري في البيانات البروتينية من Bruno1 متحولة IFM ودراسة Bruno1 تنظيم الحدث C-الطرفية لصق من سلسله الثقيلة ميوزين (Mhc). وتوضح هذه النتائج كيف يمكن لبيانات الاوميكس ان توفر فهم أعمق للظواهر البيولوجية ، بما يكمل التجارب الوراثية والبيوكيميائية.
Protocol
1. تجهيز العرائس
- رفع الذباب من النمط الجيني المطلوب في زجاجات (الشكل 1ه). اما جعل الوجه الطازجة من المخزون تشريح أو تعيين الصليب مع ما لا يقل عن 20 الذباب العذراء الإناث. الحفاظ علي الزجاجات حتى تبدا الذباب إلى الدمى.
- جمع ما قبل pupae مع فرشاه الطلاء المبللة ونقل إلى ورقه فلتر مبلله في صحن بيتري مم 60 (الشكل 1و).
- الجنس والعرائس ، وجمع الجنس المناسب للتجربة (الشكل 1ز). يتم تحديد الذكور من قبل وجود الخصيتين ، والتي تظهر ككرات شفافة في pupa مبهمه خلاف ذلك.
- تسميه طبق بيتري مع الوقت والتاريخ ، والنمط الجيني ، ثم سن عذارى إلى المرحلة المطلوبة (الشكل 1ح).
ملاحظه: الحفاظ علي الصلبان/الأسهم والدمى العمرية في حاضنه التي تسيطر عليها درجه الحرارة (اي ، 25 درجه مئوية أو 27 درجه مئوية لصلبان RNAi ، وزيادة النشاط Gal4 في درجات حرارة اعلي يزيد من ضربه إلى أسفل كفاءه40). تاكد من ان الرطوبة عاليه بما فيه الكفاية حتى العرائس لا تجف عند الشيخوخة عده أيام.
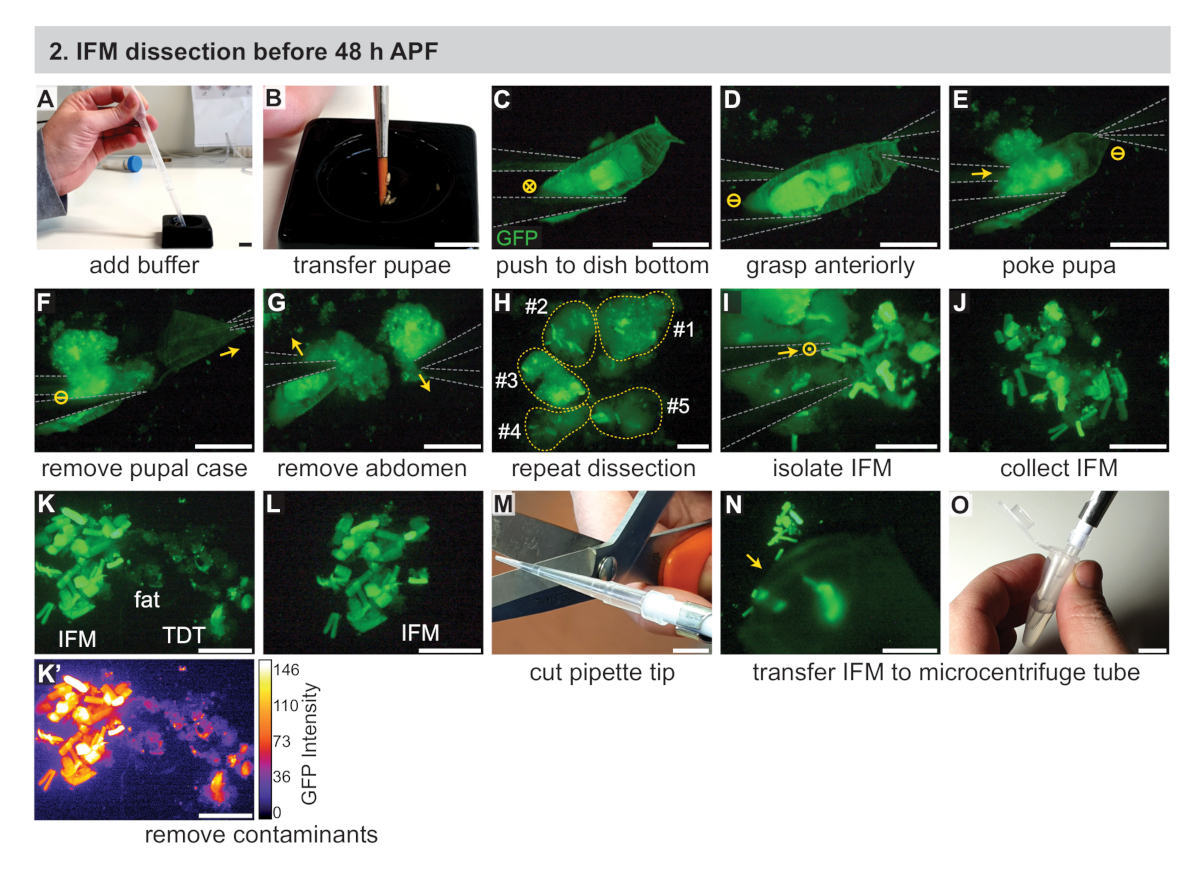
2. تشريح IFM قبل 48 h منتدى الدعوة الاتحادية
- تجميع المعدات اللازمة بما في ذلك اثنين من ال#5 البيولوجيا الصف ملقط ، ماصه ، ونصائح ماصه ، والجليد الجاف ، و (لعينات RNA) كاشف العزلة (انظر جدول المواد). الاضافه إلى ذلك ، البرد الأسود تشريح الاطباق (انظر جدول المواد) ، 1x الفوسفات-مخزنه المالحة (تلفزيوني) العازلة ، و 1.5 mL أنابيب الطرد المركزي الصغرى علي الجليد.
- باستخدام فرشاه الطلاء المبللة ، نقل العرائس المقسمة إلى صحن التشريح الأسود شغل حوالي الثلثين مع الباردة 1x تلفزيوني (الشكل 2ا ، ب). الانتقال إلى المجهر تشريح الفلورسنت.
ملاحظه: استخدام العديد من عذارى كما يمكن تشريحها في غضون 30 دقيقه نافذه الوقت. اعتمادا علي الخبرة ، وهذا يتراوح بين 3 و 15 pupae. انظر الأساليب التكميلية لمناقشه بدائل الاطباق السوداء التشريح. - باستخدام ملقط #5 ، دفع واحده من العرائس إلى الجزء السفلي من طبق تشريح السوداء وضبط التكبير المجهر والتركيز علي رؤية بوضوح pupa (الشكل 2ج).
- فهم الامامي من pupa مع ملقط واحد (الشكل 2د) ، ثم كزه العرائس مع طرف واحد من ملقط الأخرى قليلا خارج المركز في البطن ، خلف الصدر مباشره. هذا يحمل pupa في مكان ويمنع IFMs من الانتقال إلى البطن (الشكل 2ه).
ملاحظه: بدء توقيت طول تشريح من هذه النقطة ، حالما يتم تعطيل سلامه الخوادر. استخدام طول محدد من تشريح (علي سبيل المثال 20-30 دقيقه) للحد من وفاه العضلات والتغييرات المصاحبة الناسخة والبروتينية. تشريح أكبر عدد ممكن من الذباب في هذه الفترة من الزمن. - باستخدام الملقط الأول ، قم بازاله النصف الامامي من الحالة الخوادر (الشكل 2و).
- استخدم نفس الملقط لقرصه الدمى المكشوفة خلف الصدر مباشره ، وافصل البطن عن الصدر (الشكل 2ز).
- باستخدام ملقط ، ضغط برفق الجزء الامامي من الصدر (ل < 35 ح المنتدى) أو مزق فتح الصدر لفضح فلوريسسينتلي المسمي IFMs (الشكل 2ح). سيتم فصل IFMs بسهوله من البشرة ، حيث ان مرفقات الأوتار في النقاط الزمنيه المبكرة هشه. تجاهل الذبيحة المتبقية باستخدام ملقط لدفعها إلى الجانب الآخر من الطبق.
- الخطوات المكررة 2.3 – 2.7 ، تشريح العرائس الاضافيه.
- جمع ألياف IFM مع ملقط وتنظيمها في كومه في الجزء السفلي من الطبق الأسود تشريح (الشكل 2I, J). أزاله اي الحطام عن طريق دفعها للخروج من مجال الرؤية باستخدام ملقط.
ملاحظه: مع الممارسة ، يمكن جلب نصائح ملقط إلى قرب وثيق دون لمس بعضها البعض. ويمكن استخدام هذه التقنية لانتزاع بشكل فضفاض IFMs دون تدميرها. وتشمل الأساليب البديلة بلطف دفع أو رفع IFMs مع طرف واحد أو ملقط مغلقه تماما ، أو أخذ بعض الدهون أو غيرها من الانسجه مع IFMS وأزاله الدهون كما هو موضح في الخطوة 2.10. - مراقبه الجودة عينه العضلات IFM ، وذلك باستخدام ملقط لأزاله العضلات غير IFM ، والدهون ، واهاب ، وما إلى ذلك من العينة (الشكل 2K ، L).
ملاحظه: مع Mef2-Gal4, IFM هو المسمي أكثر قوه من أنواع العضلات الأخرى في وقت مبكر (الشكل 2k, k '), السماح بازاله العضلات القفزة وعضلات اليرقات علي أساس كثافة الفلورية وشكل العضلات. الدهون وبشره الانسجه تبدو مختلفه وليست المسمية بواسطة التسمية الفلورية الخاصة بالعضلات (الشكل 2k, k '). راجع قسم المناقشة للخطوط Gal4 الأخرى التي تسمي IFM. - باستخدام طرف ماصه قص ، نقل كومه من IFMs إلى 1.5 mL أنبوب الطرد المركزي الصغير مليئه 250 μL من المبردة 1x تلفزيوني (الشكل 2م-O). انتقل فورا إلى القسم 4.
ملاحظه: قد تضيع عينات IFM ببساطه عن طريق التصاق بجانب طرف الماصة. يمكن ان يجعل العازل المؤقت لاعلي ولأسفل عده مرات قبل جمع IFMs نصائح قياسيه اقل التصاقا ، ويمكن ان تساعد نصائح بيرفلوروالكوكسي (PFA) (انظر جدول المواد) ذات التوترات السطحية المنخفضة في منع فقدان العينة.
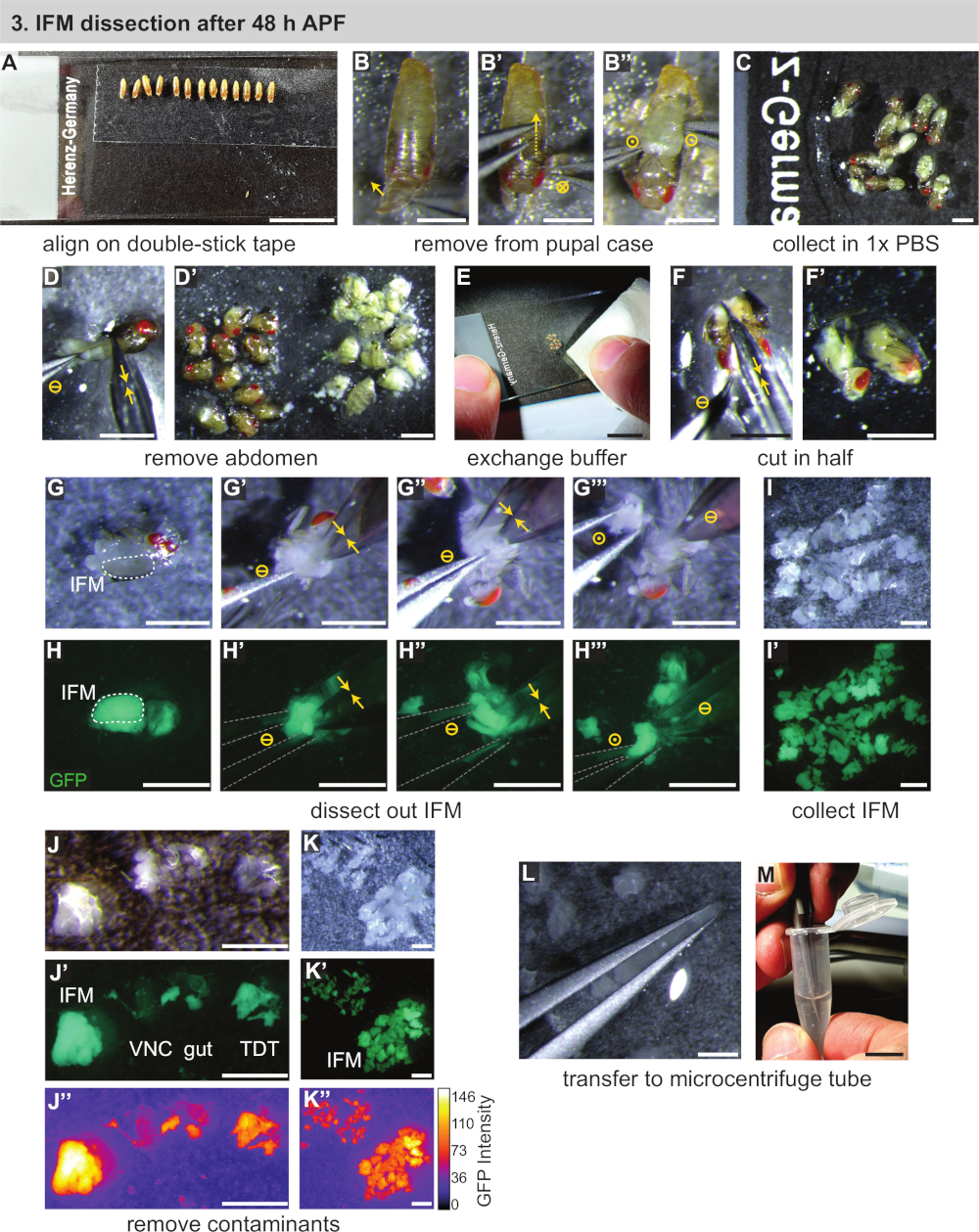
3. تشريح IFM بعد 48 h منتدى الدعوة الاتحادية
- تجميع المعدات اللازمة بما في ذلك اثنين من ال#5 البيولوجيا الصف ملقط ، مقص غرامه ، والشرائح المجهر الزجاج القياسية ، مزدوجة عصا الشريط ، ماصه ، ونصائح ماصه ، والجليد الجاف ، و (للتطبيقات RNA) العزل كاشف (انظر جدول المواد). البرد 1x تلفزيوني وأنابيب الطرد المركزي الصغير علي الجليد.
- باستخدام فرشاه الطلاء المبللة بخفه ، نقل الدمى المقسمة إلى شريط من الشريط اللاصق المزدوج علي الوجه المثبت علي شريحة المجهر (الشكل 3ا). وضع عذارى في خط المنحى في نفس الاتجاه (بطني أسفل والامامي نحو الجزء السفلي من الشريحة).
ملاحظه: يجب الحرص علي عدم استخدام الكثير من الماء علي فرشاه الطلاء أو فلتر ، أو العرائس لن عصا جيدا. إذا لم تلتصق العرائس ، جففيها بواسطة النقل الاولي إلى فلتر جاف أو مناديل ورقيه. جبل كما العديد من العرائس كما يمكن تشريحها في غضون 30 دقيقه نافذه الوقت ، من الناحية المثالية ~ 10 عذارى. - أزاله العانة من القضية pupa. استخدام ملقط لندف بعيدا وفتح القضية الخوادر فوق الحلزونيات الاماميه (الشكل 3ب).
- حرك برفق زوجا من الملقط الناتئ نحو الخلف ، وقطع الحالة بالاصبع كالملقط الحركة (الشكل 3ب '). يجب الحرص علي عدم تمزق pupa الكامنة. تحرير pupa من القضية فتح ونقلها علي الفور إلى قطره من 1x تلفزيوني علي شريحة المجهر الثاني (الشكل 3ب "، ج).
- كرر الخطوات 3.3 و 3.4 لكل عذارى في السطر ، ثم قم بتعيين شريحة الشريط المزدوج جانبا.
- باستخدام مقص غرامه ، وقطع البطن من pupa بعيدا عن الصدر ودفعها إلى كومه منفصلة (الشكل 3d ، d '). كرر لما تبقي من pupae.
ملاحظه: بدء توقيت طول تشريح مع الخطوة 3.6 ، بمجرد ان يتم تعطيل سلامه الخوادر. تشريح أكبر عدد ممكن من الذباب في 20 – 30 دقيقه لمنع موت الخلايا والتغييرات المرتبطة بالناسخ والبروتيني. عند تشريح 1 د البالغين أو > 90 ح pupae ، فانه غالبا ما تكون مريحه للخطوات في وقت لاحق لأزاله الراس بالاضافه إلى ذلك مع مقص غرامه. - باستخدام ورقه المناديل الورقية ، وأزاله غالبيه 1x تلفزيوني (غائم عموما مع الدهون المعلقة) ، فضلا عن كومه من بطون (الشكل 3ه). أضافه قطره من الطازجة ، والمبردة 1x تلفزيوني إلى الthoraxes المتبقية.
- استخدام مقص لقطع الصدر في نصف (الشكل 3f ، f ') عن طريق قطع من الراس إلى أسفل محور الجسم الطولي في حركه واحده. بالتناوب ، إذا تمت أزاله الراس ، أولا ادراج المقص حيث تم إرفاق الراس وقطع النصف العلوي من الصدر طوليا بين IFMs. ثم ، قطع الجانب البطني من الصدر مع خفض الثاني في نفس الاتجاه.
- كرر الخطوات 3.7 و 3.8 لجميع عذارى ليتم تشريحها ، وتوليد كومه من الصدر هيميسيكتيونس بالقرب من وسط الشريحة. تاكد من ان هناك ما يكفي 1x المبردة تلفزيوني علي الشريحة بحيث هيميسيكتيونس لا تجف.
ملاحظه: بعد 48 h منتدى الدعوة الاتحادية ، IFMs كبيره بما يكفي لتكون مرئية تحت المجهر تشريح القياسية للعين المدربة. عند هذه النقطة في البروتوكول ، يمكن نقل العضلات مع تسميه الفلورسنت إلى نطاق تشريح الفلورسنت للمساعدة في تحديد IFM أو لأغراض التدريب ، ولكن هذا ليس ضروريا. - تشريح ال IFMs خارج الصدر. عزل أحد هيميسيكتيونس باستخدام ملقط ال#5 (الشكل 3G ، H). ادخل برفق النصائح الخاصة بملقط واحد فوق وتحت منتصف IFMs (الشكل 3G ' ، H '). في حين عقد ملقط الاولي لا يزال ، واستخدام مقص غرامه لقطع واحده من نهاية IFM بعيدا عن اهاب والأوتار. ثم ، قطع الطرف الآخر من IFM خاليه من اهاب (الشكل 3G '' ، H '').
ملاحظه: اعتمادا علي اتجاه الصدر بعد قطع IFM الاولي ، فمن المفيد لتدوير الصدر 180 درجه بحيث قطع IFM الثاني هو أسهل لأداء. - أزاله حزمه IFM من الصدر مع ملقط (الشكل 3ز '' ، H '''') ، نقله إلى حافه فقاعه تلفزيوني لاستخدام التوتر المياه لعقد في مكان (الشكل 3ط)... دفع الذبيحة إلى الجانب الآخر من الشريحة. كرر لهيميسيكشنز الصدرية المتبقية ، وتوليد مجموعه من الifms تشريح.
ملاحظه: إذا كان IFMs لا تبقي في كومه أنيق ، وأزاله بعض من 1x تلفزيوني مع الانسجه. يجب الحرص علي عدم السماح لجميع الإذاعات التلفزيونية تتبخر ، والتاكد من ان IFMs تشريح و hemithoraxes تبقي مغطاه العازلة. - بعد تشريح جميع IFMs ، بسرعة تنفيذ مراقبه الجودة علي العضلات تشريحها. باستخدام ملقط #5 ، وأزاله اي العضلات القفز أو الشظايا اهاب التي قد وجدت طريقها إلى العينة (الشكل 3J-K '''').
ملاحظه: تظهر العضلات القفز مختلفه من IFM. إذا تشريح Mef2-Gal4 المسمي العضلات تحت الفلوري, القفز العضلات لديه فلوري أضعف وشكل مختلف والملمس. تحت الضوء العادي ، فانه يبدو شبه شفافة في حين IFMs هي مبهمه ، حليبي الأصفر (الشكل 3j-j '''' ، K). - باستخدام التوتر المياه ، والتقاط (ولكن لا squish) IFMs تشريح بين زوج من الملقط (الشكل 3L). نقل IFMs إلى أنبوب الطرد المركزي الصغير 1.5 mL معباه مسبقا مع 250 μL من المبردة 1x تلفزيوني (الشكل 3م). المضي قدما فورا مع القسم 4.
ملاحظه: عندما يتم إحضارها نصائح ملقط في القرب من بعضها البعض ورفعها من حل العازلة ، يسبب التوتر المياه فقاعه من العازلة ليتم القبض بين النصائح ملقط. إذا كانت IFMs موجودة أيضا في هذه الفقاعة ، فانه يمكن رفعها من الحل ونقلها بسهوله إلى وعاء آخر مملوء بالمخزن المؤقت. من المهم ان تضغط علي ملقط لجلب النصائح بالقرب من بعضها البعض دون لمس بعضها البعض ، لتجنب ماشيتينغ الانسجه الملتقطة في فقاعه العازلة.
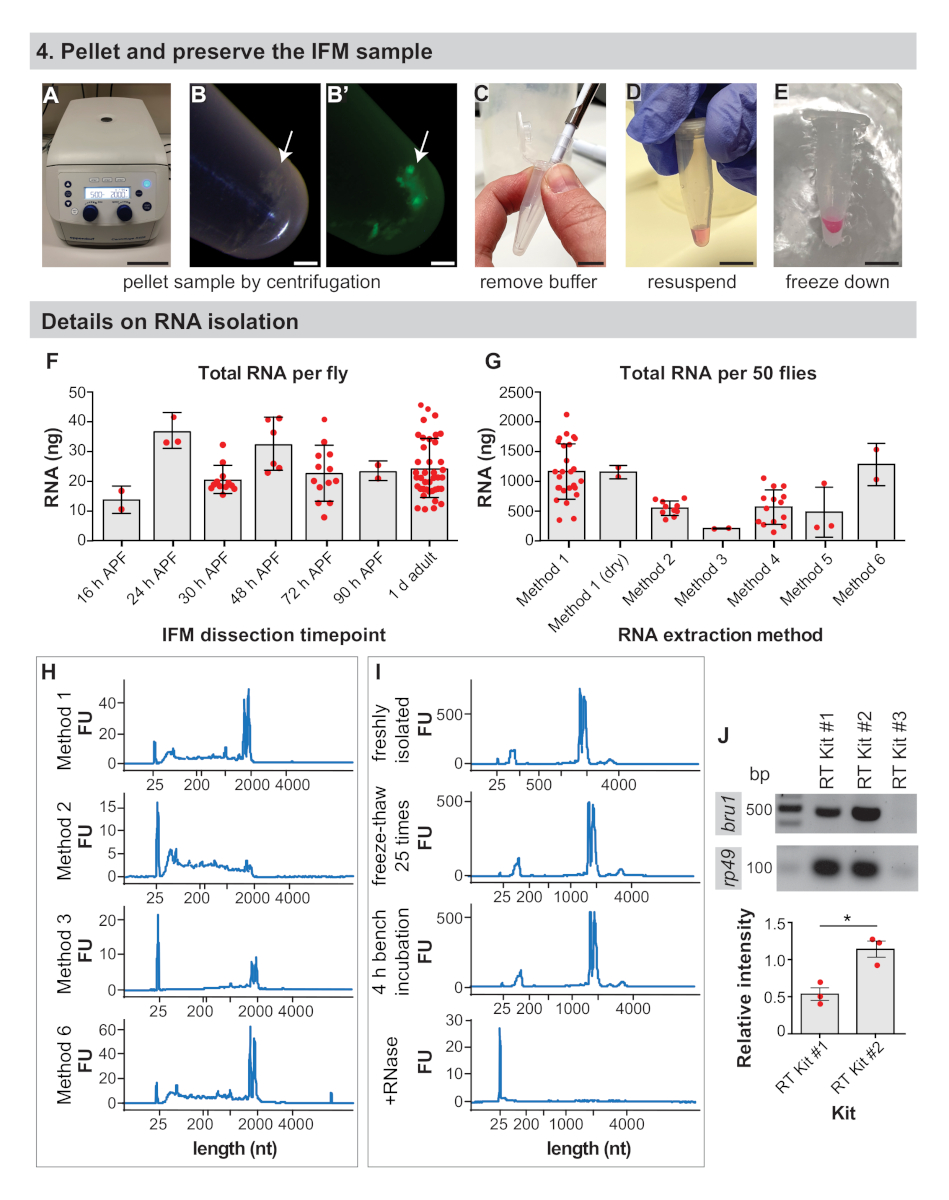
4. بيليه والحفاظ علي عينه IFM
- بيليه IFMs بواسطة يطرد 1.5 mL أنبوب الطرد المركزي لمده 3-5 دقيقه في 2,000 x g في جهاز الطرد المركزي الجدول الأعلى (الشكل 4ا ، ب).
- قم بازاله المخزن المؤقت باستخدام طرف ماصه (الشكل 4ج).
- للتطبيقات RNA ، أعاده تعليق بيليه IFM في 50-100 μL من المخزن المؤقت المطلوب العزل RNA (انظر جدول المواد، الشكل 4د). والا ، انتقل إلى الخطوة 4.4.
ملاحظه: IFMs يمكن ان تكون جافه-المجمدة بعد الخطوة 4.2 للتحضيرات قياس الطيف الكتلي أو عزل الحمض الريبي النيبالي مع مجموعات تجاريه (انظر النتائج التمثيلية). للتطبيقات RNA ، يتم الحصول علي نتائج أفضل عن طريق أعاده تعليق فورا وتجميد بيليه IFM في العازلة العزلة. - تجميد العينة علي الجليد الجاف أو تجميد المفاجئة في النيتروجين السائل (الشكل 4ه). يخزن عند-80 درجه مئوية حتى يصبح جاهزا للخطوات اللاحقة في اعداد العينة للتحليل النهائي.
ملاحظه: بعد الحجز بالتبريد ، يمكن تخزين العينات لعده أشهر قبل المعالجة للتحقيق النهائي.
النتائج
بروتوكولات التشريح المعروضة أعلاه مفيده لتوليد العينات المخصب IFM من 16 ح بعد تشكيل puparium (المنتدى) حتى مرحله الكبار. تشريح العينات العضلات الطيران يمكن استخدامها لتطبيقات متعددة ، وحتى الآن تم تطبيقها بنجاح ل RT-PCR4،17، RNA-Seq16،32، رقاقه36،37، الغربية التنقيط14،41 والتجارب الطيفية الكتلي (انظر أدناه). لمساعده المستخدمين المحتملين تشريح للتطبيقات المستندة إلى RNA ، ونحن أولا تقديم نتائجنا تسليط الضوء علي الاعتبارات الهامه علي وجه التحديد لعزل RNA من IFMs. لتوضيح أكثر علي نطاق واسع من فائده بروتوكولات التشريح لدينا ، ونحن ثم توضيح بعض التطبيقات المحتملة-omics باستخدام البيانات الخاصة بنا علي Bruno1 البروتين الحمض الريبي الملزم.
IFM التشريح البروتوكول غله عاليه الجودة RNA
من المهم تحديد عدد الذباب الذي سيتم تشريحه مسبقا ، حيث يقدر ان الترميز mRNA لا يشكل سوي 1 – 5% من إجماليال42RNA. حصلنا في المتوسط 24 ± 9 نانوغرام من الحمض الريبي النيبالي الإجمالي لكل ذبابه من IFM تشريحها من 1 د البالغين (الشكل 4F والشكل التكميلي 1a) ، مع زيادة غله عاده مع الخبرة. هذا العائد من الحمض الريبي النيبالي الإجمالي لكل ذبابه ثابته نسبيا ، وتذبذب حول 25 نانوغرام ل IFM تشريح في 16 ح منتدى المحيط الهادي ، 24 ح المنتدى ، 30 ح منتدى المحيط الهادي ، 48 ح منتدى المحيط الهادي ، 72 ح منتدياتالدفاع الجوي و 90 h (الشكل 4و ). تعكس هذه الملاحظات أيضا اي RNA معزولة عن تلويث الدهون والأوتار والقصبة الهوائية أو أنواع الخلايا الأخرى ، والتي قد تكون اعلي في عينات معزولة عن النقاط الزمنيه السابقة. وهكذا ، حصلنا علي > 1 ميكروغرام من الحمض الريبي النيبالي الإجمالي من IFM من 50 الذباب وتشريح عاده IFM من 100 − 150 الذباب لتوليد > 3 ميكروغرام من الحمض الريبي النيبالي الإجمالي للعينات الحمض الريبي النيبالي-Seq.
طريقه العزلة RNA يؤثر علي كميه ونوعيه الحمض الريبي الذي تم استرداده ، ونحن نشجع المستخدمين علي التحقق من صحة نهج العزلة. علي سبيل المثال ، في حين ان العزلة باستخدام الطريقة 1 تنتج في المتوسط 1143 ± 465 ng من إجمالي الحمض الريبي النيبالي من IFM من 50 1 د الذباب الكبار ، والعزلة مع مجموعات تجاريه مختلفه الغلة في اي مكان من 186 ± 8 نانوغرام إلى 1261 ± 355 ng من إجمالي RNA (الشكل 4ز و الشكل التكميلي 1 ج). الجيش النيبالي الريبي معزولة من مجموعات تجاريه عموما من نوعيه جيده (الشكل 4H والشكل التكميلي 1f) ، ولكن انخفاض المستردات تشير إلى ان الجيش النيبالي الريبي قد لا تكون بكفاءة التملص من الاعمده. ويمكن أيضا الكشف عن سلامه الحمض الريبي النيبالي عن طريق استخدام مجموعه كما فعلت في الطريقة 2 (الشكل 4H، المؤامرة الثانية) ، من المرجح بسبب العازلة الدستور والعلاجات الحرارية ، مما يؤدي إلى تجزئه شديده التي يمكن ان تؤثر علي التجارب المصب.
ومن المهم أيضا ان نلاحظ المناسبة تقنيه RNase خاليه عند عزل والتعامل مع عينات RNA. علي الرغم من ان دورات التجميد-ذوبان الجليد و 4 h حضانة درجه حرارة الغرفة لا تؤثر بشكل كبير علي ملامح سلامه الحمض الريبي النيبالي ، وحتى كميات صغيره من RNase تؤدي إلى تدهور الحمض الريبي النيبالي السريع (الشكل 4الأول والأساليب التكميلية). ولا يزال المستخدمون يشجعون علي العمل علي الجليد والحد من التجمد والذوبان لمنع التحلل المائي RNA والتجزؤ. لم يتم الكشف عن هذا هنا ولكن منع التلوث RNase باستخدام نصائح فلتر والمخازن المؤقتة المعالجة DEPC ضرورية للغاية.
وتؤثر كفاءه النسخ العكسي أيضا علي نجاح التطبيقات النهائية. حصلنا علي نتائج موثوق بها مع اثنين من ثلاث مجموعات RT التجارية التي اختبرناها ، والتي تضخيم العصابات RT-PCR قويه لجين ريبوسومال rp49 (الشكل 4ي). ومع ذلك ، قد تكون #2 RT Kit أكثر حساسية للكشف عن النسخ المنخفضة التي تم التعبير عنها ، كما حصلنا علي عصابات اقوي للبروتين الحمض الريبي الbru1 الربط لجميع البيولوجية الثلاثة ونسخ متماثلة (الشكل 4ياء). معا, هذه النتائج توضح ان الحمض الريبي النيبالي عاليه الجودة يمكن ان تكون معزولة من IFMs تشريح مع هذا الاجراء.
تشريح IFMs إنتاج جوده عاليه mRNA-Seq والبيانات البروتينية
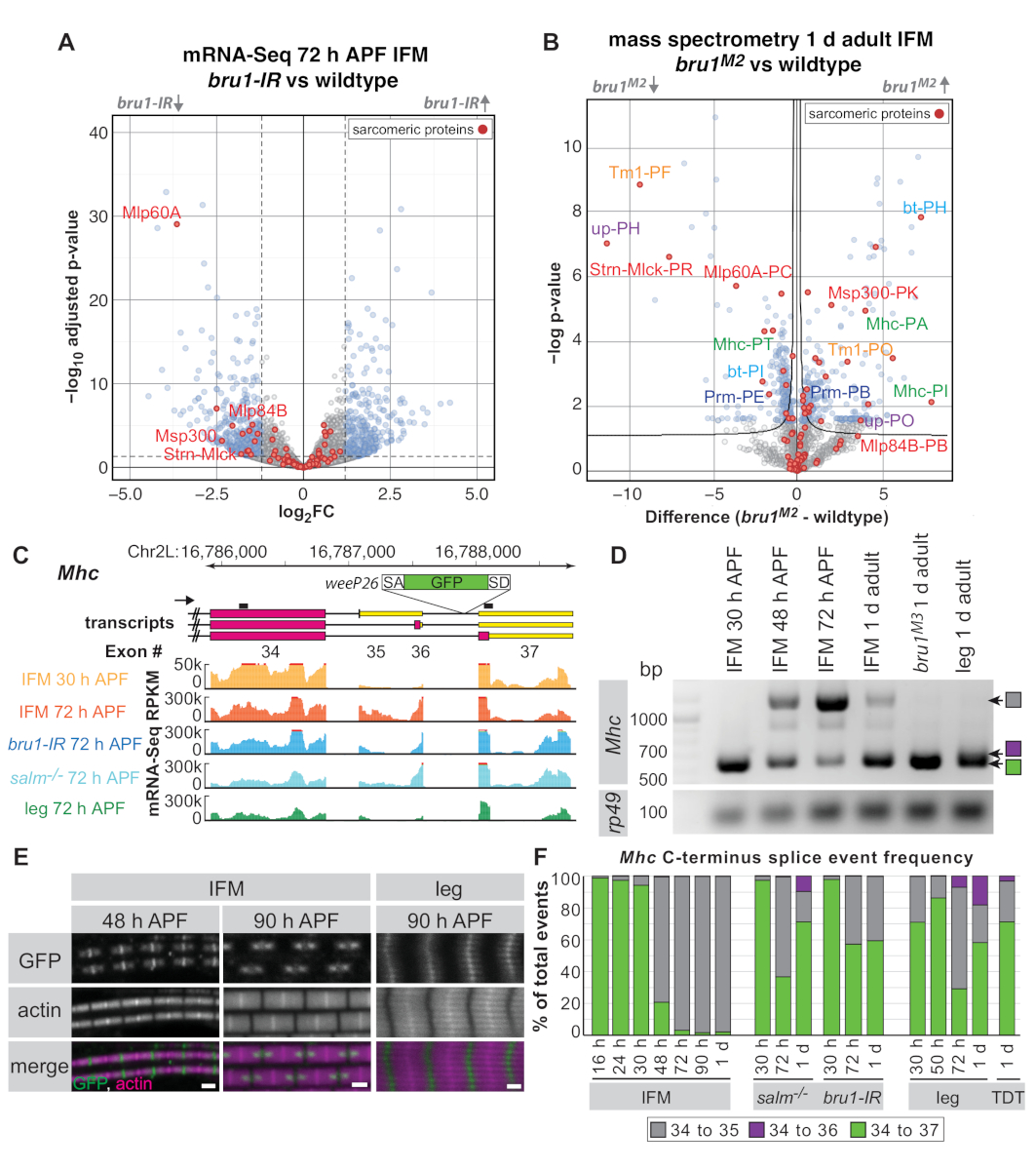
باستخدام IFM تشريح وفقا للبروتوكول أعلاه في 30 ح المنتدى المذكور ، 72 ح منتدى الدعوة والطيران من 1 د الذباب الكبار ، ونحن أظهرت سابقا ان الحمض الريبي النيبالي الملزم البروتين و CELF1-متجانسات Bruno1 (Bru1 ، الاعتقال ، أريت) يتحكم في مسار الربط IFM محدده المصب من عامل النسخ Spalt الرئيسية (سالم)16. Ifms من المسوخ خاليه وكذلك الذباب مع العضلات الخاصة bruno1 rnai (bru1) عرض العيوب النمو ساركوميري, سوء تنظيم النشاط ميوسين وفي نهاية المطاف فرط الانكماش وفقدان ألياف العضلات16,17 . أدناه ونحن نظهر فائده من تشريح IFMs لقياس الطيف الكتلة بروتينيه كامله وتظهر ان العديد من التغييرات التعبير لاحظنا علي مستوي الحمض الريبي النيبالي هي أيضا واضحة علي مستوي البروتين. ونحن نسلط الضوء كذلك علي حدث لصق التنموية محدده في Mhc التي وجدت ان تنظمها Bruno1 ، مما يدل علي ان Mrna-SEQ و RT-PCR من تشريح ifms يمكن استخدامها لإثبات تنظيم الاحداث لصق البديلة.
اعتمادا علي جوده المكتبة وعمقها ، يمكن تحليل بيانات mrna-Seq علي مستوي وحدات الجينات (المتوسط قراءه التهم علي جميع الاكسون من الجينات) ، الاكسون الفردية ، أو وصله التقاطعات. البيانات المتسلسلة من bru1 ifms بالمقارنة مع النوع البري يظهر التغيرات الضعيفة في التعبير علي مستوي وحده الجينات16 (الشكل 5ا). 300 72 bru1-IR العضلات (الشكل 5A والجدول التكميلي 1). ومع ذلك ، لقد أظهرنا سابقا ان علي مستوي exons الفردية ، وهناك المزيد من التنظيم السفلي اقوي من الجينات ساركوميري محدده16، مما يوحي بان الوظيفة الرئيسية لل Bruno1 هو التحكم البديل الربط (الجدول التكميلي 1 ).
باستخدام قياس الطيف الكتلي بالبالكامل علي الifms التشريح ، يمكننا ان نظهر تنظيما مماثلا علي مستوي البروتين (الشكل 5ب والجدول التكميلي 2). من مجموعات الببتيد 1,895 الكشف عنها ، 524 (28 ٪) منهم وسوء التنظيم في Bru1M2 متحولة ifm في 1 د البالغين (الجدول التكميلي 2). كما لوحظ انخفاض التنظيم من كل من Strn-Mlck و Mlp60A البروتين, مطابقه الملاحظات علي مستوي النص في البيانات الخاصة بنا mRNA-Seq. علي الرغم من العدد المحدود من الببتيدات قاعده البيانات التي خريطة لبروتينات معينه اشكال الاسويه (انظر الأساليب التكميلية لتحليل التفاصيل) ، للبروتينات ساركوميري تروميوبين 1 (Tm1) ، أيدت (up/TnT) ، mhc ، عازمه (bt/مشروع) و paramyosin (اليه) نحن مراقبه الحامل من الببتيدات من ايزوفورم واحد وخفض التنظيم من آخر (الشكل 5ب) ، مؤكدا ملاحظاتنا السابقة للوائح مماثله علي مستوي الحمض الريبي النيبالي16. وهذا يوضح ان IFMs التشريح مفيده لكل من mRNA-Seq والتطبيقات البروتينية.
وكمثال آخر للكيفية التي يمكن ان تكمل بها البيانات المتعلقة بالمعلومات النهج التقليدية لتعزيز وتوسيع البصيرة البيولوجية ، اخترنا التركيز علي الربط في المحطة المتوسطة لل Mhc. وقد تم ادراج خط فخ البروتين الذي كان يتميز سابقا باسم weeP26 في المرحلة النهائية من mhc43،44 (انظر الأساليب التكميلية للموقع الدقيق). weeP26 يحتوي علي متقبل لصق قويه وأدرجت في يفترض جميع النصوص Mhc (الشكل 5ج). ومع ذلك ، يتم تضمين البروتين المسمي GFP في IFM في اثنين من "النقاط" علي جانبي الخط M ، بينما في عضله الساق ، فانه يدمج بشكل موحد عبر خط M وضعيف عبر خيوط سميكه (الرقم 5ه). وأظهرت اورفانوس وسبارو هذه "النقاط" في شكل ifm بسبب التحول mhc ايزوفورم التنموية: وأعرب ايزوفورم mhc قبل 48 h منتدى التنمية الاتحادية هو gfp-وصفت بأنها ادراج weeP26 اكسون في اطار قراءه مفتوحة ، في حين ان mhc وأعرب عن ايزوفورم بعد 48 h منتدى الدعوة الاتحادية غير مسمي ، كما يتم تضمين weeP26 اكسون المصب من كودون وقف في 3 '-utr44.
وقد سمحت لنا بيانات mRNA-Seq الخاصة بنا بتوصيف تعبير ايزوفورم Mhc من المحطة C بمزيد من التفصيل. في حين تم الإبلاغ عن الإنهاءات mhc مختلفه43,44, لدينا mrna-Seq البيانات والتعليق التوضيحي الحالي flybase (FB2019_02) تشير إلى ان هناك في الواقع ثلاثه الاحداث البديلة المحتملة لصق في mhc C-تيرمينوس (exon 34-35 ، 34-36 ، أو 34-37) (الشكل 5ج) ، الذي يؤكده RT-PCR (الشكل 5د). weeP26 يتم إدخال GFP في الداخل بين Exon 36 و 37; التالي ، كما علي حد سواء exon 34-35 و exon 34-36 ايزوفورم تحتوي علي وقف المخطوطات ، يمكن ان تترجم gfp فقط في exon 34-37 اشكال الاسويه (مما ادي إلى exon 34-gfp-37). ويمكننا أيضا ان نري كلا من التنظيم الزماني والمكاني لجميع ايزواشكال Mhc . في IFM ، نلاحظ التبديل ايزوفورم Mhc من exon 34-37 إلى exon 34-35 بين 30 ح المنتدى المذكور و 48 ح منتدى الدفاع العام (الشكل 5ج ، د ، و) في 27 درجه مئوية ، علي الرغم من ان هذا ليست مرئية حتى الآن عن طريق المناعة في 48 ح منتدى اسيا والشرق (الشكل 5ه). الساقين بالفعل التعبير عن خليط من exon 34-37 و exon 34-35 في المنتدى 30 ح ، وبواسطة 72 ح منتديات الدعوة السريعة التعبير عن جميع الثلاثة mhc اشكال الاسويه (الشكل 5د ، و). الكبار العضلات القفز (TDT) كما يعبر عن جميع الثلاثة Mhc ايزواشكال (الشكل 5و) ، مما يوحي بان هذا صحيح عموما للعضلات الجسدية أنبوبي. التالي, لدينا mRNA-Seq البيانات تسمح تمديد النتائج السابقة عن طريق تضييق الإطار الزمني للتبديل ايزوفورم mhc في ifm وتميز mhc ايزوفورم استخدام في عضلات أنبوبي.
تم فحصها بعد ذلك Mhc التنظيم ايزوفورم في سالم و bru1 متحولة ifm. في كلتا الحالتين ، راينا سوء تنظيم weeP26. سالم متحولة ifms تفشل لإكمال التحول التنموي في التعبير ايزوفورم mhc وأنماط الربط الساق فينوكوبي في مراحل لاحقه ، بما في ذلك كسب الحدث Exon 34-36 (الشكل 5و). وهذا يتفق مع النتائج السابقة ان فقدان سالم النتائج في تحويل مصير شبه كامل من IFM إلى العضلات أنبوبي16. Bru1 وBru1 متحولة ifm ، علي غرار سالم-/- ifm ، يحتفظ exon 34-37 لصق الحدث من خلال مراحل الكبار (الشكل 5ه ، و) ، مما ادي إلى نمط الوسم weeP26 gfp تشبه عضله الساق ، ولكنها لا تكسب الحدث Exon 34-36. وهذا يوحي بان Bruno1 ضروري في ifm للسيطرة جزئيا علي الأقل علي مفتاح النمو في الربط البديل mhc ، ولكنه يشير إلى ان عوامل الربط اضافيه هي أيضا سوء تنظيمها في سالم-/-السياق. وعلاوة علي ذلك ، يوضح هذا المثال كيف يمكن ان تكون البيانات RT-PCR و mRNA-Seq من تشريح IFM قيمه في اكتساب فهم أعمق لليات الربط التنموية والعيوب المورفولوجية الملحوظة.

الشكل 1: ifm التنمية والتجهيز لل pupae. 3272 48 32 الأوتار في الرمادي الداكن. (ب) الصور المنسقة لifms الثابتة من الأجزاء المفتوحة من الكتاب (24 ساعة ، 32 ساعة ، 48 ساعة)19 أو هيميسيكتيونس الصدر (72 ساعة ، 1 يوم) ملطخه بماده الاكتين (روامين phخلائط ، أرجواني) و gfp (الأخضر). (جيم ، دال) صور من الفلوري GFP في الدمى الحية التي توضح الشكل IFM سليمه من خط التشريح ذبابه في الظهرية (C) أو الجانبي (D) الطائرة. علامة النجميه موقع IFM. (ه) للتحضير لفصل المواد ، يجب ان تكون الأسهم المتطايرة معكوسه أو ان تكون متقاطعة قبل 3 – 4 أيام مقدما. (و) يتم اختيار prepupae باللون الأبيض (نصال الأصفر) ومعزولة باستخدام فرشاه الطلاء المبللة (f ', f ''''). (ز) ينبغي ان تلاعب prepupae لفصل الإناث من الذكور علي أساس وجود الخصيتين التي تظهر علي النحو التالي تقع كرات شفافة (النجميه الصفراء). (H) وتتراوح اعمار العرائس علي ورق الترشيح المبلل في اطباق 60 مم. قضبان المقياس = 100 μm (B) ، 1 سم (C ، D ، E ، H) ، 1 ملم (F ، F '''' ، G). يرجى النقر هنا لعرض نسخه أكبر من هذا الرقم.
{kind=link}

الشكل 2: تشريح ifms قبل 48 h منتديات الدعوة. (ا) أضافه 1x الحاجز العازل لصحن التشريح الأسود مع ماصه النقل. (ب) نقل العرائس المقسمة باستخدام فرشاه طلاء. (ج) تحت مجهر تشريح الفلورسنت لتصور gfp ، ودفع لطيف من العانة إلى الجزء السفلي من طبق تشريح باستخدام ملقط #5 (المبينة في الرمادي). "X" في دائره يدل علي الحركة في الصورة. (دال ، هاء) استيعاب الدمى الخلفية (D) ، ثم بدس من العرائس فقط وراء الصدر (E). اندفاعه في دائره لا يدل علي اي حركه. (واو ، زاي) سحب مع الملقط الامامي (السهم) لأزاله الحالة الخوادر (F) ، ثم أزاله البطن (G). (ح) تكرار الجيم-زاي لعده بوبا. الخطوط المنقطة الصفراء مرقمه بالمساهمة بالدمى. (أولا ، ياء) استخدام ملقط (I) لعزل IFMs من الانسجه المحيطة بها (J). نقطه في دائره يدل علي الحركة من الصفحة. (ك ، ل) أزاله الملوثات بما في ذلك الدهون والقفز (TDT) العضلات (K) لتوليد عينه IFM نظيفه (L). لدي TDT تعبير GFP اقل وشكل مختلف عن ألياف IFM (K '). (M, N, O) استخدام طرف ماصه مقصوصه (M) لجمع IFMs التشريح (N) ونقله إلى أنبوب الطرد المركزي الصغير (O). قضبان المقياس = 1 سم (A ، B ، M ، O) ، 1 مم (C-G) ، 500 μm (H-L ، N). يرجى النقر هنا لعرض نسخه أكبر من هذا الرقم.
{kind=link}

الشكل 3: تشريح ifms بعد 48 h منتديات الدعوة. (ا) محاذاة العرائس علي شريط مزدوج العصا. (ب) أزاله العرائس من الحالة بالبول عن طريق فتح الجانب الامامي (باء) ، وقطع القضية (ب) ، ورفع العانة (b ''''). تمثل رموز الدائرة نفس الشكل 2. (ج) نقل العرائس إلى المخزن المؤقت. (د) أزاله البطن عن طريق قطع مع مقص (السهام الصفراء المزدوجة) والانفصال عن الصدر (D '). (هاء ، واو) أضافه العازلة النظيفة (E) ، ثم قطع من الصدر في نصف طوليا (f ، f '). (زاي ، حاء) يمكن اجراء التفكيك تحت الضوء الأبيض (G) أو فلوري لتصور GFP (H) ؛ قطع من IFMs علي جانب واحد (ز ') ، ثم الجانب الآخر (ز '') ؛ رفع من الصدر مع ملقط (المبينة في الرمادي) (ز ''). (أولا ، ياء ، كاف) جمع IFMs في المخزن المؤقت (I) وأزاله تلوث العصب البطني الملوث (VNC) ، الأمعاء ، والقفز العضلات (TDT) (J) لتوليد عينه IFMS نظيفه (K). TDT لديها اقل GFP التعبير وشكل مختلف عن ألياف IFM (J '''' ، K '). (L, M) استخدام ملقط لنقل IFMs (L) إلى أنبوب الطرد المركزي الصغير (M). قضبان المقياس = 1 سم (A, E, M), 1 مم (B-D ', F-L). يرجى النقر هنا لعرض نسخه أكبر من هذا الرقم.
{kind=link}

الشكل 4: ifm الحفظ وتفاصيل العزلة RNA. (ا) يتم وضع الطرد باليد لمده 5 دقائق في 2000 x g. (ب) ifm بيليه (السهم) وبيليه تحت فلوري (B '). (ج) أزاله جميع العازلة بطرف ماصه. (د) لاستخراج RNA ، أعاده تعليق بيليه في العازلة العزلة. يمكن تخطي هذه الخطوة إلى تجميد الجافة التي تم تشريحها IFMs. (ه) تجميد العينة في النيتروجين السائل أو علي الجليد الجاف والتخزين عند-80 درجه مئوية. قضبان المقياس = 10 سم (A) ، 1 مم (B ، B ') ، 1 سم (C ، D ، E). (و) الصور المجسمة (ng) من إجمالي الحمض الريبي النيبالي من تشريح ifm الحصول علي كل ذبابه في 16 ح منتديات الدفاع الجوي ، 24 ح المنتدى المذكور ، 30 ح المنتدى ، 48 ح المنتدى الاتحادي لأسيا ، 72 ح منتديات الدعوة ، 90 h منتدى الدفاع الاتحادي ، و 1 د الكبار. أشرطه الخطا = SD. (ز) مجموع الحمض الريبي النيبالي معزولة من ifm تشريح من 50 1 d الذباب الكبار باستخدام أساليب استخراج مختلفه. أشرطه الخطا = SD. (ح) اثار الممثل لفحص سلامه الجيش النيبالي الريبي بعد أساليب استخراج مختلفه. العصابات ريبوسومال تشغيل اقل من 2000 النيوبوتيدس (nt) والفرقة علامة في 25 nt. اثار اضافيه متوفرة في الشكل التكميلي 1. (ط) اثار تمثيليه لعينه الحمض الريبي النيبالي المعزولة حديثا (اعلي) ، عينه تجميد-أذابه 25x علي الجليد الجاف (المؤامرة الثانية) ، عينه اليسار ل 4 ح علي مقاعد البدلاء (المؤامرة الثالثة) ، وعينه تعامل مع RNase a (المؤامرة السفلي). ملاحظه التحلل الكامل لل RNA بناء علي أضافه RNase ا. (ي) هلام RT-PCR من مجموعات كما وصفت ل bru1 و rp49. يتم رسم الكثافة النسبية للنطاق bru1 الذي تمت تسويته ضد rp49 أدناه. أشرطه الخطا = SEM (unpaired t-اختبار ، p = 0.0119). يرجى النقر هنا لعرض نسخه أكبر من هذا الرقم.
{kind=link}

الشكل 5: تطبيق الأقسام ifm للتحقيق في وظيفة Bruno1 في الربط البديل. (ا) قطعه بركان من mrna-البيانات المتسلسلة (وحده الجينات) من ifms تشريح في 72 ح المنتدى الاتحادي المذكور. يتم عرض الجينات التي يتم تنظيمها بشكل مختلف إلى حد كبير بين bru1 والنوع البري ifm (padj < 0.05 ، abs (log2FC) > 1.5) باللون الأزرق ، والجينات غير الهامه في الرمادي. يتم تمييز البروتينات ساركوميري باللون الأحمر ، ويتم تسميه الجينات المختارة. (ب) الأرض بركان الكتلة البروتينية كامله نتائج الطيف من 1 د البالغين ifms. البروتينات تختلف اختلافا كبيرا بين المسوخ BruM2و wildtype (روزفلت < 0.05) وتظهر في البروتينات الزرقاء ، غير الهامه في الرمادي. ويسلط الضوء علي البروتينات sarcomeric باللون الأحمر. يتم تسميه الببتيدات المقابلة للجينات في (A) باللون الأحمر. يتم تسميه مجموعات من الببتيدات تعيين إلى ايزواشكال مختلفه من نفس البروتين في نفس اللون. (ج) مخطط خط النهاية C لل mhc الذي يوضح النماذج المتشابهة للنص وموقع الادراج في مصيدة جين weeP26 (انظر الأساليب التكميلية لنقطه الادراج). ويشار إلى الإشعال RT-PCR كخطوط سوداء فوق النصوص. قراءه التهم لكل kilobase لكل مليون قاعده (rpkm) من mrna-Seq تظهر ل ifms تشريح من البرية في 30 ح منتدى البيئة الاتحادية (البرتقالي) و 72 h منتدى ال72 بيئة الاتحادية (الأحمر) ، من bru1 (الأزرق) و سالم-/- (سماوي) في 72 h المنتدى . (د) RT-PCR مع الإشعال ضد mhc تبين التبديل ايزوفورم في ifm بين 30 ح المنتدى والنقاط الزمنيه في وقت لاحق. الحدث Exon 34-35 لصق فقط لوحظ ضعيف في BruM3متحولة ifm أو في الساق الكبار. (ه) الصور المنسقة لتوطين weeP26 gfp في النوع البري ifm ساركوميريس في 48 h منتديات الاتصالات العامة و 90 h الاتحاد الاتحادي للاتصالات بالمقارنة مع 90 h العضلات الساق. قضبان المقياس = 1 μm. (و) لصق التحديد الكمي للموصل من بيانات Mrna-Seq الخاصة بالأنماط الجينية والنقاط الزمنيه كما هو مسمي. وتقدم القراءات تقاطع كنسبه لحدث لصق محدده (اكسون 34 إلى 35 في الرمادي ، 34 إلى 36 في الأرجواني ، و 34 إلى 37 في الأخضر) إلى مجموع عدد الاحداث تقاسم اكسون 34 لصق المانحة. يرجى النقر هنا لعرض نسخه أكبر من هذا الرقم.
{kind=link}

الشكل التكميلي 1: (ا ، ب ، ج) غله الحمض الريبي النيبالي من عينات من نفس النمط الجيني الذي قام الباحث نفسه بتشريحه في نفس الأسبوع. وبعد تشريح جميع العينات ، كانت الRNA معزولة وتقاس في نفس اليوم. (ا) الصور المجسمة (ng) من الحمض الريبي الإجمالي الذي تم الحصول عليه من الأجزاء IFM لكل ذبابه بالغه 1 د. أشرطه الخطا = SEM. (ب) إجمالي الحمض الريبي الذي تم الحصول عليه من IFM التشريح لكل ذبابه في 30 ح منتديات الاعلام ، 48 ح المنتدى المذكور ، 72 ح منتديات الدعوة والبالغين 1 د. (C) مجموع الحمض الريبي النيبالي معزولة من IFM تشريح من 50 1 d الكبار الذباب باستخدام أساليب استخراج مختلفه. (د) مجموع تركيزات الحمض الريبي النيبالي لكل ذبابه من الساقين تشريح ، والقفز العضلات (tdt) و ifm. يتم الحصول علي المزيد من RNA من IFMs أكبر. أشرطه الخطا = SD. (ه) مجموع تركيزات الحمض الريبي النيبالي لكل ذبابه من ifm تشريحها من الضوابط بالمقارنة مع rnai أو عينات متحولة في 30 ح منتديات التنمية المستدامة ، 72 ح المنتديات الاتحادية والبالغين 1 د. بالنسبة للمتحولين ، تم استخدام w1118 كعنصر تحكم النوع الوحشي. يتم تجميع البيانات متحولة من bru1، سالم-/- واخر الحمض الريبي النيبالي--ربط البروتين متحولة. لاحظ ان لهذه التلاعبات ، وانخفضت غله RNA في 1 د الكبار بسبب ضمور العضلات والخسارة ، لذلك المزيد من الذباب تحتاج إلى تشريح للحصول علي كميات كافيه للنهج اوميكس. أشرطه الأخطاء = SD. (و) الآثار الاضافيه التي تبين سلامه الحمض الريبي النيبالي لأساليب عزل الحمض الريبي النيبالي المبينة في الشكل 4زاي وفي الشكل التكميلي 1 جيم. يرجى النقر هنا لعرض نسخه أكبر من هذا الرقم.
{kind=link}

الأساليب التكميلية: (ب) وصف مفصل للأساليب والكاشفات المستخدمة في جميع أجزاء النص ، وبخاصه لتوليد البيانات المبينة في الشكل 1الف-دال، والشكل 4واو-كاف ، والشكل 5، والجدول التكميلي 1، و الجدول التكميلي 2. هذه البيانات تحفز البروتوكول تشريح وإثبات فائدته للعزل RNA, mRNA-Seq, RT-PCR, وبروموميكس. الرجاء الضغط هنا لتحميل هذا الملف.
| فيما يتعلق بالشكل 5 والفقرات المرتبطة به في النص الرئيسي | |
| اسم علامة التبويب | ملخص البيانات |
| البروتينات ساركوميري | قائمه الجينات ساركوميري من الشريحة وآخرون Elife 2018 ؛ هنا نحن قائمه FBgn الحالية واسم الجينات. |
| SP الجينات units_DESeq2_72h | باستخدام البيانات من الشريحة وآخرون تيمبو Rep 2015 ، نظرنا علي وجه التحديد في الجينات ساركوميري في بيانات mRNA-Seq في 72 ح منتدى الدعوة. هذا هو من تحليل DESeq2 الكشف عن التعبير التفاضلي علي مستوي وحده الجينات بين السيطرة (Mef2-Gal4, UAS-GFM-Gma عبرت إلى w1118) وMef2-Gal4, UAS-GFM-Gma x Bruno1-IR. الصفوف المبرزة باللون الأصفر هي الجينات المنظمة بشكل لافت للأعلى أو لأسفل (فوق/تحت عتبه log2FC = abs (1.5)). هذه البيانات هي تراكب النقطة الحمراء في الشكل 5A. لكل جين ساركوميري ، ونحن نقدم معلومات المعرف ، و log2FC من DESeq2 ، p القيمة والقيمة p المعدلة ، فضلا عن DESeq2 التعبير تطبيع التهم. |
| SP exon_DEXSeq_72h | باستخدام البيانات من الشريحة وآخرون تيمبو Rep 2015 ، ونحن ننظر علي وجه التحديد في ساركوميري الجينات اكسون استخدامها في بيانات mrna-Seq في 72 ح منتدى الدعوة. هذا هو من التحليل dexseq الكشف عن التفاضلية اكسون الاستخدام بين السيطرة (Mef2-Gal4, uas-gfm-gma عبرت إلى w1118) وMef2 Gal4, uas-gfm-gma x Bruno1-IR. الصفوف التي تم تمييزها باللون الأصفر هي بشكل لافت اعلي أو أسفل الاكسون المنظمة (فوق/تحت عتبه log2FC = abs (1.5)). نحن نقدم اكسون ومعلومات معرف الجينات ، و log2FC من dexseq ، p القيمة والقيمة p المعدلة ، فضلا عن قائمه من النصوص المرتبطة بها. |
| يرجى ملاحظه ان العديد من الجينات تظهر تنظيم واحد أو أكثر من الاكسون في تحليل dexseq ، وغالبا مع قيم log2FC عاليه وقيمه p منخفضه/ضبط قيم p ، في حين ان قائمه محدوده من الجينات يظهر التغييرات في 72 h. وهذا يدعم تاثير قوي لفقدان برونو علي تنظيم الربط البديل. | |
الجدول التكميلي 1: جدول 72 h منتديات البيانات المتسلسلة لبروتينات ساركوميري التي تحدد الجينات المعرب عنها بشكل مختلف (عبر DESeq2) و الاكسون (عبر dexseq) في bru1 vs. النوع الوحشي ifms.
| الشكل 5 باء والفقرات المرتبطة به في النص الرئيسي | |
| اسم علامة التبويب | ملخص البيانات |
| الإخراج ساوس | وهذا جدول بيانات مجهز يقدم بيانات قياس الطيف الكتلي المستخدمة لتوليد الشكل 5 باء. عينات IFM هي من 1 د الكبار السيطرة (w1118) ومتحولة (bruno1) الذباب. الاعمده الهامه هي قيم الكثافة المحولة لكل من 4 نسخ متماثلة لكل عينه ، وإحصاء t-اختبار واهميه ، معرفات الببتيد وأسماء الجينات المقابلة ومعرفات Flybase. تم حساب signifance باستخدام الإعدادات القياسية في ساوس (روزفلت <. 05). هناك 1859 البروتينات/الببتيدات المكتشفة ، منها 524 (28 ٪) تختلف اختلافا كبيرا بين العينات. |
| دوونريجولاتيد | هذه هي جميع البروتينات 252/الببتيدات من الناتج ساوس التي يتم دوونريجولاتيد في bruno1 متحولة ifm. كما معرفات Flybase وأسماء الجينات عفا عليها الزمن ، ونحن نقدم بالاضافه إلى ذلك الحالية الجينات Flybase معرف واسم الجينات. |
| اوبريجولاتيد | هذه هي جميع البروتينات 272/الببتيدات من الناتج ساوس التي يتم اوبريجولاتيد في bruno1 متحولة ifm. كما معرفات Flybase وأسماء الجينات عفا عليها الزمن ، ونحن نقدم بالاضافه إلى ذلك الحالية الجينات Flybase معرف واسم الجينات. |
| يرجى ملاحظه ان البروتينات ساركوميري المبينة باللون الأحمر في الشكل 5B موجودة في القوائم المذكورة أعلاه. قائمه الجينات التي تعتبر جزءا من ساركوميري متاحه في واحده من علامات التبويب في الجدول التكميلي 1. | |
الجدول التكميلي 2: جدول البيانات الشاملة لقياس الطيف الكتلي من 1 د الكبار تحديد بشكل مختلف البروتينات والبروتين ايزواشكال في بروM2متحولة مقابل. النوع الوحشي IFMs.
Discussion
في هذا البروتوكول ، نقدم التقنية الاساسيه لتشريح دروفيا ifms من الدمى المبكرة والمتاخره المرحلة لعزل البروتين ، الحمض النووي ، RNA أو الجزيئات الأخرى. ويمكن تكييف البروتوكول بسهوله لتشريح IFM من الذباب الكبار. ونحن نظهر فائده من بروتوكول لدينا تشريح لل mRNA-Seq ، البروتينية والتطبيقات RT-PCR. مع التحسين المستمر للتكنولوجيات اوميكس للسماح بتحليل العينات مع مواد البداية اقل وتركيزات المدخلات اقل ، وهذه الأجزاء من المرجح ان تصبح قيمه للعديد من التطبيقات الاضافيه. كما ifms هي نموذج الثابتة لاعتلال العضلات البشرية4,24 والعضلية من نوع التنمية المحددة9,12, ونحن نتصور, علي سبيل المثال, ifms التخصيب الأيض, التحقيقات من التشكيل الكروماتين عبر 3C أو 4C ، والربط تقييم الشبكة عن طريق التفاعلات كليب أو فوسفوس-بروتينيه من ميوفيبريلوجينيسيس.
من المهم ان نعتبر ان هذه الأجزاء تنتج عينه المخصب ل IFM بدلا من عينه IFM نقيه. وهذا أمر لا مفر منه بسبب الحركية العصبية ، المرفقات وتر والاجتياح الرغامي من ألياف العضلات. ويمكن استخدام تحليل المعلوماتية الحيوية لتحديد الجينات المخصبة IFM أو البروتينات ، ولكن هناك حاجه إلى مزيد من التجارب لإثبات انها في الواقع IFM الخاصة. ويمكن النظر إلى نقاء العينة باستخدام علامات الانسجه المحددة المنشورة مثل45 شريطيه (وتر) ، Act79B4،44 (العضلات الانبوبيه) ، Act88F15 (ifm) ، أو syb46 (الخلايا العصبية المحددة). وقد يكون من الممكن استخدام هذه العلامات لتطبيع مجموعات البيانات مع المحتوي الخاص بالاتحاد ، ولكن المستخدمين يحذرون من ان التغيرات الزمنيه في التعبير عن الجينات المستخدمة في التطبيع ، مثل الجينات أو الأنابيب الخاصة بالاتحاد ، قد تنحاز إلى مثل هذا النهج.
وراثيا المشفرة الانسجه المحددة طرق الوسم ، علي سبيل المثال EC-وضع العلامات47،48 أو باب وسم49،50 لعزل الجيش النيبالي الريبي وقد تم تطويرها في السنوات الاخيره ، والتي قد تساعد في الحصول علي حقا عينه [رنا] خاصه بالانسجه. ومع ذلك ، الجماعة الاوروبيه-وضع العلامات يتطلب تغذيه مستمرة من الذباب47 التالي لا ينطبق خلال مراحل الخوادر. قد يكون حساسية واكتمال المدونات المسمية بابف القيود51. وتعقد نهج FACS لعزل ألياف العضلات الفردية بالحجم الكبير والطبيعة المتزامنة IFMs. 52سليمه ،يمكن تطبيق نهج نمط53 لعزل المقصورات فرعيه معينه من ifms ، والتي قد تكون مفيده لعزل السكان النقي من نوى ifms أو الميتوكوندريا. لا تزال المقاطع اليدوية هي المعيار الحالي للحصول علي انسجه IFM السليمة لمعظم التطبيقات المتلقية للمعلومات.
نوعيه العينة يعتمد علي عده خطوات حاسمه في عمليه التشريح. الأقسام هي المطالبة من الناحية الفنية ، مع سرعه التشريح ونقاء العينة زيادة مع الخبرة. تشريح لفترات قصيرة من الزمن (20-30 دقيقه) في العازلة المبردة دون المنظفات وتجميد فورا يساعد علي الحفاظ علي سلامه العينة ، كما لوحظ سابقا لعزل وتر الماوس54. IFMs يمكن ان تكون بنجاح الجافة المجمدة بعد أزاله جميع العازلة من بيليه ، ولكن علي وجه التحديد لعزل الجيش النيبالي الريبي ، وتجميد عينات في العازلة العزلة يميل إلى إنتاج نتائج أفضل. يتم الجمع بين ifms من ما يصل إلى 20 فصل منفصلة قبل الحمض الريبي النيبالي أو عزل البروتين ، مما يسمح التوسع وجمع ما يكفي من المواد ، حتى من الوقت المبكر أو المسوخ16،32، للتحليل المصب.
ل [رنا] تطبيقات, الخطوة حاسمه أكثر يمكن كنت العزلة من ال [رنا]. غوانيدينيوم ثيوسيانات-الفينول-كلوروفورم العزلة (الطريقة 1 أعلاه) يتفوق معظم مجموعات التجارية اختبارها ، وكما لوحظ سابقا ، هو اقل تكلفه بكثير55. التغير يلاحظ في [رنا] عزل غله مع مجموعه تجاريه اتفاق مع ملاحظات سابقه56,57. نضيف أيضا الجليكوجين خلال الايزوبروبانول هطول الامطار للمساعدة في استرداد جميع RNA. إلى ما بعد [رنا] غله, هو مهمة ان يتحقق [رنا] نزاهة ان يضمن ان العينة يتلقى لم يكن مجزاه أو تدهور اثناء التشريح وعزل عمليات. ومن الضروري أيضا العمل خاليه من RNase. وأخيرا ، فان اختيار RT-كيت يمكن ان تؤثر علي حساسية عمليه النسخ العكسي. وفي حين ان هذه النقاط لا تناقش بالتفصيل في كثير من الأحيان ، فانها تؤثر علي نوعيه عينه الاتحاد والبيانات التي يتم الحصول عليها من التطبيقات النهائية.
العديد من التعديلات الهامه تعيين البروتوكول باستثناء بروتوكولات تشريح IFM الموجودة. علي الرغم من وجود بروتوكول تشريح مفصل ل ifm المناعية موجود19، ويقدم هذا البروتوكول نهجا مختلفا لتفكيك الخوادر التي تسمح العزلة الأسرع من الانسجه ifm. وهذا يسمح بجمع كميات كبيره من انسجه IFM (التحدث نسبيا) مع فترات تشريح محدوده لمنع التغيرات البروتينية أو الناسخة. البروتوكولات الأخرى تصف تشريح ifm الكبار لتصور تلطيخ gfp في الفردية myoالفايليلز39 أو لتلطيخ العضلات الجسم الجدار58، ولكنها لا تعالج تشريح في مراحل الخوادر أو لعزل الحمض الريبي النيبالي أو البروتين. هذا النهج هو أيضا متميزة عن البروتوكول الحالي لتشريح المجهرية من الخوادر ifms من الحلوى38، والتي قد تولد عينه الأنقى ifms ولكن هو أكثر كثافة العمالة وتنتج مواد اقل. بالمقارنة مع غيرها من السريع الكبار ifm البروتوكولات تشريح38،39، يتم عزل ifm في المخزن المؤقت بدون المنظفات للحد من الإجهاد التعريفي وغيرها من التغييرات الرئيسية التعبير.
التقدم الرئيسي في هذا البروتوكول هو ادراج مراسل الفلورية الحية ، مما يسمح بعزل ifms في مراحل الخوادر في وقت مبكر. نحن الخسيس استخدام Mef2-GAL459 القيادة اما UAS-CD8:: gfp أو Uas-gfp:: Gma60. وهذا يسمح الوسم التفاضلية ifm (عضلات الطيران هي أكثر بقوة المسمي وشكل مختلف من العضلات الخوادر الأخرى) ، فضلا عن أداء GAL4-uas-التلاعب القائم ، علي سبيل المثال الإنقاذ أو التجارب rnai. من الممكن أيضا الجمع بين Mef2-GAL4 مع الحوض-GAL80ts لتجنب rnai المرتبطة الفتك المبكر أو مع uas-Dcr2 لزيادة الكفاءة rnai40.
هناك اضافيه GAL4 السائقين أو gfp-خطوط المتاحة التي تختلف في خصوصية نوع العضلات, نمط التعبير الزمني, وقوه السائق19,61 التي يمكن استخدامها بدلا من Mef2-GAL4. علي سبيل المثال ، يتم التعبير عن Act88F-GAL4 لأول مره حول 24 ح منتدى المحيط الهادي ، لذلك لا يمكن استخدامها للنقاط الزمنيه السابقة. ومع ذلك ، فانه بشده تسمي IFM وقد تكون مفيده لتجنب الفتك المبكر المرتبطة RNAi. له-gfp أو Act88F-GFP التسمية ifm ، مره أخرى مع القيود الزمنيه ، لكنها تجنب الاعتماد GAL4 من علامة التعبير وقد تكون مفيده في تركيبه مع خلفيه متحولة من الاهتمام. تتوفر قوائم بخطوط العلامات المحتملة الأخرى19. وتجدر الاشاره أيضا إلى ان استخدام الجينات المتحولة ونظام GAL4/UAS قد يسبب الآثار الجينية التعبير ، لذلك فمن المهم استخدام الضوابط المناسبة ، علي سبيل المثال خط السائق عبرت إلى سلاله الخلفية من نوع البرية ، بحيث يفترض ان هذه التحف نفسه في جميع العينات.
مع الفيديو المصاحبة, هذا البروتوكول المفصل يهدف إلى جعل الخوادر ifm تشريح أكثر سهوله وتعزيز استخدام النهج اوميكس لدراسة تنميه العضلات. اقتران قوه الوراثة دروفيا والبيولوجيا الخلية مع الكيمياء الحيوية واختبارات اوميكس يمكن الوصول اليها من خلال تشريح ifm لديه القدرة علي تعزيز الفهم الألى لتولد ووظيفة العضلات. ستوفر الدراسات المستقبلية التي تربط بين الملاحظات علي مستوي الانظمه الخاصة بالتدوين والتنظيم البروتيني للنواتج الايضيه والوظيفية فهم أعمق للتطور النوعي للعضلات والتسبب في اضطرابات العضلات.
Disclosures
وليس لدي المؤلفين ما يفصحون عنه.
Acknowledgements
ونحن ممتنون لاندرياس لادورنر وفرانك شونورير للدعم السخي. نشكر ساندرا يسر علي المساعدة التقنية الممتازة و Akanksha Roy لتوليد بيانات الطيف الكتلي. نحن نعترف بمراكز الأسهم في بلومنقتون وفيينا لتوفير الذباب. ونشكر المركز الأساسي للتصوير الإحيائي للمساعدة في التصوير البؤري المركزي والتحليل التحليلي لعينات قياس الطيف الكتلي ، سواء في المجمع الطبي الحيوي LMU (Martinsried, DE). وقد دعمت عملنا من قبل المانيه Forschungs Gemeinschaft (MLS ، SP 1662/3-1) ، ومركز للعلوم البروتين المتكاملة ميونيخ (سيبسم) في جامعه لودفيغ-ماكسيميليان ميونيخ (MLS) ، وفريدريتش باور ستيفتونغ (MLS) ، والدولية ماكس مدرسه بلانك للبحوث (EN).
Materials
| Name | Company | Catalog Number | Comments |
| 5x High Fidelity (HF) buffer | Thermo Fisher | F518L | |
| 60 mm culture dishes | Sigma-Aldrich | Z643084-600EA | Greiner dishes, 60 mm x 15 mM, vented |
| Black dissecting dish (glass) | Augusta Laborbedarf | 42021010 | Lymphbecken, black glass, 4 cm x 4 cm |
| Black silicon dissecting dishes: activated charcoal powder | Sigma-Aldrich | C9157 | Also available from most pharmacies |
| Black silicon dissecting dishes: Sylgard 184 | Sigma-Aldrich | 761036 | Dishes are made by mixing the Sylgard (~50g) with activated charcoal powder (200 mg) and curing it in Petri dishes (~4 60 mm dishes). |
| Blue pestle | Sigma-Aldrich | Z359947-100EA | Any pellet pestle that can sterilized, also can be used with a motor-driven grinder |
| Cell phone camera, Samsung Galaxy S9 | Samsung | SM-G960F/DS | used for photos not taken under a microscope |
| Chloroform | PanReac AppliChem | A3691,0500 | |
| Confocal microscope, Leica SP8X upright confocal | Leica | www.leica-microsystems.com | |
| Confocal microscope, Zeiss LSM 780 inverted confocal | Zeiss | www.zeiss.com | |
| Double stick tape | Scotch/3M | 3M ID 70005108587 | Double-sided tape, available at most office supply handlers |
| Dumont #5 Forceps | Fine Science Tools | 11252-20 | Inox straight tip 11 cm forceps, Biology grade with 0.05 mm x 0.02 mm tip |
| EtOH (100%, RNase free) | Sigma-Aldrich | 32205-M | |
| fluorescent dissecting microscope camera, Leica DFC310 FX camera | Leica | www.leica-microsystems.com | |
| Fluorescent dissecting microscope software, Leica Application Suite (LAS) version 4.0.0 | Leica | www.leica-microsystems.com | |
| Fluorescent dissecting microscope, Leica M165 FC | Leica | www.leica-microsystems.com | |
| Fly: Bru1[M2] | Fly stock; This paper | ||
| Fly: Bru1[M3] | Fly stock; This paper | ||
| Fly: Mef2-GAL4 | Bloomington Stock Center | BDSC:27390 | Fly stock |
| Fly: salm[1] | Bloomington Stock Center | 3274 | Fly stock |
| Fly: salm[FRT] | Fly stock; see Spletter et al., Elife, 2018 | ||
| Fly: UAS-Bru1IR | Vienna Drosophila Research Center | GD41568 | Fly stock, RNAi hairpin |
| Fly: UAS-GFP::Gma | Bloomington Stock Center | BDSC:31776 | Fly stock |
| Fly: UAS-mCD8a::GFP | Bloomington Stock Center | BDSC:5130 | Fly stock |
| Fly: w[1118] | Bloomington Stock Center | 3605 | Fly stock |
| Fly: weeP26 | Fly stock; see Clyne et al., Genetics, 2003 | ||
| GFP detection reagent, GFP-Booster | ChromoTek | gba488-100 | |
| Glycogen | Invitrogen | 10814-010 | |
| Image processing software, Photoshop CS6 | Adobe | www.adobe.com | |
| Isopropanol | Sigma-Aldrich | I9516-25ML | 2-propanol |
| Method 1 (RNA isolation): TRIzol | Life Technologies | 15596018 | Guanidinium isothiocyanate and phenol monophasic solution |
| Method 2 (RNA isolation): Method 1 + TURBO DNA-free Kit | Invitrogen | AM1907 | TRIzol isolation followed by treatment with a kit to remove DNA |
| Method 3 (RNA isolation): Direct-zol RNA Miniprep Plus Kit | Zymo Research | R2070S | RNA isolation in TRIzol, but over commercial columns instead of using phase separation. Recommended DNase treatment performed with Monarch Dnase I in Monarch DNase I Reaction buffer. |
| Method 4 (RNA isolation): RNeasy Plus Mini Kit | Qiagen | 74134 | We used the provided DNase treatment. IFM pellets were homogenized in RTL buffer as suggested for animal tissues. |
| Method 5 (RNA isolation): ReliaPrep RNA Tissue Miniprep System | Promega | Z6110 | We applied the protocol for ‘Purification of RNA from Fibrous Tissues’. |
| Method 6 (RNA isolation): Monarch Total RNA Miniprep Kit | New England Biolabs | T2010G | We applied the protocol for tissues/leukocytes and lysed in 300 μL of RNA Protection Reagent. |
| Microcentrifuge tubes | Thermo Fisher | AM12400 | RNase-free Microfuge Tubes, 1.5 mL |
| Microscope slides | Thermo Fisher | 12342108 | Standard slides, uncharged, 1 mm |
| Microtome blades | PFM Medical | 207500003 | C35 feather 80 mm |
| Monarch DNase I | New England Biolabs | T2004-21 | |
| Monarch DNase I Reaction Buffer | New England Biolabs | T2005-21 | |
| Normal goat serum | Thermo Fisher | 16210072 | |
| OneTaq Polymerase | New England Biolabs | M0480X | |
| Paintbrush | Marabu | 1910000000 | Marabu Fino Round No. 0, or similar brush from any art supply |
| Paraformaldehyde | Sigma-Aldrich | 158127 | |
| PBS buffer (1x) | Sigma-Aldrich | P4417 | Phosphate buffered saline tablets for 1 L solutions, pH 7.4 |
| PFA PureTip Pipette Tips | Elemental Scientific | ES-7000-0101 | Optional substitute for standard pipette tips to reduce sample loss; 100 mL, 0.8 mm orifice |
| Phusion High Fidelity Polymerase | Thermo Fisher | F-530XL | |
| Pipette tips | Sigma-Aldrich | P5161 | Universal 200 mL pipette tips |
| Preomics iST 8x Kit | Preomics | P.O.00001 | peptide preparation kit for mass spectrometry |
| Q Exactive mass spectrometer | Thermo Fisher | 725500 | mass spectrometry was performed at the Protein Analysis Unit of the LMU Biomedical Center |
| Qubit RNA Assay Kit | Life Technologies | Q32855 | |
| Rhodamine-phalloidin | Invitrogen, Molecular Probes | 10063052 | |
| RNA concentration Approach 1 & RNA integrity traces, Bioanalyzer | Agilent Technologies | G2939BA | |
| RNA concentration Approach 2, Nanodrop | Thermo Fisher | ND-2000 | |
| RNA concentration Approach 3, Qubit 4 Fluorometer | Invitrogen | Q33238 | |
| RNA Pico Chips | Agilent Technologies | 5067-1513 | |
| RNase A | Promega | A7937 | |
| RNase-free water, Diethyl pyrocarbonate (DEPC) | Sigma-Aldrich | D5758 | DEPC treat water overnight and then autoclave, to remove all RNase. |
| RT Kit #1: Super Script III Reverse Transcriptase Kit | Invitrogen | 18080-044 | reverse transcription kit |
| RT Kit #2: LunaScript | New England Biolabs | E3010S | reverse transcription kit |
| RT Kit #3: QuantiNova Reverse Transcription Kit | Qiagen | 205410 | reverse transcription kit |
| Slide mounting buffer, Vectashield | Vector Laboratories | H-1200 | containing DAPI |
| Statistical software: GraphPad Prism | GraphPad Prism | www.graphpad.com | |
| Statistical software: Microscoft Excel | Microsoft | Purchased as part of the bundle: Office Home & Student 2019 | |
| Table-top centrifuge | Eppendorf | 5405000760 | Eppendorf Centrifuge 5425 or equivalent |
| Tissue/ Kimwipes | Sigma-Aldrich | Z188956 | Standard tissue wipes |
| Transfer pipette | Sigma-Aldrich | Z350796 | Plastic pipette |
| Triton-X100 | Sigma-Aldrich | T9284-500ML | |
| Vannas spring scissors | Fine Science Tools | 15000-00 | 3 mm cutting edge, tip diameter 0.05 mm, length 8 cm |
| Whatman paper | Sigma-Aldrich | 1004-070 | Filter paper circles, Grade 4, 70 mm |
References
- Rexiati, M., Sun, M., Guo, W. Muscle-Specific Mis-Splicing and Heart Disease Exemplified by RBM20. Genes. 9 (1), 18 (2018).
- Guo, W., et al. RBM20, a gene for hereditary cardiomyopathy, regulates titin splicing. Nature Medicine. 18 (5), 766-773 (2012).
- Guo, W., et al. Splicing Factor RBM20 Regulates Transcriptional Network of Titin Associated and Calcium Handling Genes in The Heart. International Journal of Biological Sciences. 14 (4), 369-380 (2018).
- Nikonova, E., Kao, S. -. Y., Ravichandran, K., Wittner, A., Spletter, M. L. Conserved functions of RNA-binding proteins in muscle. The International Journal of Biochemistry & Cell Biology. 110, 29-49 (2019).
- Wang, E. T., et al. Dysregulation of mRNA Localization and Translation in Genetic Disease. The Journal of Neuroscience. 36 (45), 11418-11426 (2016).
- Wang, E. T., et al. Antagonistic regulation of mRNA expression and splicing by CELF and MBNL proteins. Genome Research. 25 (6), 858-871 (2015).
- Kalsotra, A., et al. A postnatal switch of CELF and MBNL proteins reprograms alternative splicing in the developing heart. Proceedings of the National Academy of Sciences of the United States of America. 105 (51), 20333-20338 (2008).
- Ho, T. H., et al. Muscleblind proteins regulate alternative splicing. The EMBO Journal. 23 (15), 3103-3112 (2004).
- Lemke, S. B., Schnorrer, F. Mechanical forces during muscle development. Mechanisms of Development. 144, 92-101 (2017).
- Iwamoto, H. Structure, function and evolution of insect flight muscle. Biophysics. 7, 21-28 (2011).
- Schnorrer, F., Dickson, B. J. Muscle building; mechanisms of myotube guidance and attachment site selection. Developmental Cell. 7 (1), 9-20 (2004).
- Spletter, M. L., Schnorrer, F. Transcriptional regulation and alternative splicing cooperate in muscle fiber-type specification in flies and mammals. Experimental Cell Research. 321 (1), 90-98 (2014).
- Benoist, P., Mas, J. A., Marco, R., Cervera, M. Differential muscle-type expression of the Drosophila troponin T gene. A 3-base pair microexon is involved in visceral and adult hypodermic muscle specification. Journal of Biological Chemistry. 273 (13), 7538-7546 (1998).
- Schönbauer, C., et al. Spalt mediates an evolutionarily conserved switch to fibrillar muscle fate in insects. Nature. 479 (7373), 406-409 (2011).
- Bryantsev, A. L., et al. Extradenticle and Homothorax Control Adult Muscle Fiber Identity in Drosophila. Developmental Cell. 23 (3), 664-673 (2012).
- Spletter, M. L., et al. The RNA-binding protein Arrest (Bruno) regulates alternative splicing to enable myofibril maturation in Drosophila flight muscle. EMBO Reports. 16 (2), 178-191 (2015).
- Oas, S. T., Bryantsev, A. L., Cripps, R. M. Arrest is a regulator of fiber-specific alternative splicing in the indirect flight muscles of Drosophila. The Journal of Cell Biology. 206 (7), 895-908 (2014).
- Kim, J. H., Jin, P., Duan, R., Chen, E. H. ScienceDirect Mechanisms of myoblast fusion during muscle development. Current Opinion in Genetics & Development. 32, 162-170 (2015).
- Weitkunat, M., Schnorrer, F. A guide to study Drosophila muscle biology. Methods. 68 (1), 2-14 (2014).
- Rai, M., Nongthomba, U., Grounds, M. D. Skeletal Muscle Degeneration and Regeneration in Mice and Flies. Mechanisms of Regeneration. 108, 247-281 (2014).
- Swank, D. M., Wells, L., Kronert, W. A., Morrill, G. E., Bernstein, S. I. Determining structure/function relationships for sarcomeric myosin heavy chain by genetic and transgenic manipulation of Drosophila. Microscopy Research and Technique. 50 (6), 430-442 (2000).
- de Joussineau, C., Bataillé, L., Jagla, T., Jagla, K. Diversification of muscle types in Drosophila: upstream and downstream of identity genes. Current Topics in Developmental Biology. 98, 277-301 (2012).
- Maqbool, T., Jagla, K. Genetic control of muscle development: learning from Drosophila. Journal of Muscle Research and Cell Motility. 28 (7-8), 397-407 (2008).
- Jagla, K., Kalman, B., Boudou, T., Hénon, S., Batonnet-Pichon, S. Beyond mice: Emerging and transdisciplinary models for the study of early-onset myopathies. Seminars in Cell & Developmental Biology. 64, 171-180 (2017).
- Haigh, S. E., et al. Drosophila indirect flight muscle specific Act88F actin mutants as a model system for studying congenital myopathies of the human ACTA1 skeletal muscle actin gene. Neuromuscular Disorders. 20 (6), 363-374 (2010).
- Batonnet-Pichon, S., et al. Myofibrillar Myopathies: New Perspectives from Animal Models to Potential Therapeutic Approaches. Journal of Neuromuscular Diseases. 4 (1), 1-15 (2017).
- Kreipke, R. E., Kwon, Y. V., Shcherbata, H. R., Ruohola-Baker, H. Drosophila melanogaster as a Model of Muscle Degeneration Disorders. Current Topics in Developmental Biology. 121, 83-109 (2017).
- Souidi, A., Zmojdzian, M., Jagla, K. Dissecting Pathogenetic Mechanisms and Therapeutic Strategies in Drosophila Models of Myotonic Dystrophy Type 1. International Journal of Molecular Sciences. 19 (12), 4104 (2018).
- Sparrow, J., Hughes, S. M., Segalat, L. Other Model Organisms for Sarcomeric Muscle Diseases. Advances in Experimental Medicine and Biology. 642, 192-206 (2008).
- Lloyd, T. E., Taylor, J. P. Flightless flies: Drosophila models of neuromuscular disease. Annals of the New York Academy of Sciences. 1184, 1-20 (2010).
- Swank, D. M. Mechanical analysis of Drosophila indirect flight and jump muscles. Methods. 56 (1), 69-77 (2012).
- Spletter, M. L., et al. A transcriptomics resource reveals a transcriptional transition during ordered sarcomere morphogenesis in flight muscle. eLife. 7, 1361 (2018).
- Weitkunat, M., Kaya-Çopur, A., Grill, S. W., Schnorrer, F. Tension and force-resistant attachment are essential for myofibrillogenesis in Drosophila flight muscle. Current Biology. 24 (7), 705-716 (2014).
- Gunage, R. D., Dhanyasi, N., Reichert, H., VijayRaghavan, K. Drosophila adult muscle development and regeneration. Seminars in Cell & Developmental Biology. 72, 56-66 (2017).
- Weitkunat, M., Brasse, M., Bausch, A. R., Schnorrer, F. Mechanical tension and spontaneous muscle twitching precede the formation of cross-striated muscle in vivo. Development. 144 (7), 1261-1272 (2017).
- Zappia, M. P., Rogers, A., Islam, A. B. M. M. K., Frolov, M. V. Rbf Activates the Myogenic Transcriptional Program to Promote Skeletal Muscle Differentiation. Cell Reports. 26 (3), 702-719 (2019).
- Zappia, M. P., Frolov, M. V. E2F function in muscle growth is necessary and sufficient for viability in Drosophila. Nature Communications. 7 (1), 10509 (2016).
- Bryantsev, A. L., et al. Myogenesis in Drosophila melanogaster: Dissection of Distinct Muscle Types for Molecular Analysis. Methods in Molecular Biology. 1889 (5), 267-281 (2019).
- Xiao, Y. S., Schöck, F., González-Morales, N. Rapid IFM Dissection for Visualizing Fluorescently Tagged Sarcomeric Proteins. Bio-Protocol. 7 (22), (2017).
- Kaya-Çopur, A., Schnorrer, F. RNA Interference Screening for Genes Regulating Drosophila Muscle Morphogenesis. Myogenesis. 1889, 331-348 (2019).
- Chechenova, M. B., et al. Functional redundancy and non-redundancy between two Troponin C isoforms in Drosophila adult muscles. Molecular Biology of the Cell. 28 (6), 760-770 (2017).
- Alberts, B. . Molecular Biology of the Cell. , (2017).
- Clyne, P. J., Brotman, J. S., Sweeney, S. T., Davis, G. Green fluorescent protein tagging Drosophila proteins at their native genomic loci with small P elements. Genetics. 165 (3), 1433-1441 (2003).
- Orfanos, Z., Sparrow, J. C. Myosin isoform switching during assembly of the Drosophila flight muscle thick filament lattice. Journal of Cell Science. 126 (1), 139-148 (2013).
- Volohonsky, G., Edenfeld, G., Klambt, C., Volk, T. Muscle-dependent maturation of tendon cells is induced by post-transcriptional regulation of stripeA. Development. 134 (2), 347-356 (2007).
- Estes, P. S., Ho, G. L., Narayanan, R., Ramaswami, M. Synaptic localization and restricted diffusion of a Drosophila neuronal synaptobrevin--green fluorescent protein chimera in vivo. Journal of Neurogenetics. 13 (4), 233-255 (2000).
- Hida, N., et al. EC-tagging allows cell type-specific RNA analysis. Nucleic Acids Research. 45 (15), 138 (2017).
- Thomas, A., et al. A versatile method for cell-specific profiling of translated mRNAs in Drosophila. PLoS ONE. 7 (7), 40276 (2012).
- Yang, Z. Isolation of mRNA from specific tissues of Drosophila by mRNA tagging. Nucleic Acids Research. 33 (17), 148 (2005).
- Jiao, Y., Moon, S. J., Montell, C. A Drosophila gustatory receptor required for the responses to sucrose, glucose, and maltose identified by mRNA tagging. Proceedings of the National Academy of Sciences of the United States of America. 104 (35), 14110-14115 (2007).
- Blazie, S. M., et al. Comparative RNA-Seq analysis reveals pervasive tissue-specific alternative polyadenylation in Caenorhabditis elegans intestine and muscles. BMC Biology. 13 (1), 1775-1821 (2015).
- Henry, G. L., Davis, F. P., Picard, S., Eddy, S. R. Cell type-specific genomics of Drosophila neurons. Nucleic Acids Research. 40 (19), 9691-9704 (2012).
- Deal, R. B., Henikoff, S. The INTACT method for cell type-specific gene expression and chromatin profiling in Arabidopsis thaliana. Nature Protocols. 6 (1), 56-68 (2011).
- Grinstein, M., Dingwall, H. L., Shah, R. R., Capellini, T. D., Galloway, J. L. A robust method for RNA extraction and purification from a single adult mouse tendon. PeerJ. 6 (8), 4664 (2018).
- Green, M. R., Sambrook, J. . Molecular Cloning. , (2012).
- Brown, R. A. M., et al. Total RNA extraction from tissues for microRNA and target gene expression analysis: not all kits are created equal. BMC Biotechnology. 18 (1), 16 (2018).
- Ford, K. L., et al. Optimisation of laboratory methods for whole transcriptomic RNA analyses in human left ventricular biopsies and blood samples of clinical relevance. PLoS ONE. 14 (3), 02136855 (2019).
- Ramachandran, P., Budnik, V. Dissection of Drosophila larval body-wall muscles. Cold Spring Harbor Protocols. (8), 5469 (2010).
- Ranganayakulu, G., Schulz, R. A., Olson, E. N. Wingless signaling induces nautilus expression in the ventral mesoderm of the Drosophila embryo. Developmental Biology. 176 (1), 143-148 (1996).
- Dutta, D., Bloor, J. W., Ruiz-Gómez, M., VijayRaghavan, K., Kiehart, D. P. Real-time imaging of morphogenetic movements in Drosophila using Gal4-UAS-driven expression of GFP fused to the actin-binding domain of moesin. Genesis. 34 (1-2), 146-151 (2002).
- Lemke, S. B., Schnorrer, F. In Vivo Imaging of Muscle-tendon Morphogenesis in Drosophila Pupae. Journal of Visualized Experiments. (132), e57312 (2018).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved