
A subscription to JoVE is required to view this content. Sign in or start your free trial.
Method Article
مطيافية نيوترونية عالية الدقة لدراسة ديناميكيات بيكو ثانية-نانوثانية للبروتينات ومياه الترطيب
In This Article
Summary
يوفر التحليل الطيفي للتشتت العكسي للنيوترونات وصولا غير مدمر وخالي من الملصقات إلى ديناميكيات ps-ns للبروتينات ومياه ترطيبها. يتم تقديم سير العمل مع دراستين حول بروتينات الأميلويد: حول ديناميكيات الليزوزيم التي تم حلها زمنيا أثناء التجميع وعلى ديناميكيات الماء المائية لتاو عند تكوين الألياف.
Abstract
يوفر تشتت النيوترونات إمكانية سبر الديناميكيات داخل العينات لمجموعة واسعة من الطاقات بطريقة غير مدمرة وبدون تسمية بخلاف الديوتيريوم. على وجه الخصوص ، يسجل التحليل الطيفي للتشتت العكسي للنيوترونات إشارات التشتت بزوايا تشتت متعددة في وقت واحد وهو مناسب تماما لدراسة ديناميكيات الأنظمة البيولوجية على مقياس ps-ns الزمني. من خلال استخدام D2O - وربما مكونات عازلة متوقفة - تسمح الطريقة بمراقبة كل من انتشار مركز الكتلة وحركات العمود الفقري والسلسلة الجانبية (الديناميات الداخلية) للبروتينات في الحالة السائلة.
بالإضافة إلى ذلك ، يمكن دراسة ديناميكيات مياه الترطيب عن طريق استخدام مساحيق من البروتينات المثبطة المرطبة ب H2O. تقدم هذه الورقة سير العمل المستخدم على الأداة IN16B في معهد Laue-Langevin (ILL) للتحقيق في ديناميكيات البروتين والماء المائي. يتم شرح تحضير عينات المحلول وعينات مسحوق البروتين المائي باستخدام تبادل البخار. يتم وصف إجراء تحليل البيانات لكل من ديناميات البروتين والماء الإماهة لأنواع مختلفة من مجموعات البيانات (الأطياف شبه المرنة أو عمليات المسح ذات النافذة الثابتة) التي يمكن الحصول عليها على مطياف التشتت العكسي للنيوترونات.
يتم توضيح الطريقة من خلال دراستين تتضمنان بروتينات الأميلويد. يظهر أن تجميع الليزوزيم إلى مجاميع كروية بحجم ميكرومتر - يشار إليها بالجسيمات - يحدث في عملية من خطوة واحدة على النطاق المكاني والزمني الذي تم فحصه في IN16B ، بينما تظل الديناميكيات الداخلية دون تغيير. علاوة على ذلك ، تمت دراسة ديناميات ماء الترطيب في تاو على مساحيق رطبة من البروتين perdeuteated. يتبين أن الحركات الانتقالية للماء يتم تنشيطها عند تكوين ألياف الأميلويد. أخيرا ، تتم مناقشة الخطوات الحاسمة في البروتوكول حول كيفية وضع تشتت النيوترونات فيما يتعلق بدراسة الديناميات فيما يتعلق بالطرق الفيزيائية الحيوية التجريبية الأخرى.
Introduction
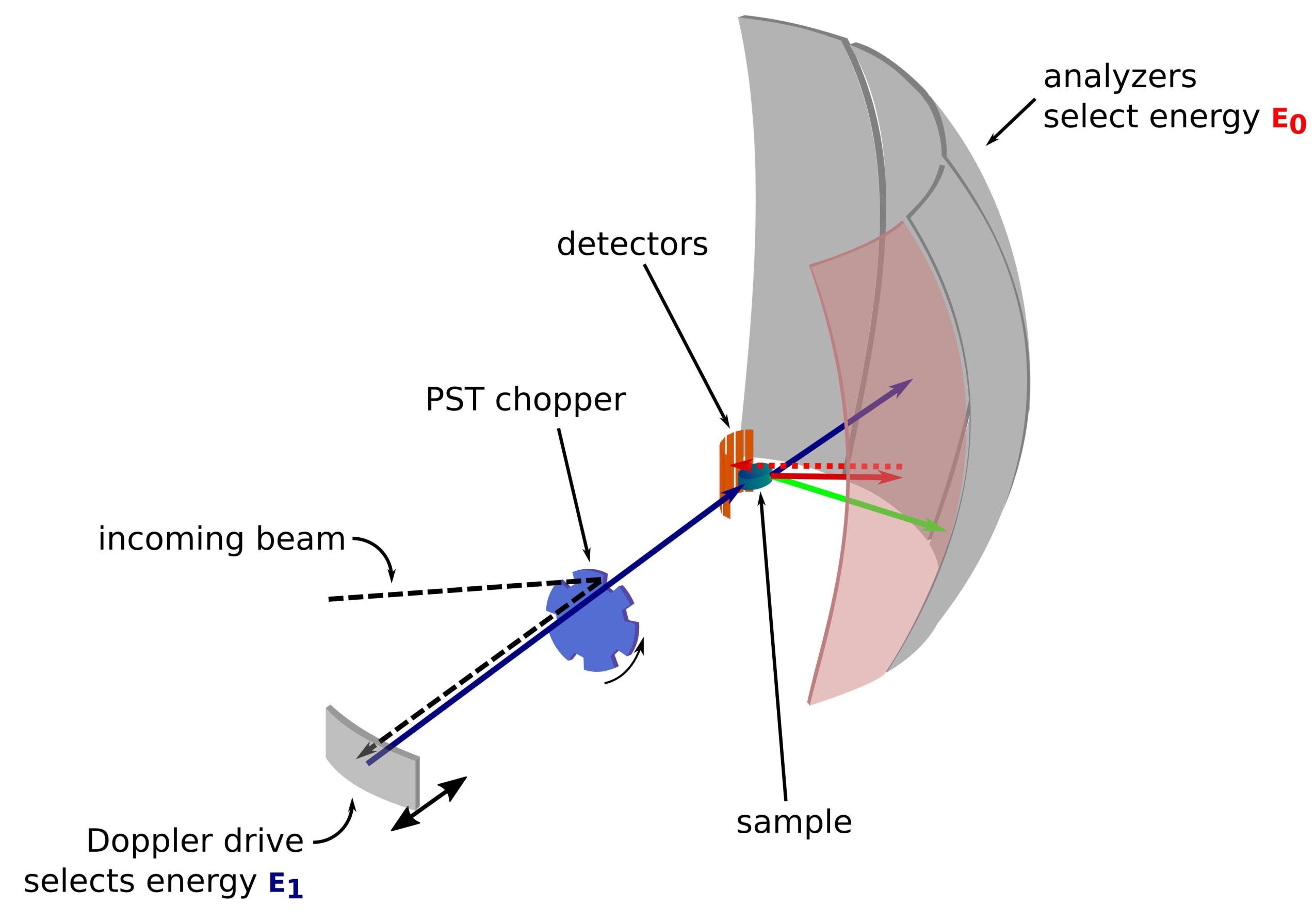
النيوترون هو جسيم ضخم بدون شحنة تم استخدامه بنجاح على مر السنين لفحص العينات في مختلف المجالات من الفيزياء الأساسية إلى علم الأحياء1. بالنسبة للتطبيقات البيولوجية ، يتم استخدام تشتت النيوترونات بزاوية صغيرة ، وتشتت النيوترونات غير المرن ، وعلم البلورات النيوتروني وقياس الانعكاس على نطاق واسع2،3،4. يوفر تشتت النيوترونات غير المرن قياسا متوسطا للديناميكيات دون الحاجة إلى وضع علامات محددة في حد ذاتها ، وجودة إشارة لا تعتمد على الحجم أو البروتين5. يمكن إجراء القياس باستخدام بيئة معقدة للغاية للبروتين قيد الدراسة الذي يحاكي الوسط داخل الخلايا ، مثل المحللة البكتيرية المنزوعة أو حتى في الجسم الحي3،6،7. يمكن استخدام إعدادات تجريبية مختلفة لدراسة الديناميات ، وهي i) وقت الرحلة الذي يمنح الوصول إلى ديناميكيات ps-ps الفرعية ، ii) الوصول إلى التشتت العكسي إلى ديناميكيات ps-ns ، و iii) الوصول إلى الصدى المغزلي إلى الديناميكيات من ns إلى مئات ns. يستخدم التشتت العكسي للنيوترونات قانون براغ 2d sinθ = nλ ، حيث d هي المسافة بين المستويات في البلورة ، و θ زاوية التشتت ، و n ترتيب التشتت ، و λ الطول الموجي. يسمح استخدام البلورات للتشتت العكسي نحو أجهزة الكشف بتحقيق دقة عالية في الطاقة ، عادة ~ 0.8 μeV. لقياس تبادل الطاقة ، يتم استخدام إما محرك دوبلر يحمل بلورة في التشتت العكسي لتحديد وضبط الطول الموجي النيوتروني الوارد8،9،10 (الشكل 1) ، أو يمكن استخدام إعداد وقت الرحلة على حساب انخفاض في دقة الطاقة11.

الشكل 1: رسم مطياف التشتت العكسي للنيوترونات بمحرك دوبلر. يصطدم الشعاع الوارد بمروحية تحويل فضاء الطور (PST)42 ، مما يزيد من التدفق في موضع العينة. ثم يتم تشتيتها مرة أخرى نحو العينة بواسطة محرك دوبلر ، الذي يختار الطاقة E1 (السهم السماوي). ثم يتم تشتيت النيوترونات بواسطة العينة (مع طاقات مختلفة ممثلة بلون الأسهم) والمحللين ، المصنوع من بلورات Si 111 ، سوف ينتشر فقط النيوترونات ذات الطاقة المحددة E0 (الأسهم ذات اللون الأحمر هنا). ومن ثم ، يتم الحصول على نقل الزخم q من الموضع المكتشف للنيوترون على صفيف الكاشف ، ويتم الحصول على نقل الطاقة من الفرق E1- E0. يتم استخدام وقت الرحلة المتوقع لنبضة النيوترون التي تنتجها PST لتجاهل الإشارة من النيوترونات المنتشرة مباشرة نحو أنابيب الكاشف. اختصار: PST = تحويل مساحة الطور. يرجى النقر هنا لعرض نسخة أكبر من هذا الرقم.
{kind=link}
بالنسبة للتحليل الطيفي للتشتت العكسي ، تأتي المساهمة الرئيسية في الإشارة من العينات الغنية ببروتون الهيدروجين ، مثل البروتينات ، من التشتت غير المترابط ، حيث تظهر شدة التشتت Sinc (q ، ω) بواسطة Eq (1) 12
 (1)
(1)
حيث σinc هو المقطع العرضي غير المترابط للعنصر المعتبر ، k 'هو معيار المتجه الموجي المتناثر ، k هو معيار المتجه الموجي الوارد ، q (= k - k') نقل الزخم ، r j (t) متجه موضع الذرة j في الوقت t ، و ω التردد المقابل لنقل الطاقة بين النيوترون الوارد والنظام. تشير الأقواس الزاوية إلى متوسط المجموعة. ومن ثم ، فإن التشتت غير المترابط يسبر الارتباط الذاتي للجسيم الواحد متوسط المجموعة لمواقع الذرة مع الوقت ويعطي متوسط الديناميكيات الذاتية على جميع الذرات في النظام وأصول زمنية مختلفة (متوسط المجموعة). دالة التشتت هي تحويل فورييه في زمن دالة التشتت الوسيطة I (q، t) ، والتي يمكن اعتبارها تحويل فورييه في الفضاء لدالة ارتباط فان هوف الموضحة بواسطة Eq (2):
 (2)
(2)
حيث ρ (r، t) هي كثافة احتمال إيجاد ذرة في الموضع r والوقت t 13.
بالنسبة لعملية الانتشار الويكي ، تنتج دالة الانتشار الذاتي (انظر Eq (3)) بعد تحويل فورييه المزدوج في دالة تشتت تتكون من لورنتزيان بعرض الخط المعطى بواسطة γ = Dq2.
 (3)
(3)
تم تطوير نماذج أكثر تطورا ووجدت مفيدة مثل نموذج انتشار القفز بواسطة Singwi و Sjölander لديناميات البروتين الداخليةps-ns 14 أو نموذج الدوران بواسطة Sears لمياه الترطيب15،16،17.
على أداة التشتت العكسي للنيوترونات (NBS) IN16B 8,9 في ILL، غرونوبل، فرنسا (الشكل التكميلي S1)، يتكون الإعداد الذي يشيع استخدامه مع البروتينات من بلورات Si 111 للمحللات مع محرك دوبلر لضبط الطول الموجي الوارد (الشكل التكميلي S2A)، مما يتيح الوصول إلى نطاق نقل الزخم ~ 0.2 Å-1 < q < ~ 2 Å-1 ونطاق نقل الطاقة -30 μeV <  < 30 μeV المقابلة لجداول زمنية تتراوح من بضعة ps إلى عدد قليل من ns ومسافات قليلة Å. بالإضافة إلى ذلك ، يوفر IN16B إمكانية إجراء عمليات مسح مرنة وغير مرنة للنافذة الثابتة (E / IFWS) 10 ، والتي تشمل الحصول على البيانات عند نقل الطاقة الثابتة. نظرا لأن التدفق محدود عند العمل مع النيوترونات ، فإن E / IFWS يسمح بتعظيم التدفق لنقل طاقة واحد ، وبالتالي تقليل وقت الاكتساب اللازم للحصول على نسبة إشارة إلى ضوضاء مرضية. والخيار الأحدث هو النمط11 لمطياف التشتت العكسي ووقت الطيران (BATS) ، والذي يسمح بقياس مجموعة واسعة من عمليات نقل الطاقة ، (على سبيل المثال ، -150 μeV < < 150 μeV) ، مع تدفق أعلى من محرك دوبلر ، ولكن على حساب دقة طاقة أقل (الشكل التكميلي S2B).
< 30 μeV المقابلة لجداول زمنية تتراوح من بضعة ps إلى عدد قليل من ns ومسافات قليلة Å. بالإضافة إلى ذلك ، يوفر IN16B إمكانية إجراء عمليات مسح مرنة وغير مرنة للنافذة الثابتة (E / IFWS) 10 ، والتي تشمل الحصول على البيانات عند نقل الطاقة الثابتة. نظرا لأن التدفق محدود عند العمل مع النيوترونات ، فإن E / IFWS يسمح بتعظيم التدفق لنقل طاقة واحد ، وبالتالي تقليل وقت الاكتساب اللازم للحصول على نسبة إشارة إلى ضوضاء مرضية. والخيار الأحدث هو النمط11 لمطياف التشتت العكسي ووقت الطيران (BATS) ، والذي يسمح بقياس مجموعة واسعة من عمليات نقل الطاقة ، (على سبيل المثال ، -150 μeV < < 150 μeV) ، مع تدفق أعلى من محرك دوبلر ، ولكن على حساب دقة طاقة أقل (الشكل التكميلي S2B).
من الخصائص المهمة لتشتت النيوترونات أن المقطع العرضي غير المترابط σinc له قيمة أعلى 40 مرة للهيدروجين من الديوتيريوم ولا يكاد يذكر بالنسبة للعناصر الأخرى التي توجد عادة في العينات البيولوجية. لذلك ، يمكن دراسة ديناميكيات البروتينات في بيئة سائلة باستخدام مخزن مؤقت مقسم ، وتسمح حالة المسحوق بدراسة إما الديناميات الداخلية للبروتين مع مسحوق البروتين المهدرج المبلل ب D 2 O ، أودراسة ماء الترطيب لمسحوق البروتين perdeuterated المائي باستخدام H2O. في الحالة السائلة ، يسمح التشتت العكسي للنيوترونات عادة بالوصول في وقت واحد إلى الانتشار الذاتي لمركز الكتلة للبروتينات (الانتشار من نوع Fickian) وديناميكياتها الداخلية. هذه الأخيرة هي حركات العمود الفقري والسلسلة الجانبية التي يصفها عادة ما يسمى نموذج انتشار القفز أو غيره 3,18. في مساحيق البروتين المهدرجة ، يكون انتشار البروتين غائبا ويجب نمذجة الديناميات الداخلية فقط. بالنسبة لمياه الإماهة ، فإن مساهمات الحركات الانتقالية والدورانية لجزيئات الماء تقدم اعتمادا مختلفا على نقل الزخم q ، مما يسمح بتمييزها في عملية تحليل البيانات17.
توضح هذه الورقة طريقة التشتت العكسي للنيوترونات مع دراسة البروتينات التي وجد أنها قادرة على التكشف ، وتتجمع في شكل قانوني يتكون من أكوام من خيوط β - ما يسمى بنمط β المتقاطع19،20 - وتشكل أليافا ممدودة. هذا هو ما يسمى بتجميع الأميلويد ، والذي تمت دراسته على نطاق واسع بسبب دوره المركزي في الاضطرابات التنكسية العصبية مثل مرض الزهايمر أو مرض باركنسون21,22. إن دراسة بروتينات الأميلويد مدفوعة أيضا بالدور الوظيفي الذي يمكن أن تلعبه 23,24 أو إمكاناتها العالية لتطوير مواد حيوية جديدة25. لا تزال المحددات الفيزيائية والكيميائية لتجميع الأميلويد غير واضحة ، ولا تتوفر نظرية عامة لتجميع الأميلويد ، على الرغم من التقدم الهائل خلال السنوات الماضية21,26.
ينطوي تجميع الأميلويد على تغييرات في بنية البروتين واستقراره مع مرور الوقت ، والتي تنطوي دراستها بشكل طبيعي على ديناميكيات مرتبطة باستقرار تكوين البروتين ، ووظيفة البروتين ، ومشهد طاقة البروتين27. ترتبط الديناميكيات ارتباطا مباشرا باستقرار حالة معينة من خلال المساهمة الإنتروبية لأسرع الحركات28 ، ويمكن الحفاظ على وظيفة البروتين من خلال الحركات على نطاقات زمنية مختلفة من sub-ps للبروتينات الحساسة للضوء29 إلى ms لحركات المجال ، والتي يمكن تسهيلها بواسطة ديناميكيات بيكو ثانيةنانو ثانية 30.
سيتم تقديم مثالين على استخدام التحليل الطيفي للتشتت العكسي للنيوترونات لدراسة بروتينات الأميلويد ، أحدهما في الحالة السائلة لدراسة ديناميكيات البروتين والآخر في حالة المسحوق المائي لدراسة ديناميكيات الماء المماهة. يتعلق المثال الأول بتجميع الليزوزيم في كرات بحجم ميكرومتر (تسمى الجسيمات) متبوعة في الوقت الفعلي5 ، والثاني مقارنة لديناميكيات المياه في الحالات الأصلية والمجمعة للبروتين البشري تاو31.
الليزوزيم هو إنزيم يشارك في الدفاع المناعي ويتكون من 129 من بقايا الأحماض الأمينية. يمكن أن يشكل الليزوزيم جسيمات في المخزن المؤقت المثبط عند pD 10.5 وعند درجة حرارة 90 درجة مئوية. مع تشتت النيوترونات ، أظهرنا أن التطور الزمني لمعامل انتشار مركز الكتلة للليزوزيم يتبع الحركية الأسية المفردة لتألق ثيوفلافين T (مسبار فلوري يستخدم لمراقبة تكوين أنماط β المتقاطع الأميلويد32) ، مما يشير إلى أن الهياكل الفوقية للجسيمات وأنماط β المتقاطع تحدث في خطوة واحدة بنفس المعدل. علاوة على ذلك ، ظلت الديناميات الداخلية ثابتة طوال عملية التجميع ، والتي يمكن تفسيرها إما من خلال تغيير توافقي سريع لا يمكن ملاحظته على أدوات NBS ، أو من خلال عدم وجود تغيير كبير في الطاقة الداخلية للبروتين عند التجميع.
البروتين البشري تاو هو بروتين مضطرب جوهريا (IDP) يتكون من 441 من الأحماض الأمينية لما يسمى ب 2N4R isoform ، والذي يشارك بشكل خاص في مرض الزهايمر33. باستخدام التشتت العكسي للنيوترونات على مساحيق بروتين تاو perdeuterated ، أظهرنا أن ديناميكيات الماء المائيات تزداد في حالة الألياف ، مع وجود عدد أكبر من جزيئات الماء التي تخضع لحركات متعدية. تشير النتيجة إلى أن الزيادة في إنتروبيا الماء المائي قد تؤدي إلى رجفان الأميلويد في تاو.
Protocol
1. إعداد المخزن المؤقت deuterated للبروتينات في الحالة السائلة
- حل جميع مكونات المخزن المؤقت في D2O النقي.
- إذا تمت معايرة قطب الأس الهيدروجيني في H2O ، فاضبط pD وفقا للصيغة pD = pH + 0.4 باستخدام NaOD أو DCl34.
ملاحظة: قد يؤثر استخدام D 2O بدلا من H2O على قابلية ذوبان البروتين وقد تحتاج الظروف العازلة إلى التكيف (على سبيل المثال ، عن طريق تغيير طفيف في تركيز الملح).
2. تحضير مساحيق H2O المائية من البروتين perdeuterated
- تحضير حامل العينة.
- قم بتنظيف حامل عينة الألمنيوم المسطح تماما باستخدام ختم سلك الإنديوم والبراغي بالماء والإيثانول واتركه يجف.
ملاحظة: يتم استخدام حامل عينة مسطح بحيث يمكن توزيع المسحوق بشكل متجانس على السطح. يجب أن تكون كمية المسحوق كافية بحيث يمكن الحفاظ عليها بين الجدران ولا تسقط عند وضع حامل العينة عموديا. - قم بوزن الأجزاء المختلفة من قاع حامل العينة والغطاء وسلك الإنديوم بشكل منفصل على ميزان دقيق.
- ضع ختم سلك الإنديوم 1 مم في أخدود الجزء السفلي من حامل العينة ، تاركا تداخلا صغيرا حيث ينضم الطرفان (الشكل 2 أ).
- ضع كمية مناسبة من البروتين المجفف بالتجميد (عادة ~ 100 ملغ من البروتين) بحيث يملأ السطح الداخلي للجزء السفلي من حامل العينة.
- قم بتنظيف حامل عينة الألمنيوم المسطح تماما باستخدام ختم سلك الإنديوم والبراغي بالماء والإيثانول واتركه يجف.
- رطب مسحوق البروتين.
- ضع حامل العينة في مجفف مع طبق بتري يحتوي على مسحوق P2O5 لمدة 24 ساعة لتجفيف مسحوق البروتين تماما35 (الشكل 2B). قم بوزن الجزء السفلي الجاف من حامل العينة الذي يحتوي على ختم الإنديوم والمسحوق الجاف للحصول علىم جاف.
تنبيه: مسحوق P2O5 تآكل للغاية. - أخرج P 2 O5 من المجفف وضع طبق بتري مع D2O بالداخل. تحكم في كتلة المسحوق بانتظام للتحقق من مستوى الترطيب h = m hyd / m الجاف حيث mhyd و mdry هما كتلة المسحوق المائي والمسحوقالجاف ، على التوالي.
ملاحظة: بالنسبة للبروتينات الكارهة للماء مثل الأنسولين ، قد يكون من الضروري زيادة درجة الحرارة داخل المجفف للحصول على ضغط بخار أعلى والوصول إلى مستوى الترطيب المطلوب h. - كرر الخطوتين 2.2.1 و 2.2.2 ثلاث مرات على الأقل لتحويل جميع الهيدروجين القابل للتبديل إلى الديوترونات بشكل صحيح.
ملاحظة: بدلا من ذلك ، يمكن استخدام دورات التجفيف بالتجميد والذوبان في D2O النقي لتحسين تبادل H / D بشرط ألا يتأثر البروتين به. - قم بترطيب المسحوق إلى أعلى قليلا من المستوى المطلوب ، واترك الجزء السفلي من حامل العينة بسلك الإنديوم والمسحوق المائي على ميزان الدقة ، وانتظر حتى تنخفض الكتلة ببطء إلى القيمة المطلوبة للحصول على الهدف h (عادة 0.2-0.4 إذا كان البروتين الكروي متوسط الحجم سيتم تغطيته بطبقة ترطيب كاملة واحدة).
- ضع الغطاء بسرعة على الجزء السفلي وأغلق حامل العينة أولا بأربعة براغي لإيقاف تبادل البخار (الشكل التكميلي S3A).
- ضع جميع البراغي المتبقية وأحكمها حتى لا تظهر فجوة بين الجزء السفلي والغطاء (الشكل التكميلي S3B).
- قم بوزن حامل العينة المختوم للتحقق من أي فقد محتمل للترطيب عن طريق التسريبات بعد تجربة النيوترونات.
- ضع حامل العينة في مجفف مع طبق بتري يحتوي على مسحوق P2O5 لمدة 24 ساعة لتجفيف مسحوق البروتين تماما35 (الشكل 2B). قم بوزن الجزء السفلي الجاف من حامل العينة الذي يحتوي على ختم الإنديوم والمسحوق الجاف للحصول علىم جاف.
3. إجراء تجربة تشتت النيوترونات غير المتماسكة
- ناقش وتحقق مرة أخرى من تكوين الأداة اللازمة للتجربة مع جهة الاتصال المحلية قبل بضعة أسابيع من وقت الحزمة المعين.
- تحضير عينة الحالة السائلة.
- حل البروتين في المخزن المؤقت deuteated.
- تحديد الحجم المناسب للسائل الذي سيتم وضعه في حامل العينة باستخدام الماء (تأكد من عدم وجود فائض عند إغلاق حامل العينة ؛ الشكل 2 ج).
ملاحظة: تصف الخطوتان التاليتان (3.3 و 3.4) تجربة أجريت على مطياف NBS IN16B في ILL 8,9 ، باستخدام فرن التبريد كبيئة عينة. سيتغير نظام التحكم في الأداة من أداة إلى أخرى ، لكن مبادئ العمل تظل كما هي.
- أدخل العينة.
- جفف عصا العينة تماما (الشكل 2D) ، وأزل العينة السابقة ، إن وجدت ، بعد التحقق من أن جرعة الإشعاع المؤين أقل من 100 μSv / h قبل التعامل مع أي مادة (في ILL).
- ضع العينة ، وتحقق من التمركز المناسب بالنسبة لمركز الحزمة (الشكل التكميلي S4) ، وأدخل عصا العينة في فرن التبريد (الشكل 2D). قم بتشغيل مضخة التفريغ للوصول إلى أقل من 10-3 بار ، واطرد الهواء داخل فرن التبريد عن طريق تكرار ثلاث مرات التالية: املأ فرن التبريد بغاز الهيليوم حتى يتم الوصول إلى الضغط الجوي ، وقم بإزالة الغاز مرة أخرى باستخدام مضخة التفريغ.
ملاحظة: في حالة حامل العينة المسطح ، يجب توجيه حامل العينة بزاوية 45 درجة بالنسبة إلى الحزمة الواردة. يمكن تقليل نطاق نقل الزخم المفيد بسبب الامتصاص والتشتت بواسطة الخلية. يمكن استخدام ممتص نيوتروني قوي مثل الكادميوم لإخفاء أجزاء معينة من حامل العينة (على سبيل المثال ، البراغي والأجزاء السميكة). - أدخل بعض غاز الهيليوم في فرن التبريد بحيث يكون الضغط ~ 0.05 بار.
- الحصول على البيانات (على سبيل المثال ، باستخدام NOMAD على IN16B في ILL ، يفترض أن المستخدم يفضل درجة حرارة 200 K قبل الحصول على طيف طيف نيوتروني شبه مرن (QENS) ، ثم E / IFWS أثناء منحدر درجة الحرارة إلى 310 K عند 0.5 K في الدقيقة وأخيرا QENS عند 310 K).
- باستخدام NOMAD ، في علامة تبويب التنفيذ ، اسحب وحدة تحكم FurnaceCryostat وأفلتها في لوحة التشغيل. اضبط درجة الحرارة على 200 كلفن. استخدم الوضع السريع ومهلة 30 دقيقة بحيث يكون لدرجة الحرارة وقت للاستقرار. انقر على أيقونة الأسهم الدوارة لتشغيلها في الخلفية بحيث يمكن الحصول على البيانات أثناء انخفاض درجة الحرارة.
- اسحب وحدة التحكم IN16DopplerSettings وأفلتها، واضبط ملف تعريف السرعة على السرعة الدقيقة التي تم تعيينها بواسطة Max ΔE، وقيمة 0.00 μeV و128 قناة للحصول على تكوين EFWS.
- قم بسحب وإسقاط وحدة تحكم العد ، واملأ حقل العنوان الفرعي باسم يسمح بسهولة التعرف على البيانات ، وقم بتعيين 60 تكرارا لمسح 30 ثانية (الشكل التكميلي S5A).
- اسحب وحدة تحكم IN16DopplerSettings وأفلتها، واضبط ملف تعريف السرعة على Sine الذي تم تعيينه بواسطة Speed بقيمة 4.5 m/s و2048 قناة للحصول على تكوين QENS.
- قم بسحب وإسقاط وحدة تحكم العد مع 4 تكرارات لمدة 30 دقيقة من عمليات المسح الضوئي (الشكل التكميلي S5B).
- بالنسبة لمنحدر درجة الحرارة ، اسحب وحدة تحكم FurnaceCryostat وأفلتها ، واضبط درجة الحرارة على 310 K ، واضبط المنحدر على SetPoint مع Δ = 0.05 K و 6 ثوان. استخدم وقتا من 220 دقيقة (الشكل التكميلي S6A).
- استخدم حلقة for مع 65 تكرارا. في الداخل ، أدخل وحدة تحكم IN16DopplerSettings كما في الخطوة 3.4.2 ، متبوعة بعدد واحد يبلغ 30 ثانية. بعد ذلك ، أدخل IN16DopplerSettings ، كما هو موضح سابقا ولكن باستخدام إزاحة طاقة تبلغ 1.5 μeV و 1024 قناة متبوعة بعدد واحد قدره 3 دقائق (الشكل التكميلي S6B).
- للحصول على آخر QENS عند 310 كلفن، قم بسحب وإفلات وحدات التحكم IN16DopplerSettings وCount التي تم تكوينها كما هو موضح في الخطوتين 3.4.4 و3.4.5 على التوالي.
- اضغط على زر البدء (مثلث قائم الزاوية في أسفل النافذة) لتشغيل البرنامج النصي.
ملاحظة: ستتطلب كل تجربة الحصول على بيانات المعايرة. أي الخلية الفارغة لتصحيحات الطرح أو الامتصاص ، والمخزن المؤقت وحده في درجات الحرارة المختلفة المستخدمة لنمذجة الخلفية ، وقياس الفاناديوم (أو ما يعادله ، العينة عند درجة حرارة 10 كلفن أو أقل) للحصول على وظيفة دقة الجهاز.
4. تحليل البيانات - QENS
- قم باستيراد مجموعة البيانات باستخدام طريقة 'IN16B_QENS.process()' في برنامج Python nPDyn v3.x36
>>>> من nPDyn.dataParsers استيراد IN16B_QENS
>>> العينة = IN16B_QENS(
... <المسار إلى ملفات البيانات>
... [detGroup = < عدد صحيح أو ملف تجميع كاشف في XML
... الشكل>]
... ). عملية ()
>>> عينة = sample.get_q_range (0.3 ، 1.8) - قم بإجراء تصحيحات البيانات (اختياري) باستخدام الأوامر التالية (انظر وثائق nPDyn لمزيد من المعلومات ، الشكل 3):
يفترض #it أن بيانات الخلية الفارغة والفاناديوم والمخزن المؤقت
# تم استيرادها بالفعل في مجموعة بيانات تسمى "empty_cell" ، "الفاناديوم" ،
# و "المخزن المؤقت" ، على التوالي.
# لطرح الخلية الفارغة بعامل تحجيم
# (يتم نشر الأخطاء تلقائيا)
>>> عينة = عينة - 0.95 * empty_cell
# للتصحيح باستخدام معامل بالمان بينغ
# (حصري بشكل متبادل مع المثال أعلاه)
>>> العينة = sample.absorptionCorrection(empty_cell)
# للتطبيع
>>> عينة = عينة.تطبيع (الفاناديوم)
# للتجميع على طول المحور الذي يمكن ملاحظته
# يمكن ملاحظته هو وقت التجميع هنا
>>> العينة = العينة .bin (3 ، المحور = 0) - تناسب بيانات المعايرة. يمكن تركيب عينات مجموعة البيانات والخلية الفارغة والمخزن المؤقت المخفف (إذا لزم الأمر) والفاناديوم باستخدام نماذج مدمجة أو نموذج محدد من قبل المستخدم (انظر وثائق nPDyn):
>>> من استيراد nPDyn.models.builtins (
... نموذجPVoigt,
... نموذجالمياه,
... نموذجمعايرةD2O,
... )
# تستخدم النماذج المدمجة متجه عمود للزخم
# نقل قيم q
>>> q = الفاناديوم.q[:, لا يوجد]
# يتم تركيب الفاناديوم باستخدام ملف تعريف زائف Voigt
>>> الفاناديوم.فيت (نموذجPVoigt (ف)) - استخدم النموذج المدمج لمياه الترطيب المسمى "modelWater". يقرأ هذا النموذج كما هو موضح بواسطة مكافئ (4) 17
 (4)
(4)
حيث0 وr وt هي أعداد قياسية تمثل المساهمة النسبية للإشارة المرنة والحركات الدورانية والحركات الانتقالية ، على التوالي ؛ j1 (qd) هي دالةبيسل الكروية من الدرجة l th ، مع q هي نقل الزخم ؛ د مسافة O-H في جزيء الماء ؛ δ (ω) هي دلتا ديراك ، والتي يتم ضربها في EISF هنا ؛ N هو أعلى ترتيب لدالة Bessel الكروية المستخدمة (عادة ~ 5) ؛ وهي
وهي  الحركات الدورانية والانتقالية لورنتزيان ، على التوالي ؛ ب (ف) هو مصطلح خلفية مسطح. تعطي دوال Bessel الكروية المساهمة النسبية لكل حالة زخم زاوية لجزيئات الماء ، ويتم تحديد الرقم N بناء على نطاق q لنقل الزخم. في حالة مطياف NBS النموذجي ، فإن المصطلحات حتى N = 4 تشرح الإشارة بالكامل تقريبا (الشكل التكميلي S7).
الحركات الدورانية والانتقالية لورنتزيان ، على التوالي ؛ ب (ف) هو مصطلح خلفية مسطح. تعطي دوال Bessel الكروية المساهمة النسبية لكل حالة زخم زاوية لجزيئات الماء ، ويتم تحديد الرقم N بناء على نطاق q لنقل الزخم. في حالة مطياف NBS النموذجي ، فإن المصطلحات حتى N = 4 تشرح الإشارة بالكامل تقريبا (الشكل التكميلي S7).
# هنا ، يتم استخدام المعادلة 2 لمياه الترطيب
# الالتفاف مع وظيفة القرار وإضافة
# تتم خلفية D2O تلقائيا باستخدام ملف
# الحجج المقدمة
>>> sample.fit (modelWater (q) ،
... الدقة = الفاناديوم ،
... bkgd = العازلة ،
... volume_fraction_bkgd=0.95
... )
ملاحظة: يجب أن تكون مساهمات الحركات الدورانية والانتقالية معقدة لتكون صارمة تماما. يعزى نجاح النموذج الإضافي إلى وجود مجموعات متميزة من الماء على سطح البروتين ونطاق الطاقة المحدود الذي يمكن الوصول إليه. - استخدم ما يلي لرسم البيانات (الشكل 4):
>>> من قطعة أرض استيراد nPDyn.plot
مؤامرة >>> (عينة)
5. تحليل البيانات - منحدر درجة الحرارة ، مسح النافذة الثابتة المرنة (EFWS)
- استخدم إجراء مشابها للقسم 4 لتطبيع بيانات منحدر درجة الحرارة بواسطة الإشارة عند أدنى درجة حرارة (عادة 10 كلفن):
>>> من nPDyn.dataParsers استيراد IN16B_FWS
>>> عينة = IN16B_FWS(
... <المسار إلى ملفات البيانات> ،
... detGroup=[detGroup=<ملف تجميع عدد صحيح أو كاشف بتنسيق XML>]
... ). عملية ()
# التطبيع مع النقاط ال 5 الأولى على ما يمكن ملاحظته
# المحور ، والتي تتوافق مع درجة الحرارة
>>> عينة /= عينة [:5].يعني (0)
# نطاق Q لنقل الزخم المستخدم هنا أصغر
# حيث أن النموذج المستخدم صالح ل Low Q فقط
>>> عينة = sample.get_q_range (0.2 ، 0.8) - استخدم نموذجا غاوسيا بسيطا للبدء ، يتم إعطاء عرضه بواسطة ما يسمى بمتوسط الإزاحة التربيعية (MSD). قم ببناء النموذج وملاءمته باستخدام الأوامر التالية:
>>> استيراد numpy ك np
>>> من nPDyn.models استيراد المعلمات والنموذج والمكون
# أ هو عامل التحجيم
>>> المعلمات = المعلمات (
... a={'value': 1، 'bounds': (0, np.inf)},
... msd = {'القيمة': 1 ، 'الحدود': (0 ، np.inf)}
... )
>>> نموذج = نموذج (معلمات)
>>> model.addComponent(Component(
... "غاوسي" ،
... لامدا س ، أ ، إم إس دي: أ * NP.exp (-x ** 2 * MSD / 6)
... ))
>>> sample.fit (نموذج ، x = sample.q [: ، لا شيء])
مؤامرة >>> (عينة)
ملاحظة: ينطبق تقريب Gaussian دائما على q2MSD << 1 ، ولكن يمكن استخدام نطاق نقل زخم أوسع للمقارنة النسبية بين العينات. تم تطوير نماذج أكثر تطورا ، والتي تتجاوز التقريب الغاوسي ،37،38،39.
6. تحليل البيانات - عمليات المسح المرنة وغير المرنة للنوافذ الثابتة (E / IFWS)
- على غرار الخطوة 4 ، قم باستيراد مجموعة البيانات ولكن باستخدام الفئة "IN16B_FWS":
>>> من nPDyn.dataParsers استيراد IN16B_FWS
>>> عينة = IN16B_FWS(
... <المسار إلى ملفات البيانات>
... [detGroup = <ملف تجميع عدد صحيح أو كاشف بتنسيق XML>]
... ). عملية ()
>>> عينة = sample.get_q_range (0.3 ، 1.8) - تناسب بيانات المعايرة وبيانات العينة.
- قم بتحليل بيانات E / IFWS باستخدام MSD40 المعمم أو من خلال اعتبارها أطياف QENS الخشنة (مع وجود عدد قليل فقط من نقاط البيانات على محور الطاقة). عندما ينظر إلى E / IFWS على أنه QENS-خشن ، يتم استخدام النماذج المستخدمة في QENS لتناسب مجموعة بيانات E / IFWS بأكملها في وقت واحد (التوافق العالمي لعمليات نقل الطاقة وتحويلات الزخم).
ملاحظة: يتم استخدام نماذج استخدام الحل الأخير ل QENS على بيانات E / IFWS هنا حيث يتم فرض اعتماد نقل الزخم لانتشار مركز الكتلة والديناميات الداخلية للبروتين. - نموذج ديناميكيات البروتين في السوائل باستخدام مكافئ البروتين التالي (5) ("modelProteinJumpDiff" في nPDyn):
 (5)
(5)
حيث R (q، ω) هي دالة الدقة ؛ β عدد قياسي مستقل لكل نقل زخم q ؛ 0 هو عامل البنية غير المتماسكة المرنة (EISF) ؛ حساب لورنتزيان لانتشار مركز الكتلة بعرض معطى بواسطة Eq (6) ؛
حساب لورنتزيان لانتشار مركز الكتلة بعرض معطى بواسطة Eq (6) ؛  هو لورنتزيان يتضمن انتشار مركز الكتلة ومساهمة بعد نموذج انتشار القفز14 الذي يمثل الديناميكيات الداخلية (Eq (7) ؛
هو لورنتزيان يتضمن انتشار مركز الكتلة ومساهمة بعد نموذج انتشار القفز14 الذي يمثل الديناميكيات الداخلية (Eq (7) ؛  كونها الإشارة المجهزة من D2O المعاد قياسها بواسطة جزء حجمها في العينة.
كونها الإشارة المجهزة من D2O المعاد قياسها بواسطة جزء حجمها في العينة.
γ = دقس 2 (6)
Ds هو معامل الانتشار الذاتي. (7)
(7)
Di هو معامل الانتشار الظاهر للديناميكيات الداخلية و τ وقت استرخاء للحركات الانتشارية.
>>> من استيراد nPDyn.models.builtins (
... نموذجPVoigt,
... modelProteinJumpDiff,
... نموذجمعايرةD2O,
... )
# تستخدم النماذج المدمجة متجه عمود للزخم
# نقل قيم q
>>> q = الفاناديوم.q[:, لا يوجد]
# يتم تركيب الفاناديوم باستخدام ملف تعريف زائف Voigt
>>> الفاناديوم.فيت (نموذجPVoigt (ف))
# ل D2O النقي ، نموذج بعرض خط معاير
# لدرجات حرارة مختلفة يتم تضمينها في nPDyn
>>> buffer.fit (modelCalibratedD2O (q ، temp = 363))
# هنا ، يتم استخدام المعادلة 3 للعينات السائلة
# الالتفاف مع وظيفة القرار وإضافة
# تتم خلفية D2O تلقائيا باستخدام # الوسيطات المقدمة
>>> sample.fit (modelProteinJumpDiff (q) ،
... الدقة = الفاناديوم ،
... bkgd = العازلة ،
... volume_fraction_bkgd=0.95
... )
- قم بتحليل بيانات E / IFWS باستخدام MSD40 المعمم أو من خلال اعتبارها أطياف QENS الخشنة (مع وجود عدد قليل فقط من نقاط البيانات على محور الطاقة). عندما ينظر إلى E / IFWS على أنه QENS-خشن ، يتم استخدام النماذج المستخدمة في QENS لتناسب مجموعة بيانات E / IFWS بأكملها في وقت واحد (التوافق العالمي لعمليات نقل الطاقة وتحويلات الزخم).
- ارسم البيانات المجهزة باستخدام:
>>> من قطعة أرض استيراد nPDyn.plot
مؤامرة >>> (عينة)
النتائج
تم إجراء تجميع الليزوزيم في الجسيمات عند 90 درجة مئوية بتركيز بروتين 50 مجم / مل في مخزن مؤقت مقسم (0.1 M NaCl عند pD 10.5). يتم تشغيل تكوين الجسيمات من خلال زيادة درجة الحرارة إلى 90 درجة مئوية ويحدث في غضون 6 ساعات (الشكل التكميلي S8). تم الحصول على البيانات على IN16B ، كما هو موضح في البروتوكول أع?...
Discussion
التحليل الطيفي النيوتروني هو الطريقة الوحيدة التي تسمح بفحص ديناميكيات ps-ns ذات المتوسط الجماعي لعينات البروتين بغض النظر عن حجم البروتين أو تعقيد المحلول عند استخدام deuteration6. على وجه التحديد ، من خلال التحقيق في الانتشار الذاتي لمجموعات البروتين في المحلول ، يمكن تحديد الحجم...
Disclosures
ليس لدى المؤلفين أي تضارب في المصالح للكشف عنه.
Acknowledgements
يعرب المؤلفون عن امتنانهم لميكايلا زامبوني من مركز يوليش لعلوم النيوترون في هاينز ماير لايبنيتز سنتروم ، جارشينج ، ألمانيا ، لجزء من تجارب تشتت النيوترونات التي أجريت على أداة SPHERES. وقد استفاد هذا العمل من أنشطة اتحاد مختبرات Deuteration (DLAB) الذي يموله الاتحاد الأوروبي بموجب العقدين HPRI-2001-50065 و RII3-CT-2003-505925 ، ومن النشاط الممول من مجلس أبحاث العلوم الهندسية والفيزيائية في المملكة المتحدة (EPSRC) داخل معهد Laue Langevin EMBL DLAB بموجب المنح GR / R99393/01 و EP / C015452 / 1. الدعم المقدم من المفوضية الأوروبية في إطار البرنامج الإطاري السابع من خلال الإجراء الرئيسي: تعزيز منطقة البحوث الأوروبية ، يتم الاعتراف بالبنى التحتية البحثية [العقد 226507 (NMI3)]. يشكر كيفن بونوت وكريستيان بيك الوزارة الفيدرالية للتعليم والبحث (BMBF ، رقم المنحة 05K19VTB) على تمويل زمالات ما بعد الدكتوراه.
Materials
| Name | Company | Catalog Number | Comments |
| Aluminum sample holder | Not commercially available. Either the local contact on the instrument can provide them or they can be manufactured based on a technical drawing that can be provided by the local contact. | ||
| Deuterium chloride, 35 wt. % in D2O, ≥99 atom % D | Sigma-Aldrich | 543047 | |
| Deuterium oxide (D, 99.9%) | Eurisotop | DLM-4DR-PK | |
| Dow Corning high-vacuum silicone grease | Sigma-Aldrich | Z273554-1EA | |
| Ethanol 96%, EMSURE Reag. Ph Eur | Sigma-Aldrich | 1.5901 | |
| Glass dessicator | VWR | 75871-660 | |
| Glass dessicator plate, 140 mm | VWR | 89038-068 | |
| Indium wire, 1.0 mm (0.04 in) dia, Puratronic, 99.999% | Alfa Aesar | 00470.G1 | |
| Lysozyme from chicken egg white dialyzed, lyophilized, powder, ~100,000 U/mg | Sigma-Aldrich | 62970 | |
| nPDyn | v3.x | see github.com/kpounot/nPDyn, model functions fot fitting also included in the software | |
| OHAUS AX324 Adventurer balance, internal calibration | Dutscher | 92641 | |
| Phosphorus pentoxide, ReagentPlus, 99% | Sigma-Aldrich | 214701 | |
| Pipette ErgoOne 0.5-10 μL | Starlab | S7100-0510 | |
| Pipette ErgoOne 100-1,000 μL | Starlab | S7100-1000 | |
| Pipette ErgoOne 20-200 μL | Starlab | S7100-2200 | |
| Pipette tip TipOne 1,000 μL | Starlab | S1111-6001 | |
| Pipette tip TipOne 10 μL | Starlab | S1111-3200 | |
| Pipette tip TipOne 200 μL | Starlab | S1111-0206 | |
| Sodium deuteroxide solution, 40 wt. % in D2O, 99.5 atom % D | Sigma-Aldrich | 372072 |
References
- Jacrot, B. Des neutrons pour la science: Histoire de l'Institut Laue-Langevin. Des neutrons pour la science. EDP Sciences. , (2021).
- Mahieu, E., Gabel, F. Biological small-angle neutron scattering: recent results and development. Acta Crystallographica Section D. 74 (8), 715-726 (2018).
- Grimaldo, M., Roosen-Runge, F., Zhang, F., Schreiber, F., Seydel, T. Dynamics of proteins in solution. Quarterly Reviews of Biophysics. 52, 7 (2019).
- Martel, A., et al. Membrane permeation versus amyloidogenicity: A multitechnique study of islet amyloid polypeptide interaction with model membranes. Journal of the American Chemical Society. 139 (1), 137-148 (2017).
- Pounot, K., et al. Tracking internal and global diffusive dynamics during protein aggregation by high-resolution neutron spectroscopy. The Journal of Physical Chemistry Letters. 11 (15), 6299-6304 (2020).
- Grimaldo, M., et al. Protein short-time diffusion in a naturally crowded environment. The Journal of Physical Chemistry Letters. 10 (8), 1709-1715 (2019).
- Jasnin, M., Stadler, A., Tehei, M., Zaccai, G. Specific cellular water dynamics observed in vivo by neutron scattering and NMR. Physical Chemistry Chemical Physics. 12 (35), 10154-10160 (2010).
- Frick, B. The neutron backscattering spectrometer IN16 at ILL-high energy resolution with high intensity and excellent signal-to-noise ratio. Neutron News. 13 (2), 15-22 (2002).
- Frick, B., Mamontov, E., van Eijck, L., Seydel, T. Recent backscattering instrument developments at the ILL and SNS. Zeitschrift für Physikalische Chemie. 224 (1-2), 33-60 (2010).
- Frick, B., Combet, J., van Eijck, L. New possibilities with inelastic fixed window scans and linear motor Doppler drives on high resolution neutron backscattering spectrometers. Nuclear Instruments and Methods in Physics Research Section A: Accelerators, Spectrometers, Detectors and Associated Equipment. 669, 7-13 (2012).
- Appel, M., Frick, B., Magerl, A. A flexible high speed pulse chopper system for an inverted neutron time-of-flight option on backscattering spectrometers. Scientific Reports. 8 (1), 13580 (2018).
- Squires, G. L. . Introduction to the theory of thermal neutron scattering. , (1996).
- Singwi, K. S., Sjölander, A. Diffusive motions in water and cold neutron scattering. Physical Review. 119 (3), 863-871 (1960).
- Sears, V. F. Theory of cold neutron scattering by homonuclear diatomic liquids: i. free rotation. Canadian Journal of Physics. 44 (6), 1279-1297 (1966).
- Sears, V. F. Theory of cold neutron scattering by homonuclear liquid: ii. hindered rotation. Canadian Journal of Physics. 44 (6), 1299-1311 (1966).
- Schirò, G., et al. Translational diffusion of hydration water correlates with functional motions in folded and intrinsically disordered proteins. Nature Communications. 6, 6490 (2015).
- Grimaldo, M., et al. Hierarchical molecular dynamics of bovine serum albumin in concentrated aqueous solution below and above thermal denaturation. Physical Chemistry Chemical Physics. 17 (6), 4645-4655 (2015).
- Eanes, E. D., Glenner, G. G. X-ray diffraction studies on amyloid filaments. Journal of Histochemistry & Cytochemistry. 16 (11), 673-677 (1968).
- Bonar, L., Cohen, A. S., Skinner, M. M. Characterization of the Amyloid Fibril as a Cross-β Protein. Proceedings of the Society for Experimental Biology and Medicine. 131 (4), 1373-1375 (1969).
- Chiti, F., Dobson, C. M. Protein Misfolding, Amyloid Formation, and Human Disease: A Summary of Progress Over the Last Decade. Annual Review of Biochemistry. 86 (1), 27-68 (2017).
- Knowles, T. P. J., Vendruscolo, M., Dobson, C. M. The amyloid state and its association with protein misfolding diseases. Nature Reviews Molecular Cell Biology. 15 (6), 384-396 (2014).
- Maji, S. K., et al. Functional amyloids as natural storage of peptide hormones in pituitary secretory granules. Science. 325 (5938), 328-332 (2009).
- Li, J., et al. The RIP1/RIP3 necrosome forms a functional amyloid signaling complex required for programmed necrosis. Cell. 150 (2), 339-350 (2012).
- Knowles, T. P. J., Mezzenga, R. Amyloid fibrils as building blocks for natural and artificial functional materials. Advanced Materials. 28 (31), 6546-6561 (2016).
- Stephens, A. D., Kaminski Schierle, G. S. The role of water in amyloid aggregation kinetics. Current Opinion in Structural Biology. 58, 115-123 (2019).
- Adamcik, J., Mezzenga, R. Amyloid polymorphism in the protein folding and aggregation energy landscape. Angewandte Chemie International Edition. 57 (28), 8370-8382 (2018).
- Liu, Z., et al. Entropic contribution to enhanced thermal stability in the thermostable P450 CYP119. Proceedings of the National Academy of Sciences of the United States of America. 115 (43), 10049-10058 (2018).
- Coquelle, N., et al. Chromophore twisting in the excited state of a photoswitchable fluorescent protein captured by time-resolved serial femtosecond crystallography. Nature Chemistry. 10 (1), 31-37 (2018).
- Henzler-Wildman, K. A., et al. A hierarchy of timescales in protein dynamics is linked to enzyme catalysis. Nature. 450 (7171), 913-916 (2007).
- Fichou, Y., et al. Hydration water mobility is enhanced around tau amyloid fibers. Proceedings of the National Academy of Sciences of the United States of America. 112 (20), 6365-6370 (2015).
- Burns, J., Pennock, C. A., Stoward, P. J. The specificity of the staining of amyloid deposits with thioflavine T. The Journal of Pathology and Bacteriology. 94 (2), 337-344 (1967).
- Iqbal, K., Liu, F., Gong, C. -. X., Grundke-Iqbal, I. Tau in Alzheimer disease and related tauopathies. Current Alzheimer Research. 7 (8), 656-664 (2010).
- Krȩżel, A., Bal, W. A formula for correlating pKa values determined in D2O and H2O. Journal of Inorganic Biochemistry. 98 (1), 161-166 (2004).
- Dolman, M., Halling, P. J., Moore, B. D., Waldron, S. How dry are anhydrous enzymes? Measurement of residual and buried 18O-labeled water molecules using mass spectrometry. Biopolymers. 41 (3), 313-321 (1997).
- Pounot, K. kpounotnPDyn: v3.0.0. Zenodo. , (2021).
- Yi, Z., Miao, Y., Baudry, J., Jain, N., Smith, J. C. Derivation of mean-square displacements for protein dynamics from elastic incoherent neutron scattering. Journal of Physical Chemistry B. 116 (16), 5028-5036 (2012).
- Peters, J., Kneller, G. R. Motional heterogeneity in human acetylcholinesterase revealed by a non-Gaussian model for elastic incoherent neutron scattering. The Journal of Chemical Physics. 139 (16), 165102 (2013).
- Zeller, D., Telling, M. T. F., Zamponi, M., García Sakai, V., Peters, J. Analysis of elastic incoherent neutron scattering data beyond the Gaussian approximation. The Journal of Chemical Physics. 149 (23), 234908 (2018).
- Roosen-Runge, F., Seydel, T. A generalized mean-squared displacement from inelastic fixed window scans of incoherent neutron scattering as a model-free indicator of anomalous diffusion confinement. EPJ Web of Conferences. 83, 02015 (2015).
- Ortega, A., Amorós, D., García de la Torre, J. Prediction of hydrodynamic and other solution properties of rigid proteins from atomic- and residue-level models. Biophysical Journal. 101 (4), 892-898 (2011).
- Hennig, M., Frick, B., Seydel, T. IUCr Optimum velocity of a phase-space transformer for cold-neutron backscattering spectroscopy. Journal of Applied Crystallography. 44 (3), 467-472 (2011).
- Paalman, H. H., Pings, C. J. Numerical evaluation of X-ray absorption factors for cylindrical samples and annular sample cells. Journal of Applied Physics. 33 (8), 2635-2639 (1962).
- Ow, S. -. Y., Dunstan, D. E. The effect of concentration, temperature and stirring on hen egg white lysozyme amyloid formation. Soft Matter. 9 (40), 9692-9701 (2013).
- Tominaga, T., Sahara, M., Kawakita, Y., Nakagawa, H., Yamada, T. Evaluation of sample cell materials for aqueous solutions used in quasi-elastic neutron scattering measurements. Journal of Applied Crystallography. 54 (6), 1631-1640 (2021).
- Beck, C., et al. Following protein dynamics in real time during crystallization. Crystal Growth & Design. 19 (12), 7036-7045 (2019).
- Smith, A. A., Testori, E., Cadalbert, R., Meier, B. H., Ernst, M. Characterization of fibril dynamics on three timescales by solid-state NMR. Journal of Biomolecular NMR. 65 (3-4), 171-191 (2016).
- Wang, T., Jo, H., DeGrado, W. F., Hong, M. Water distribution, dynamics, and interactions with Alzheimer's β-amyloid fibrils investigated by solid-state NMR. Journal of the American Chemical Society. 139 (17), 6242-6252 (2017).
- Rezaei-Ghaleh, N., Giller, K., Becker, S., Zweckstetter, M. Effect of zinc dinding on β-amyloid structure and dynamics: Implications for Aβ aggregation. Biophysical Journal. 101 (5), 1202-1211 (2011).
- Vugmeyster, L., et al. Fast motions of key methyl groups in amyloid-β fibrils. Biophysical Journal. 111 (10), 2135-2148 (2016).
- Yang, X., Wang, B., Hoop, C. L., Williams, J. K., Baum, J. NMR unveils an N-terminal interaction interface on acetylated-α-synuclein monomers for recruitment to fibrils. Proceedings of the National Academy of Sciences of the United States of America. 118 (18), (2021).
- Tuttle, M. D., et al. Solid-state NMR structure of a pathogenic fibril of full-length human α-synuclein. Nature Structural & Molecular Biology. 23 (5), 409-415 (2016).
- Karamanos, T. K., Kalverda, A. P., Thompson, G. S., Radford, S. E. Mechanisms of amyloid formation revealed by solution NMR. Progress in Nuclear Magnetic Resonance Spectroscopy. 88-89, 86-104 (2015).
- Lai, Y. -. C., Kuo, Y. -. H., Chiang, Y. -. W. Identifying protein conformational dynamics using spin-label ESR. Chemistry - An Asian Journal. 14 (22), 3981-3991 (2019).
- Franck, J. M., Han, S. Overhauser dynamic nuclear polarization for the study of hydration dynamics, explained. Methods in Enzymology. 615, 131-175 (2019).
- Pavlova, A., et al. Protein structural and surface water rearrangement constitute major events in the earliest aggregation stages of tau. Proceedings of the National Academy of Sciences of the United States of America. 113 (2), 127-136 (2016).
- Lin, Y., et al. Liquid-liquid phase separation of tau driven by hydrophobic interaction facilitates fibrillization of tau. bioRxiv. , (2020).
- Decatur, S. M. Elucidation of residue-level structure and dynamics of polypeptides via isotope-edited infrared spectroscopy. Accounts of Chemical Research. 39 (3), 169-175 (2006).
- Chatani, E., Tsuchisaka, Y., Masuda, Y., Water Tsenkova, R. molecular system dynamics associated with amyloidogenic nucleation as revealed by real time near infrared spectroscopy and aquaphotomics. PLoS One. 9 (7), 101997 (2014).
- Goret, G., Aoun, B., Pellegrini, E. MDANSE: An interactive analysis environment for molecular dynamics simulations. Journal of Chemical Information and Modeling. 57 (1), 1-5 (2017).
- Fujiwara, S., et al. Internal dynamics of a protein that forms the amyloid fibrils observed by neutron scattering. Journal of the Physical Society of Japan. 82, (2013).
- Schiró, G., et al. Neutron scattering reveals enhanced protein dynamics in concanavalin a amyloid fibrils. Journal of Physical Chemistry Letters. 3 (8), 992-996 (2012).
- Pounot, K., et al. Zinc determines dynamical properties and aggregation kinetics of human insulin. Biophysical Journal. 120 (5), 886-898 (2021).
- Fujiwara, S., et al. Dynamic properties of human α-synuclein related to propensity to amyloid fibril formation. Journal of Molecular Biology. 431 (17), 3229-3245 (2019).
- Sanz, A., et al. High-pressure cell for simultaneous dielectric and neutron spectroscopy. Review of Scientific Instruments. 89 (2), 023904 (2018).
- Adams, M. A., et al. Simultaneous neutron scattering and Raman scattering. Applied Spectroscopy. 63 (7), 727-732 (2009).
Reprints and Permissions
Request permission to reuse the text or figures of this JoVE article
Request PermissionExplore More Articles
This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. All rights reserved