
Method Article
黄曲霉毒素在花生RNAi介导的控制:方法分析霉菌毒素的产生和转基因表达的花生/
摘要
我们证明了包含RNA干扰信号沉默中黄曲霉毒素合成基因真菌黄曲霉黄曲霉毒素的转基因表达的花生种子的分析方法。真菌毒素的植物中RNAi介导的控制以前没有被报道。
摘要
联合国粮食和农业组织估计,粮食作物在世界上25%的污染黄曲霉毒素。表示亿吨食物被破坏或每年改行非人类消费。黄曲霉毒素是致癌物质强大通常由真菌黄曲霉和寄生 A.谷物,坚果,块根作物等农产品积累。五黄曲霉毒素合成基因在植物花生RNA干扰(RNAi)沉默使用以下接种以 A来控制黄曲霉毒素积累菌 。以前,没有方法存在来分析的RNAi在个别花生转基因事件的有效性,因为这些通常会产生黄曲霉毒素有利的条件下几个种子,和传统的大视场实验方法进行了不是一个选项。在该领域,寻找自然污染的种子的概率是经常1/100至1/1,000。另外,黄曲霉毒素污染是不均匀分布的。我们的方法使用每个转基因事件的种子数,以处理用于实时PCR(RT-PCR)小块或小RNA测序,和黄曲霉毒素的积累通过超高效液相色谱(UPLC)分析。的RNAi表达花生线288-72和288-74,显示出高达100%的减少(P≤0.01)中黄曲霉毒素B 1和B 2相比,累加到14000纳克控制。克黄曲霉毒素1时-1接种是否产A.菌 。作为参考,最大总黄曲霉毒素的允许供人食用的在美国为20纳克。克-1。本协议描述的黄曲霉毒素的转基因花生种子和其评价方法RNAi介导的控制中的应用。我们相信,它在花生等作物的育种中的应用将带来快速的进步在科学这一重要领域,医学和人体营养,而且会显著促进国际努力控制黄曲霉毒素,以及潜在的其他真菌毒素的主要粮食作物。
引言
约4.5十亿人长期接触黄曲霉毒素1,在大自然2已知最强大的致癌物质。这些真菌毒素污染的粮食作物的25%的世界3,包括玉米,木薯,大米,坚果,谷类和香料。4。黄曲霉毒素导致发育迟缓的儿童5,损害免疫系统 6,存在于肝细胞,癌58%的人活检7,8,并在对黄曲霉毒素中毒9,10周期性爆发杀死数百人。黄曲霉毒素是通常由黄曲霉菌 和 A.生产的聚酮化合物来源的霉菌毒素寄生 ;黄曲霉毒素B 1和B 2是由 A.产生黄曲霉 ,而A.寄生还生产G1和G2 2。这些化合物和一个色谱图,显示它们的分离通过UPLC的化学结构示于图1。

图1. 黄曲霉毒素和RNAi插入 顶部 :四种最常见的聚酮化合物衍生黄曲霉毒素的化学结构(左)和例子色谱图(右):B 1,B 2,G 1和G 2, 寄生曲霉 , 所生产的。 黄曲霉产生B 1和B 2底部 :示意图中所述RNAi基因片段的构建p5XCAPD用于花生变换,下箭头号在黄曲霉基因组基因片段的登录号。 PIV2:马铃薯内含子;沸点:碱基对; RT_5X_1和RT_5X_2:实时荧光定量PCR引物网站请点击此处查看该图的放大版本。
{kind=link}
在出口因花生黄曲霉毒素的经济损失就超过$ 4.5亿美元如果基于4计算纳克例 -1黄曲霉毒素的限制允许用于人类消费在欧盟11。黄曲霉毒素已经知道60年12;然而,尽管许多农业实践的开发是为了减轻其影响,包括应用程序的其他真菌菌株13,14,控制没有一致的方式存在,抗性植物品种不具备的。测试植物种质的抗黄曲霉毒素是特别困难的,因为即使在病原体入侵有利的条件,毒素积累是不可预测的,不符合正态分布。因此,实验通常需要大面积种植,数百种子和100-1,700克多个样本以减小数据15,16的可变性。
RNA干扰是发现于1998年17;和"沉默"的好处目前正在探讨在许多新的应用,例如,在针对转移性乳腺癌18,肝癌19,粒细胞性白血病20,和在植物保护,防止昆虫21和线虫22人类疗法。在植物中,RNA干扰信号可行驶细胞与细胞,小干扰RNA(siRNA)和高分子量的RNA负责全身转录后基因沉默23,24,甚至内部真菌病原体是与植物宿主25紧密接触。 RNAi对真菌病原体的基因植物介导的沉默的有效性已经在几个植物pathosystems了说明,对于这些,在植物(叶)的地上部分症状目视检查允许病量化, 即 ,卵菌Bremia莴苣26 , 锈病小麦 27和枯萎的香蕉28。更加困难的是评估RNAi的有效性来控制霉菌毒素的植物,特别是黄曲霉毒素在花生的叶子表明没有感染症状,侵入(种子)的器官是下几英寸的土壤,感染的发生是不可预知的,只有化学分析可确定的黄曲霉毒素的存在。此外,在花生每个转基因事件通常会产生一些种子(4-6每株植物);因此,传统的测试无黄曲霉毒素积累特征在大视场地块,持续整个收获季节,用上百种的是不可行的。一种方法,在此描述不到一周来分析,RNA干扰花生种子转基因的存在和用于无黄曲霉毒素积累性状,只用几个种子。
研究方案
1.分子构造和花生转型
- 结合五A. DNA片段黄曲霉基因,AFL2G_07223(aflS或aflJ),AFL2G_07224(aflR),AFL2G_07228(AFLC / PKSA / pksL1),AFL2G_07731(Pes1低 )和AFL2G_05027(黄曲霉毒素外排泵,aflep)。为此,请使用以下的引物和ultramers:DIR-1,短期方向1-R,DIR-2反转,短期方向2-R,DirAll-NCO-RV和DirAll-BamEco-FW, 见表1。
- 使DIR-1双链由5个PCR循环(25微升反应; 95℃2分钟,然后5个循环的94℃45秒,55℃30秒,68℃15秒)根据制造商的使用DNA聚合酶指令和底漆短Dir1中-R留下3'突出CCCGT。重复这些步骤,使用引物的短方向2-R留下3'突出ACGGG互补DIR-1,使DIR-2反转双链。
- 结扎两个199 bp的fragmeNTS用T4 DNA连接酶按制造商的说明。 PCR扩增得到的393碱基对片段作为在1.1.1使用引物DirAll-CACC-FW和DirAll-NCO-RV(表1)表示,并且使用标准技术进入pENTR1A使质粒P2 + 4ENTR克隆的产物。
- 重组P2 + 4ENTR成pCAPD 29(NCBI登录:KC176455.1)根据制造商的说明进行质粒p5XCAPD使用LR克隆酶II酶混合物,并用标准技术后跟部分测序它转化大肠杆菌DH5α。注意:完整的RNAi插入物示于表1。
- 变换农杆菌菌株 C58C1 30用质粒p5XCAPD如先前报道30,并使用所产生的细菌如下转化花生植物:
- 生长在30℃的农杆菌窝藏p5XCAPD,要用于此,将50ml的LB肉汤补充有500微克毫升-1链霉素,25微克毫升-1庆大霉素,10微克毫升-1卡那霉素,并摇动培养在250转,直到达到1 OD 260。
- 收获农杆菌细胞通过离心(6000×g离心)10分钟,重新悬浮在50ml AB基本培养基31与100微米乙酰丁香酮的1小时,并发生在细菌悬液10-14天龄幼苗Exp27-1516,转轮的外植体型花生育种线。吸干的3MM吸墨纸30分钟后,外植体,并将其置于枪战诱导培养基(SIM)MS盐32,3%蔗糖,20微米苄氨基嘌呤(BAP),10微米噻苯隆(TDZ),pH值5.8,0.3 %吉兰糖胶],而不在黑暗中三天抗生素。
- 做组织选择和再生为33之前的报道。移动的组织到SIM(500μM头孢噻肟和100μM的卡那霉素)进行枝条形成,以每两周传输2个月。然后将扩大对芽伸长培养基(SEM)拍摄[5微米BAP,1微米赤霉素(GA 3),双周数月。
- 放置单个枝条,在大小为2厘米,在根诱导培养基(RIM)[1/2 MS,1.5%蔗糖,5μM的α - 萘乙酸(NAA),2.5μM吲哚丁酸(IBA)],再适应的苗,转移到温室。
2.确定花生植物窝藏RNAi技术沉默中黄曲霉毒素合成基因
- 在一个机器人工作站根据制造商的说明,从幼叶花生植物都受转化的过程中提取的DNA200μl的洗脱使用植物小试剂盒(如前面所述)与RNAi构建p5XCAPD(图1),其具有作为主链质粒pCAPD 29为基因沉默。
- 筛选通过单管巢式PCR(STN-PCR)的DNA样品所描述PREVIously 34以检测可选择标记的NPTⅡ和所述RNAi从p5XCAPD插入。从切口(3-4节点)PCR阳性的植物无性繁殖产生足够的种子进行测试的第一代。
- 使用的DNA的4个2倍稀释液(50-100纳克微升-1稀释前)中的所有的STN-PCR反应。用于NPTII,使用外部引物PCAPD 5714F:5'-AGGCTATTCGGCTATGACTG-3'和PCAPD 6446R:5'-CGTCAAGAAGGCGATAGAAG-3',和内部引物PCAPD 5730F:5'-ACTGGGCACAACAGACAATC-3'和PCAPD 6249R:5'-ATATTCGGCAAGCAGGCATC- 3'。
- 用于检测的RNAi插入在STN-PCR反应的使用外部引物35S-PDSFw:5'-CCTAACAGAACTCGCCGTAA-3'和DirAll-NCO-RV:5'-ATGCCATGGGGTTATTGGGTGCAGAATGG-3',和内部引物Probe_5027_Fw:5'-gtatttgtgaccatgtttctg -3- '和Probe_7228_Rv:5'-GGACGGATAGTAAACTGCGG-3'。
- 从CON的STN-PCR阳性植株和收获的花生荚控制植物生长在相同的条件。关键的一步:确保控制植物生长在相同的条件和相同的季节作为RNAi的植物。水高炉用压力垫圈荚果以低强度或用手刮掉通过将舱体上的到期板和独立荚基(黄色,橙色,棕色和黑色)35(图除去外果皮,确定中果皮的颜色2)。

图2.准备花生荚进行分析。左:发现在收获花生是一种无限生长的植物不同的花生大小;中心:将花生在金属篮水压除去外果皮; 右:成熟群体通过在花生纹板(黄色,橙色,棕色和黑色)中果皮颜色。98fig2large.jpg"目标="_空白">点击此处查看该图的放大版本。
3.实验装置
- 拆下外壳,工艺黄色和棕色种子分开。计算种子在根据表2中的实验使用的数量。另外,最少三个种块(1种子一块=半子叶)每花生线路和取样时间,需要进行统计分析,并降低的标准误差。
| 名称 | 序列 | ||
| DIR-1 | 5'- GCCAGCTCAAAAGTGCGATGCACCAAGGAGAAACCGGCCTGTGCT CGGTGTATCGAACGTGGTCTTGCCTGTCAATACATGGTCGTATTTG TGACCATGTTTCTGGTGGCATTGGACCGTCTTGTCATCTCTACAGCC ATTCCCCAGATCACGGACGAATCCCCTGCATCTACGCGCACGCATC ACTTGGGGTACCCGT-3' | ||
| 短期方向1-R | 5'-PHOS-TACCCCAAGTGATGCGTGCGCG-3' | ||
| DIR-2反转 | 5'- GGTTATTGGGTGCAGAATGGTAAACCACCCAACAGTACGCGAAATG TCAATTCCAGAGTCCCAAACCTCCCTACCGTGGCCTGGACGGATAG TAAACTGCGGAGCTTGGGAACAAAATCCGCTGTCTGATCGCCGAAG AGAAAGAGTTGCCTTGATTGAGCCGCATCGAGGACAGGTTGTGTTG CTGTTGATAGACGGG-3' | ||
| 短期方向2-R | 5'-PHOS-CTATCAACAGCAACACAACC-3' | ||
| DirAll-CACC-FW | 5'-CACCGCCAGCTCAAAAGTGCGATGC-3' | ||
| DirAll-NCO-RV | 5'-ATGCCATGGGGTTATTGGGTGCAGAATGG-3' | ||
| DirAll-BamEco-FW | 5'-ATGGGATCCGAATTCGCCAGCTCAAAAGTGCGATGC-3' | ||
| 完整的RNA干扰插入 | 5'- GCCAGCTCAAAAGTGCGATGCACCAAGGAGAAACCGGCCTGTGCT CGGTGTATCGAACGTGGTCTTGCCTGTCAATACATGGTC / GTATTTG TGACCATGTTTCTGGTGGCATTGGACCGTCTTGTCATCTCTACAGCC ATTCCCCAGATCACGGACGAAT / CCCCTGCATCTACGCGCACGCAT CACTTGGGGTACCCGTCTATCAACAGCAACACAACCTGTCCTCGAT GCG / GCTCAATCAAGGCAACTCTTTCTCTTCGGCGATCAGACAGCG GATTTTGTTCCCAAGCTCCGCAGTTTACTATCCGTCCA / GGCCACGG TAGGGAGGTTTGGGACTCTGGAATTGACATTTCGCGTACTGTTGGG TGGTTTACCATTCTGCACCCAATAACC-3' | ||
表 1.寡核苷酸和ultramers用于构建RNAi构建p5XCAPD磷酸:磷酸5'端; "/"分隔用五个基因片段;完整的RNA干扰插入:序列作为2反向重复序列,形成p5XCAPD。
- 放置整个花生种子在一个单一的层,使得它们覆盖的无菌烧杯的底部。添加75%乙醇/水(v / v)溶液以覆盖种子,然后添加相同的溶液的等体积。孵育在室温下进行30秒,然后用无菌的去离子水(SDW)。
- 添加2%次氯酸盐到含有乙醇处理过的种子如下烧杯:补充足够的2%次氯酸盐溶液以覆盖的种子,再加入相同的溶液等体积并孵育5分钟。关键的一步:与卷的SDW相当于彻底冲洗三次,5倍的次氯酸盐用量达到( 图3)。

图3实验装置。上图:花生种子前和次氯酸盐后表面杀菌,去除种皮(种皮)的; 中间:去除胚胎和胚胎的密切观,再切子叶一半; 下图:半子叶无菌蒸馏水,印迹在无菌吸水造纸,水琼脂,置半子叶(切方向上)琼脂表面凹痕。 请点击此处查看该图的放大版本。
{kind=link}
- 允许在表面消毒的种子,以吸收浸入SDW 2小时。将种子上的无菌培养皿,用钳子除去种皮,分离子叶,并用解剖刀除去胚胎。注意:胚胎可以丢弃或用于再生的新植物。
- 切割用手术刀在各一半子叶。为了避免脱水,保持切种块在无菌水中,直到所有的种子处理。
- 已经准备含有无菌水琼脂培养皿(1.5%琼脂/水;重量/体积),一个用于每个3种块。使在琼脂小凹痕使用镊子,简要地吸干的种块在无菌纸巾的过量的水,然后将种块(切割面向上)上的水/琼脂平板上。
- 从是否产黄曲霉 NRRL 3357在察氏琼脂培养基一个新鲜的培养,在25℃生长10天,制备的每微升SDW 50000孢子,用血球计数的悬浮液。
- 放置2微升孢子悬浮液的每一个半子叶片避免在侧面径流的切割表面上,以确保得到孢子暴露于种子组织该港口的RNAi(图4)。

图4.接种和培养黄曲霉毒素的分析。上图:半子叶,接种孢子悬浮液,和黄曲霉经过24小时培养上半子叶菌丝生长下图:左起:48小时的潜伏期在1.5%琼脂;中心:孵化72小时1.5%的琼脂;右:不正确的实验装置的例子中,培养72小时,在0.5%的琼脂请点击此处查看该图的放大版本。
{kind=link}
- 孵育含有接种和未接种的半子叶培养皿在30℃下在黑暗中直到采样。
4.抽样黄曲霉毒素分析和基因表达
- 收集样本一式三份在培养24,48和72小时(可选96小时),两个用于RT-PCR和黄曲霉毒素分析,每个复制为一种片(半子叶)。抽取样品来自不同的板块上的每个采样日期随机。用纸巾,将种块在小瓶或试管前,轻轻地取出琼脂和过量的真菌孢子。
- 黄曲霉毒素分析,将每个重复(一张)在4毫升玻璃螺旋盖小瓶中,并储存在-80℃。注意:给定的黄曲霉毒素的化学稳定性,将样品可以存储在此条件下数月(在当前的实验一般1-2个月)。
- 用于RT-PCR的地方每个重复(一张)中已制备的2 ml的研磨含有两个不锈钢珠(2.5毫米直径)和三个锆珠(2mm直径)管中。
- 立即冻结(优选在液氮中)所有样品并保持在-80℃,直到处理。
个别半的C 5黄曲霉毒素分析otyledon件
- 使用A.黄曲霉接种样本这一分析。携带样品到环境温度约30分钟,加入四卷(通常为2-3毫升;重量/体积)的甲醇(保持便于以后计算记录),合上盖子,孵化澳/ N(〜16小时)的暗无需搅拌。
- 放置一个玻璃料成匹配1.5 ml的丙烯微柱,加入200毫克的Al基本2 O 3和用如先前36描述的另一种玻璃料盖住它;然后,将柱足够接近下一个超高效液相Chomatographer(UPLC)自动进样瓶中,以避免洗脱物可能蒸发。
- 在一次性玻璃试管(不使用塑料),放置0.5毫升步骤4.1中得到的甲醇提取物,加0.5毫升乙腈,用移液管混合并应用0.5毫升混合物进入在步骤4.2中制备的柱。允许洗脱自动进样器小瓶通过重力(不施加压力)。洗脱通常需要2-4分钟,接近日Ë小瓶立即用UPLC兼容的帽隔。保持小瓶在环境温度下,并分析它们放在UPLC在同一天。
- 对于黄曲霉毒素的分离,含有样品的洗脱液和自动进样器小瓶黄曲霉毒素标准的UPLC仪器配备有一个匹配的UPLC四元溶剂管理器,UPLC样品管理器,超高效液相色谱荧光检测器(B 1和B 2使用黄曲霉时)的地方自动进样瓶,和Ç17 2.1毫米×50毫米,1.7微米柱。
- 在0.30 ml的分钟的流速使用的水/甲醇/ CH 3 CN(64:23:13,体积/体积/体积)混合物组成的等度流动相-1。获得的色谱确保一个稳定的基线分离黄曲霉毒素浓度的精确计算,根据制造商的说明37。
- 通过获得它们的质量光谱数据,并将其与公布的数据38进行比较确认黄曲霉毒素的身份。使用I上阱质谱仪配备有电喷雾接口,并根据制造商的说明相应的软件。
- 参照通过注射不同量的相应的黄曲霉毒素B 1,B 2商业标准的获得的校准曲线确定的黄曲霉毒素的浓度中 ,G 1和G 2所建议的UPLC制造商,由软件37来确定。
- 将种块已经用甲醇提取,在个别玻璃小瓶的O / N(〜16小时),冻干,以确定其干重。然后,计算NG。种子一片干重量(g)-1黄曲霉毒素浓度。
- 对数据进行分析,转换黄曲霉毒素的结果来登录(纳克。克-1 1),接着Tukey检验进行平均值的比较。
6.基因表达,样品的RT-PCR法处理中
- 取2 ml的研磨管中含荷兰国际集团的样品从-80℃冷冻,并立即(不解冻)在3100转进行40秒研磨它们在珠磨机均化,然后使用Trizol根据制造商的说明进行到RNA提取。
- 使用1微克RNA和寡聚dT和随机六聚体的相等量的每种样品制备cDNA,使1:8稀释的cDNA,并使用每在RT-PCR反应2微升(如前所述39)。
- 用于检测的RNAi插入件使用的引物表达的:RT_5X_1_105F:5'GGTGGCATTGGACCGTCTTG-3',RT_5X_1_232R:5'-CGCATCGAGGACAGGTTGTG-3';和RT_5X_2_95F:5'-CCATGTTTCTGGTGGCATTG-3',RT_5X_2_229R:5'-ATCGAGGACAGGTTGTGTTG-3'。
- 用于检测选择性标记的NPTⅡ的表达,使用引物:RT_NPTII_1_6871F:5'-CTCGCTCGATGCGATGTTTC-3',RT_NPTII_1_7004R:5'-GCAGGATCTCCTGTCATCTC-3'。使用看家基因肌动蛋白标准化,和引物:肌动蛋白-FW:CACATGCCATCCTTCGATTG;肌动蛋白-RV:CCAAGGCAACATATGCAAGCT 40。
- 分析由三角洲三角洲(C T)方法41标准化的肌动蛋白表达的结果。代表结果通过控制倍。
结果
质粒p5XCAPD被制成pCAPD 29的衍生物,并用它来 转化花生植物;这个载体携带的五个小片段反向重复A.黄曲霉毒素合成的基因,每70-80基点, 黄曲霉由一个内含子(图1)隔开。 AFL2G_07224(aflR),AFL2G_07223(aflS或aflJ),AFL2G_05027(黄曲霉毒素外排泵,aflep),AFL2G_07228(AFLC / PKSA / pksL1),和AFL2G_07731(Pes1低 )的片段用于构建物, 在图1上对应编号,以甲。菌基因组注释的Broad研究所,马萨诸塞州剑桥市,和文学42。共有99花生线经过改造过程后再生,50 PCR阳性的NPTII由STN-PCR方法检测,以及33行PCR阳性而产生的种子。只有七个PCR阳性细胞系无性繁殖和由presen测试吨方法黄曲霉毒素积累,所有七个显示60%至100%以下的黄曲霉毒素积累比对照。在这里,我们展示了其中的两个七行的结果。作为单独的转基因事件通常会产生一些种子,一种方法是开发利用,同时仍然能够做参数统计分析的种子的最小数目。样品制备和实验装置的流程图示于图 5和表 1,虽然第一代转基因种子的通常是半合子,因此预 计在细胞与细胞间和小干扰RNA的全身运动(RNA干扰)产生的通过RNA干扰应赋予黄曲霉素的合成沉默整个工厂。

该方法的 图5. 概要流程图来分析的RNAi沉默的有效性 花生种子 黄曲霉 黄曲霉毒素合成的基因 。工作流程的处理花生样品进行基因表达或黄曲霉毒素分析时图形表示。 请点击此处查看该图的放大版本。
{kind=link}
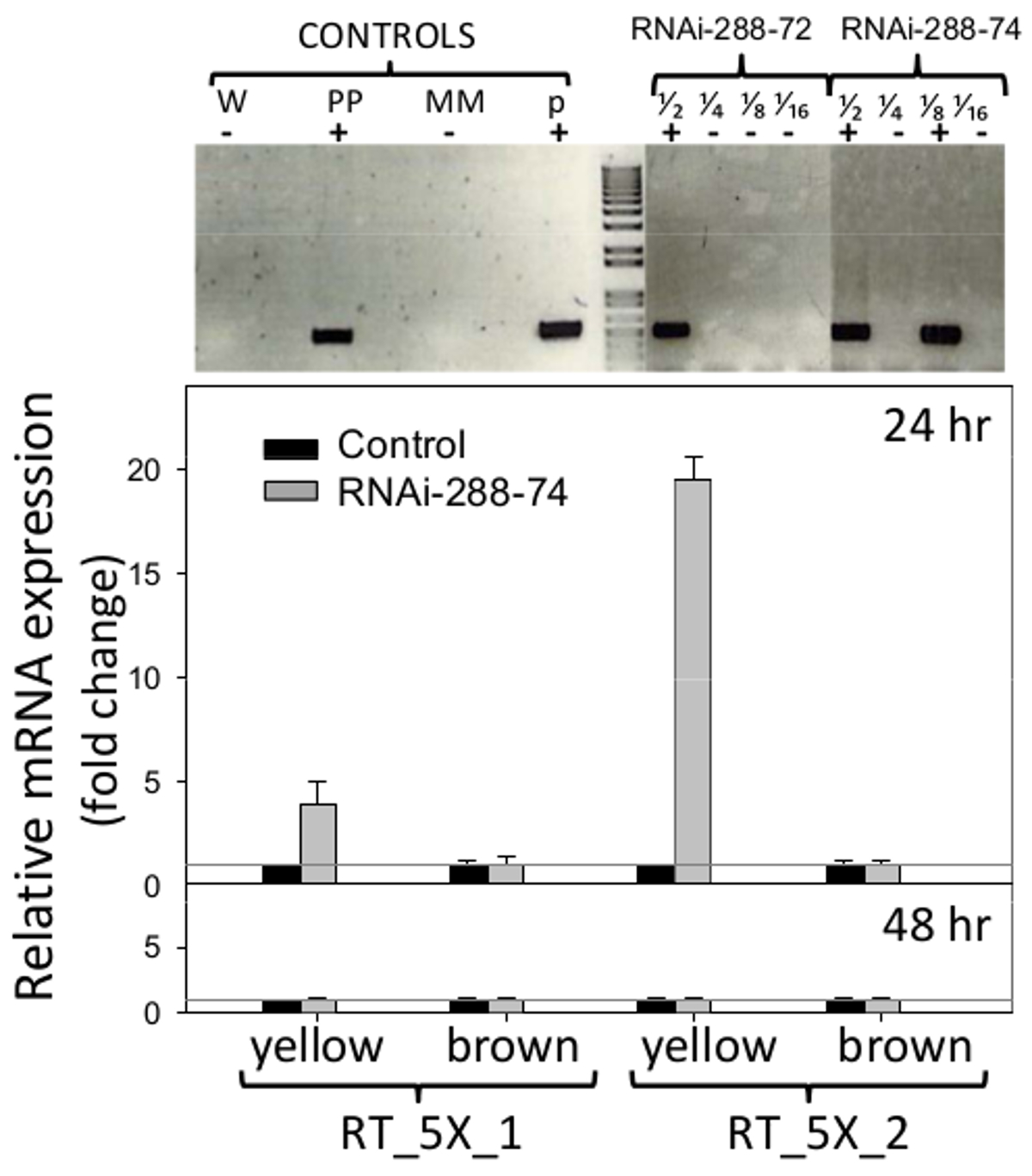
RNAi的花生线288-72和288-74显示,NPTII选择标记的存在时通过的STN-PCR检测,凝胶部分被示于图6(顶),原始图片可从根据要求作者。因为它编码两个基因CMR(氯霉素抗性)和对植物的未知影响CCDB(毒素)的反向重复质粒pCAPD不作为转型的控制。 PCR阴性花生线路288-9,而另一些通过再生过程去,并种植在相同的条件下的RNAi线,用作阴性CON组控制。

图 6. 检测转基因和RNA干扰的实时表情插入 顶部:单管,巢式PCR检测转基因花生线的RNAi 288-72和RNAi-288-74,阳性植物。控制:W(水),PP(阳性植物),MM(主混合物)中,p(质粒p5XCAPD);级分1/2 1/4 1/8 1/16 个代表的DNA 的底部 2倍稀释:实时PCR检测所述RNAi表达的插入(引物组:RT_5X_1,RT_5X_2,如在图1中,对未成熟(黄色)和成熟的转基因品系(棕色)子叶在24和48小时温育;灰线:(C T)= 1。直方图表示三次生物样品的平均值和标准误差线(T)的有三个技术复制。 RNA干扰插入的相对定量进行标准化相对于管家基因肌动蛋白作为内部控制,并在5.2.2和5.3中所述计算出的转基因的比较倍的表达。 请点击此处查看该图的放大版本。
{kind=link}
为了测试植物宿主RNAi介导的潜在黄曲霉毒素积累的控制的有效性,黄曲霉NRRL 3357的新鲜收获的分生孢子施加在半子叶从中胚胎和种皮被除去切断面( 图3,图4)。 黄曲霉 NRRL 3357,对于其基因组已被测序,并且是基础设计p5XCAPD,被亲切地用喇叭博士USDA-ARS-NPRL提供。所得真菌侵袭半子叶后24,48和72小时,在30℃下是显示中的n 图4。的接种半子叶收集样品在24,48,72,96小时温育,并且通过LC-MS分析为主要4黄曲霉毒素B 1,B 2,G 1和G 2使用UPLC和确认;结果示于图6中。黄曲霉毒素浓度使用公开的方法进行了修改36确定。经过96小时培养,子叶启动,因为真菌感染的瓦解。在未成熟的子叶和在成熟的人最采样日期所有采样日期的RNAi线288-72展出黄曲霉毒素比对照的显著较低水平。 RNA干扰线288-74表现出黄曲霉毒素显著较低水平顶多采样日期。的Tukey检验显着性水平标以在图7的图形星号。
g7.jpg"/>
图 7. 中的黄曲霉毒素 孵育(24,48,72和96小时)与 黄曲霉 后 B 1和B 2 中的一半花生子叶控制:288-9非转基因系种子; RNA干扰:从RNA干扰-288-72和RNAi-288-74,转基因的RNAi p5XCAPD种子沉默5黄曲霉毒素合成的基因。 ( 一)黄曲霉毒素1的成熟种子(褐色); ( 二)黄曲霉毒素B 2的成熟种子;在未成熟种子(黄色)和未成熟种子(D)中黄曲霉毒素的 B 2(C)的黄曲霉毒素乙1。具有重复生物样品的相应的标准误差线(T)的平均值表示。据统计显著差异Tukey检验*:P≤0.05,**:P≤0.01,***:P≤0.001。S://www.jove.com/files/ftp_upload/53398/53398fig7large.jpg"目标="_空白">点击此处查看该图的放大版本。
整体的RNAi-288-72表明整个实验(24至96小时培养),在黄曲霉毒素B 2 94%-100%减少,以及90%-100%的减少黄曲霉毒素1与对照相比。的RNAi-288-74显示,黄曲霉毒素B 2和60%-100%减少63%-100%的减少黄曲霉毒素B 1, 图7。
用于实时PCR检测所述RNAi刀片的表达的引物示于图1。无胚花生种子的子叶进行了分析,以除去其自然防御和能够检测大多数暴露在区域的RNAi的潜 在影响真菌入侵,子叶。 RNA干扰的表达符合288-74未成熟子叶(黄色)用引物RT_5X_1插入检测是四折比(C T) = 1阈阴性对照,并通过引物组RT_5X_2为19倍以上的阈值,都在24小时温育。至少四分之三的在花生转化中使用五个连续的基因片段,5027,7223和7228(aflep,aflS / aflJ,和AFLC / PKSA,分别)通过RT-PCR检测(图1,6)。在成熟子叶在24小时,或在成熟的或未成熟的子叶在48小时温育后, 如图6(底部)未检测到的RNAi刀片的表达。
1| 花生线 | 采样时间 | 样品用于RT-PCR | 样品黄曲霉毒素分析(接种) | ñ种子红棕色 | ||||
| (非接种) | ||||||||
| 众议员1 | 众议员2 | 众议员3 | 众议员1 | 众议员2 | 众议员3 | |||
| RNA干扰(黄色) | 24小时 | 1 | 1 | 1 | 1 | 1 | 1 | 4.5 |
| 48小时 | 1 | 1 | 1 | 1 | 1 | |||
| 72小时 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| 控制 | 24小时 | 1 | 1 | 1 | 1 | 1 | 1 | 4.5 |
| (黄色) | 48小时 | 1 | 1 | 1 | 1 | 1 | 1 | |
| 72小时 | 1 | 1 | 1 | 1 | 1 | 1 | ||
表2中。 小样本设置实施例来分析在RNAi的花生种子的基因表达和黄曲霉毒素的积累。表达的完整的分析和黄曲霉毒素一到期组( 即,黄色),有三个采样时间(24,24,48和72小时) ,一式三份,将需要4.5种子,每个数字一个在表表示半子叶。
讨论
植物宿主RNAi介导的基因在真菌病原体的沉默已证明27,43,但是,也有没有出版物示出在植物中真菌毒素积累的RNAi介导的控制的可行性。这些研究花生的一个限制因素是缺乏以评估个别植株无黄曲霉毒素积累型的方法,因为叶出示附有地下荚真菌感染没有症状。此外,黄曲霉毒素的不正态分布积累,以及需要大的样本,进行化学分析15,16阻碍潜在的RNAi效应的定量上的单个植物。这里介绍的方法包括使用五个种子进行一式三份3 24小时间隔取样(表1, 图7)72小时实验。相比典型的黄曲霉毒素分析,无需小于100g的种子,我们的方法是特别适合于individua花生植物最初产生不超过两个或三个荚多个升转基因事件。
黄曲霉毒素合成的RNA介导的沉默已证明通过遗传转化黄曲霉 和 A. 寄生 。因为aflR是黄曲霉毒素的产生在A的主调节器黄曲霉及 寄生 44,45,就变成为在植物中的RNA介导的沉默一个有趣的目标。然而,遗传变异aflR已经显示曲霉种 46之间,而这些基因变异可能逃脱沉默如果没有完美的序列与在植物宿主中产生的RNAi的信号相匹配。因此,aflR是对沉默的载体p5XCAPD的目标之一,但不是唯一的一个。该aflR基因引入A.反向重复序列黄曲霉及 寄生通过转化导致沉默和最小或没有产品黄曲霉毒素47的离子(McDonald等。,2005年b)。另外,沉默AFLD基因防止黄曲霉毒素产量高达98% 在 A中黄曲霉及 寄生在直接转化48。为了增加在我们的系统中的成功的概率,花生转化用5个基因参与黄曲霉毒素生产A中反向重复片段菌 。这表明,使用p5XCAPD靶向用于沉默的几个基因在黄曲霉毒素合成途径,90%黄曲霉毒素B 1和B 2的下部-100%水平在线路288-72实现的,和较低的60-100%的水平积累线288-74与对照相比,当半子叶接种A.黄曲霉, 图4,图7,最重要的是,这种方法检测由线288-72,288-74 与控制整个实验中的黄曲霉毒素积累统计学显著差异通过应用参数统计的抽动, 图7。鉴于小样本大小,它以强调需要使用功能强大的方法来检测黄曲霉毒素,这些实验是由UPLC具有高的分辨率,5倍高的性能和比灵敏度高三倍分析是很重要的HPLC 49。
RNA干扰的表达插在288-74未成熟子叶(黄色),在24小时培养才被发现。通过RT-PCR在24小时未检测到对288-74成熟子叶所述RNAi插入件,或在任何成熟组在48小时, 图6,这同样的现象中观察到的RNAi其他转基因花生线(阿里亚斯,RS,2015未公布),其中只在24小时检测未成熟子叶通常RNAi的成绩单。 RNA样品与cDNA合成之前DNA酶处理后,数据标准化为肌动蛋白表达的水平,也没有证据DNA污染进行了观察。如若DNA已在样品中一直存在,它应该在48小时的样品中被检测为好,但始终事实并非如此。在35S启动子的控制下表达并不总是均匀的;它可受环境条件50,组织和发育阶段51,52的类型。与此同时,在RNA干扰的通路,mRNA降解的速率和siRNA衰减率可以显著53有所不同。这可能是与mRNA的RNA干扰的机构的快速降解可能阻止mRNA的检测在48小时温育。无论是在缺席48小时的表达是由于低35S启动子驱动的转录,或双链RNA被Dicer快速降解有待回答的问题。因此,检测的小RNA高通量测序将使通过RNA干扰54发生的过程更好地了解 在这些实验中。然而,由于RNA沉默扩散全身,主要是通过从photosynt韧皮部讨厌源蔗糖汇(在这种情况下,花生种子)55,黄曲霉毒素合成的沉默可以发生在种子未经所述RNAi的局部表达插入物。很多研究工作要做,以确定需要防止黄曲霉毒素积累在种子小干扰RNA(siRNA)的阈值水平。必须强调的事实,即,mRNA的所述RNAi的表达构建(图6)是重要的,和黄曲霉毒素B 1和B 2(图7)的累加结果显示不同的结果为未成熟(黄色)与。成熟(棕色)子叶。花生植物具有无限生长,也就是说,它们提出在收获的范围成熟荚果的, 图2,另外,从不同的成熟群体的种子会有不同的化学组合物,例如,2.4%的蔗糖在未成熟的种子,和1.9%在同场的条件56,57成熟种子。因此,要了解实际efficienc黄曲霉毒素积累的RNA介导的控制Y,分别分析成熟群体是重要的。
花生种子的天然防御是生产植物抗毒素,而变化所产生的化合物和它们的依赖于种子和环境条件58-61的成熟相对量的多样性,并且特别在更高的胚胎相比子叶62。胚胎也有显著较高浓度的核酸,DNA和RNA比子叶(Arias的RS,未发表)。由于花生种子成熟,改变他们的生理和化学成分发生63。酚类抗氧剂花生种皮形式缩合单宁具有抑菌活性的64;这也是在中果皮颜色反映成熟期,黄色到黑色35显而易见的,如单宁酸和酚类化合物的其内容与成熟65增大。因此,存在的t埃斯塔或在实验胚胎,由于其抗微生物性质,可能具有有限的真菌的生长,因此高估的RNAi沉默的作用,因此,它们被拆除。此外,去除种皮和胚胎有助于限制中的变化分析的来源,因为半子叶承载胚胎将有更多的植物防御素,更RNA含量。
除了由成熟群体和除去种皮和胚芽在这些实验的分析,它指出几个观察是很重要的:1)虽然结果示长达96小时培养时,建议使用不超过72小时,得到一致的结果,因为种子得到了96小时降解;和b),而从同一种子半子叶,虽然随机取样,不构成完全独立样本,RT-PCR和转基因事件中的黄曲霉毒素的积累表明种子间的最小变化。此外,精确的真菌孢子数,接种量第2微升,与应用避免滴落在两侧的子叶的切割表面孢子是重要的,以确保该发芽的孢子暴露于植物组织。在板上的水/琼脂应为1.5%(重量/体积),较软琼脂使孢子径流,如图4(底)的最后一帧上。 ;应该从一个特定的转基因事件的种子的可用性是有限的,采样可在重复的,而不是一式三份得到类似的结果(即,图7)来完成然而,一式三份样本将有助于降低标准误差。该方法的唯一限制是它要求一个高度敏感的系统(UPLC)黄曲霉毒素检测/定量,但在同一时间这降低高估的RNAi的效果的不应该被较不敏感的方法来检测的黄曲霉毒素的概率。
总之,本方法提供了在第一时间来研究的可靠方法RNA干扰中黄曲霉毒素的控制效果。减少的时间,从整个种植季节进行实验,以不到一周的时间,这种方法将极大地加快对缓解和/或消除黄曲霉毒素的RNA干扰花生/ 曲霉菌病害系统研究。
披露声明
The authors have nothing to disclose or any conflicts of interest.
致谢
This work received the financial support of USDA-ARS CRIS project 6604-21000-004-00D, CRIS project 6604-42000-008-00D, and USAID Feed-the-Future program Agreement number 58-0210-3-012. We thank Valerie Orner, LaTanya Johnson, Joseph Powell and Kathy Gray for their technical assistance. Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the US Department of Agriculture.
材料
| Name | Company | Catalog Number | Comments |
| Primers, oligonucleotides | DNA Technologies, Coralville, IA, USA | n/a | |
| Dneasy Plant Mini Kit | Qiagen, Valencia, CA | 69106 | |
| Czapek Dox agar medium | Oxoid, by Thermo Fisher Scientific, Waltham, MA | CM0095 | |
| Agar | Thermo Fisher Scientific, Waltham, MA | BP 1423 | |
| Freezer -80 °C | n/a | n/a | |
| Aluminum Oxide, Al2O3 | Fisher Scientific | A941 | |
| SPE Reservoirs 1.5 ml | Grace Davison Discovery Scientific | 210011 | |
| Frits for 1.5 ml SPE reservoir | Grace Davison Discovery Scientific | 211401 | |
| Autosampler vials | Waters Corporation, Milford, MA | 186005221 | |
| Waters Acquity Ultra-Performance Liquid-Chromatography (UPLC) instrument; UPLC-H-Class Quaternary Solvent Manager; UPLC Sample Manager; UPLC Fluorescent detector (FLR); UPLC BEH C18 2.1 mm x 50 mm, 1.7 mm column | Waters Corporation, Milford, MA | ||
| Finnigan LCQ Advantage MAX ion trap mass spectrometer, with Xcalibur version 1.4 software | Thermo Electron Corp., San Jose, CA | ||
| Aflatoxin standards, B1, B2, G1 and G2 | Sigma-Aldrich, St. Louis, MO | A6636; A9887; A0138; A0263 | |
| Systat Software 12.2 | SYSTAT Software Inc., Point Richmond, CA | ||
| Trizol reagent | Invitrogen, CA | 15596-018 | |
| SuperScript III First Strand Synthesis Super Mix | Invitrogen, CA | 11752-050 | |
| ABI 7500 Real-Time PCR | Lifetechnologies, Grand Island, NY | 4406984 | |
| Luria Broth-Miller | Fisher Scientific | R453642 | |
| pENTR1A | Invitrogen, CA | A10462 | |
| LR Clonase II enzyme mix | Invitrogen, CA | 11791-020 | |
| T4 DNA Ligase | NEB Biolabs | M0202L | |
| Gelrite | Sigma-Aldrich, St. Louis, MO | G1919 | |
| Acetosyringone | Sigma-Aldrich, St. Louis, MO | D134406 | |
| QIAcube robot workstation | Qiagen, Valencia, CA | 9001292 | |
| Antibiotics: kanamycin, cefotaxime, gentamicin; streptomycin | Goldbio, St. Louis, MO | cef.: C-104-25; kan: K-120-5; gent.: G-400-1; strep.: S-150-50 | |
| Platinum Taq DNA Polymerase High Fidelity | Invitrogen, CA | 11304-029 |
参考文献
- Williams, J. H., et al. Human aflatoxicosis in developing countries: a review of toxicology, exposure, potential health consequences, and interventions. The American Journal of Clinical Nutrition. 80, 1106-1122 (2004).
- American Association for Cancer Research: AACR. An evaluation of chemicals and industrial processes associated with cancer in humans based on human and animal data: IARC Monographs Volumes 1 to 20. Cancer Research. 40, 1-12 (1980).
- Turner, P. C. The molecular epidemiology of chronic aflatoxin driven impaired child growth. Scientifica. , (2013).
- Rasooly, R., Hernlem, B., He, X., Friedman, M. Non-linear relationships between aflatoxin B1 levels and the biological response of monkey kidney vero cells. Toxins (Basel). 5, 1447-1461 (2013).
- Gong, Y. Y., et al. Determinants of aflatoxin exposure in young children from Benin and Togo, West Africa: the critical role of weaning. International Journal of Epidemiology. 32, 556-562 (2003).
- Eaton, D. L., Groopman, J. D. The toxicology of aflatoxins: human health, veterinary, and agricultural significance. , Academic Press. (1994).
- Murugavel, K. G., et al. Prevalence of aflatoxin B1 in liver biopsies of proven hepatocellular carcinoma in India determined by an in-house immunoperoxidase test. Journal of Medical Microbiology. 56, 1455-1459 (2007).
- Wang, J. S., et al. Hepatocellular carcinoma and aflatoxin exposure in Zhuqing Village, Fusui County, People's Republic of China. Cancer Epidemiology, Biomarkers & Prevention. 10, American Association for Cancer Research. 143-146 (2001).
- Azziz-Baumgartner, E., et al. Case-control study of an acute aflatoxicosis outbreak, Kenya, 2004. Environmental Health Perspectives. 113, 1779-1783 (2005).
- Lye, M. S., Ghazali, A. A., Mohan, J., Alwin, N., Nair, R. C. An outbreak of acute hepatic encephalopathy due to severe aflatoxicosis in Malaysia. American Journal of Tropical Medicine and Hygiene. 53, 68-72 (1995).
- Villers, P. Aflatoxins and safe storage. Frontiers in Microbiology. 5, 158(2014).
- Kensler, T. W., Roebuck, B. D., Wogan, G. N., Groopman, J. D. Aflatoxin: A 50-year odyssey of mechanistic and translational toxicology. Toxicological Sciences. 120, S28-S48 (2011).
- Dorner, J. W., Cole, R. J., Wicklow, D. T. Aflatoxin reduction in corn through field application of competitive fungi. Journal of Food Protection. 62, 650-656 (1999).
- Cotty, P. J., Bhatnagar, D. Variability among atoxigenic Aspergillus flavus strains in ability to prevent aflatoxin contamination and production of aflatoxin biosynthetic-pathway enzymes. Applied and Environmental Microbiology. 60, 2248-2251 (1994).
- Whitaker, T. B. Standardisation of mycotoxin sampling procedures: an urgent necessity. Food Control. 14, 233-237 (2003).
- Whitaker, T. B., Dorner, J. W., Giesbrecht, F. G., Slate, A. B. Variability among aflatoxin test results on runner peanuts harvested from small field plots. Peanut Science. 31, 59-63 (2004).
- Fire, A., et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 391, 806-811 (1998).
- Rafael, D., et al. EMT blockage strategies: Targeting Akt dependent mechanisms for breast cancer metastatic behaviour modulation. Current Gene Therapy. , (2015).
- Li, G., Chang, H., Zhai, Y. P., Xu, W. Targeted silencing of inhibitors of apoptosis proteins with siRNAs: a potential anti-cancer strategy for hepatocellular carcinoma. Asian Pacific. Journal of Cancer Prevention: APJCP. 14, 4943-4952 (2013).
- Koldehoff, M. Targeting bcr-abl transcripts with siRNAs in an imatinib-resistant chronic myeloid leukemia patient: challenges and future directions. Methods in Molecular Biology. 1218, 277-292 (2015).
- Zhang, J., et al. Pest control. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science. 347, 991-994 (2015).
- Ajjappala, H., Chung, H. Y., Sim, J. S., Choi, I., Hahn, B. S. Disruption of prefoldin-2 protein synthesis in root-knot nematodes via host-mediated gene silencing efficiently reduces nematode numbers and thus protects plants. Planta. 241, 773-787 (2015).
- Jose, A. M., Hunter, C. P. Transport of sequence-specific RNA interference information between cells. Annual Review of Genetics. 41, 305-330 (2007).
- Vazquez, F., Hohn, T. Biogenesis and biological activity of secondary siRNAs in plants. Scientifica. , Hindawi Publishing Corporation. (2013).
- Tinoco, M. L. P., Dias, B. B. A., Dall'Astta, R. C., Pamphile, J. A., Aragao, F. J. L. In vivo trans-specific gene silencing in fungal cells by in planta expression of a double-stranded RNA. BMC Biology. 8, (2010).
- Govindarajulu, M., Epstein, L., Wroblewski, T., Michelmore, R. W. Host-induced gene silencing inhibits the biotrophic pathogen causing downy mildew of lettuce. Plant Biotechnology Journal. , (2014).
- Yin, C., Jurgenson, J. E., Hulbert, S. H. Development of a host-induced RNAi system in the wheat stripe rust fungus Puccinia striiformis f. sp. tritici. Molecular Plant-Microbe Interactions. 24, 554-561 (2011).
- Ghag, S. B., Shekhawat, U. K., Ganapathi, T. R. Host-induced post-transcriptional hairpin RNA-mediated gene silencing of vital fungal genes confers efficient resistance against Fusarium. wilt in banana. Plant Biotechnology Journal. 12, 541-553 (2014).
- Filichkin, S. A., et al. Efficiency of gene silencing repeats vs. transitive RNAi in Arabidopsis: direct inverted vectors. Plant Biotechnology Journal. 5, 615-626 (2007).
- Sciaky, D., Montoya, A. L., Chilton, M. D. Fingerprints of Agrobacterium Ti Plasmids. Plasmid. 1, 238-253 (1978).
- Clark, D. J., Maaloe, O. DNA Replication and Division Cycle in Escherichia coli. Journal of Molecular Biology. 23, 99-112 (1967).
- Murashige, T., Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plantarum. 15, 473-497 (1962).
- Srinivasan, T., Kumar, K. R. R., Kirti, P. B. Establishment of efficient and rapid regeneration system for some diploid wild species of Arachis. Plant Cell Tissue and Organ Culture. 101, 303-309 (2010).
- Gomes, A. L. V., et al. Single-tube nested PCR using immobilized internal primers for the identification of dengue virus serotypes. Journal of Virology Methods. 145, 76-79 (2007).
- Williams, E. J., Drexler, J. S. A non-destructive method for determining peanut pod maturity. Peanut Science. 8, 134-141 (1981).
- Sobolev, V. S., Dorner, J. W. Cleanup procedure for determination of aflatoxins in major agricultural commodities by liquid chromatography. Journal of AOAC International. 85, 642-645 (2002).
- Empower Software, Getting Started Guide. , Waters Corporation. Milford, MA. Available from: http://sites.chem.colostate.edu/diverdi/C431/experiments/high%20pressure%20liquid%20chromatography/references/Empower%20getting%20started%2071500031203rA.pdf (2002).
- Biselli, S., Hartig, L., Wegner, H., Hummert, C. Analysis of Fusarium. toxins using LC-MS-MS: Application to various food and feed matrices. LC GC North America. 23, 404-413 (2005).
- Arias, R. S., Sobolev, V. S., Orner, V. A., Dang, P. M., Lamb, M. C. Potential involvement of Aspergillus flavus laccases in peanut invasion at low water potential. Plant Pathology. 63, 353-363 (2014).
- Dang, P. M., Chen, C. Y., Holbrook, C. C. Evaluation of five peanut (Arachis hypogaea) genotypes to identify drought responsive mechanisms utilising candidate-gene approach. Functional Plant Biology. 40, 1323-1333 (2013).
- Schmittgen, T. D., Livak, K. J. Analyzing real-time PCR data by the comparative C-T method. Nature Protocols. 3, 1101-1108 (2008).
- Amaike, S., Keller, N. P. Aspergillus flavus. Annual Review of Phytopathology. 49, 107-133 (2011).
- Nowara, D., et al. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell. 22, 3130-3141 (2010).
- Woloshuk, C. P., et al. Molecular characterization of aflR, a regulatory locus for aflatoxin biosynthesis. Applied and Environmental Microbiology. 60, 2408-2414 (1994).
- Price, M. S., et al. The aflatoxin pathway regulator AflR induces gene transcription inside and outside of the aflatoxin biosynthetic cluster. FEMS Microbiology Letters. 255, 275-279 (2006).
- Ehrlich, K. C., Montalbano, B. G., Cotty, P. J. Sequence comparison of aflR from different Aspergillus. species provides evidence for variability in regulation of aflatoxin production. Fungal Genetics and Biology. 38, 63-74 (2003).
- McDonald, T., Brown, D., Keller, N. P., Hammond, T. M. RNA silencing of mycotoxin production in Aspergillus and Fusarium species. Molecular Plant Microbe Interactions. 18, 539-545 (2005).
- Abdel-Hadi, A. M., Caley, D. P., Carter, D. R., Magan, N. Control of aflatoxin production of Aspergillus flavus. and Aspergillus parasiticus. using RNA silencing technology by targeting aflD. (nor-1) gene. Toxins (Basel). 3, 647-659 (2011).
- Swartz, M. E. Ultra performance liquid chromatography (UPLC): An introduction: Separation Science Redefined. LCGC North America. , Suppl ement 8. 8-14 (2005).
- Maghuly, F., Khan, M. A., Fernandez, E. B., Druart, P., Watillon, B., Laimer, M. Stress regulated expression of the GUS-marker gene (uidA) under the control of plant calmodulin and viral 35S promoters in a model fruit tree rootstock: Prunus incisa x serrula. Journal of Biotechnology. 135, 105-116 (2008).
- de Mesa, M. C., Santiago-Doménech, N., Pliego-Alfaro, F., Quesada, M. A., Mercado, J. A. The CaMV 35S promoter is highly active on floral organs and pollen of transgenic strawberry plants. Plant Cell Reports. 23, 32-38 (2004).
- Sunilkumar, G., Mohr, L., Lopata-Finch, E., Emani, C., Rathore, K. S. Developmental and tissue-specific expression of CaMV 35S promoter in cotton as revealed by GFP. Plant Molecular Biology. 50, 463-474 (2002).
- Groenenboom, M. A. C., Maree, A. F. M., Hogeweg, P. The RNA silencing pathway: The bits and pieces that matter. PLoS Computational Biology. 1, 155-165 (2005).
- Zhao, D., Song, G. Q. High-throughput sequencing as an effective approach in profiling small RNAs derived from a hairpin RNA expression vector in woody plants. Plant Science: an International Journal of Experimental Plant Biology. 228, 39-47 (2014).
- Kamthan, A., Chauduri, A., Kamthan, M., Datta, A. Small RNAs in plants: recent development and application for crop improvement. Frontiers in Plant Science. 6, 208(2015).
- Manda, A., Bodapati, P. N., Rachaputi, N. C., Wright, G., Fukai, S. Aflatoxins and their relationship with sugars in peanut (Arachis hypogaea L). 4th International Crop Science Congress, 2004, , Available from: http://www.cropscience.org.au/icsc2004/poster/5/1/3/625_manda.htm (2004).
- Uppala, S. S. Factors affecting pre-harvest aflatoxin contamination of peanut (Arachis hypogaea L). , Auburn University. (2011).
- Sobolev, V. S. Localized production of phytoalexins by peanut (Arachis hypogaea) kernels in response to invasion by Aspergillus species. Journal of Agricultural and Food Chemistry. 56, 1949-1954 (2008).
- Sobolev, V. S., Guo, B. Z., Holbrook, C. C., Lynch, R. E. Interrelationship of phytoalexin production and disease resistance in selected peanut genotypes. Journal of Agricultural and Food Chemistry. 55, 2195-2200 (2007).
- Sobolev, V. S., Neff, S. A., Gloer, J. B. New stilbenoids from peanut (Arachis hypogaea) seeds challenged by an Aspergillus caelatus strain. Journal of Agricultural and Food Chemistry. 57, 62-68 (2009).
- Dorner, J. W., Cole, R. J., Sanders, T. H., Blankenship, P. D. Interrelationship of kernel water activity, soil temperature, maturity, and phytoalexin production in preharvest aflatoxin contamination of drought-stressed peanuts. Mycopathologia. 105, 117-128 (1989).
- Sobolev, V. S. Production of phytoalexins in peanut (Arachis hypogaea) seed elicited by selected microorganisms. Journal of Agricultural and Food Chemistry. 61, 1850-1858 (2013).
- Basha, S. M. M., Cherry, J. P., Young, C. T. Changes in free amino acids, carbohydrates, and proteins of maturing seeds from various peanut (Arachis hypogaea L.) cultivars. Cereal Chemistry. 53, 586-596 (1976).
- Lansden, J. A. Aflatoxin inhibition and fungistasis by peanut tannins. Peanut Science. 9, 17-20 (1982).
- Yen, G. C., Duh, P. D., Tsai, C. L. Relationships between antioxidant activity and maturity of peanut hulls. Journal of Agricultural and Food Chemistry. 41, 67-70 (1993).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。