
Method Article
Controle mediado por RNAi de aflatoxinas em amendoim: método para analisar a produção de micotoxinas e Transgene Expression na Amendoim /
Neste Artigo
Resumo
Nós demonstramos um método para a análise de aflatoxinas e a expressão do transgene em sementes de amendoim que contêm sinais de ARN de interferência para silenciar genes aflatoxina-síntese no fungo Aspergillus flavus. Controlo mediada por ARNi de micotoxinas em plantas não foi reportado anteriormente.
Resumo
A Organização para a Alimentação e Agricultura das Nações Unidas estima que 25% das lavouras de alimentos do mundo estão contaminados com aflatoxinas. Isso representa 100 milhões de toneladas de alimentos a ser destruídos ou desviados para consumo não humano a cada ano. As aflatoxinas são substâncias cancerígenas poderosos normalmente acumulados pelos fungos Aspergillus flavus e A. parasiticus em cereais, nozes, tubérculos e outros produtos agrícolas. O silenciamento de genes cinco aflatoxina-síntese através de interferência de RNA (RNAi) em plantas de amendoim foi usada para controlar a acumulação de aflatoxina a seguir à inoculação com A. flavus. Anteriormente, existia nenhum método para analisar a eficácia de RNAi em eventos transgênicos amendoim indivíduo, uma vez que estes geralmente produzem poucas sementes e os métodos tradicionais de grandes experimentos de campo em condições de aflatoxinas-propício não foram uma opção. No campo, a probabilidade de encontrar sementes naturalmente contaminadas é muitas vezes 1/100 a 1/1,000. Além disso, a contaminação por aflatoxinas não é uniformemente distribuída. O nosso método utiliza algumas sementes por modificação genética, com pequenos pedaços processados para PCR em tempo real (RT-PCR) ou pequena sequenciação de ARN, e para a análise da acumulação de aflatoxinas por cromatografia líquida de ultra-desempenho (UPLC). Linhagens de amendoim expressando RNAi 288-72 e 288-74, mostrou redução de até 100% (p≤0.01) em aflatoxina B 1 e B 2 em comparação com o controle que acumulou até 14.000 ng. G -1 de aflatoxinas B 1, quando inoculadas com A. aflatoxigênicos flavus. Como referência, o total máximo de aflatoxinas permitidos para consumo humano nos Estados Unidos é de 20 ng g -1.. Este protocolo descreve a aplicação do controlo mediado por RNAi de aflatoxinas em sementes de amendoim transgénicos e métodos para a sua avaliação. Acreditamos que a sua aplicação no melhoramento de amendoim e outras culturas trará rápido avanço nesta importante área da ciência, Medicina e nutrição humana, e irá contribuir significativamente para o esforço internacional para controlar aflatoxinas e, potencialmente, outras micotoxinas nos principais culturas alimentares.
Introdução
Cerca de 4,5 bilhões de pessoas estão cronicamente expostos a aflatoxinas 1, os agentes cancerígenos mais poderosas conhecidas na natureza 2. Estas micotoxinas contaminam 25% das culturas alimentares no mundo 3, incluindo milho, mandioca, arroz, nozes, cereais e especiarias. 4. As aflatoxinas causam raquitismo em crianças de 5, prejudicar o sistema imunológico 6, estão presentes em 58% dos carcinomas hepatocelulares-em biópsias humanos 7,8, e matar centenas de pessoas durante os surtos periódicos de aflatoxicosis 9,10. As aflatoxinas são micotoxinas derivadas de polic�ido normalmente produzidas por Aspergillus flavus e A. parasiticus; aflatoxinas B 1 e B 2 são produzidos por A. flavus, enquanto A. parasiticus também produz L 1 e L 2. A estrutura química destes compostos e um cromatograma que mostra a separação por UPLC são mostrados na Figura 1.

Figura 1. As aflatoxinas e RNAi inserir Top:. Estrutura química (esquerda) e exemplo de cromatograma (à direita) dos quatro aflatoxinas derivados de polic�ido mais comuns: B 1, B 2, G 1 e G 2, produzida por Aspergillus parasiticus, A . flavus produz B 1 e B 2 inferior: Esquema de fragmentos de genes no RNAi construir p5XCAPD utilizado para transformação de amendoim, números sob setas são números de genes-fragmento de adesão no genoma Aspergillus flavus;. PIV2: intrão de batata; pares de bases;: pb RT_5X_1 e RT_5X_2:. Real-Time PCR locais iniciadores Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}
Sozinho perdas econômicas nas exportações devido a aflatoxinas em amendoim ultrapassar US $ 450 milhões de dólares norte-americanos, se calculado com base no 4 ng. G -1 limite de aflatoxina permitidos para consumo humano na União Europeia 11. As aflatoxinas são conhecidos há 60 anos 12; no entanto, embora muitas práticas agrícolas foram desenvolvidos para mitigar seus efeitos, incluindo a aplicação de outras cepas fúngicas 13,14, nenhum método coerente de controlo existe, e variedades de plantas resistentes não estão disponíveis. Teste de germoplasma de plantas para resistência a aflatoxinas é particularmente difícil, porque mesmo sob condições propícias para a invasão do patógeno, a acumulação de micotoxinas é imprevisível e não segue uma distribuição normal. Assim, as experiências geralmente requerem grandes áreas de plantação, centenas de sementes e várias amostras de 100-1,700 g para reduzir a variabilidade de dados a 15,16.
Interferência de ARN foidescobriram em 1998 17; e os benefícios de "silenciar" estão actualmente a ser explorada em uma série de novas aplicações, por exemplo., em terapias humanas contra o câncer de mama metastático 18, câncer de fígado 19, leucemia mielóide 20, e na protecção das plantas contra insetos e nematóides 22 21. Nas plantas, sinais de interferência de RNA pode viajar uma célula para outra, com pequeno RNA de interferência (siRNA) e RNA de elevado peso molecular sendo responsável pelo pós-transcricional gene silenciamento sistêmica 23,24, mesmo dentro de fungos patogénicos que estão em contato próximo com planta hospedeira 25. A eficácia do RNAi na silenciamento mediado por planta de genes de fungos-patógeno foi descrito em alguns patossistemas de plantas, para estes, exame visual dos sintomas na parte aérea das plantas (folhas) permitiu a quantificação da doença, ou seja, oomiceto Bremia em alface 26 , no trigo Puccinia 27 e Fusarium em 28 de banana. Muito mais difícil é para avaliar a eficácia de ARNi para controlar micotoxinas em plantas, particularmente as aflatoxinas em amendoins como as folhas não apresentam sintomas de infecção, os órgãos invadidos (sementes) estão sob várias polegadas de solo, a ocorrência de infecção é imprevisível, e apenas química análise pode determinar a presença de aflatoxinas. Além disso, cada evento transgênico no amendoim normalmente produz poucas sementes (4-6 por planta); Portanto, o teste tradicional para um não-acumulação de aflatoxina traço em grandes parcelas de terreno, com duração de safras inteiras, e usando centenas de sementes não é viável. É descrito um método para analisar aqui em menos de uma semana, de ARNi sementes de amendoim para a presença de transgene e para um não-aflatoxina acumulação traço, usando apenas poucas sementes.
Protocolo
1. Construir Molecular e amendoim Transformation
- Combinar fragmentos de DNA de A. cinco genes flavus, AFL2G_07223 (AFLs ou aflJ), AFL2G_07224 (AFLR), AFL2G_07228 (AFLC / pksA / pksL1), AFL2G_07731 (PES1) e AFL2G_05027 (bomba de efluxo aflatoxina, aflep). Para isso, use os seguintes iniciadores e ultramers: DIR-1, Short-Dir1-R, DIR-2-invertido, Short-Dir2-R, DirAll-NCO-Rv, e DirAll-BamEco-Fw, a Tabela 1.
- Adicione DIR-1 de cadeia dupla por 5 ciclos de PCR (reacção em 25 mL; 95 ° C 2 min, seguido de 5 ciclos de 94 ° C 45 seg, 55 ° C 30 seg, 68 ° C 15 seg) utilizando polimerase de ADN de acordo com o fabricante do instruções e iniciador Short-Dir1-R para deixar um CCCGT 3 'overhang. Repita essas etapas para fazer DIR-2-invertida cadeia dupla utilizando o iniciador de curto Dir2-R para deixar um ACGGG 3 'overhang complementar a DIR-1.
- Ligadura da Fragme dois 199 pbnts com T4 DNA ligase como por instruções do fabricante. PCR amplificar o fragmento de 393 pb resultante como indicado em 1.1.1, utilizando iniciadores DirAll-CACC-FW e DirAll-NCO-RV (Tabela 1), e clonar o produto usando técnicas convencionais em pENTR1A fazer plasmídeo p2 + 4ENTR.
- Reunir em p2 + 4ENTR em pCAPD 29 (Acesso NCBI: KC176455.1) usando clonase LR II de mistura de enzimas de acordo com as instruções do fabricante para fazer plasmídeo p5XCAPD, e transformá-la em Escherichia coli DH5a, utilizando técnicas convencionais seguidas por sequenciação parcial. Nota: A inserção de ARNi completa é mostrada na Tabela 1.
- Transformar Agrobacterium estirpe C58C1 30 com o plasmídeo p5XCAPD como relatado anteriormente 30, e utilizar a bactéria resultante para transformar plantas de amendoim da seguinte forma:
- Crescer a 30 ° C, a Agrobacterium abrigando p5XCAPD, usar-se para esta, 50 ml de caldo LB suplementado com 500ng ml -1 estreptomicina, 25 ug ml -1 gentamicina, 10 ug ml -1 canamicina e agitar a cultura a 250 rpm até atingir uma OD 260.
- Colher as células de Agrobacterium por centrifugação (6000 x g) durante 10 minutos, ressuspender em 50 ml AB meio mínimo 31 com 100 acetosiringona mM durante 1 hora e lugar na suspensão bacteriana nos explantes de 10-14 dias da germinação Exp27-1516, corredor -tipo linhagem de amendoim. Secar os explantes em 3MM papel absorvente após 30 min, e colocá-los em meio shoot-indução (SIM) [MS sal de 32, 3% de sacarose, 20 mM benzilaminopurina (BAP), thidiazuron 10 mM (TDZ), pH 5,8, 0,3 % de goma de gel] sem antibióticos no escuro por três dias.
- Faça a seleção e regeneração tecidual, como relatado anteriormente 33. Mova tecidos para o SIM (cefotaxima 500? M e 100 M canamicina) para a formação de filmagem, com transferências bi-semanais durante 2 meses. Então lugarexpansão brotos em meio shoot-alongamento (SEM) [5 mM BAP, giberelina 1 PM (GA 3)], bi-semanal por vários meses.
- Coloque tiros individuais, 2 cm de tamanho, em meio-indução de raízes (RIM) [1/2 MS, 1,5% de sacarose, ácido α-5? M-naftaleno acético (ANA), 2,5 mM de ácido indolbutírico (IBA)], em seguida, se aclimatar as mudas e transferi-los para o efeito estufa.
2. Identificação de plantas de amendoim abrigar RNAi para silenciar genes aflatoxina Síntese
- Uso planta mini-kit em uma estação de trabalho do robot com 200 eluição ul de acordo com as instruções do fabricante para extrair DNA a partir de folhas jovens de plantas de amendoim que eram sujeitas ao processo de transformação (conforme descrito anteriormente) com o RNAi construir p5XCAPD (Figura 1) que tem como espinha dorsal plasmídeo pCAPD 29 para o silenciamento do gene.
- Tela as amostras de DNA de um único tubo de nested PCR (STN-PCR), como descrito PREVIously 34 para detectar o marcador selecionável NPTII e o RNAi inserir a partir p5XCAPD. Clonal propagar de cortes (3-4 nós) de PCR positivos plantas para produzir sementes suficientes para testes nesta primeira geração.
- Use quatro diluições de 2 vezes de ADN (50-100 ng mL -1 antes da diluição) em todas as reacções de PCR-STN. Para NPTII, utilizar os iniciadores externos PCAPD 5714F: 5'- AGGCTATTCGGCTATGACTG-3 'e 6446R PCAPD: 5'-CGTCAAGAAGGCGATAGAAG-3', e os iniciadores internos PCAPD 5730F: 5'- ACTGGGCACAACAGACAATC-3 'e 6249R PCAPD: 5'-ATATTCGGCAAGCAGGCATC- 3 '.
- Para a detecção de ARNi inserir em reacções de STN-PCR utilizam iniciadores externos 35S-PDSFw: 5'-CCTAACAGAACTCGCCGTAA-3 'e DirAll-NCO-RV: 5'-ATGCCATGGGGTTATTGGGTGCAGAATGG-3' iniciadores internos, e Probe_5027_Fw: 5'-3-gtatttgtgaccatgtttctg 'e Probe_7228_Rv: 5'-GGACGGATAGTAAACTGCGG-3'.
- Colheita das vagens das plantas positivas STN-PCR e da con trolo plantas cultivadas nas mesmas condições. Passo crítico: Certifique-se de que o controle de plantas são cultivadas nas mesmas condições e na mesma estação como as plantas RNAi. Explosão da água das vagens com uma lavadora de alta pressão em baixa intensidade ou raspar com a mão para remover exocarp, determinar a cor do mesocarpo, colocando as vagens em uma placa de maturidade e vagens separados em grupos (amarelo, laranja, marrom e preto) 35 (Figura 2).

Figura 2. Preparação de vagens de amendoim para análise. Esquerda: amendoim tamanhos diferentes encontrados na colheita como amendoim é uma planta de crescimento indeterminado; Center: colocação de amendoim na cesta do metal para a remoção de pressão de água de exocarp; Direita: grupos de maturação de cor mesocarpo em uma placa de perfil de amendoim (amarelo, laranja, marrom e preto)."Target =" _ blank 98fig2large.jpg "> Clique aqui para ver uma versão maior desta figura.
3. Setup Experimental
- Remover a casca, o processo de amarelo e castanho sementes separadamente. Calcule o número de sementes para uso no experimento de acordo com a Tabela 2. Note-se que, no mínimo, três pedaços de sementes (parte 1 semente = metade de cotilédones) por linha de amendoim e data de amostragem é necessário para realizar uma análise estatística e reduzir o erro padrão.
| Nome | Seqüência | ||
| DIR-1 | 5'- GCCAGCTCAAAAGTGCGATGCACCAAGGAGAAACCGGCCTGTGCT CGGTGTATCGAACGTGGTCTTGCCTGTCAATACATGGTCGTATTTG TGACCATGTTTCTGGTGGCATTGGACCGTCTTGTCATCTCTACAGCC ATTCCCCAGATCACGGACGAATCCCCTGCATCTACGCGCACGCATC ACTTGGGGTACCCGT-3 ' | ||
| Curto-R-Dir1 | 5'-Fosfato-TACCCCAAGTGATGCGTGCGCG-3 ' | ||
| DIR-2-invertida | 5'- GGTTATTGGGTGCAGAATGGTAAACCACCCAACAGTACGCGAAATG TCAATTCCAGAGTCCCAAACCTCCCTACCGTGGCCTGGACGGATAG TAAACTGCGGAGCTTGGGAACAAAATCCGCTGTCTGATCGCCGAAG AGAAAGAGTTGCCTTGATTGAGCCGCATCGAGGACAGGTTGTGTTG CTGTTGATAGACGGG-3 ' | ||
| Curto-R-Dir2 | 5'-Fosfato-CTATCAACAGCAACACAACC-3 ' | ||
| DirAll-CACC-FW | 5'-CACCGCCAGCTCAAAAGTGCGATGC-3 ' | ||
| DirAll-NCO-Rv | 5'-ATGCCATGGGGTTATTGGGTGCAGAATGG-3 ' | ||
| DirAll-BamEco-FW | 5'-ATGGGATCCGAATTCGCCAGCTCAAAAGTGCGATGC-3 ' | ||
| Inserção RNAi completa | 5'- GCCAGCTCAAAAGTGCGATGCACCAAGGAGAAACCGGCCTGTGCT CGGTGTATCGAACGTGGTCTTGCCTGTCAATACATGGTC / GTATTTG TGACCATGTTTCTGGTGGCATTGGACCGTCTTGTCATCTCTACAGCC ATTCCCCAGATCACGGACGAAT / CCCCTGCATCTACGCGCACGCAT CACTTGGGGTACCCGTCTATCAACAGCAACACAACCTGTCCTCGAT GCG / GCTCAATCAAGGCAACTCTTTCTCTTCGGCGATCAGACAGCG GATTTTGTTCCCAAGCTCCGCAGTTTACTATCCGTCCA / GGCCACGG TAGGGAGGTTTGGGACTCTGGAATTGACATTTCGCGTACTGTTGGG TGGTTTACCATTCTGCACCCAATAACC-3 ' | ||
. Tabela 1. Oligonucleotides e ultramers utilizados para construir o RNAi construir p5XCAPD Phos: fosforilada extremidade 5 '; "/" Separa os fragmentos de genes cinco utilizado; Inserção completa RNAi: sequência utilizada como 2 repetições invertidas para formar p5XCAPD.
- Coloque sementes de amendoim inteiros em uma única camada de forma a cobrir a parte inferior de um copo estéril. Adicionar solução de 75% de etanol / água (v / v) para cobrir as sementes e, em seguida, adicionar um volume igual da mesma solução. Incubar à temperatura ambiente por 30 segundos e, em seguida, enxágüe com água deionizada estéril (ADE).
- Adicionar 2% de hipoclorito para a proveta contendo as sementes de etanol tratada como se segue: adicionar suficiente solução de hipoclorito de 2% para cobrir as sementes, em seguida, adicionar um volume igual da mesma solução e incubar durante 5 min. Passo crítico: Lavar cuidadosamente três vezes com um volume de SDW equivalente a 5 vezes o volume de hipoclorito usado (Figura 3).

Figura 3. Instalação experimental. Top: Superfície de esterilização de sementes de amendoim antes e depois de hipoclorito, a remoção do tegumento (testa); Oriente: remoção de embrião e ver de perto embrião, em seguida, corte cotilédones ao meio; inferior: cotilédones meio em água destilada estéril, apagando no absorvente estéril papel, mossas em ágar água, e colocação dos cotilédones meio (corte lateral para cima) sobre a superfície do ágar. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}
- Permitir que as sementes esterilizadas de superfície para absorver submersa em SDW durante 2 horas. Coloque as sementes em uma placa de Petri estéril, remover os casacos de semente com uma pinça, separe os cotilédones, e com um bisturi remover os embriões. Nota:Os embriões podem ser descartados ou usados para regenerar plantas novas.
- Corte cada cotilédones ao meio usando um bisturi. Para evitar desidratação, manter as peças de sementes corte em água estéril até que todas as sementes são processadas.
- Ter preparado placas de Petri contendo ágar água estéril (1,5% de agar / água; w / v), um para cada um de três pedaços de sementes. Faça pequenas amolgadelas no agar usando uma pinça, seque rapidamente o excesso de água dos pedaços de sementes em toalhas de papel estéreis, e em seguida, coloque os pedaços de sementes (corte rosto para cima) nas placas de agar de água /.
- A partir de uma cultura fresca de aflatoxigênicos Aspergillus flavus NRRL 3357 num meio de agar de Czapek cultivadas a 25 ° C durante 10 dias, preparar uma suspensão de 50.000 esporos por mL SDW, contadas com um hemocitómetro.
- Coloque 2 ml da suspensão de esporos sobre as superfícies de corte de cada peça semi-cotilédone evitando o escoamento nos lados, para garantir que os esporos se exposto ao tecido da semente que abriga RNAi (Figura 4).

Figura 4. Inoculação e incubação para a análise das aflatoxinas. Top: Metade cotilédones, a inoculação com suspensão de esporos, e Aspergillus flavus crescimento micelial em metade cotilédone após 24 horas de incubação inferior: esquerda:. Incubação de 48 h em 1,5% agar; Centro: incubação durante 72 horas em 1,5% agar; direita:. exemplo de configuração experimental incorreta, a incubação durante 72 horas em 0,5% agar Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}
- Incubar as placas de Petri contendo os cotilédones inoculados meio e não inoculadas a 30 ° C no escuro até à colheita.
4. A amostragem para análise de aflatoxina e Expressão Gênica
- Coletar amostras emtriplicado, a 24, 48 e 72 h (96 h opcional) de incubação, tanto por RT-PCR e para a análise das aflatoxinas, cada replicar sendo uma peça (metade cotilédone). Escolha amostras aleatoriamente em diferentes placas em cada data de amostragem. Utilizando um tecido, suavemente remover esporos de fungos e excesso de ágar antes de colocar pedaços de sementes em frascos ou tubos de ensaio.
- Para a análise das aflatoxinas, coloque cada repetição (uma peça) em um 4 ml parafuso tampa do frasco de vidro e armazenar a -80 ° C. Nota: Dada a estabilidade química de aflatoxinas, as amostras podem ser armazenados nesta condição durante vários meses (em experiências actuais normalmente 1-2 meses).
- Para RT-PCR lugar cada repetição (uma peça) já preparada em 2 ml tubos contendo duas esferas de aço inoxidável (2,5 mm diam) e três esferas de zircônio (2 mm diam) moagem.
- Congelar imediatamente (de preferência em azoto líquido) todas as amostras e mantê-las a -80 ° C até processamento.
5. Aflatoxina Análise Individual de Meio Cotyledon Pieces
- Use a A. flavus inoculadas amostras para esta análise. Trazer amostras à temperatura ambiente durante cerca de 30 min, adicionar quatro volumes (geralmente 2-3 ml; w / v) de metanol (manter registro para cálculos posteriores), feche as tampas, e incubar O / N (~ 16 horas) no escuro sem agitação.
- Coloque uma frita numa minicoluna combinando 1,5 mL de propileno, adicionar 200 mg de base Al 2 O 3 e cap com outro filtro poroso 36 como descrito anteriormente; em seguida, coloque um Ultra-High Performance Liquid Chomatographer (UPLC) frasco autosampler sob a coluna de perto o suficiente para evitar possível evaporação do eluato.
- Num tubo de ensaio de vidro descartável (não usar plástico), coloque 0,5 ml de extracto de metanol obtido no passo 4.1, adicionar 0,5 mL de acetonitrilo, mistura com uma pipeta e aplicar 0,5 ml da mistura para a coluna, preparada no passo 4.2. Permitir eluição no frasco amostrador automático por gravidade (não aplique pressão). Eluição normalmente leva 2-4 min, perto the frasco imediatamente usando um boné compatível UPLC com septos. Manter os frascos à temperatura ambiente e analisá-los no mesmo dia em um UPLC.
- Para a separação de aflatoxinas, frascos lugar amostrador automático de amostras contendo eluatos e frascos de auto-amostragem com padrões de aflatoxinas (B 1 e B 2 quando se utiliza A. flavus) em um instrumento UPLC equipado com uma correspondência Gerente UPLC Quaternário solvente, UPLC Sample Manager, UPLC fluorescente Detector, e um 2.1 mm x 50 mm 18 C, de 1,7? m.
- Usar uma fase móvel isocrática constituída por água / MeOH / CH3CN (64:23:13, v / v / v) de mistura a uma taxa de fluxo de 0,30 ml min -1. Obter cromatogramas que assegurem uma separação da linha de base estabilizada para o cálculo preciso da concentração de aflatoxina, de acordo com as instruções do fabricante 37.
- Confirmar a identidade de aflatoxinas através da obtenção de seus dados em massa espectral e comparando-os com os dados publicados 38. Use um ino espectrómetro de massa de armadilha equipado com uma interface de ESI e software correspondente de acordo com as instruções do fabricante.
- Determinar as concentrações de aflatoxinas por referência a curvas de calibração obtidas por injecção de diferentes quantidades de correspondentes padrões comerciais de aflatoxinas B 1, B 2, G 1 e G 2, tal como sugerido pelo fabricante e UPLC determinado pelo software 37.
- Coloque pedaços de sementes já extraída com metanol, em frascos de vidro individuais para O / N (~ 16 horas) liofilização para determinar o seu peso seco. Em seguida, calcular a concentração de aflatoxina em ng. G -1 de peso seco de parte da semente.
- Para a análise dos dados, converter resultados de aflatoxinas para login (ng. G -1 +1), seguido pelo teste de Tukey para comparação média.
6. Gene Expression, Processamento de RT-PCR de amostras
- Pegue os 2 ml de moagem tubos contêming amostras do freezer -80 ° C, e imediatamente (sem descongelar) triturá-los num homogeneizador moinho de esferas em 3100 rpm durante 40 segundos, em seguida, proceder à extracção de ARN usando Trizol de acordo com as instruções do fabricante.
- Preparar a partir de cada amostra de ADNc utilizando ARN de 1 ug e quantidades iguais de oligo dT e hexâmeros aleatórios, efectuar uma diluição de 1: 8 e o ADNc de usar 2 ul por reacção de RT-PCR (como descrito anteriormente 39).
- Para a detecção da expressão de ARNi usar INSERT iniciadores: RT_5X_1_105F: 5'GGTGGCATTGGACCGTCTTG-3 ', RT_5X_1_232R: 5'-CGCATCGAGGACAGGTTGTG-3'; e RT_5X_2_95F: 5'-CCATGTTTCTGGTGGCATTG-3 ', RT_5X_2_229R: 5'-ATCGAGGACAGGTTGTGTTG-3'.
- Para a detecção de expressão do marcador seleccionável NPTII, utilizar iniciadores: RT_NPTII_1_6871F: 5'-CTCGCTCGATGCGATGTTTC-3 ', RT_NPTII_1_7004R: 5'-GCAGGATCTCCTGTCATCTC-3'. Use o gene de manutenção actina para a normalização, e primers: actina-Fw: CACATGCCATCCTTCGATTG; Actina-Rv: CCAAGGCAACATATGCAAGCT 40.
- Analisar os resultados por Delta-delta método C T 41 padronizado para a expressão de actina. Representam os resultados como vezes de aumento sobre o controlo.
Resultados
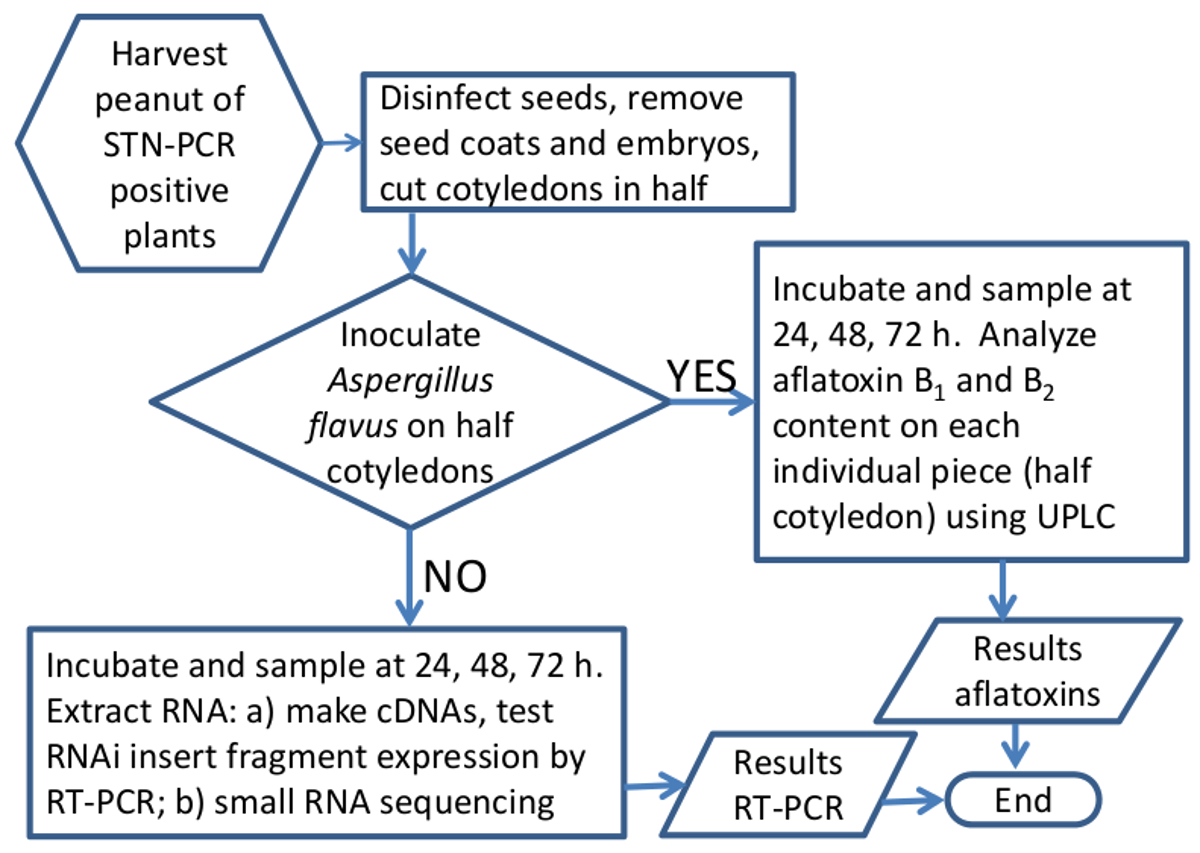
Plasmídeo p5XCAPD foi feito como um derivado do pCAPD 29, e é usado para transformar plantas de amendoim; este vector carrega repetições invertidas de cinco pequenos fragmentos, 70-80 pb cada, de genes aflatoxina-síntese de A. flavus separados por um intrão (Figura 1). Fragmentos de AFL2G_07224 (AFLR), AFL2G_07223 (AFLS ou aflJ), AFL2G_05027 (bomba de efluxo de aflatoxina, aflep), AFL2G_07228 (AFLC / pksA / pksL1), e AFL2G_07731 (PES1) foram utilizados para a construção, os números na Figura 1 corresponder a um . flavus genoma anotação no Broad Institute, Cambridge, MA, ea literatura 42. Um total de 99 linhas de amendoim foram regenerados depois de passar pelo processo de transformação, 50 foram PCR positivo para NPTII detectado pela STN-PCR, e 33 linhas foram PCR positivos e sementes produzidas. Apenas sete linhas positivas de PCR foram propagadas clonalmente e testado pela presenmétodo de t para a acumulação de aflatoxina, todos os sete apresentaram entre 60% e% menor acúmulo de aflatoxina 100 do que o controle. Aqui nós mostramos os resultados de duas destas sete linhas. Como os eventos transgênicos individuais geralmente produzem poucas sementes, foi desenvolvido um método para utilizar um número mínimo de sementes e ainda ser capaz de fazer análise estatística paramétrica. Um fluxograma da preparação da amostra e a configuração experimental é mostrado na Figura 5 e na Tabela 1. Embora a primeira geração de sementes transgénicas é tipicamente hemizigótico, espera-se que a célula-a-célula e circulação sistémica de ARN interferente pequeno (ARNsi) gerado através de interferência de RNA deve conferir aflatoxina-síntese silenciamento toda a planta.

Figura 5. fluxograma esquemático do método para analisar a eficácia dos ARNi em silenciamento Aspergillus genes aflatoxina-síntese em sementes de amendoim. Representação gráfica do fluxo de trabalho durante o processamento de amostras de amendoim para a expressão do gene ou a análise das aflatoxinas. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}
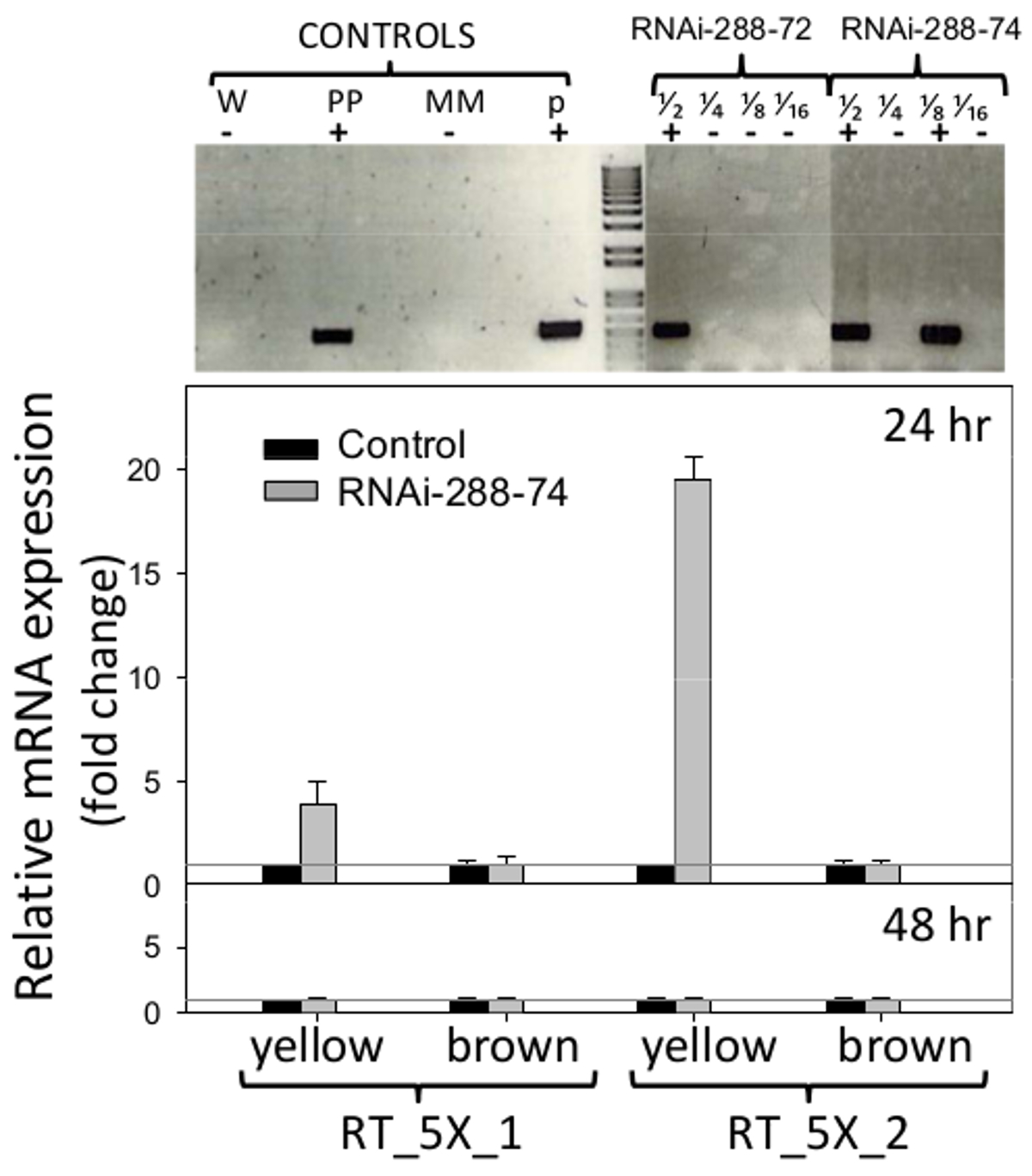
ARNi linhas 288-72 e 288-74 amendoim mostrou a presença do fabricante seleccionável NPTII quando testado pela STN-PCR, secções de gel são mostradas na Figura 6 (parte superior), as imagens originais estão disponíveis a partir dos autores, a pedido. O plasmídeo pCAPD não foi utilizada como um controlo de transformação, uma vez que codifica repetições invertidas de dois genes CMR (resistência ao cloranfenicol) e ccdB (toxina) de efeito desconhecido em plantas. PCR negativo 288-9 linha de amendoim e outros, que passaram pelo processo de regeneração e foi cultivada em condições idênticas às linhas de ARNi, foram usados como con negativatrolo.

. Figura 6. A detecção de transgênicos e de expressão Tempo real de RNAi inserção Top: Single-tubo, detecção de Nested PCR de linhas de amendoim transgênicas RNAi-288-72 e 288-74-RNAi, plantas positivos. Controles: W (água), PP (planta positivo), MM (mistura principal), p (plasmídeo p5XCAPD); As fracções 1/2 1/4 1/8 1/16 representam as diluições de 2 vezes do ADN de fundo:. Real-Time PCR detecção de expressão do ARNi inserir (conjuntos de iniciadores: RT_5X_1, RT_5X_2, como na Figura 1, em imatura ( amarelo) e maduro () cotilédones marrom de linhagens transgênicas em 24 e 48 h de incubação; linha cinza: C T = 1. Os histogramas representam os meios e barras de erro padrão (T) de três amostras biológicascom três repetições técnicas. A quantificação relativa de inserção RNAi foi normalizado em relação ao gene housekeeping actina como um controlo interno e de expressão dobra comparativa do transgene calculada conforme descrito no ponto 5.2.2 e 5.3. Por favor clique aqui para ver uma versão maior desta figura.
{kind=link}
Para testar a eficácia do controle potencial planta-hospedeiro RNAi mediada de acumulação de aflatoxina, conídios recém-colhida de Aspergillus flavus NRRL 3357 foram aplicadas na superfície de corte de cotilédones meio do qual os embriões e testa havia sido removido (Figuras 3, 4). A. flavus NRRL 3357, para o qual o genoma foi seqüenciado e foi a base para a concepção de p5XCAPD, foi gentilmente cedido pelo Dr. Horn em USDA-ARS-NPRL. A invasão de fungos resultante da metade cotilédone após 24, 48 e 72 horas a 30 ° C é mostrar N na Figura 4. As amostras de cotilédones meio inoculadas foram colhidas às 24, 48, 72, 96 hr de incubação, e analisada para o principal quatro aflatoxinas B 1, B 2, G 1 e G 2, utilizando UPLC e confirmado por LC-MS ; os resultados estão apresentados na Figura 6. As concentrações de aflatoxina foram determinados usando um método publicado com modificações 36. Em 96 horas de incubação, cotilédones começam a desintegrar-se devido à infecção fúngica. Linha de RNAi 288-72 apresentaram níveis significativamente mais baixos de aflatoxinas do que o controle em todas as datas de amostragem em cotilédones imaturos e na maioria das datas de amostragem nas mais maduras. Linha de RNAi 288-74 apresentaram níveis significativamente mais baixos de aflatoxinas na maioria das datas de amostragem. Os níveis de significância de teste de Tukey são indicados com asteriscos no gráfico da Figura 7.
g7.jpg "/>
. Figura 7. As aflatoxinas B 1 e B 2 em cotilédones meio de amendoim após incubação (24, 48, 72 e 96 h) com Aspergillus flavus Controle: 288-9 sementes de linha não-transgênico; RNAi: sementes de ARNi-288-72 e 288-74 RNAi, transgénicos para RNAi p5XCAPD para silenciar genes cinco aflatoxina-síntese. (A) A aflatoxina B 1 em sementes maduras (marrom); (B) A aflatoxina B 2 sementes maduras; (C) A aflatoxina B 1 em sementes imaturas (amarelo) e (D) Aflatoxina B 2 em sementes imaturas. Os valores médios com as correspondentes barras de erro padrão (T) de amostras biológicas duplicados são representados. Diferenças estatisticamente significativas de Tukey teste *: p ≤ 0,05, **: p ≤ 0,01, ***: p ≤ 0,001.s: //www.jove.com/files/ftp_upload/53398/53398fig7large.jpg "target =" _ blank "> Clique aqui para ver uma versão maior desta figura.
Em geral, RNAi-288-72 mostrou ao longo da experiência (24 horas de incubação a 96), uma redução de 94% -100% em aflatoxina B 2, e uma redução de 90% -100% em aflatoxina B 1 em comparação com o controlo. ARNi-288-74 mostrou uma redução de 63% -100% em aflatoxina B 2 e 60% -100% de redução em aflatoxina B 1, Figura 7.
Os iniciadores utilizados para a detecção de PCR em tempo real de expressão do inserto de RNAi são mostrados na Figura 1. Cotilédones amendoim-semente foram analisadas sem embriões, para remover as defesas naturais e ser capaz de detectar o efeito potencial de ARNi na área mais exposta ao invasão de fungos, os cotilédones. Expressão do ARNi inserção detectada em cotilédones imaturos (amarelo) de linha pelo conjunto de iniciadores 288-74 RT_5X_1 foi de quatro vezes em relação ao C T 1 = limiar do controlo negativo, e pelo conjunto de iniciadores foi RT_5X_2 19 vezes acima do limiar, todos em 24 horas de incubação. Pelo menos três dos cinco fragmentos de genes consecutivos utilizados na transformação de amendoim, 5027, 7223 e 7228 (aflep, AFLS / aflJ, e AFLC / pksA, respectivamente) foram detectados por RT-PCR (Figura 1, 6). Expressão de inserção RNAi não foi detectada em cotilédones maduros às 24 horas, ou no cotilédones maduras ou imaturas em 48 horas de incubação, a Figura 6 (parte inferior).
1| Linha de amendoim | Tempo de amostragem | As amostras para RT-PCR | As amostras para análise de aflatoxinas (inoculado) | Number de sementes | ||||
| (não inoculado) | ||||||||
| Rep 1 | Rep 2 | Rep 3 | Rep 1 | Rep 2 | Rep 3 | |||
| RNAi (amarelo) | 24 hr | 1 | 1 | 1 | 1 | 1 | 1 | 4,5 |
| 48 hr | 1 | 1 | 1 | 1 | 1 | |||
| 72 hr | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Ao controle | 24 hr | 1 | 1 | 1 | 1 | 1 | 1 | 4,5 |
| (amarelo) | 48 hr | 1 | 1 | 1 | 1 | 1 | 1 | |
| 72 hr | 1 | 1 | 1 | 1 | 1 | 1 | ||
Tabela 2. Exemplo de configuração pequena amostra para analisar a expressão gênica e acumulação de aflatoxina em RNAi amendoim sementes. Análise completa de expressão e de aflatoxinas para um grupo de maturação (ie., Amarelo), com três tempos de amostragem (24, 48 e 72 horas) , em triplicado, exigiria 4,5 sementes, cada número um na tabela representa metade cotilédones.
Discussão
Planta-hospedeiro RNAi mediada por silenciamento de genes em fungos patogénicos foi demonstrada 27,43, no entanto, não existem publicações que mostram a viabilidade do controlo de RNAi mediada por acumulação de micotoxinas em plantas. Um factor limitante para estes estudos em amendoim foi a falta de um método para avaliar um não-aflatoxina fenótipo acumulação em plantas individuais, como as folhas não apresentam sintomas após infecção fúngica das vagens subterrâneas. Além disso, o acúmulo de não-normalmente distribuídos de aflatoxinas, ea necessidade de grandes amostras para análise química 15,16 dificultaram a quantificação do potencial efeito RNAi em uma única planta. O método aqui apresentado consiste em 72 experiências de RH utilizando cinco sementes para executar três amostragens de 24 horas de intervalo, em triplicado (Tabela 1, Figura 7). Em comparação com a análise típica aflatoxina que requer não menos do que 100 g de sementes, o nosso método é particularmente adequado para individuaL eventos transgénicas de plantas de amendoim que inicialmente não produzem mais do que dois ou três vagens.
Silenciamento mediado ARN-síntese de aflatoxina foi demonstrada por transformar geneticamente Aspergillus flavus e A. parasiticus. Desde AFLR é um regulador principal de produção de aflatoxinas em A. flavus e A. parasiticus 44,45, torna-se um alvo interessante para o silenciamento mediado ARN em plantas. No entanto, variações genéticas nos AFLR foram mostrados entre espécies Aspergillus 46, e essas variantes genéticas poderiam escapar silenciamento se não existe uma sequência perfeita correspondência com o sinal de ARNi produzido no hospedeiro planta. Assim, AFLR foi um dos alvos para silenciar no vetor p5XCAPD, mas não foi o único. Repetições invertidas do gene AFLR introduzidos A. flavus e A. parasiticus por transformação resultou em silenciar e mínimo ou nenhum produtoion de aflatoxinas 47 (McDonald et al., 2005b). Além disso, o silenciamento do gene Afld impedido a produção de aflatoxina em até 98% em A. flavus e A. parasiticus em transformação direta 48. Para aumentar a probabilidade de sucesso no nosso sistema, de amendoim foi transformada com fragmentos de repetição invertida de cinco genes envolvidos na produção de aflatoxinas em A. flavus. Aqui mostra-se que o uso de p5XCAPD que tem como alvo para silenciar vários genes na via de síntese de aflatoxina, 90% -100% de níveis mais baixos de aflatoxina B 1 e B 2 foram obtidos de acordo 288-72, e níveis mais baixos de 60-100% em acumulada A linha 288-74 em comparação com o controlo, quando cotilédones meio foram inoculados com A. flavus, as Figuras 4, 7. Mais importante ainda, este método detectou diferenças estatisticamente significativas na acumulação de aflatoxina por linhas 288-72, 288-74 contra o controle durante o experimento, aplicando statis paramétricotiques, a Figura 7. Dado o pequeno tamanho da amostra, é importante destacar a necessidade de usar um método poderoso para detectar aflatoxinas, estas experiências foram analisadas por UPLC que tem uma alta resolução, cinco vezes maior desempenho e três vezes maior sensibilidade do que HPLC 49.
Expressão da RNAi inserção em 288-74 só foi detectada em cotilédones imaturos (amarelo) em 24 horas de incubação. A inserção RNAi não foi detectado por RT-PCR em cotilédones maduros de 288-74 às 24 horas, ou em qualquer grupo de maturação em 48 horas, a Figura 6. Este mesmo fenômeno foi observado em outras linhas de amendoim transgênicas de RNAi (Arias, RS, 2015 não publicado), onde normalmente transcritos de ARNi foram detectadas apenas em cotilédones imaturos às 24 h. As amostras de RNA foram tratadas com DNase antes da síntese de ADNc, os dados foram normalizados para o nível de expressão de actina e nenhuma evidência de contaminação do ADN foi observada. No caso de o ADN ter estado presente nas amostras, que deveriaforam detectados nas amostras de 48 h, bem como, de forma consistente, mas que não foi o caso. A expressão sob o controlo do promotor 35S-nem sempre é uniforme; ele pode ser afetada pelas condições ambientais 50, tipo de tecido e estágio de desenvolvimento 51,52. Ao mesmo tempo, na via de interferência de ARN, a taxa de decaimento do mRNA e a taxa de decaimento siARN pode variar significativamente 53. É possível que a rápida degradação do ARNm por o mecanismo de interferência de ARN poderia ter evitado a detecção de ARNm de 48 horas de incubação. Se a ausência de expressão em 48 horas foi devido a baixa 35S-promotor impulsionado transcrição, ou a degradação rápida de dsRNA por Dicer continua a ser respondida. Assim, a detecção de pequenos RNAs de alto rendimento de sequenciamento daria uma melhor visão sobre os processos que ocorrem por meio de RNAi 54 Nestas experiências. No entanto, desde RNA silenciamento espalha sistemicamente, principalmente através do floema de photosyntodeio fontes à sacarose pias (neste caso, amendoim sementes) 55, o silenciamento de aflatoxina-síntese pode ocorrer em sementes sem expressão local do RNAi inserção. Muita pesquisa ainda precisa ser feito para determinar o nível de limiar de pequenos RNAs de interferência (siRNAs) necessários para evitar a acumulação de aflatoxina em sementes. É importante ressaltar o fato de que ambos, a expressão do mRNA do RNAi construir (Figura 6), ea acumulação de aflatoxinas B 1 e B 2 (Figura 7) apresentaram resultados diferentes para imaturo (amarelo) vs. madura (marrom) cotilédones. Plantas de amendoim têm crescimento indeterminado, isto é, eles apresentam no momento da colheita de uma gama de vagens de maturidade, Figura 2. Além disso, as sementes a partir de diferentes grupos de maturação diferem na sua composição química, por ex., 2,4% de sacarose nas sementes imaturas, e 1,9% em sementes maduras, nas mesmas condições de campo 56,57. Assim, para entender a efic realy de controlo mediada por ARN-acumulação de aflatoxina, é importante analisar separadamente grupos de maturação.
A defesa natural de sementes de amendoim é a produção de fitoalexinas, que varia na diversidade de compostos produzidos e suas quantidades relativas, dependendo da maturidade das sementes e das condições ambientais 58-61, e é particularmente maior em embriões em comparação com 62 cotilédones. Os embriões também têm concentrações significativamente maiores de ácidos nucleicos, tanto DNA e RNA do que os cotilédones (RS Arias, não publicados). Como sementes de amendoim amadurecer, mudanças em sua fisiologia e composição química ocorrer 63. Antioxidantes fenólicos em forma de amendoim testa taninos condensados com atividade fungistática 64; este é também evidente na cor mesocarpo que reflete estágios de maturidade, amarelo para preto 35, como o seu teor de taninos e compostos fenólicos aumenta com a maturidade de 65. Assim, a presença de tESTA ou embriões no experimento, dadas as suas propriedades antimicrobianas, pode ter limitado o crescimento de fungos e, portanto, sobrestimada o efeito de silenciamento de RNAi, por conseguinte, eles foram removidos. Além disso, a remoção dos embriões Testa e ajuda a limitar as fontes de variação na análise, como a metade cotilédone que transporta o embrião terá mais fitoalexinas e mais conteúdo de RNA.
Em adição à análise por grupos de maturação e remoção do tegumento e embrião nestas experiências, é importante salientar mais algumas observações: a) embora os resultados são mostrados por até 96 horas de incubação, é recomendado o uso de não mais do que 72 hr para obter resultados consistentes, como sementes se degradado por 96 horas; e b) Considerando cotilédones meia da mesma semente, embora selecionadas aleatoriamente, não constituem amostras perfeitamente independentes, RT-PCR e acumulação de aflatoxina dentro de eventos transgênicos mostrou pouca variação entre as sementes. Além disso, contar um esporo do fungo precisa, volume de inóculoS de 2 mL, e a aplicação de esporos sobre a superfície de corte dos cotilédones evitando gotejamento sobre os lados são importantes para assegurar que os esporos germinados são expostos para o tecido da planta. A água / agar nas placas deve ser de 1,5% (w / v), mais macio faz com que o escoamento de agar de esporos, como mostrado no último quadro da Figura 4 (em baixo). Deve disponibilidade semente transgénica a partir de um evento especial, ser limitado, a amostragem pode ser feita em duplicado em triplicado, em vez de se obterem resultados semelhantes (ou seja, Figura 7); no entanto, as amostras em triplicado vai ajudar a reduzir o erro padrão. A única limitação deste método é que ele exige um sistema altamente sensível (UPLC) para a detecção de aflatoxina / quantificação, mas, ao mesmo tempo que reduz a probabilidade de sobrestimar o efeito de RNAi deve aflatoxinas não ser detectado por meio de métodos menos sensíveis.
Em conclusão, este método oferece pela primeira vez um método fiável para estudar oefeito de RNAi no controlo das aflatoxinas. Reduzindo o tempo para uma experiência de uma safra inteira para menos de uma semana, este método irá tremendamente acelerar a investigação sobre RNAi-amendoim / Aspergillus patossistema para a mitigação e / ou eliminação de aflatoxinas.
Divulgações
The authors have nothing to disclose or any conflicts of interest.
Agradecimentos
This work received the financial support of USDA-ARS CRIS project 6604-21000-004-00D, CRIS project 6604-42000-008-00D, and USAID Feed-the-Future program Agreement number 58-0210-3-012. We thank Valerie Orner, LaTanya Johnson, Joseph Powell and Kathy Gray for their technical assistance. Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the US Department of Agriculture.
Materiais
| Name | Company | Catalog Number | Comments |
| Primers, oligonucleotides | DNA Technologies, Coralville, IA, USA | n/a | |
| Dneasy Plant Mini Kit | Qiagen, Valencia, CA | 69106 | |
| Czapek Dox agar medium | Oxoid, by Thermo Fisher Scientific, Waltham, MA | CM0095 | |
| Agar | Thermo Fisher Scientific, Waltham, MA | BP 1423 | |
| Freezer -80 °C | n/a | n/a | |
| Aluminum Oxide, Al2O3 | Fisher Scientific | A941 | |
| SPE Reservoirs 1.5 ml | Grace Davison Discovery Scientific | 210011 | |
| Frits for 1.5 ml SPE reservoir | Grace Davison Discovery Scientific | 211401 | |
| Autosampler vials | Waters Corporation, Milford, MA | 186005221 | |
| Waters Acquity Ultra-Performance Liquid-Chromatography (UPLC) instrument; UPLC-H-Class Quaternary Solvent Manager; UPLC Sample Manager; UPLC Fluorescent detector (FLR); UPLC BEH C18 2.1 mm x 50 mm, 1.7 mm column | Waters Corporation, Milford, MA | ||
| Finnigan LCQ Advantage MAX ion trap mass spectrometer, with Xcalibur version 1.4 software | Thermo Electron Corp., San Jose, CA | ||
| Aflatoxin standards, B1, B2, G1 and G2 | Sigma-Aldrich, St. Louis, MO | A6636; A9887; A0138; A0263 | |
| Systat Software 12.2 | SYSTAT Software Inc., Point Richmond, CA | ||
| Trizol reagent | Invitrogen, CA | 15596-018 | |
| SuperScript III First Strand Synthesis Super Mix | Invitrogen, CA | 11752-050 | |
| ABI 7500 Real-Time PCR | Lifetechnologies, Grand Island, NY | 4406984 | |
| Luria Broth-Miller | Fisher Scientific | R453642 | |
| pENTR1A | Invitrogen, CA | A10462 | |
| LR Clonase II enzyme mix | Invitrogen, CA | 11791-020 | |
| T4 DNA Ligase | NEB Biolabs | M0202L | |
| Gelrite | Sigma-Aldrich, St. Louis, MO | G1919 | |
| Acetosyringone | Sigma-Aldrich, St. Louis, MO | D134406 | |
| QIAcube robot workstation | Qiagen, Valencia, CA | 9001292 | |
| Antibiotics: kanamycin, cefotaxime, gentamicin; streptomycin | Goldbio, St. Louis, MO | cef.: C-104-25; kan: K-120-5; gent.: G-400-1; strep.: S-150-50 | |
| Platinum Taq DNA Polymerase High Fidelity | Invitrogen, CA | 11304-029 |
Referências
- Williams, J. H., et al. Human aflatoxicosis in developing countries: a review of toxicology, exposure, potential health consequences, and interventions. The American Journal of Clinical Nutrition. 80, 1106-1122 (2004).
- American Association for Cancer Research: AACR. An evaluation of chemicals and industrial processes associated with cancer in humans based on human and animal data: IARC Monographs Volumes 1 to 20. Cancer Research. 40, 1-12 (1980).
- Turner, P. C. The molecular epidemiology of chronic aflatoxin driven impaired child growth. Scientifica. , (2013).
- Rasooly, R., Hernlem, B., He, X., Friedman, M. Non-linear relationships between aflatoxin B1 levels and the biological response of monkey kidney vero cells. Toxins (Basel). 5, 1447-1461 (2013).
- Gong, Y. Y., et al. Determinants of aflatoxin exposure in young children from Benin and Togo, West Africa: the critical role of weaning. International Journal of Epidemiology. 32, 556-562 (2003).
- Eaton, D. L., Groopman, J. D. The toxicology of aflatoxins: human health, veterinary, and agricultural significance. , Academic Press. (1994).
- Murugavel, K. G., et al. Prevalence of aflatoxin B1 in liver biopsies of proven hepatocellular carcinoma in India determined by an in-house immunoperoxidase test. Journal of Medical Microbiology. 56, 1455-1459 (2007).
- Wang, J. S., et al. Hepatocellular carcinoma and aflatoxin exposure in Zhuqing Village, Fusui County, People's Republic of China. Cancer Epidemiology, Biomarkers & Prevention. 10, American Association for Cancer Research. 143-146 (2001).
- Azziz-Baumgartner, E., et al. Case-control study of an acute aflatoxicosis outbreak, Kenya, 2004. Environmental Health Perspectives. 113, 1779-1783 (2005).
- Lye, M. S., Ghazali, A. A., Mohan, J., Alwin, N., Nair, R. C. An outbreak of acute hepatic encephalopathy due to severe aflatoxicosis in Malaysia. American Journal of Tropical Medicine and Hygiene. 53, 68-72 (1995).
- Villers, P. Aflatoxins and safe storage. Frontiers in Microbiology. 5, 158(2014).
- Kensler, T. W., Roebuck, B. D., Wogan, G. N., Groopman, J. D. Aflatoxin: A 50-year odyssey of mechanistic and translational toxicology. Toxicological Sciences. 120, S28-S48 (2011).
- Dorner, J. W., Cole, R. J., Wicklow, D. T. Aflatoxin reduction in corn through field application of competitive fungi. Journal of Food Protection. 62, 650-656 (1999).
- Cotty, P. J., Bhatnagar, D. Variability among atoxigenic Aspergillus flavus strains in ability to prevent aflatoxin contamination and production of aflatoxin biosynthetic-pathway enzymes. Applied and Environmental Microbiology. 60, 2248-2251 (1994).
- Whitaker, T. B. Standardisation of mycotoxin sampling procedures: an urgent necessity. Food Control. 14, 233-237 (2003).
- Whitaker, T. B., Dorner, J. W., Giesbrecht, F. G., Slate, A. B. Variability among aflatoxin test results on runner peanuts harvested from small field plots. Peanut Science. 31, 59-63 (2004).
- Fire, A., et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 391, 806-811 (1998).
- Rafael, D., et al. EMT blockage strategies: Targeting Akt dependent mechanisms for breast cancer metastatic behaviour modulation. Current Gene Therapy. , (2015).
- Li, G., Chang, H., Zhai, Y. P., Xu, W. Targeted silencing of inhibitors of apoptosis proteins with siRNAs: a potential anti-cancer strategy for hepatocellular carcinoma. Asian Pacific. Journal of Cancer Prevention: APJCP. 14, 4943-4952 (2013).
- Koldehoff, M. Targeting bcr-abl transcripts with siRNAs in an imatinib-resistant chronic myeloid leukemia patient: challenges and future directions. Methods in Molecular Biology. 1218, 277-292 (2015).
- Zhang, J., et al. Pest control. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science. 347, 991-994 (2015).
- Ajjappala, H., Chung, H. Y., Sim, J. S., Choi, I., Hahn, B. S. Disruption of prefoldin-2 protein synthesis in root-knot nematodes via host-mediated gene silencing efficiently reduces nematode numbers and thus protects plants. Planta. 241, 773-787 (2015).
- Jose, A. M., Hunter, C. P. Transport of sequence-specific RNA interference information between cells. Annual Review of Genetics. 41, 305-330 (2007).
- Vazquez, F., Hohn, T. Biogenesis and biological activity of secondary siRNAs in plants. Scientifica. , Hindawi Publishing Corporation. (2013).
- Tinoco, M. L. P., Dias, B. B. A., Dall'Astta, R. C., Pamphile, J. A., Aragao, F. J. L. In vivo trans-specific gene silencing in fungal cells by in planta expression of a double-stranded RNA. BMC Biology. 8, (2010).
- Govindarajulu, M., Epstein, L., Wroblewski, T., Michelmore, R. W. Host-induced gene silencing inhibits the biotrophic pathogen causing downy mildew of lettuce. Plant Biotechnology Journal. , (2014).
- Yin, C., Jurgenson, J. E., Hulbert, S. H. Development of a host-induced RNAi system in the wheat stripe rust fungus Puccinia striiformis f. sp. tritici. Molecular Plant-Microbe Interactions. 24, 554-561 (2011).
- Ghag, S. B., Shekhawat, U. K., Ganapathi, T. R. Host-induced post-transcriptional hairpin RNA-mediated gene silencing of vital fungal genes confers efficient resistance against Fusarium. wilt in banana. Plant Biotechnology Journal. 12, 541-553 (2014).
- Filichkin, S. A., et al. Efficiency of gene silencing repeats vs. transitive RNAi in Arabidopsis: direct inverted vectors. Plant Biotechnology Journal. 5, 615-626 (2007).
- Sciaky, D., Montoya, A. L., Chilton, M. D. Fingerprints of Agrobacterium Ti Plasmids. Plasmid. 1, 238-253 (1978).
- Clark, D. J., Maaloe, O. DNA Replication and Division Cycle in Escherichia coli. Journal of Molecular Biology. 23, 99-112 (1967).
- Murashige, T., Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plantarum. 15, 473-497 (1962).
- Srinivasan, T., Kumar, K. R. R., Kirti, P. B. Establishment of efficient and rapid regeneration system for some diploid wild species of Arachis. Plant Cell Tissue and Organ Culture. 101, 303-309 (2010).
- Gomes, A. L. V., et al. Single-tube nested PCR using immobilized internal primers for the identification of dengue virus serotypes. Journal of Virology Methods. 145, 76-79 (2007).
- Williams, E. J., Drexler, J. S. A non-destructive method for determining peanut pod maturity. Peanut Science. 8, 134-141 (1981).
- Sobolev, V. S., Dorner, J. W. Cleanup procedure for determination of aflatoxins in major agricultural commodities by liquid chromatography. Journal of AOAC International. 85, 642-645 (2002).
- Empower Software, Getting Started Guide. , Waters Corporation. Milford, MA. Available from: http://sites.chem.colostate.edu/diverdi/C431/experiments/high%20pressure%20liquid%20chromatography/references/Empower%20getting%20started%2071500031203rA.pdf (2002).
- Biselli, S., Hartig, L., Wegner, H., Hummert, C. Analysis of Fusarium. toxins using LC-MS-MS: Application to various food and feed matrices. LC GC North America. 23, 404-413 (2005).
- Arias, R. S., Sobolev, V. S., Orner, V. A., Dang, P. M., Lamb, M. C. Potential involvement of Aspergillus flavus laccases in peanut invasion at low water potential. Plant Pathology. 63, 353-363 (2014).
- Dang, P. M., Chen, C. Y., Holbrook, C. C. Evaluation of five peanut (Arachis hypogaea) genotypes to identify drought responsive mechanisms utilising candidate-gene approach. Functional Plant Biology. 40, 1323-1333 (2013).
- Schmittgen, T. D., Livak, K. J. Analyzing real-time PCR data by the comparative C-T method. Nature Protocols. 3, 1101-1108 (2008).
- Amaike, S., Keller, N. P. Aspergillus flavus. Annual Review of Phytopathology. 49, 107-133 (2011).
- Nowara, D., et al. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell. 22, 3130-3141 (2010).
- Woloshuk, C. P., et al. Molecular characterization of aflR, a regulatory locus for aflatoxin biosynthesis. Applied and Environmental Microbiology. 60, 2408-2414 (1994).
- Price, M. S., et al. The aflatoxin pathway regulator AflR induces gene transcription inside and outside of the aflatoxin biosynthetic cluster. FEMS Microbiology Letters. 255, 275-279 (2006).
- Ehrlich, K. C., Montalbano, B. G., Cotty, P. J. Sequence comparison of aflR from different Aspergillus. species provides evidence for variability in regulation of aflatoxin production. Fungal Genetics and Biology. 38, 63-74 (2003).
- McDonald, T., Brown, D., Keller, N. P., Hammond, T. M. RNA silencing of mycotoxin production in Aspergillus and Fusarium species. Molecular Plant Microbe Interactions. 18, 539-545 (2005).
- Abdel-Hadi, A. M., Caley, D. P., Carter, D. R., Magan, N. Control of aflatoxin production of Aspergillus flavus. and Aspergillus parasiticus. using RNA silencing technology by targeting aflD. (nor-1) gene. Toxins (Basel). 3, 647-659 (2011).
- Swartz, M. E. Ultra performance liquid chromatography (UPLC): An introduction: Separation Science Redefined. LCGC North America. , Suppl ement 8. 8-14 (2005).
- Maghuly, F., Khan, M. A., Fernandez, E. B., Druart, P., Watillon, B., Laimer, M. Stress regulated expression of the GUS-marker gene (uidA) under the control of plant calmodulin and viral 35S promoters in a model fruit tree rootstock: Prunus incisa x serrula. Journal of Biotechnology. 135, 105-116 (2008).
- de Mesa, M. C., Santiago-Doménech, N., Pliego-Alfaro, F., Quesada, M. A., Mercado, J. A. The CaMV 35S promoter is highly active on floral organs and pollen of transgenic strawberry plants. Plant Cell Reports. 23, 32-38 (2004).
- Sunilkumar, G., Mohr, L., Lopata-Finch, E., Emani, C., Rathore, K. S. Developmental and tissue-specific expression of CaMV 35S promoter in cotton as revealed by GFP. Plant Molecular Biology. 50, 463-474 (2002).
- Groenenboom, M. A. C., Maree, A. F. M., Hogeweg, P. The RNA silencing pathway: The bits and pieces that matter. PLoS Computational Biology. 1, 155-165 (2005).
- Zhao, D., Song, G. Q. High-throughput sequencing as an effective approach in profiling small RNAs derived from a hairpin RNA expression vector in woody plants. Plant Science: an International Journal of Experimental Plant Biology. 228, 39-47 (2014).
- Kamthan, A., Chauduri, A., Kamthan, M., Datta, A. Small RNAs in plants: recent development and application for crop improvement. Frontiers in Plant Science. 6, 208(2015).
- Manda, A., Bodapati, P. N., Rachaputi, N. C., Wright, G., Fukai, S. Aflatoxins and their relationship with sugars in peanut (Arachis hypogaea L). 4th International Crop Science Congress, 2004, , Available from: http://www.cropscience.org.au/icsc2004/poster/5/1/3/625_manda.htm (2004).
- Uppala, S. S. Factors affecting pre-harvest aflatoxin contamination of peanut (Arachis hypogaea L). , Auburn University. (2011).
- Sobolev, V. S. Localized production of phytoalexins by peanut (Arachis hypogaea) kernels in response to invasion by Aspergillus species. Journal of Agricultural and Food Chemistry. 56, 1949-1954 (2008).
- Sobolev, V. S., Guo, B. Z., Holbrook, C. C., Lynch, R. E. Interrelationship of phytoalexin production and disease resistance in selected peanut genotypes. Journal of Agricultural and Food Chemistry. 55, 2195-2200 (2007).
- Sobolev, V. S., Neff, S. A., Gloer, J. B. New stilbenoids from peanut (Arachis hypogaea) seeds challenged by an Aspergillus caelatus strain. Journal of Agricultural and Food Chemistry. 57, 62-68 (2009).
- Dorner, J. W., Cole, R. J., Sanders, T. H., Blankenship, P. D. Interrelationship of kernel water activity, soil temperature, maturity, and phytoalexin production in preharvest aflatoxin contamination of drought-stressed peanuts. Mycopathologia. 105, 117-128 (1989).
- Sobolev, V. S. Production of phytoalexins in peanut (Arachis hypogaea) seed elicited by selected microorganisms. Journal of Agricultural and Food Chemistry. 61, 1850-1858 (2013).
- Basha, S. M. M., Cherry, J. P., Young, C. T. Changes in free amino acids, carbohydrates, and proteins of maturing seeds from various peanut (Arachis hypogaea L.) cultivars. Cereal Chemistry. 53, 586-596 (1976).
- Lansden, J. A. Aflatoxin inhibition and fungistasis by peanut tannins. Peanut Science. 9, 17-20 (1982).
- Yen, G. C., Duh, P. D., Tsai, C. L. Relationships between antioxidant activity and maturity of peanut hulls. Journal of Agricultural and Food Chemistry. 41, 67-70 (1993).
Reimpressões e Permissões
Solicitar permissão para reutilizar o texto ou figuras deste artigo JoVE
Solicitar PermissãoThis article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. Todos os direitos reservados