
Method Article
ピーナッツにおけるアフラトキシンのRNAi仲介制御:メソッドは、ピーナッツにマイコトキシン生産と導入遺伝子発現を分析します/
要約
我々は、真菌アスペルギルス・ フラバスでアフラトキシン合成遺伝子をサイレンシングするためのRNA干渉信号を含むピーナッツ種子でアフラトキシン及び導入遺伝子発現の分析のための方法を実証します。植物におけるマイコトキシンのRNAi媒介性制御は、以前に報告されていません。
要約
国連食糧農業機関は、世界の食糧作物の25%がアフラトキシンに汚染されていると推定しています。それは毎年の非ヒトの消費に破壊されたり流用されている食品の億トンを表します。アフラトキシンは、通常、穀類、ナッツ、根菜類や他の農産物におけるカビアスペルギルス・ フラバス と A. parasiticusにより蓄積された強力な発癌物質です。ピーナッツ植物におけるRNA干渉(RNAi)による5アフラトキシン合成遺伝子のサイレンシングは 、Aを接種した後アフラトキシンの蓄積を制御するために使用されましたフラバス 。以前は、何の方法は、これらは通常、いくつかの種子を生産するように、個々のピーナッツトランスジェニック事象においてRNAiの有効性を分析するために存在しなかった、とアフラトキシン導く条件下で大規模なフィールド実験の伝統的な方法は、オプションではありませんでした。フィールドに、自然に汚染種を発見する確率は1/1にしばしば1/100また000は、アフラトキシン汚染が均一に分布されていません。我々の方法は、リアルタイムPCR(RT-PCR)のために処理小片または小RNAシークエンシングを用いて、トランスジェニック事象当たり数種を使用し、超高性能液体クロマトグラフィー(UPLC)によってアフラトキシン蓄積の分析のために。 RNAiの発現ピーナッツライン288から72と288から74には、14,000 NGまで蓄積し、対照と比較してアフラトキシンB 1が100%削減(p≤0.01)とB 2に現れた。グラム-1アフラトキシンB 1のアフラトキシン生産A.を接種フラバス 。参考までに、米国では、人間の消費の許容アフラトキシンの最大合計は、20 ngのです。G -1。このプロトコルは、トランスジェニックピーナッツの種子とその評価方法におけるアフラトキシンのRNAi仲介制御の適用を説明しています。私たちは、ピーナッツや他の作物の育種への応用は、科学のこの重要な分野で急速な進歩をもたらすと考えています、医学、ヒトの栄養は、大幅主要食用作物でアフラトキシン、および潜在的に他のマイコトキシンを制御するために、国際的な取り組みに貢献していきます。
概要
約4.5億人が慢性的にアフラトキシン1、自然2で知られている最も強力な発がん物質にさらされています。これらのマイコトキシンは、トウモロコシ、キャッサバ、米、ナッツ、穀物やスパイスなど世界3に食用作物の25%を汚染する。4。子供5で発育阻害原因アフラトキシン、免疫系6を損ない 、人間の生検7,8における肝細胞癌-の58%に存在し、アフラトキシン中毒症9,10の定期的な大流行時に数百人を殺します。アフラトキシンは、通常、 アスペルギルス・ フラバス と A.によって生成ポリケチド由来のマイコトキシンですparasiticus;アフラトキシンB 1及びB 2は、A によって製造されていますA.一方フラバス 、 parasiticusも 、G 1およびG 2が生成されます。これらの化合物の化学構造およびUPLCによりそれらの分離を示すクロマトグラムを図1に示します。

図1. アフラトキシンおよびRNAi トップを 挿入します。化学構造(左)とクロマトグラムの例(右)4つの最も一般的なポリケチド由来のアフラトキシンの: アスペルギルス parasiticusにより産生され、B 1、B 2、G 1、G 2を 、 。。 フラバスは、B 1およびB 2が生成 下:RNAiの中の遺伝子断片の概略はp5XCAPDはピーナッツ変換に使用される構築、矢印の下の数字は、 アスペルギルス・フラバス のゲノム中の遺伝子断片アクセッション番号です。 PIV2:ジャガイモのイントロン。 BP:塩基対; RT_5X_1とRT_5X_2:リアルタイムPCRのプライマー部位は、 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
4 NGに基づいて算出した場合ピーナッツ中のアフラトキシンによる輸出の経済的損失だけでは$ 450万ドルを超えています。欧州連合(EU)11で人間の消費のために許可されたアフラトキシンのグラム-1限界。アフラトキシンは、60年12知られています。しかし、多くの農業は他の真菌株13,14の適用を含め、それらの影響を軽減するために開発されたものの、制御の一貫した方法が存在せず、耐性植物品種は使用できません。でも、病原体の侵入のための助けとなる条件下で、マイコトキシン蓄積が予測不可能であり、正規分布に従わないため、アフラトキシンに対する抵抗性試験植物遺伝資源は、特に困難です。このように、実験は通常、種子や100-1,700グラムの複数のサンプルの何百ものデータ15,16のばらつきを低減するために、大規模な植林面積を必要とします。
RNA干渉がありました1998年17で発見されました。そして、「サイレンシング」の利点は、現在、新しいアプリケーションの数で 、例えば、探索されている。、転移性乳癌18、肝臓癌19、骨髄性白血病20、および昆虫21と線虫22に対して植物保護におけるに対するヒトの治療に。植物では、RNA干渉信号も植物宿主25に密着している真菌病原体の内部に、23,24サイレンシング全身転写後遺伝子を担当する低分子干渉RNA(siRNA)および高分子量RNAを、細胞に細胞を移動することができます。レタス26における真菌病原体の遺伝子の植物媒介サイレンシングのRNAiの有効性植物の地上部における症状のこれらの、目視検査(葉)は、疾患の定量化を可能にするために、いくつかの植物pathosystemsに記載されている、 すなわち 、卵菌類Bremia小麦で、 プクキニア 28のp> 27とフザリウム 。はるかに難しいが、葉は、(種子)侵略器官は土壌の数インチ下にある感染症の症状を示さないようピーナッツの工場でマイコトキシン、特にアフラトキシンを制御するためのRNAi効果を評価することであり、感染症の発生は予測できない、唯一の化学物質であります分析では、アフラトキシンの存在を決定することができます。また、ピーナッツの各トランスジェニックイベントは、通常、数種子(植物当たり4-6)を生成し;そのため、大規模なフィールドプロットで無アフラトキシン蓄積特性、全体の作付季節を持続し、種子の数百を使用するための伝統的なテストは現実的ではありません。この方法は、わずか数種を使用して、一週間以内で、RNAiのピーナッツ種子導入遺伝子の存在について、および無アフラトキシン蓄積形質について分析するために、ここで説明されています。
プロトコル
1.分子構築物およびピーナッツ変換
- 5 Aの DNA断片を結合フラバス遺伝子 、AFL2G_07223(AFLSまたはaflJ)、AFL2G_07224(aflR)、AFL2G_07228(AFLC / pksA / pksL1)、AFL2G_07731(PES1)とAFL2G_05027(アフラトキシン排出ポンプ、aflep)。このために、以下のプライマーおよびultramersを使用:DIR-1、短期Dir1内-R、DIR-2反転、ショートDIR2-R、DirAll-NCO-RV、およびDirAll-BamEco-FW、 表1。
- メーカーのに従って、DNAポリメラーゼを用いて、(94℃45秒の5サイクル、55℃30秒、68℃15秒、続いて95℃で2分25μlの反応)5回のPCRサイクルによってDIR-1二本鎖を作ります命令とプライマー短期Dir1内-Rが3 'オーバーハングCCCGTを終了します。 DIR-1と相補的な3 'オーバーハングのACGGGを残すために、プライマー短期DIR2-Rを使用してDIR-2反転二本鎖を作るためにこれらのステップを繰り返します。
- 2 199 bpのfragmeを連結製造元の指示に従ってT4 DNAリガーゼでNTS。 PCRは、1.1.1プライマーを用いDirAll-CACC-FWとDirAll-NCO-RV( 表1)に示すように、得られた393 bpの断片を増幅し、プラスミドP2 + 4ENTRを作るためにpENTR1Aに、標準的な技術を使用して製品のクローンを作成。
- プラスミドp5XCAPDを行い、部分配列が続く、標準的な技術を用いて大腸菌DH5αに変換するために、製造業者の指示に従って、LRクロナーゼII酵素ミックスを使用して:pCAPD 29(KC176455.1のNCBIアク)にP2 + 4ENTRを再結合します。注:完全なRNAiインサートを表1に示します。
- 以前に30を報告したように、プラスミドp5XCAPDでアグロバクテリウム株 C58C1 30を変換して、次のようにピーナッツ植物を形質転換するために、得られた細菌を使用します。
- 50ミリリットルのLBブロスは500を補足し、30℃p5XCAPDを保有するアグロバクテリウムで生育このために使用μgのml -1のストレプトマイシン、25μgのml -1のゲンタマイシン、10μgのml -1のカナマイシン、および1 OD 260に達するまで、250 rpmで文化を振ります。
- 10〜14日齢の苗Exp27-1516、ランナーから外植細菌懸濁液中の100μMの1時間アセトシリンゴン、と場所で50mlのAB最小培地31で再懸濁し、10分間の遠心分離(6,000×gで)によってアグロバクテリウム細胞を収穫-typeピーナッツ育種系統。ブロットは、[30分後3MM吸い取り紙の外植片を乾燥し、シュート誘導培地(SIM)の上に置き、MS塩32、3%スクロース、20μMのベンジルアミノプリン(BAP)、10μMのチジアズロン(TDZ)、pHは5.8、0.3 3日間暗所で抗生物質を含まない%のジェランガム]。
- 33前に報告されているような組織の選択と再生を行います。 2ヶ月の隔週転送に、シュート形成のためにSIM(500μMのセフォタキシムおよび100μMカナマイシン)に組織を移動します。その後、場所数ヶ月シュート伸長培地(SEM)の新芽[5μMBAP、1μMのジベレリン酸(GA 3)]、隔週の拡大。
- 、個々の芽、根誘導培地(RIM)でサイズが2センチ、[1/2 MS、1.5%スクロース、5μMのα-ナフタレン酢酸(NAA)、2.5μMのインドール酪酸(IBA)]を配置しますその後、苗を馴化し、温室に転送します。
沈黙アフラトキシン合成遺伝子にRNAiを保有する2識別ピーナッツの植物
- RNAiを用いた形質転換のプロセスの対象となったピーナッツ植物の若い葉からDNAを抽出するために、製造業者の説明書に従って、200μlの溶出での使用ロボットワークステーションでの植物ミニキットは、(前述のように)バックボーンとして持つp5XCAPD( 図1)を構築遺伝子サイレンシングのためのプラスミドpCAPD 29。
- 説明previとしてシングルチューブネステッドPCR(STN-PCR)によってDNAサンプルのスクリーニングously 34は、選択可能なマーカーNPTIIを検出し、RNAiはp5XCAPDから挿入します。クローンこの第一世代ではテストのための十分な種子を生産するためにカット(3-4節)PCR陽性植物から伝播します。
- すべてのSTN-PCR反応におけるDNAの4つの2倍希釈(50〜100 ngのμL-1希釈前)を使用します。 5'-AGGCTATTCGGCTATGACTG-3 'とPCAPD 6446R:5'-CGTCAAGAAGGCGATAGAAG-3'、および内部プライマーPCAPD 5730F:5'-ACTGGGCACAACAGACAATC-3 'とPCAPD 6249R:5'-ATATTCGGCAAGCAGGCATC- NPTIIは、外部プライマーPCAPD 5714Fを使用3 '。
- 5'-CCTAACAGAACTCGCCGTAA-3 'およびDirAll-NCO-RV:5'-ATGCCATGGGGTTATTGGGTGCAGAATGG-3' RNAiの検出のために外部プライマー35S-PDSFw使用STN-PCR反応に挿入し、内部プライマーをProbe_5027_Fw:5'-gtatttgtgaccatgtttctg-3 'とProbe_7228_Rv:5'-GGACGGATAGTAAACTGCGG-3'。
- STN-PCR陽性植物からと詐欺の収穫ピーナッツポッド TROL植物は、同じ条件で成長させました。重要なステップ:対照植物は、同じ条件およびRNAiの植物と同じ季節で栽培されていることを確認してください。低強度で水の圧力洗浄機での爆風ポッドまたは、exocarpを削除(茶色と黒、黄色、オレンジ、)グループ内の成熟・ボードと個別のポッドにポッドを配置することにより、果皮の色を決定するために、手でこすり35( 図2)。

分析のためのピーナッツポッドの調製図。左:ピーナッツは不確定な成長植物であるとして、収穫時に見られる異なるピーナッツサイズ。センター:exocarpの水圧を除去するために金属かごにピーナッツを置く; 右:ピーナッツプロファイルボード(黄色、オレンジ、茶色と黒)の果皮色によって満期グループ。98fig2large.jpg「ターゲット= "_空白">この図の拡大版をご覧になるにはこちらをクリックしてください。
3.実験セットアップ
- 別途種を殻を外し、プロセス黄色と茶色。 表2によれば、実験で使用する種子の数を計算ピーナッツラインとサンプリング日ごとに3つの種子片(1種ピース=半子葉)の最小値が統計分析を行い、標準誤差を低減することが要求されることに注意してください。
| 名 | シーケンス | ||
| DIR-1 | 5'- GCCAGCTCAAAAGTGCGATGCACCAAGGAGAAACCGGCCTGTGCT CGGTGTATCGAACGTGGTCTTGCCTGTCAATACATGGTCGTATTTG TGACCATGTTTCTGGTGGCATTGGACCGTCTTGTCATCTCTACAGCC ATTCCCCAGATCACGGACGAATCCCCTGCATCTACGCGCACGCATC ACTTGGGGTACCCGT-3 ' | ||
| 短期Dir1内-R | 5'-フォス-TACCCCAAGTGATGCGTGCGCG-3 ' | ||
| DIR-2反転 | 5'- GGTTATTGGGTGCAGAATGGTAAACCACCCAACAGTACGCGAAATG TCAATTCCAGAGTCCCAAACCTCCCTACCGTGGCCTGGACGGATAG TAAACTGCGGAGCTTGGGAACAAAATCCGCTGTCTGATCGCCGAAG AGAAAGAGTTGCCTTGATTGAGCCGCATCGAGGACAGGTTGTGTTG CTGTTGATAGACGGG-3 ' | ||
| 短期DIR2-R | 5'-フォス-CTATCAACAGCAACACAACC-3 ' | ||
| DirAll-CACC-FW | 5'-CACCGCCAGCTCAAAAGTGCGATGC-3 ' | ||
| DirAll-NCO-RV | 5'-ATGCCATGGGGTTATTGGGTGCAGAATGG-3 ' | ||
| DirAll-BamEco-FW | 5'-ATGGGATCCGAATTCGCCAGCTCAAAAGTGCGATGC-3 ' | ||
| 完全なRNAiを挿入 | 5'- GCCAGCTCAAAAGTGCGATGCACCAAGGAGAAACCGGCCTGTGCT CGGTGTATCGAACGTGGTCTTGCCTGTCAATACATGGTC / GTATTTG TGACCATGTTTCTGGTGGCATTGGACCGTCTTGTCATCTCTACAGCC ATTCCCCAGATCACGGACGAAT / CCCCTGCATCTACGCGCACGCAT CACTTGGGGTACCCGTCTATCAACAGCAACACAACCTGTCCTCGAT GCG / GCTCAATCAAGGCAACTCTTTCTCTTCGGCGATCAGACAGCG GATTTTGTTCCCAAGCTCCGCAGTTTACTATCCGTCCA / GGCCACGG TAGGGAGGTTTGGGACTCTGGAATTGACATTTCGCGTACTGTTGGG TGGTTTACCATTCTGCACCCAATAACC-3 ' | ||
p5XCAPDをRNAi構築物を構築するために使用される 表1 オリゴヌクレオチドおよびultramersフォス:5 '末端をリン酸化し、。 「/」を使用し5遺伝子断片を分離し;完全なRNAiインサート:p5XCAPDを形成するために2逆方向反復として使用される配列。
- 彼らは無菌ビーカーの底をカバーするように、単層で全体ピーナッツの種子を置きます。種子を被覆した後、同じ溶液の等量を追加し、75%エタノール/水(v / v)の溶液を加えます。 30秒間周囲温度でインキュベートした後、滅菌脱イオン水(SDW)ですすいでください。
- 次のようにエタノール処理種子を含有するビーカーに2%次亜塩素酸を追加し、同じ溶液の等量を添加し、5分間インキュベートし、シードをカバーするために十分な2%次亜塩素酸溶液を添加します。重要なステップ:( 図3使用次亜塩素酸塩の5倍のボリュームにSDWと同等の容量で十分に3回すすぎ)。

3.実験のセットアップ図。トップ:前と次亜塩素酸後のピーナッツ種子の表面滅菌、種皮(種皮)の除去; 中央:胚と胚の近景の除去は、その後、半分に子葉をカット; 下:半子葉を滅菌蒸留水に、無菌の吸収性にブロッティング紙、水寒天上のへこみ、および寒天表面上半分子葉(上向きカット側)を配置。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
- 表面殺菌種子を2時間SDWに沈め吸収するようにします。滅菌シャーレに種子を置き、ピンセットで種皮を除去し、子葉を分離し、メスで胚を削除します。注意:胚は廃棄されるか、または新しい植物を再生するために使用することができます。
- メスを用いて半分に各子葉をカット。脱水を避けるために、すべての種子が処理されるまで滅菌水中カット種子片を保持します。
- 、各3種子片のための1つ、滅菌水寒天を含むペトリ皿(W / V 1.5%寒天/水)を用意しました。ピンセットを用いて寒天に小さなへこみを行い、水/寒天プレート上(上向き切断面)を簡単に滅菌ペーパータオルの種子片の余分な水分をブロットした後、種子片を配置します。
- 10日間25℃で増殖させツァペックの寒天培地でアフラトキシン生産アスペルギルス・ フラバス NRRL 3357の新鮮な培養から、血球計で計数μlのSDWあたり50,000胞子の懸濁液を調製。
- 胞子はRNAiを( 図4)を保有種子組織に暴露され得ることを確認するために、両側に流出を避ける各半子葉片の切断面に胞子懸濁液の2μLを置 きます。

アフラトキシンの分析については、図4の接種およびインキュベーション。トップ:ハーフ子葉、胞子懸濁液を接種し、24時間のインキュベーション後の半子葉のアスペルギルス・フラバス菌糸成長下:左:1.5%寒天培地上、48時間のインキュベーション;センター:1.5%寒天上で72時間のインキュベーション;右:誤った実験の例、0.5%寒天の72時間のインキュベーションこの図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
- サンプリングまで暗所で30℃で接種し、非接種半子葉を含むペトリ皿をインキュベートします。
アフラトキシン分析および遺伝子発現のために4サンプリング
- サンプルを収集中RT-PCR用とアフラトキシン分析用の両方のインキュベーションの24、48および72時間で三連(オプションの96時間)は、それぞれが1枚(半子葉)である複製します。各サンプリング日の異なるプレートからランダムにサンプルを選んでください。組織を使用して、そっとバイアルまたは試験管内で種子片を配置する前に、寒天と過剰真菌胞子を削除します。
- アフラトキシン分析のために、-80℃で4ミリリットルガラススクリューキャップバイアルとストア内の各反復(1枚)を配置。注:アフラトキシンの化学的安定性を考慮すると、試料は(通常1-2ヶ月現在の実験において)、数ヶ月間、この状態で保存することができます。
- RT-PCRの場所については各反復(1枚)で、すでに2つのステンレス鋼ビーズ(2.5ミリメートルDIAM)と3つのジルコニウムビーズ(2mmのDIAM)を含有するチューブを研削2ミリリットルを用意しました。
- 直ちに、すべてのサンプル(好ましくは液体窒素中で)凍結し、処理するまで-80℃で保管してください。
個々の半分のCの5アフラトキシン分析otyledon小品
- A.を使用してくださいフラバスは、この分析のためのサンプルを接種しました。約30分間周囲温度にサンプルを持参、4つのボリュームを追加(通常は2〜3ミリリットルを、w / v)のメタノールのキャップを閉め、(後の計算のための記録を保持)、および中のO / N(〜16時間)インキュベート撹拌せずに暗いです。
- 、マッチング1.5ミリリットルプロピレンミニカラムにフリットを配置した Al 2 O 3の基本を200mgを追加し、以前に36に従い、別のフリットとそれをキャップ。その後、溶出液の可能性蒸発を避けるために十分に近い列の下に超高速液体Chomatographer(UPLC)オートサンプラーバイアルを置きます。
- 使い捨てガラス試験管(プラスチックを使用していない)では、ステップ4.1で得られたメタノール抽出物0.5ミリリットルを配置0.5ミリリットルのアセトニトリルを加え、ピペットで混合し、ステップ4.2で調製したカラムに混合物の0.5ミリリットルを適用します。 (圧力をかけないでください)、重力によってオートサンプラーバイアルに溶出を許可します。溶出は、通常2-4分、近くに目を取りEバイアルは、直ちにセプタムとUPLC互換性のキャップを使用。周囲温度でバイアルを維持し、UPLCで同じ日にそれらを分析。
- アフラトキシンを分離するため、マッチングUPLC第四紀ソルベントマネージャ、UPLCサンプルマネージャ、UPLC蛍光検出器を搭載したUPLC機器に(A.フラバスを使用した場合 、B 1およびB 2)アフラトキシン標準とサンプルの溶出液とオートサンプラーバイアルを含む場所のオートサンプラーバイアル、 C 18 2.1ミリメートル×50mmの、1.7μmのカラム。
- 0.30 mlの分の流速で水/メタノール/ CH 3 CN(64:23:13、v / v / v)の混合物からなる定組成移動相を使用して、-1。製造元の指示37に従って、アフラトキシン濃度の正確な計算のための安定したベースライン分離を保証するクロマトグラムを取得します。
- その質量スペクトルデータを取得し、公開されたデータ38とそれらを比較することにより、アフラトキシンの身元を確認してください。私を使用してくださいトラップ質量分析計でESIインターフェイス及び製造者の指示に従って、対応するソフトウェアを備えました。
- アフラトキシンB 1、B 2の商業規格に対応し、異なる量を注入することにより得られた較正曲線を参照することにより、アフラトキシンの濃度を決定し、G 1およびG 2 UPLC製造元が推奨およびソフトウェア37によって決定されます。
- 場所種子片はすでに乾燥重量を決定するために、O / N(〜16時間)、凍結乾燥のための個々のガラスバイアルに、メタノールで抽出しました。そして、中にNG。種子片の乾燥重量のグラム-1アフラトキシン濃度を計算します。
- 平均比較のためTukeyの検定に続いて、データの分析のために、(NG。グラム-1 1)ログインするアフラトキシン結果を変換します。
6.遺伝子発現、サンプルのRT-PCR処理
- チューブが含まれている研削2ミリリットルを取ります40秒間3,100 rpmでビーズミルホモジナイザーでそれらを挽く(解凍せずに)直ちに-80℃の冷凍庫からサンプルをる、とし、製造元の指示に従ってトリゾール用いてRNA抽出に進みます。
- 1μgのRNAおよびオリゴdTおよびランダムヘキサマーの等量を使用して、各サンプルからcDNAを準備し、1メイク:cDNAの8希釈し、(以前に39を説明したように)、RT-PCR反応あたり2μLを使用しています。
- RNAiの挿入用プライマーの発現の検出のために:RT_5X_1_105F:5'GGTGGCATTGGACCGTCTTG-3 '、RT_5X_1_232R:5'-CGCATCGAGGACAGGTTGTG-3'。そしてRT_5X_2_95F:5'-CCATGTTTCTGGTGGCATTG-3 '、RT_5X_2_229R:5'-ATCGAGGACAGGTTGTGTTG-3'。
- NPTII選択マーカーの発現を検出するための、使用プライマー:RT_NPTII_1_6871F:5'-CTCGCTCGATGCGATGTTTC-3 '、RT_NPTII_1_7004R:5'-GCAGGATCTCCTGTCATCTC-3'。アクチン-FW:CACATGCCハウスキーピング遺伝子の標準化のためのアクチン、およびプライマーを使用してくださいATCCTTCGATTG;アクチン-RV:CCAAGGCAACATATGCAAGCT 40。
- アクチン発現のための標準化デルタ・デルタC T法 41で結果を分析します。対照に対する倍の増加などの結果を示しています。
結果
プラスミドp5XCAPDはpCAPD 29の誘導体としてなされたものであり、ピーナッツ植物を形質転換するためにそれを使用しました。このベクターは、Aのアフラトキシン合成遺伝子の5小さな断片の逆方向反復、70〜80塩基対のそれぞれを、運びますイントロン( 図1)によって分離フラバス 。 AFL2G_07224(aflR)、AFL2G_07223(AFLSまたはaflJ)、AFL2G_05027(アフラトキシン排出ポンプ、aflep)、AFL2G_07228(AFLC / pksA / pksL1)、およびAFL2G_07731(PES1)は構造物に使用された、 図1相当の数のフラグメントに。 BROAD研究所、ケンブリッジ、マサチューセッツ州、および文献42でフラバスのゲノムアノテーション。 99ピーナッツ行の合計は、変換プロセスを経て再生させた、50は、PCRは、STN-PCRによって検出NPTII陽性であった、と33行がPCR陽性と生産種子でした。のみ7 PCR陽性ラインがクローン増殖とPRESENによって試験しましたアフラトキシンの蓄積のためのT法は、すべての7の間には、コントロールよりも60%と100%未満アフラトキシンの蓄積を示しました。ここでは、それらの7行の2の結果を示します。個々のトランスジェニック事象は、通常、いくつかの種子を生産するように、この方法は、依然としてパラメトリック統計解析を行うことができながら、種子数の最小値を使用するために開発されました。試料調製および実験のフローチャートを図5 及び表1に示されている。トランスジェニック種子の最初の世代は、典型的には、ヘミ接合であるが、それが細胞の細胞及び低分子干渉RNA(siRNA)の全身の動きが発生することを期待されていますRNA干渉を介してプラント全体のアフラトキシン合成サイレンシングを与える必要があります。

方法の概略 図5 のフローチャートは、サイレンシングにおいてRNAiの有効性を分析します ピーナッツ種子における アスペルギルス アフラトキシン合成遺伝子 。ワークフローのグラフィック表現遺伝子発現またはアフラトキシン分析のためのピーナッツサンプルを処理する。 この図の拡大版をご覧になるにはこちらをクリックしてください。
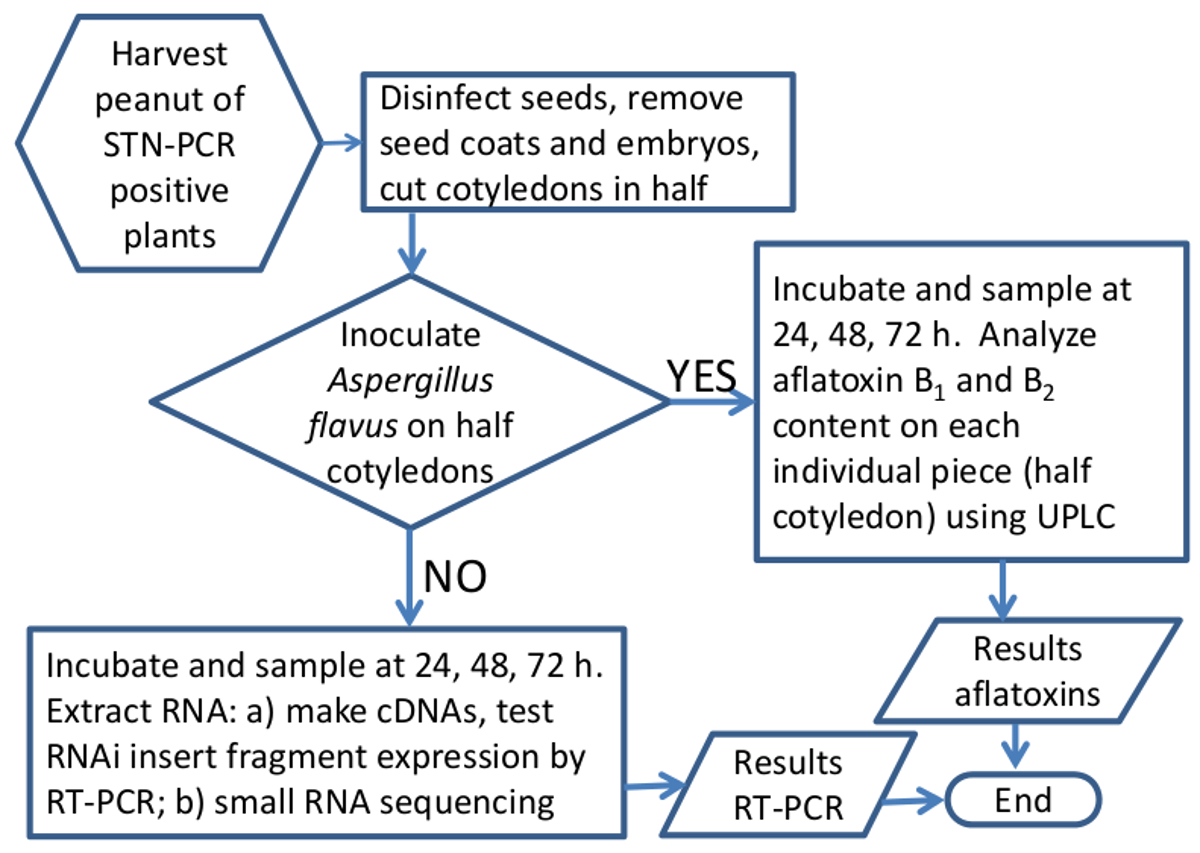{kind=link}
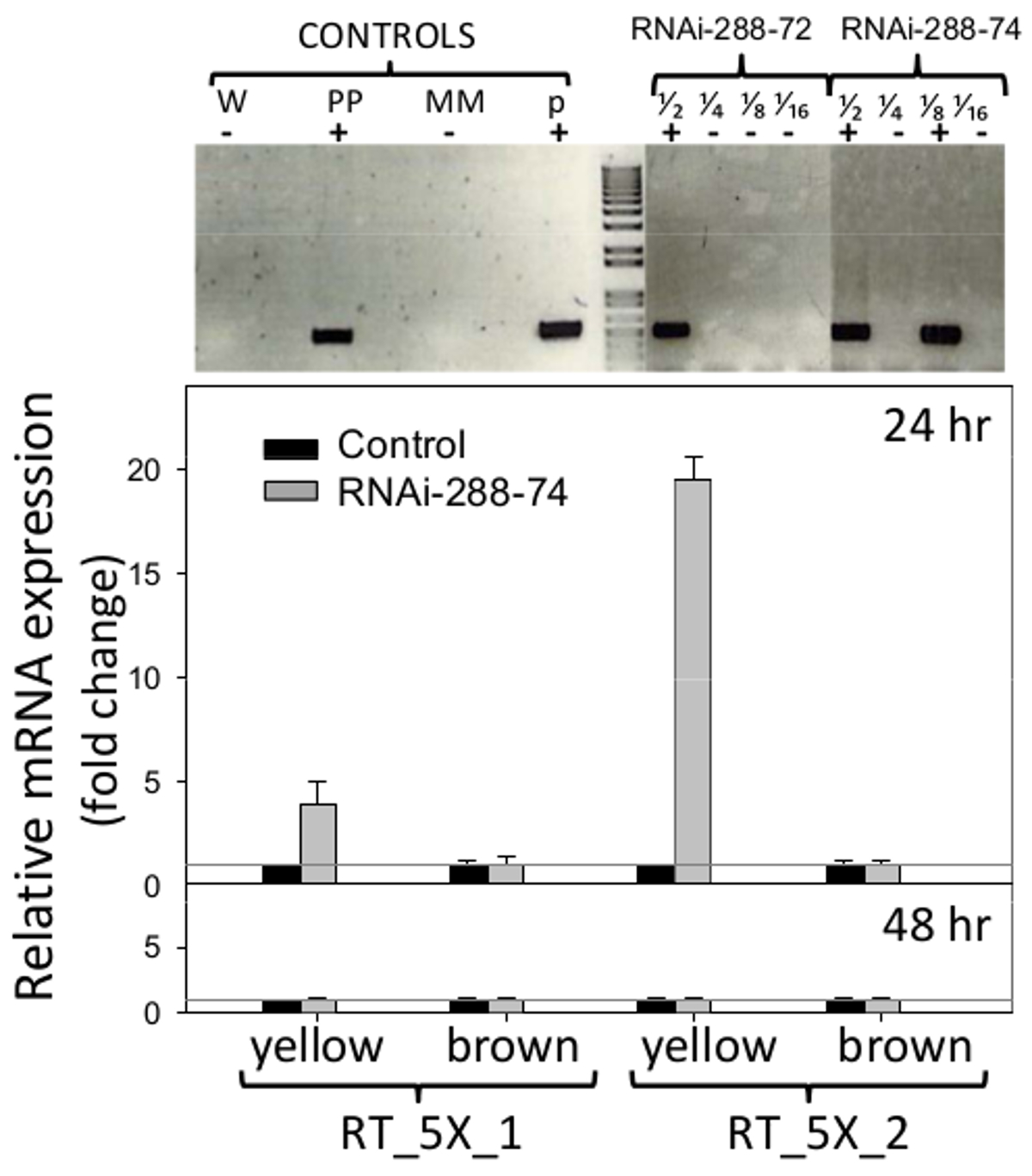
RNAiのピーナッツライン288から72と288から74ゲル部分は、図6(上)に示されているSTN-PCRによって試験したとき、NPTII選択可能メーカーの存在を示した、オリジナルの画像は、要求に応じて作成者から入手できます。それは二つの遺伝子には、cmr(クロラムフェニコール耐性)および植物に未知の効果のCCDB(毒素)の逆方向反復をコードしているので、プラスミドpCAPDは、形質転換のコントロールとして使用されませんでした。 PCRネガティブピーナッツライン288から9と再生プロセスを経て、RNAiのラインと同じ条件で増殖させた他は、負の詐欺として使用しましたトロール。

トランスジェニックの 図6. 検出とRNAiのリアルタイム表現挿入 トップ :シングルチューブ、トランスジェニックピーナッツラインのRNAi-288から72およびRNAi-288から74のネストされたPCR検出、陽性植物。コントロール:W(水)、PP(正の植物)、MM(マスターミックス)、P(プラスミドp5XCAPD)。画分を1/2〜1/4〜1/8 1/16 DNAの2倍希釈を表す下 :(未熟で、 図1のようRT_5X_1、RT_5X_2:RNAiの発現のリアルタイムPCR検出は、(プライマーセットを挿入しますイエロー)、24および48時間のインキュベーションでのトランスジェニック系統の(茶色)子葉を成熟;灰色の線:C T = 1ヒストグラムが3生物学的サンプルの平均および標準誤差バー(T)を表します3つの技術の反復で。 RNAiのインサートの相対的な定量化は、内部統制と5.2.2と5.3で説明したように計算された導入遺伝子の比較倍式としてハウスキーピング遺伝子アクチンに関して標準化した。 この図の拡大版をご覧になるにはこちらをクリックしてください。
{kind=link}
アフラトキシン蓄積の植物宿主のRNAi媒介電位制御の有効性を試験するために、 アスペルギルス・ フラバス NRRL 3357の新たに採取した分生子は、胚および種皮を除去した半子葉の切断面に塗布した( 図3、図4)。ゲノムが配列決定され、p5XCAPDを設計するための基礎となったされたA. フラバスの NRRL 3357は、親切にUSDA-ARS-NPRL博士ホーンにより提供されました。 30℃で24、48および72時間後の半子葉の結果として真菌の侵入はショーです nは、図4に。接種半子葉のサンプルは、72、48、24で収集したインキュベーションの96時間、およびLC-MSによって主4アフラトキシンB 1、B 2、G 1及びG 2 UPLCを使用するために分析し、確認されました; 結果を図6に示す 。アフラトキシン濃度は変更36で公開された方法を用いて決定しました。 96時間のインキュベーションでは、子葉が原因真菌感染症の崩壊を開始します。 RNAiのライン288から72には、未熟子葉内のすべてのサンプリング日における成熟したものの中で最もサンプリング日における対照よりもアフラトキシンの有意に低いレベルを示しました。 RNAiのライン288から74には、ほとんどのサンプリング日のアフラトキシンの有意に低いレベルを示しました。 Tukeyの検定の有意のレベルは、図7のグラフにアスタリスクで示されています。
g7.jpg "/>
アスペルギルス・フラバス とのインキュベーション(24、48、72および96時間)の後半分ピーナッツ子葉図 7. アフラトキシンB 1 およびB 2コントロール:。288から9非トランスジェニックラインの種子; RNAi:RNAiを288から72およびRNAi-288から74、RNAiのp5XCAPDためのトランスジェニックからの種子は5アフラトキシン合成遺伝子を沈黙させます。 (A)成熟種子中のアフラトキシンB 1(ブラウン)。 (B)アフラトキシンB 2成熟種子。未熟種子中の(C)未熟種子中のアフラトキシンB 1(黄色)と(D)アフラトキシンB 2。重複した生物学的サンプルの対応する標準エラーバー(T)との平均値を表しています。統計的に有意な差Tukeyの検定*:0.05≤P、**:P≤0.01、***:P≤0.001。S://www.jove.com/files/ftp_upload/53398/53398fig7large.jpg「ターゲット= "_空白">この図の拡大版をご覧になるにはこちらをクリックしてください。
全体的に、RNAiを288から72には、コントロールと比較した実験(24〜96時間のインキュベーション)、アフラトキシンB 2の 94%-100%の減少、及びアフラトキシンB 1が90%-100%の減少を通じて示しました。 RNAiを288から74には、アフラトキシンB 1、 図7のアフラトキシンB 2、60%-100%の減少で63%-100%の減少を示しました。
RNAiの挿入物の発現のリアルタイムPCR検出のために使用したプライマーは、図1に示されている。ピーナッツ種子の子葉は、それらの自然の防御を除去し、最もにさらされる領域にRNAiの潜在的影響を検出することができるように、胚ずに分析しました真菌の侵入、子葉。挿入RT_5X_1をプライマーセットによってライン288から74の未熟子葉(黄色)で検出されたRNAiの発現は、C Tの上に4倍でした = 1ネガティブコントロールのしきい値、およびRT_5X_2をプライマーセットによっては、すべての24時間のインキュベーションで、しきい値を超えて19倍でした。ピーナッツ変換に使用される5つの連続した遺伝子断片のうち、少なくとも3、5027、7223および7228(それぞれaflep、AFLS / aflJ、及びAFLC / pksAは、)、RT-PCR( 図1,6)によって検出しました。 RNAiのインサートの発現は24時間で、または48時間のインキュベーション、 図6(下)での成熟または未熟子葉の成熟した子葉では検出されませんでした。
1| ピーナッツライン | サンプリングの時間 | RT-PCR用のサンプル | アフラトキシン分析用サンプル(接種) | N種子のアンバー | ||||
| (非接種) | ||||||||
| 担当者1 | 担当者2 | 担当者3 | 担当者1 | 担当者2 | 担当者3 | |||
| RNAiの(黄色) | 24時間 | 1 | 1 | 1 | 1 | 1 | 1 | 4.5 |
| 48時間 | 1 | 1 | 1 | 1 | 1 | |||
| 72時間 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| 制御 | 24時間 | 1 | 1 | 1 | 1 | 1 | 1 | 4.5 |
| (黄) | 48時間 | 1 | 1 | 1 | 1 | 1 | 1 | |
| 72時間 | 1 | 1 | 1 | 1 | 1 | 1 | ||
表2。 小サンプルのセットアップの例は、RNAiピーナッツ種子における遺伝子発現とアフラトキシンの蓄積を分析した。完全な発現の解析と1満期グループのアフラトキシンを( すなわち、黄色)、3サンプリング時間(24、48および72時間)で、三連で、4.5種子を必要とする、テーブル内の各ナンバーワンは半分子葉を表します。
ディスカッション
真菌病原体における遺伝子の植物宿主のRNAi媒介サイレンシングは、27,43を実証されている、しかし、植物中のマイコトキシン蓄積のRNAi仲介制御の実現可能性を示す何の出版物はありません。葉は地下ポッドの真菌感染時には症状を示さないようピーナッツにおけるこれらの研究のための一つの制限要因は、個々の植物における無アフラトキシン蓄積表現型を評価する方法がないことでした。また、アフラトキシンのない正規分布の蓄積、および化学分析15,16のための大規模なサンプルの必要性は単一の植物の潜在的なRNAi効果の定量化を妨げています。ここで紹介する方法は三通り3 24時間間隔サンプリング( 表1、 図7)を実行するために5種を使用して、72時間の実験で構成されています。種子の劣らず100 g以上を必要とする典型的なアフラトキシン分析と比較して、私たちの方法はindividuaに特に適しています最初に2つ以下または3ポッドを生産ピーナッツ植物のLトランスジェニックイベント。
アフラトキシン合成のRNA媒介サイレンシングは、遺伝的にアスペルギルス・フラバス と A. を形質転換することにより実証されていますparasiticus。 aflRは Aのアフラトキシン生産の主要な調節因子であるので、 フラバス と A. parasiticus 44,45、それは植物におけるRNA媒介サイレンシングのための興味深い対象となります。しかし、aflRにおける遺伝的変異は、 アスペルギルス種 46の間で示されており、植物宿主で生産するRNAi信号と一致する完全なシーケンスが存在しない場合には、これらの遺伝的変異は、サイレンシングから逃れることができました。したがって、aflRは、ベクトル p5XCAPDにサイレンシングのための目標の一つでしたが、一つだけではありませんでした。 A.に導入aflR遺伝子の逆方向反復フラバス と A.変換によるparasiticusはサイレンシングし、最小限または全くない製品が得られましたアフラトキシン47のイオン(マクドナルドら 、2005B)。また、サイレンシングAFLD遺伝子は、Aに 98%までによってアフラトキシン生産を防止しましたフラバス と A.直接変換48でparasiticus。我々のシステムの成功の確率を高めるために、ピーナッツはAにアフラトキシン産生に関与する5つの遺伝子の逆方向反復断片で形質転換しましたフラバス 。ここでは、アフラトキシン合成経路においていくつかの遺伝子をサイレンシングするためのターゲット、アフラトキシンB 1の90%-100%低いレベルおよびB 2は、ライン 288から72に達成されたことをp5XCAPDを使用して、とに蓄積された百分の60から100まで低いレベルことが示されています半子葉がAで接種した対照と比較してライン288から74、 フラバスは 、7,4を図 。最も重要なのは、この方法は、ライン288から72によってアフラトキシンの蓄積に統計的に有意な差異を検出した288から74 対パラメトリックSTATISを適用することにより、実験を通して制御チック、 図7。小さなサンプルサイズを考えると、それは、これらの実験は、高解像度、より高い性能と比べて3倍の感度を5倍を有しUPLCで分析したアフラトキシンを検出するための強力な方法を使用する必要性を強調することが重要ですHPLC 49。
RNAiの発現は、インサート288から74にのみ24時間のインキュベーションでの未熟子葉(黄色)で検出されました。 RNAiのインサートは、図6、48時間で24時間で288から74の成熟子葉に、または任意の満期のグループにRT-PCRによって検出されなかった。これと同じ現象は他のRNAiトランスジェニックピーナッツ株において観察された(アリアス、RS、2015未発表)、通常のRNAi転写物はわずか24時間で未熟子葉上で検出された場所。 RNAサンプルは、データは、アクチンの発現レベルに対して標準化し、DNAの混入の証拠は観察されなかった、cDNA合成の前にDNアーゼで処理しました。 DNAが試料中に存在していたならば、それはすべき同様に48時間のサンプルで検出されたが、一貫して、それはそうではありませんでしたされています。 35Sプロモーターの制御下の発現は必ずしも一様ではありません。それは、環境条件50、組織や発達段階51,52の種類によって影響を受けることができます。同時に、RNA干渉の経路では、mRNAの崩壊速度およびsiRNAの減衰率が大きく53を変化させることができます 。これは、RNA干渉の機構によるmRNAの急速な分解は、48時間のインキュベーションでのmRNAの検出を妨げている可能性があります。 48時間での発現の欠如が低い35Sプロモーター駆動の転写によるものであったかどうか、またはダイサーによりdsRNAの速い劣化に答えられることを残ります。このように、ハイスループットシークエンシングによって、小さなRNAの検出は、RNAi 54を介して行われているプロセスのより深い洞察を与えるだろう これらの実験です。しかし、RNAサイレンシングは、主にphotosyntから師部を通って、全身に広がるので、(この場合はピーナッツ種子中)ショ糖シンク55にソースを嫌い、アフラトキシン合成のサイレンシングは、RNAiインサートの局所発現せずに種子に発生する可能性があります。多くの研究は、種子中のアフラトキシンの蓄積を防止するために必要な小さな干渉RNA(siRNA)の閾値レベルを決定するために行われていません。これは、RNAiの両方のmRNAの発現は( 図6)を構成するという事実を強調することが重要であり、アフラトキシンB 1及びB 2( 図7)の蓄積は、 対未成熟(黄色)に対して異なる結果を示しました。成熟した(茶色)子葉。ピーナッツ植物はつまり、彼らは収穫満期ポッドの範囲、 図2に提示し、不確定な成長を持っている。また、異なる満期グループからの種子は、その化学組成が異なる例えば 、2.4%の未熟種子中のショ糖、および1.9%で同じフィールド条件56,57の下で成熟した種子。したがって、実際のefficiencを理解しますアフラトキシン蓄積のRNA媒介性制御のyは、別途成熟グループを分析することが重要です。
ピーナッツ種子の自然な防衛は種子や環境条件58-61の成熟度に応じて製造された化合物およびそれらの相対的な量の多様性に変化ファイトアレキシンの生産、であり、子葉62に比べて胚において特に高いです。胚はまた、子葉(アリアスRS、未発表)よりもDNAとRNAの両方を、核酸の有意に高い濃度を有します。ピーナッツ種子が成熟するにつれ、その生理学および化学組成の変化が63を発生します。静真菌活性を64とピーナッツ種皮フォーム縮合型タンニンでフェノール系酸化防止剤;タンニンとフェノール化合物のその含有量は、満期65が増加するにつれて、これは、また、35黒黄色成熟段階を反映中果皮の色が明らかです。トンのこのように、存在その抗菌性所与の実験でESTAまたは胚は、制限された真菌の成長を持っているので、のRNAiサイレンシングの効果を過大評価ができ、従って、これらを除去しました。また、種皮および胚の除去は、胚がよりファイトアレキシンとより多くのRNA含量を持つことになります運ぶ半分子葉として、分析のばらつきの原因を制限することができます。
これらの実験における成熟グループと種皮と胚芽を除去することによって分析に加えて、さらにいくつかの観測を指摘することは重要である。結果は最大96時間のインキュベーションのために示されているもののA)は、72以下で使用しないことが推奨されます種子は96時間によって分解を受けるように、一貫性のある結果を得るための時間。およびb)同じシードから半子葉は、ランダムにサンプリングされたものの、完全に独立したサンプルを構成しないのに対し、RT-PCRおよびトランスジェニック事象の中アフラトキシンの蓄積が種子の間の最小変動を示しました。また、正確な真菌の胞子数、接種量2μLのS、および側面に滴下を回避子葉の切断面上の胞子のアプリケーションは、発芽胞子を植物組織にさらされていることを確認してくださいすることが重要です。 図4(下)の最後のフレームに示すように、プレート上に水/寒天を1.5%(w / v)のにする必要があり、より柔らかい寒天は胞子の流出が発生します。制限され、特定のトランスジェニック事象から可用性をシードする必要があり、サンプリングではなく、同様の結果( すなわち 、 図7)を得た三連の複製で行うことができます。しかし、三連のサンプルは、標準誤差を減らすのに役立ちます。この方法の唯一の制限は、それがアフラトキシン検出/定量化のための高感度システム(UPLC)が必要なことであるが、同時にこれはあまり高感度な方法によってアフラトキシン検出されてはならないRNAiの効果を過大評価の可能性を低減します。
結論として、この方法は、初めて勉強する信頼性の高いアプローチを提供していますアフラトキシンの制御におけるRNAiの効果。一週間以内に全体の作付けシーズンから実験のための時間を削減する、この方法は、途方もなく緩和および/ またはアフラトキシンの排除に向けたRNAi-ピーナッツ/ アスペルギルス pathosystemの研究を加速していきます。
開示事項
The authors have nothing to disclose or any conflicts of interest.
謝辞
This work received the financial support of USDA-ARS CRIS project 6604-21000-004-00D, CRIS project 6604-42000-008-00D, and USAID Feed-the-Future program Agreement number 58-0210-3-012. We thank Valerie Orner, LaTanya Johnson, Joseph Powell and Kathy Gray for their technical assistance. Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the US Department of Agriculture.
資料
| Name | Company | Catalog Number | Comments |
| Primers, oligonucleotides | DNA Technologies, Coralville, IA, USA | n/a | |
| Dneasy Plant Mini Kit | Qiagen, Valencia, CA | 69106 | |
| Czapek Dox agar medium | Oxoid, by Thermo Fisher Scientific, Waltham, MA | CM0095 | |
| Agar | Thermo Fisher Scientific, Waltham, MA | BP 1423 | |
| Freezer -80 °C | n/a | n/a | |
| Aluminum Oxide, Al2O3 | Fisher Scientific | A941 | |
| SPE Reservoirs 1.5 ml | Grace Davison Discovery Scientific | 210011 | |
| Frits for 1.5 ml SPE reservoir | Grace Davison Discovery Scientific | 211401 | |
| Autosampler vials | Waters Corporation, Milford, MA | 186005221 | |
| Waters Acquity Ultra-Performance Liquid-Chromatography (UPLC) instrument; UPLC-H-Class Quaternary Solvent Manager; UPLC Sample Manager; UPLC Fluorescent detector (FLR); UPLC BEH C18 2.1 mm x 50 mm, 1.7 mm column | Waters Corporation, Milford, MA | ||
| Finnigan LCQ Advantage MAX ion trap mass spectrometer, with Xcalibur version 1.4 software | Thermo Electron Corp., San Jose, CA | ||
| Aflatoxin standards, B1, B2, G1 and G2 | Sigma-Aldrich, St. Louis, MO | A6636; A9887; A0138; A0263 | |
| Systat Software 12.2 | SYSTAT Software Inc., Point Richmond, CA | ||
| Trizol reagent | Invitrogen, CA | 15596-018 | |
| SuperScript III First Strand Synthesis Super Mix | Invitrogen, CA | 11752-050 | |
| ABI 7500 Real-Time PCR | Lifetechnologies, Grand Island, NY | 4406984 | |
| Luria Broth-Miller | Fisher Scientific | R453642 | |
| pENTR1A | Invitrogen, CA | A10462 | |
| LR Clonase II enzyme mix | Invitrogen, CA | 11791-020 | |
| T4 DNA Ligase | NEB Biolabs | M0202L | |
| Gelrite | Sigma-Aldrich, St. Louis, MO | G1919 | |
| Acetosyringone | Sigma-Aldrich, St. Louis, MO | D134406 | |
| QIAcube robot workstation | Qiagen, Valencia, CA | 9001292 | |
| Antibiotics: kanamycin, cefotaxime, gentamicin; streptomycin | Goldbio, St. Louis, MO | cef.: C-104-25; kan: K-120-5; gent.: G-400-1; strep.: S-150-50 | |
| Platinum Taq DNA Polymerase High Fidelity | Invitrogen, CA | 11304-029 |
参考文献
- Williams, J. H., et al. Human aflatoxicosis in developing countries: a review of toxicology, exposure, potential health consequences, and interventions. The American Journal of Clinical Nutrition. 80, 1106-1122 (2004).
- American Association for Cancer Research: AACR. An evaluation of chemicals and industrial processes associated with cancer in humans based on human and animal data: IARC Monographs Volumes 1 to 20. Cancer Research. 40, 1-12 (1980).
- Turner, P. C. The molecular epidemiology of chronic aflatoxin driven impaired child growth. Scientifica. , (2013).
- Rasooly, R., Hernlem, B., He, X., Friedman, M. Non-linear relationships between aflatoxin B1 levels and the biological response of monkey kidney vero cells. Toxins (Basel). 5, 1447-1461 (2013).
- Gong, Y. Y., et al. Determinants of aflatoxin exposure in young children from Benin and Togo, West Africa: the critical role of weaning. International Journal of Epidemiology. 32, 556-562 (2003).
- Eaton, D. L., Groopman, J. D. The toxicology of aflatoxins: human health, veterinary, and agricultural significance. , Academic Press. (1994).
- Murugavel, K. G., et al. Prevalence of aflatoxin B1 in liver biopsies of proven hepatocellular carcinoma in India determined by an in-house immunoperoxidase test. Journal of Medical Microbiology. 56, 1455-1459 (2007).
- Wang, J. S., et al. Hepatocellular carcinoma and aflatoxin exposure in Zhuqing Village, Fusui County, People's Republic of China. Cancer Epidemiology, Biomarkers & Prevention. 10, American Association for Cancer Research. 143-146 (2001).
- Azziz-Baumgartner, E., et al. Case-control study of an acute aflatoxicosis outbreak, Kenya, 2004. Environmental Health Perspectives. 113, 1779-1783 (2005).
- Lye, M. S., Ghazali, A. A., Mohan, J., Alwin, N., Nair, R. C. An outbreak of acute hepatic encephalopathy due to severe aflatoxicosis in Malaysia. American Journal of Tropical Medicine and Hygiene. 53, 68-72 (1995).
- Villers, P. Aflatoxins and safe storage. Frontiers in Microbiology. 5, 158(2014).
- Kensler, T. W., Roebuck, B. D., Wogan, G. N., Groopman, J. D. Aflatoxin: A 50-year odyssey of mechanistic and translational toxicology. Toxicological Sciences. 120, S28-S48 (2011).
- Dorner, J. W., Cole, R. J., Wicklow, D. T. Aflatoxin reduction in corn through field application of competitive fungi. Journal of Food Protection. 62, 650-656 (1999).
- Cotty, P. J., Bhatnagar, D. Variability among atoxigenic Aspergillus flavus strains in ability to prevent aflatoxin contamination and production of aflatoxin biosynthetic-pathway enzymes. Applied and Environmental Microbiology. 60, 2248-2251 (1994).
- Whitaker, T. B. Standardisation of mycotoxin sampling procedures: an urgent necessity. Food Control. 14, 233-237 (2003).
- Whitaker, T. B., Dorner, J. W., Giesbrecht, F. G., Slate, A. B. Variability among aflatoxin test results on runner peanuts harvested from small field plots. Peanut Science. 31, 59-63 (2004).
- Fire, A., et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 391, 806-811 (1998).
- Rafael, D., et al. EMT blockage strategies: Targeting Akt dependent mechanisms for breast cancer metastatic behaviour modulation. Current Gene Therapy. , (2015).
- Li, G., Chang, H., Zhai, Y. P., Xu, W. Targeted silencing of inhibitors of apoptosis proteins with siRNAs: a potential anti-cancer strategy for hepatocellular carcinoma. Asian Pacific. Journal of Cancer Prevention: APJCP. 14, 4943-4952 (2013).
- Koldehoff, M. Targeting bcr-abl transcripts with siRNAs in an imatinib-resistant chronic myeloid leukemia patient: challenges and future directions. Methods in Molecular Biology. 1218, 277-292 (2015).
- Zhang, J., et al. Pest control. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science. 347, 991-994 (2015).
- Ajjappala, H., Chung, H. Y., Sim, J. S., Choi, I., Hahn, B. S. Disruption of prefoldin-2 protein synthesis in root-knot nematodes via host-mediated gene silencing efficiently reduces nematode numbers and thus protects plants. Planta. 241, 773-787 (2015).
- Jose, A. M., Hunter, C. P. Transport of sequence-specific RNA interference information between cells. Annual Review of Genetics. 41, 305-330 (2007).
- Vazquez, F., Hohn, T. Biogenesis and biological activity of secondary siRNAs in plants. Scientifica. , Hindawi Publishing Corporation. (2013).
- Tinoco, M. L. P., Dias, B. B. A., Dall'Astta, R. C., Pamphile, J. A., Aragao, F. J. L. In vivo trans-specific gene silencing in fungal cells by in planta expression of a double-stranded RNA. BMC Biology. 8, (2010).
- Govindarajulu, M., Epstein, L., Wroblewski, T., Michelmore, R. W. Host-induced gene silencing inhibits the biotrophic pathogen causing downy mildew of lettuce. Plant Biotechnology Journal. , (2014).
- Yin, C., Jurgenson, J. E., Hulbert, S. H. Development of a host-induced RNAi system in the wheat stripe rust fungus Puccinia striiformis f. sp. tritici. Molecular Plant-Microbe Interactions. 24, 554-561 (2011).
- Ghag, S. B., Shekhawat, U. K., Ganapathi, T. R. Host-induced post-transcriptional hairpin RNA-mediated gene silencing of vital fungal genes confers efficient resistance against Fusarium. wilt in banana. Plant Biotechnology Journal. 12, 541-553 (2014).
- Filichkin, S. A., et al. Efficiency of gene silencing repeats vs. transitive RNAi in Arabidopsis: direct inverted vectors. Plant Biotechnology Journal. 5, 615-626 (2007).
- Sciaky, D., Montoya, A. L., Chilton, M. D. Fingerprints of Agrobacterium Ti Plasmids. Plasmid. 1, 238-253 (1978).
- Clark, D. J., Maaloe, O. DNA Replication and Division Cycle in Escherichia coli. Journal of Molecular Biology. 23, 99-112 (1967).
- Murashige, T., Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plantarum. 15, 473-497 (1962).
- Srinivasan, T., Kumar, K. R. R., Kirti, P. B. Establishment of efficient and rapid regeneration system for some diploid wild species of Arachis. Plant Cell Tissue and Organ Culture. 101, 303-309 (2010).
- Gomes, A. L. V., et al. Single-tube nested PCR using immobilized internal primers for the identification of dengue virus serotypes. Journal of Virology Methods. 145, 76-79 (2007).
- Williams, E. J., Drexler, J. S. A non-destructive method for determining peanut pod maturity. Peanut Science. 8, 134-141 (1981).
- Sobolev, V. S., Dorner, J. W. Cleanup procedure for determination of aflatoxins in major agricultural commodities by liquid chromatography. Journal of AOAC International. 85, 642-645 (2002).
- Empower Software, Getting Started Guide. , Waters Corporation. Milford, MA. Available from: http://sites.chem.colostate.edu/diverdi/C431/experiments/high%20pressure%20liquid%20chromatography/references/Empower%20getting%20started%2071500031203rA.pdf (2002).
- Biselli, S., Hartig, L., Wegner, H., Hummert, C. Analysis of Fusarium. toxins using LC-MS-MS: Application to various food and feed matrices. LC GC North America. 23, 404-413 (2005).
- Arias, R. S., Sobolev, V. S., Orner, V. A., Dang, P. M., Lamb, M. C. Potential involvement of Aspergillus flavus laccases in peanut invasion at low water potential. Plant Pathology. 63, 353-363 (2014).
- Dang, P. M., Chen, C. Y., Holbrook, C. C. Evaluation of five peanut (Arachis hypogaea) genotypes to identify drought responsive mechanisms utilising candidate-gene approach. Functional Plant Biology. 40, 1323-1333 (2013).
- Schmittgen, T. D., Livak, K. J. Analyzing real-time PCR data by the comparative C-T method. Nature Protocols. 3, 1101-1108 (2008).
- Amaike, S., Keller, N. P. Aspergillus flavus. Annual Review of Phytopathology. 49, 107-133 (2011).
- Nowara, D., et al. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell. 22, 3130-3141 (2010).
- Woloshuk, C. P., et al. Molecular characterization of aflR, a regulatory locus for aflatoxin biosynthesis. Applied and Environmental Microbiology. 60, 2408-2414 (1994).
- Price, M. S., et al. The aflatoxin pathway regulator AflR induces gene transcription inside and outside of the aflatoxin biosynthetic cluster. FEMS Microbiology Letters. 255, 275-279 (2006).
- Ehrlich, K. C., Montalbano, B. G., Cotty, P. J. Sequence comparison of aflR from different Aspergillus. species provides evidence for variability in regulation of aflatoxin production. Fungal Genetics and Biology. 38, 63-74 (2003).
- McDonald, T., Brown, D., Keller, N. P., Hammond, T. M. RNA silencing of mycotoxin production in Aspergillus and Fusarium species. Molecular Plant Microbe Interactions. 18, 539-545 (2005).
- Abdel-Hadi, A. M., Caley, D. P., Carter, D. R., Magan, N. Control of aflatoxin production of Aspergillus flavus. and Aspergillus parasiticus. using RNA silencing technology by targeting aflD. (nor-1) gene. Toxins (Basel). 3, 647-659 (2011).
- Swartz, M. E. Ultra performance liquid chromatography (UPLC): An introduction: Separation Science Redefined. LCGC North America. , Suppl ement 8. 8-14 (2005).
- Maghuly, F., Khan, M. A., Fernandez, E. B., Druart, P., Watillon, B., Laimer, M. Stress regulated expression of the GUS-marker gene (uidA) under the control of plant calmodulin and viral 35S promoters in a model fruit tree rootstock: Prunus incisa x serrula. Journal of Biotechnology. 135, 105-116 (2008).
- de Mesa, M. C., Santiago-Doménech, N., Pliego-Alfaro, F., Quesada, M. A., Mercado, J. A. The CaMV 35S promoter is highly active on floral organs and pollen of transgenic strawberry plants. Plant Cell Reports. 23, 32-38 (2004).
- Sunilkumar, G., Mohr, L., Lopata-Finch, E., Emani, C., Rathore, K. S. Developmental and tissue-specific expression of CaMV 35S promoter in cotton as revealed by GFP. Plant Molecular Biology. 50, 463-474 (2002).
- Groenenboom, M. A. C., Maree, A. F. M., Hogeweg, P. The RNA silencing pathway: The bits and pieces that matter. PLoS Computational Biology. 1, 155-165 (2005).
- Zhao, D., Song, G. Q. High-throughput sequencing as an effective approach in profiling small RNAs derived from a hairpin RNA expression vector in woody plants. Plant Science: an International Journal of Experimental Plant Biology. 228, 39-47 (2014).
- Kamthan, A., Chauduri, A., Kamthan, M., Datta, A. Small RNAs in plants: recent development and application for crop improvement. Frontiers in Plant Science. 6, 208(2015).
- Manda, A., Bodapati, P. N., Rachaputi, N. C., Wright, G., Fukai, S. Aflatoxins and their relationship with sugars in peanut (Arachis hypogaea L). 4th International Crop Science Congress, 2004, , Available from: http://www.cropscience.org.au/icsc2004/poster/5/1/3/625_manda.htm (2004).
- Uppala, S. S. Factors affecting pre-harvest aflatoxin contamination of peanut (Arachis hypogaea L). , Auburn University. (2011).
- Sobolev, V. S. Localized production of phytoalexins by peanut (Arachis hypogaea) kernels in response to invasion by Aspergillus species. Journal of Agricultural and Food Chemistry. 56, 1949-1954 (2008).
- Sobolev, V. S., Guo, B. Z., Holbrook, C. C., Lynch, R. E. Interrelationship of phytoalexin production and disease resistance in selected peanut genotypes. Journal of Agricultural and Food Chemistry. 55, 2195-2200 (2007).
- Sobolev, V. S., Neff, S. A., Gloer, J. B. New stilbenoids from peanut (Arachis hypogaea) seeds challenged by an Aspergillus caelatus strain. Journal of Agricultural and Food Chemistry. 57, 62-68 (2009).
- Dorner, J. W., Cole, R. J., Sanders, T. H., Blankenship, P. D. Interrelationship of kernel water activity, soil temperature, maturity, and phytoalexin production in preharvest aflatoxin contamination of drought-stressed peanuts. Mycopathologia. 105, 117-128 (1989).
- Sobolev, V. S. Production of phytoalexins in peanut (Arachis hypogaea) seed elicited by selected microorganisms. Journal of Agricultural and Food Chemistry. 61, 1850-1858 (2013).
- Basha, S. M. M., Cherry, J. P., Young, C. T. Changes in free amino acids, carbohydrates, and proteins of maturing seeds from various peanut (Arachis hypogaea L.) cultivars. Cereal Chemistry. 53, 586-596 (1976).
- Lansden, J. A. Aflatoxin inhibition and fungistasis by peanut tannins. Peanut Science. 9, 17-20 (1982).
- Yen, G. C., Duh, P. D., Tsai, C. L. Relationships between antioxidant activity and maturity of peanut hulls. Journal of Agricultural and Food Chemistry. 41, 67-70 (1993).
転載および許可
このJoVE論文のテキスト又は図を再利用するための許可を申請します
許可を申請さらに記事を探す
This article has been published
Video Coming Soon
Copyright © 2023 MyJoVE Corporation. All rights reserved