
Method Article
땅콩에서 아플라톡신의 RNAi를 매개 제어 : 방법은 땅콩에 곰팡이 독소 생산 및 형질 전환 유전자 발현을 분석 /
요약
우리는 균류 아스 퍼질러 flavus를 아플라톡신에 합성 유전자 사일런 위해 RNA 간섭 신호를 포함하는 땅콩 종자 아플라톡신 및 유전자 발현 분석을위한 방법을 보여준다. 식물에서 곰팡이 독소의 RNAi를 매개 제어는 이전에보고되지 않았습니다.
초록
국제 연합 식량 농업기구는 세계에서 식량 작물의 25 %는 아플라톡신에 오염되는 것으로 추정하고있다. 즉 음식의 100,000,000톤 파괴 또는 매년 인간이 아닌 소비로 전환되는 것을 나타냅니다. 아플라톡신은 일반적으로 곡물, 견과류, 뿌리 작물과 다른 농산물의 곰팡이 아스 페르 길 루스 flavus를하고 A를 parasiticus에 의해 축적 된 강력한 발암 물질이다. 땅콩 식물에서 RNA 간섭 (RNAi에)에 의해 다섯 아플라톡신 합성 유전자의 침묵은 A. 접종 다음 아플라톡신 축적을 제어하는 데 사용 된 flavus를. 이들은 보통 아플라톡신-공헌하는 조건 하에서 몇 종자 및 큰 필드 실험의 전통적인 제조 방법을 앞서, 어떠한 방법은, 개별 땅콩에서의 RNAi 형질 전환 이벤트의 효과를 분석하는 옵션이 존재하지 아니 하였다. 필드에서 자연적으로 오염 된 종자를 발견 할 확률은 1/1로 종종 1/100또한 000은, 아플라톡신의 오염이 균일하게 분포되지 않습니다. 우리의 방법은 실시간 PCR (RT-PCR)에 대한 처리 된 작은 조각 또는 작은 RNA 서열로, 형질 전환 이벤트 당 몇 종자를 사용하고, 초 고성능 액체 크로마토 그래피 (UPLC)에 의해 아플라톡신 축적 분석. RNAi의 발현 땅콩 라인 288-72와 288-74은 14,000 NG까지 축적 된 제어에 비해 아플라톡신 B 1의 100 % 감소 (p≤0.01)와 B 2로 나타났다. G -1 아플라톡신 B 1 aflatoxigenic A. 접종 flavus를. 참고로, 미국에서 인간의 소비에 대한 허용 아플라톡신의 최대 총 20 NG입니다. G -1. 이 프로토콜은 유전자 변형 땅콩 씨앗과 그 평가 방법에 아플라톡신의 RNAi를 매개 제어의 응용 프로그램을 설명합니다. 우리는 땅콩 및 기타 작물의 육종에의 응용 과학의 중요한 분야에서 급속한 발전을 가져올 것이라고 믿는다의학 및 인간의 영양은 상당히 중요한 농작물 아플라톡신 및 잠재적으로 다른 곰팡이 독소를 제어하는 국제적인 노력에 기여할 것이다.
서문
약 45억명은 만성적으로 아플라톡신 1, 자연 2에 알려진 가장 강력한 발암 물질에 노출되어있다. 이 곰팡이 독소는 옥수수, 카사바, 쌀, 견과류, 곡물, 향신료 등 세계 3에 식량 작물의 25 %를 오염. 4. 어린이 5 성장을 방해 원인 아플라톡신은 면역 체계 (6)을 손상 인간의 생체 검사 7, 8에서 간세포 - 암의 58 %에서 존재하고, aflatoxicosis 9,10 주기적으로 발생하는 동안 수백 명의 사람들을 죽인다. 아플라톡신은 일반적으로 아스 페르 길 루스 flavus를하고 A로 생산 된 폴리 케 타이드 파생 곰팡이 독소이다 parasiticus; 아플라톡신의 B 1, B 2는 A. 의해 일어난다 flavus를, A. 반면, parasiticus 또한 G 1 및 G 2 생산하고 있습니다. 이들 화합물 및 UPLC 의해 그들의 분리를 보여주는 크로마토 그램의 화학 구조는도 1에 도시된다.

그림 1. 아플라톡신 및 RNAi에 삽입 맨 :. 4 개의 가장 일반적인 폴리 케 타이드 파생 된 아플라톡신의 화학 구조 (왼쪽)와 (오른쪽) 크로마토 그램의 예 : B 1, B 2, G 1, G 2, 아스 페르 길 루스 parasiticus, 생산 .. flavus를은 B 1, B 2 바닥을 생산 : RNAi의 유전자 단편의 도식은 화살표 아래 번호는 아스 페르 길 루스 flavus를 게놈의 유전자 단편 가입 번호는, p5XCAPD 땅콩 변환에 사용되는 구성; PIV2 : 감자 인트론; BP : 염기쌍; RT_5X_1 및 RT_5X_2 :. 실시간 PCR 프라이머 사이트는 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
계산 4를 기반으로하는 경우로 인해 땅콩에서 아플라톡신 수출에 경제 손실이 단독으로 $ 450 만 달러를 초과 NG. G -1 유럽 연합 (EU) (11) 인간의 소비를 위해 허용 된 아플라톡신의 한계. 아플라톡신은 60년 12 알려져있다; 많은 농업 관행이 다른 곰팡이 균주 (13, 14)의 응용 프로그램을 포함하여 그 효과를 완화하기 위해 개발되었습니다 불구하고 그러나, 제어의 일관된 방법은 존재하지 않고, 저항하는 식물 품종을 사용할 수 없습니다. 심지어 병원균 침입에 대한 도움이되는 조건에서, 곰팡이 독소 축적이 예측할 수와 정규 분포를 따르지 않기 때문에 아플라톡신에 저항 테스트 식물 생식 질은 특히 어렵다. 따라서 실험은 보통 종자 및 100-1,700 g의 여러 샘플 수백 데이터 15,16의 변동성을 감소시키는 큰 재배 지역을 필요로한다.
RNA 간섭이었다1998 (17)에서 발견; 과 "침묵"의 장점은 현재 새로운 애플리케이션의 숫자, 예를 들어, 탐구되고있다. 전이성 유방암 (18), 간암 (19), 골수성 백혈병 (20), 곤충 (21)와 선충 (22)에 대한 식물 보호에 대 한 인간의 치료에. 식물에서 RNA 간섭 신호도 공장 호스트 (25)에 밀착되어 곰팡이 병원균 내부 23,24 입을 전신 전사 후 유전자에 대한 책임되는 작은 간섭 RNA (siRNA와)과 고 분자량 RNA와, 셀 사이를 이동할 수있다. 곰팡이 병원체 유전자의 식물 매개 침묵에의 RNAi의 효과가 이들에 대한 몇 가지 식물 pathosystems에 설명 된, 식물 (잎)의 공중 부분에서 증상의 육안 검사 양상추 (26)에 질병 정량화, 즉, 난균 강 Bremia 허용 밀에서 푸치니 (28)에 P> 27 푸사. 훨씬 더 어려운 잎, 씨앗 침략 장기 토양의 여러 인치 미만 감염의 증상을 보여주지로 땅콩 식물의 곰팡이 독소, 특히 아플라톡신을 제어하는 RNAi의 효과를 평가하는 것입니다, 감염의 발생은 예측할 수없는, 오직 화학 물질 분석은 아플라톡신의 존재를 확인할 수 있습니다. 또한, 땅콩의 각 형질 전환 이벤트는 일반적으로 몇 가지 씨앗 (공장 당 4-6)를 생산; 따라서, 큰 필드 플롯에 노 아플라톡신 축적 특성, 전체 자르기 시즌을 지속하고, 씨앗의 수백을 사용하는 기존의 테스트는 가능하지 않습니다. 방법 미만 1 주일 분석 여기에 설명되어 있습니다 만 몇 가지 씨앗을 사용하여 형질 전환 유전자의 존재와 무 아플라톡신 축적 특성에 대한 RNAi의 땅콩 씨앗.
프로토콜
1. 분자 구성하고 땅콩 변환
- 다섯 A.의 DNA 단편을 결합 flavus를 유전자, AFL2G_07223 (AFL이 또는 aflJ), AFL2G_07224 (aflR), AFL2G_07228 (AFLC / pksA / pksL1), AFL2G_07731 (pes1) 및 AFL2G_05027 (아플라톡신 유출 펌프, aflep). 이 내용은 다음 프라이머 및 ultramers를 사용 DIR-1, 짧은있는 Dir1-R, DIR-2 반전, 단기 Dir2라는-R, DirAll-NCO-, 계, 및 DirAll-BamEco-FW, 표 1.
- 제조업체에 따라 DNA 중합 효소를 사용하여 (94 ℃, 45 초 5 회, 55 ℃ 30 초, 68 ℃, 15 초 다음에, 95 ° C 2 분 25 μL 반응) 5 PCR 사이클에 의해 DIR-1 이중 가닥을 확인 지침 및 프라이머 짧은있는 Dir1-R은 3 '오버행 CCCGT을 떠날 수 있습니다. 이 단계를 반복 DIR-1에 상보적인 3 '오버행 ACGGG를 떠나 프라이머 짧은 Dir2라는-R을 사용하여 DIR-2 반전 이중 가닥을 확인합니다.
- 두 199 염기쌍의 fragme을 결찰제조업체의 지침에 따라 T4 DNA 리가와 NTS. PCR 1.1.1 이용한 프라이머 DirAll-CACC-FW 및 DirAll-NCO-, 계 (표 1)에 나타낸 바와 같이 얻어진 393 bp의 단편을 증폭하고, 플라스미드 P2 + 4ENTR 있도록 pENTR1A으로 표준 기술을 이용하여 제품을 복제.
- pCAPD (29)에 P2 + 4ENTR 재결합 : 플라스미드 p5XCAPD을 만드는 제조업체의 지시에 따라 LR clonase II 효소 믹스를 사용하여 (NCBI 수탁 KC176455.1), 및 부분 서열이어서 표준 기법을 이용하여 대장균 DH5α로 변환. 참고 : 완전한 RNAi의 인서트를 표 1에 나타낸다.
- 이전에 (30)를보고 플라스미드 p5XCAPD과 아그로 박테 리움 균주 C58C1 30 변환, 다음과 같이 땅콩 공장을 변환하는 결과 박테리아를 사용 :
- 50 ㎖ LB-국물 500 보충, 30 ° C p5XCAPD을 품고 아그로 박테 리움에서 성장이 사용μg의 ml의 -1 스트렙토 마이신, 25 μg의 ml의 -1 겐타 마이신, 10 μg의 ml의 -1 카나마이신, 1 OD (260)에 도달 할 때까지 250 rpm에서 문화를 흔들.
- 세균 현탁액 100 μM의 1 시간 동안 시린 및 장소 50 ㎖ AB 최소 배지 (31)에 10 분에 resuspend 원심 분리 (6,000 XG)에 의한 아그로 박테 리움 세포를 수확 10 ~ 14 일 이전 모종 Exp27-1516, 주자의 외식 형 땅콩 사육 라인. 얼룩은 3MM 30 분 후 압지에 외식을 건조하고, 차기 유도 매체 (SIM) MS 소금 (32), 3 %의 자당, 20 μM의 benzylaminopurine (BAP), 10 μM의 티 디아 주론 (TDZ), pH가 5.8, 0.3에 배치 3 일 동안 어둠 속에서 항생제없이 %의 젤란 검].
- 33 이전에보고 된 조직의 선택 및 재생 작업을 수행합니다. 2개월을 위해 격주로 전송하여, 촬영 형성을 위해 (500 μM의 세포 탁심 100 μm의 카나마이신을) SIM하기 위해 조직을 이동합니다. 그런 다음 장소촬영 - 신장 매체 (SEM)에 촬영 확대 [5 μM의 BAP, 1 μm의 gibberellic 산 (GA 3)], 몇 개월 동안 격주.
- 개별 쏘고, 2cm 크기, 뿌리 유도 배지 (RIM) 장소 1/2 MS, 1.5 % 수 크로스, 5 μM의 α - 나프탈렌 초산 (NAA)를 2.5 μM 인돌 부티르산 (IBA)], 다음 모종을 적응 및 온실로 전송.
침묵 아플라톡신 합성 유전자의 RNAi를 품고 땅콩 식물 (2) 확인
- RNAi의과 (전술 한 바와 같이) 200 μL 용출이 변화의 과정에 따라했다 땅콩 식물의 어린 잎에서 DNA를 추출하는 제조업체의 지침에 따라와 로봇 워크 스테이션에서 사용 공장 미니 키트는 백본으로이 p5XCAPD (그림 1) 구성 유전자 침묵에 대한 플라스미드 pCAPD 29.
- 설명 previ으로 PCR (STN-PCR) 중첩 된 하나의 튜브에 의해 DNA 샘플을 화면ously 34은 선별 마 아커 NPTII를 검출하고,이 RNAi의 p5XCAPD로부터 삽입한다. 클론 적이 첫 번째 세대에 테스트를위한 충분한 종자를 생산하는 컷 (3-4 노드) PCR 긍정적 인 식물에서 전파.
- DNA의 네 2 배 희석액을 사용합니다 (50 ~ 100 NG μL -1 희석 전) 모든 STN-PCR 반응에. 5'-AGGCTATTCGGCTATGACTG-3 '과 PCAPD 6446R : 5'-CGTCAAGAAGGCGATAGAAG-3'내부 프라이머 PCAPD 5730F : 5'-ACTGGGCACAACAGACAATC-3 '과 PCAPD 6249R : 5'-ATATTCGGCAAGCAGGCATC- NPTII를 들어, 외부 프라이머 PCAPD 5714F를 사용 3'.
- 5'-CCTAACAGAACTCGCCGTAA-3 '과 DirAll-NCO-, 계 : 5'-ATGCCATGGGGTTATTGGGTGCAGAATGG-3', 및 내부 Probe_5027_Fw 프라이머 : 5'-gtatttgtgaccatgtttctg -3- STN-PCR 반응에 삽입 RNAi를 검출하기위한 외부 프라이머 35S-PDSFw를 사용 '및 Probe_7228_Rv : 5'-GGACGGATAGTAAACTGCGG-3'.
- STN-PCR 긍정적 인 식물과 죄수의 수확 땅콩 포드 트롤 식물을 동일한 조건에서 재배. 중요한 단계 : 식물의 RNAi 식물과 동일한 조건과 동일한 절기에서 재배되는 제어를 확인합니다. 물 폭발 압력 세탁기와 포드 낮은 강도에서 35 (그림 (갈색과 검은 색 노란색, 오렌지색,) 그룹에서 성숙 보드와 별도의 포드 포드를 배치하여 mesocarp의 색상을, exocarp을 제거 결정하기 위해 손으로 긁어 2).

분석을위한 땅콩 포드 2. 준비 그림. 왼쪽 : 땅콩은 불확정 성장 식물로 수확에서 발견 된 다른 땅콩 크기; 센터 : exocarp의 수압 제거를위한 금속 바구니에 땅콩을 배치, 오른쪽 : 땅콩 프로필 보드 (노란색, 오렌지색, 갈색과 검은 색)에 mesocarp 색으로 성숙 그룹.98fig2large.jpg "대상 ="_ 빈 ">이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
3. 실험 설정
- 노란색 껍질, 프로세스를 제거하고 별도 갈색 씨앗. . 표 2에 따른 실험에서 사용하는 종자의 수를 계산 땅콩 라인과 샘플링 기간 당 세 시드 조각 (1 시드 조각 = 절반 자엽)의 최소가 통계 분석을 수행하고, 표준 오차를 줄이기 위해 필요합니다.
| 이름 | 순서 | ||
| DIR-1 | 5' GCCAGCTCAAAAGTGCGATGCACCAAGGAGAAACCGGCCTGTGCT CGGTGTATCGAACGTGGTCTTGCCTGTCAATACATGGTCGTATTTG TGACCATGTTTCTGGTGGCATTGGACCGTCTTGTCATCTCTACAGCC ATTCCCCAGATCACGGACGAATCCCCTGCATCTACGCGCACGCATC ACTTGGGGTACCCGT-3 ' | ||
| 짧은있는 Dir1-R | 5'-PHOS-TACCCCAAGTGATGCGTGCGCG-3 ' | ||
| DIR-2 반전 | 5' GGTTATTGGGTGCAGAATGGTAAACCACCCAACAGTACGCGAAATG TCAATTCCAGAGTCCCAAACCTCCCTACCGTGGCCTGGACGGATAG TAAACTGCGGAGCTTGGGAACAAAATCCGCTGTCTGATCGCCGAAG AGAAAGAGTTGCCTTGATTGAGCCGCATCGAGGACAGGTTGTGTTG CTGTTGATAGACGGG-3 ' | ||
| 짧은 Dir2라는-R | 5'-PHOS-CTATCAACAGCAACACAACC-3 ' | ||
| DirAll-CACC-FW | 5'-CACCGCCAGCTCAAAAGTGCGATGC-3 ' | ||
| DirAll-NCO-, 계 | 5'-ATGCCATGGGGTTATTGGGTGCAGAATGG-3 ' | ||
| DirAll-BamEco-FW | 5'-ATGGGATCCGAATTCGCCAGCTCAAAAGTGCGATGC-3 ' | ||
| 전체의 RNAi 삽입 | 5' GCCAGCTCAAAAGTGCGATGCACCAAGGAGAAACCGGCCTGTGCT CGGTGTATCGAACGTGGTCTTGCCTGTCAATACATGGTC / GTATTTG TGACCATGTTTCTGGTGGCATTGGACCGTCTTGTCATCTCTACAGCC ATTCCCCAGATCACGGACGAAT / CCCCTGCATCTACGCGCACGCAT CACTTGGGGTACCCGTCTATCAACAGCAACACAACCTGTCCTCGAT GCG / GCTCAATCAAGGCAACTCTTTCTCTTCGGCGATCAGACAGCG GATTTTGTTCCCAAGCTCCGCAGTTTACTATCCGTCCA / GGCCACGG TAGGGAGGTTTGGGACTCTGGAATTGACATTTCGCGTACTGTTGGG TGGTTTACCATTCTGCACCCAATAACC-3 ' | ||
. 표 RNAi의 p5XCAPD를 구성 PHOS를 구축하는 데 사용 1. 올리고 뉴클레오티드 및 ultramers는 5 '말단을 인산화; "/"는 다섯 유전자 단편이 사용 분리; 전체의 RNAi 삽입 : p5XCAPD를 형성하기 위해 2 역 반복으로 사용 순서.
- 그들은 멸균 비커의 바닥을 커버하도록 단일 층에 전체 땅콩 씨앗을 놓습니다. 씨앗을 포함하고 동일한 솔루션의 동일한 볼륨을 추가 75 % 에탄올 / 물 (v / v)의 솔루션을 추가합니다. 30 초 동안 상온에서 부화 한 후 멸균 탈 이온수 (SDW) 씻어.
- 다음과 같이 처리 된 종자의 에탄올을 함유하는 비이커에 2 % 차아 염소산을 추가 한 다음, 종자를 피복 동일한 용액의 동량을 추가 5 분 동안 배양 충분히 2 % 차아 염소산 용액을 추가한다. 중요한 단계 : SDW에 상당 체적 충분히 세번 씻어 사용 염소산의 부피 (도 3 5 배).

3. 실험 설정 그림. 탑 : 이전과 차아 염소산 후 땅콩 씨앗의 표면 살균, 종자 코팅 (종피)의 제거, 중동 : 배아 및 배아의 가까이보기의 제거는, 다음 반으로 자엽을 절단, 아래 : 반 자엽을 멸균 증류수에, 멸균 흡수에 블로 팅 종이, 물 한천과 한천 표면에 반 자엽의 배치 (절단 측 위쪽)에 움푹는. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
- 허용 표면 살균 씨는 2 시간 동안 SDW에 잠긴 마시다합니다. 멸균 페트리 접시에 씨앗을 놓고 집게와 씨앗 코트를 제거, 자엽을 분리하고, 메스와 배아를 제거합니다. 노트 :배아는 폐기되거나 새로운 식물을 재생하는데 사용될 수있다.
- 메스를 사용하여 절반의 각 자엽을 잘라. 탈수를 방지하기 위해, 모든 씨앗이 처리 될 때까지 멸균 물에 잘라 씨 부분을 유지.
- 멸균 한천 포함 된 배양 접시 준비했습니다 (1.5 % 한천 / 물 / W V), 각각의 세 가지 씨앗 조각 하나. 집게를 사용하여 한천의 작은 찌그러짐을하기 위해선, 물 / 한천 플레이트에 (위쪽 컷 얼굴)을 간략하게 멸균 종이 타월에 씨앗 조각의 여분의 물을 가릴하고 종자 조각을 배치합니다.
- 십일 25 ℃에서 성장을 Czapek의 한천 배지에 aflatoxigenic 아스 페르 길 루스 flavus를 NRRL 3357의 새로운 문화에서, 혈구로 계산 μL SDW 50,000 포자의 현탁액을 준비합니다.
- 포자의 RNAi (그림 4) 항구 시드 조직에 노출 얻을 확인하기 위해, 측면에 흐르는 빗물을 피하고 각 반 자엽 조각의 절단 표면에 포자 현탁액의 2 μL를 놓습니다.

아플라톡신 분석을위한 그림 4. 접종 및 배양. 탑 : 하프 자엽, 포자 현탁액으로 접종하고, 아스 페르 길 루스는 24 시간 배양 후 반 자엽에 균사 생장을 flavus를 바닥이 : 왼쪽 :. 48 시간의 배양을 1.5 % 한천에; 센터 : 1.5 % 한천에 72 시간 동안 배양; 오른쪽 :. 잘못된 실험 장치의 예, 0.5 % 한천에 72 시간 동안 배양 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
{kind=link}
- 샘플링 할 때까지 어둠 속에서 30 ℃에서 접종 및 비 접종 반 자엽을 포함하는 페트리 접시를 품어.
아플라톡신 분석 및 유전자 발현 4. 샘플링
- 에서 샘플을 수집각각 한 조각 (반 자엽) 인 복제, RT-PCR과 아플라톡신 분석을 위해 모두, 배양 24, 48 및 72 시간 (옵션 96 시간)에서 세중. 각 샘플링 날짜에 다른 판에서 무작위로 샘플을 선택합니다. 조직을 사용하여 부드럽게 유리 병 또는 테스트 튜브에 씨앗 조각을 배치하기 전에 한천과 초과 곰팡이 포자를 제거합니다.
- 아플라톡신 분석을 위해, -80 ° C에서 4 ㎖의 유리 스크류 캡 유리 병 및 상점의 각 복제 (한 조각)을 배치합니다. 참고 : 아플라톡신의 화학적 안정성을 감안할 때, 샘플 (현재 실험 보통 1-2개월에) 몇 개월 동안이 상태로 저장할 수 있습니다.
- RT-PCR의 장소를 이미 각 복제 (한 조각)이 스테인리스 구슬 (2.5 밀리미터 DIAM)와 세 지르코늄 구슬 (2mm의 DIAM)를 포함하는 튜브를 연마 2 mL로 준비했다.
- 즉시 모든 샘플 (바람직하게는 액체 질소) 동결 및 처리 할 때까지 -80 ℃에서 보관하십시오.
개별 절반 C 5. 아플라톡신 분석otyledon 조각
- A를 사용하여 flavus를이 분석을 위해 샘플을 접종. 약 30 분 동안 주위 온도에 샘플을 가져와 추가 4 권 (보통 2 ~ 3 ml의 W / V) 메탄올, (나중에 계산을위한 기록을 유지) 뚜껑을 닫고, O를 품어의 / N (~ 16 시간)에서 동요없이 어두운.
- , 일치하는 1.5 ml의 프로필렌 minicolumn에 프릿을 놓고 알 2 O 3 기본 200 밀리그램을 추가하고 이전 36 설명 된대로 다른 프릿으로 캡; 다음, 용출액의 가능한 증발을 방지하기 위해 충분히 가까이 열에서 초 고성능 액체 Chomatographer (UPLC) 오토 샘플러의 병을 배치합니다.
- 일회용 유리 시험관 (플라스틱을 사용하지 않는)에서, 단계 4.1에서 수득 한 메탄올 추출물 0.5 ml에 놓고 0.5 mL의 아세토 니트릴을 추가 피펫으로 혼합 단계 4.2에서 제조 컬럼으로 혼합물을 0.5ml의 적용. (압력을 적용하지 않음) 중력에 의해 자동 시료 주입기의 유리 병에 용출을 허용합니다. 용출은 일반적으로 2 ~ 4 분, 가까운 일 소요E 유리 병은 즉시 격막와 UPLC 호환 캡을 사용. 상온에서 튜브로 유지하고 UPLC에 당일들을 분석한다.
- 아플라톡신의 분리, 일치하는 UPLC 사기 용매 관리자, UPLC 샘플 관리자, UPLC 형광 검출기가 장착 된 UPLC 악기에 (A. flavus를 사용시 B 1, B 2) 아플라톡신 기준 샘플 용출액 및 자동 시료 주입기 튜브를 포함하는 장소 자동 시료 주입기 튜브, 와 C (18) 2.1 mm X 50mm, 1.7 μm의 열입니다.
- 0.30 mL의 분의 유속으로 물 / 메탄올 / CH 3 CN (64:23:13, V / V / V)의 혼합물로 이루어진 등용 매 이동상을 사용 -1. 제조업체의 지침 (37)에 따라, 아플라톡신 농도의 정확한 계산을위한 안정 기준 분리를 보장 크로마토 그램을 얻습니다.
- 자신의 질량 스펙트럼 데이터를 획득 및 게시 된 데이터 (38)와 비교하여 아플라톡신의 신원을 확인합니다. 내가 사용트랩 질량 분석계에 ESI 인터페이스와 제조자의 지시에 따라 대응하는 소프트웨어를 구비.
- 아플라톡신의 B 1, B 2의 상업적인 표준 대응 상이한 양을 주입하여 얻은 검량선을 참조하여 아플라톡신의 농도를 결정 G 1 및 G 2 UPLC 제조사가 제시 및 소프트웨어 (37)에 의해 측정.
- 장소 시드 조각들은 이미 건조 중량을 결정하는 O / N (~ 16 시간)의 동결 건조에 대한 개별 유리 바이알에, 메탄올로 추출 하였다. 그런 다음에 NG. 종자 조각의 건조 중량의 G -1 아플라톡신의 농도를 계산한다.
- 데이터의 분석을 위해 로그인 할 아플라톡신 결과를 변환 (NG. G -1 +1), 평균 비교를 위해 Tukey에의 테스트 하였다.
6. 유전자 발현, 샘플의 RT-PCR 처리
- 튜브가 포함 연마 2 mL를 취하여40 초 동안 3,100 rpm에서 비드 밀 균질에서 그들을 분쇄 (해동없이) 즉시 -80 ° C의 냉동고에서 샘플을 보내고, 그리고 다음 제조업체의 지침에 따라 트리 졸 사용하여 RNA 추출을 진행합니다.
- 는 1, 1 μg의 RNA를하고 올리고 DT 랜덤 헥사 같은 양을 사용하여 각 샘플에서의 cDNA를 준비합니다 cDNA를 8 희석하고 (이전 39 설명) RT-PCR의 반응 당 2 μL를 사용합니다.
- RNAi에 삽입 사용 프라이머의 발현 검출 : RT_5X_1_105F : 5'GGTGGCATTGGACCGTCTTG-3 ', RT_5X_1_232R : 5'-CGCATCGAGGACAGGTTGTG-3'; 및 RT_5X_2_95F : 5'-CCATGTTTCTGGTGGCATTG-3 ', RT_5X_2_229R : 5'-ATCGAGGACAGGTTGTGTTG-3'.
- 선별 마커의 발현 NPTII 검출에 사용 된 프라이머 : RT_NPTII_1_6871F : 5'-CTCGCTCGATGCGATGTTTC-3 ', RT_NPTII_1_7004R : 5'-GCAGGATCTCCTGTCATCTC-3'. 굴지-FW : 하우스 키핑 유전자 굴지 표준화 및 프라이머를 사용 CACATGCC를ATCCTTCGATTG; 액틴, 계 : CCAAGGCAACATATGCAAGCT 40.
- 굴지의 표현에 대한 표준화 델타 - 델타 C T 방법 (41)에 의해 결과를 분석합니다. 제어를 통해 배 증가 등의 결과를 나타냅니다.
결과
플라스미드는 p5XCAPD pCAPD 29의 유도체로 만들고, 땅콩 식물을 변환하는 데 사용되었다; 이 벡터는 A의 아플라톡신 합성 유전자의 다섯 작은 조각의 반전 반복, 70 ~ 80 bp의 각을 운반 인트론 (그림 1)로 구분 flavus를. AFL2G_07224 (aflR), AFL2G_07223 (AFL이 또는 aflJ), AFL2G_05027 (아플라톡신 유출 펌프, aflep), AFL2G_07228 (AFLC / pksA / pksL1), 및 AFL2G_07731 (pes1)의 조각에 그림 1 대응에 숫자, 구조를 사용 하였다 . BROAD 연구소, 캠브리지, MA, 그리고 문학 (42)에 flavus를 게놈 주석. 땅콩 99 라인의 총 변환 과정을 거쳐 재생하고, (50) PCR는 STN-PCR에 의해 검출 NPTII 양성이었고, 33 라인 PCR 양성 및 종자 생산 하였다. 단지 7 PCR 긍정적 인 줄은 클론 적 전파 및 presen와 테스트했다아플라톡신 축적 T 방법은 일곱 60 % 및 제어 100 %보다 적은 아플라톡신 축적 사이에 나타났다. 여기에서 우리는 그 일곱 줄의 두 가지의 결과를 보여줍니다. 각각의 형질 전환은 일반적으로 이벤트를 생성 몇몇 종자로서, 파라 메트릭 방법들은 여전히 통계적 분석을 수행 할 수있는 동안 씨의 최소 수를 사용하기 위해 개발되었다. 샘플 준비 및 실험 장치의 흐름도,도 5 및 표 1에 도시되어있다. 형질 전환 종자의 1 세대 전형적 hemizygous이지만, 이는이 세포 간 및 작은 간섭 RNA의 전신 운동 (의 siRNA)이 발생하는 것이 예상된다 RNA 간섭을 통해 공장 전체에 아플라톡신 합성 침묵을 부여해야합니다.

방법 그림 5. 도식 플로우 차트는 침묵에서의 RNAi의 효과를 분석 땅콩 종자에서 아스 페르 길 루스 아플라톡신 합성 유전자. 워크 플로우의 그래픽 표현 유전자 발현 또는 아플라톡신 분석을 위해 땅콩 샘플을 처리. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
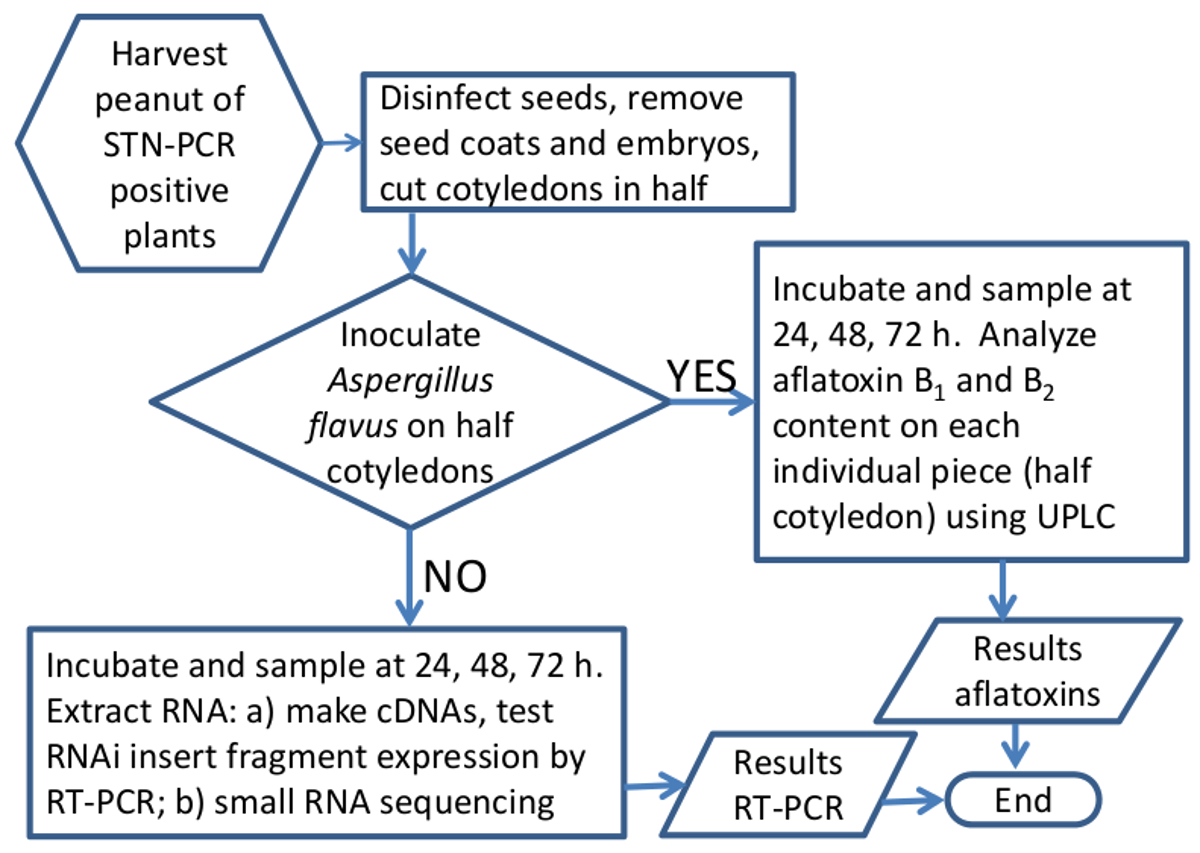{kind=link}
STN-PCR에 의해 시험했을 때의 RNAi 땅콩 라인 288-72와 288-74는 NPTII 선택할 메이커의 존재를 보여 주었다, 젤 섹션이 그림 6 (위)에 표시됩니다, 원본 사진 정보는 요청시 저자에서 사용할 수 있습니다. 이 두 유전자 CMR (클로람페니콜 저항)과 식물에 알 수없는 효과의 CcdB (독소)의 반전 반복을 암호화하기 때문에 플라스미드 pCAPD는 변화의 대조군으로 사용되지 않았다. 땅콩 PCR 네거티브 라인 2백88에서 9 사이 및 재생 공정을 통해 가서 RNAi의 라인과 동일한 조건에서 배양 된 다른 음의 콘으로 사용했다트롤.

. 그림 형질 전환 6. 탐지 및 RNAi를 실시간으로 표현 삽입 위 : 단일 튜브, 유전자 변형 땅콩 라인의 RNAi-288-72 및 RNAi의-288-74, 긍정적 인 식물의 중첩 된 PCR 검출. 컨트롤 : W (물), PP (긍정적 인 공장), MM (마스터 믹스), P (플라스미드 p5XCAPD); . 분수 / 2 4분의 1 팔분의 일 십육분의 일 1 DNA 하단의 2 배 희석을 나타냅니다 실시간 PCR은 RNAi의 발현을 검출 삽입 (프라이머 세트 : RT_5X_1, RT_5X_2을, 그림 1에서와 같이, 미성숙에 ( 노란색), 24 및 48 시간 배양에서 형질 전환 라인 (갈색) 자엽 성숙, 회색 라인 : C T = 1 히스토그램은 세 가지 생물학적 시료의 수단과 표준 오차 막대 (T)를 나타냅니다세 기술 복제와. RNAi의 삽입의 상대적 정량은 내부 통제 및 5.2.2 및 5.3에 설명 된대로 계산 된 형질 전환 유전자의 비교 배 표현식으로 하우스 키핑 유전자 굴지에 대한 표준화했다. 이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
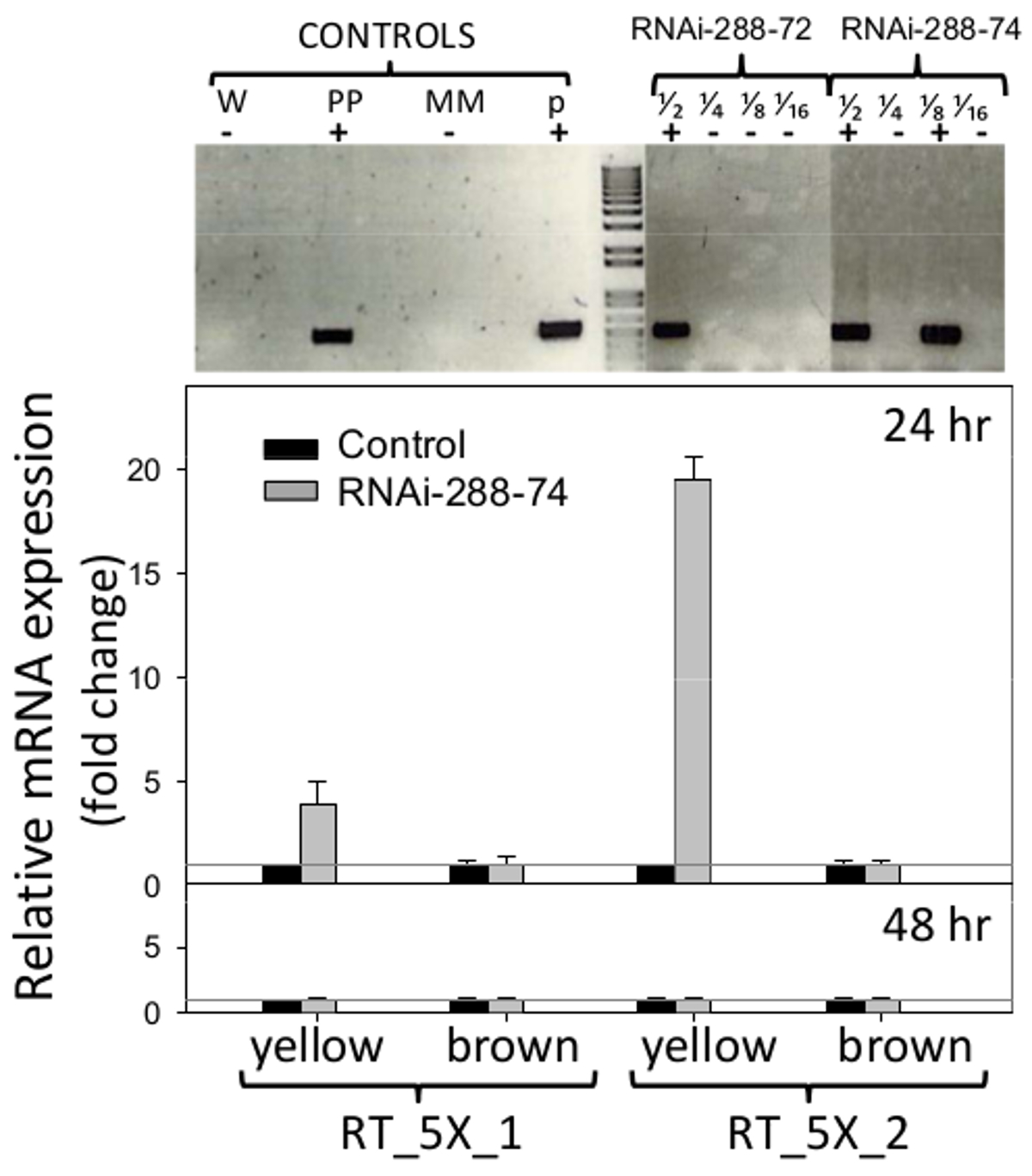{kind=link}
아플라톡신 축적 식물 숙주 RNAi를 매개 전위 제어의 효과를 시험하기 위해, 아스 퍼질러 flavus를 NRRL 3357의 갓 수확 분생는 배아 및 종피가 제거 된되는 반 자엽의 절단면에 도포 하였다 (도 3,도 4). 게놈 시퀀싱 및 p5XCAPD 설계를위한 기초이었다 된 대한 A. flavus를의 NRRL 3357은, 친절 USDA-ARS-NPRL 박사 혼에 의해 제공되었다. 30 ° C에서 24, 48 및 72 시간 후 절반 자엽의 결과 곰팡이 침공 쇼 N 그림 4. 접종 반 자엽의 샘플, 72, 48, 24에서 수집 된 배양 96 시간, 및 LC-MS에 의해 주 네 아플라톡신의 B 1, B 2, G 1 및 G 2 UPLC를 사용하여 분석 및 확인 ; 그 결과를도 6에 나타낸다. 아플라톡신 농도 수정 36 번역 방법을 사용하여 결정 하였다. 96 시간 배양에서, 자엽 때문에 곰팡이 감염의 붕괴 시작합니다. RNAi의 라인 288-72 미숙 자엽과 성숙한 사람에서 가장 샘플링 날짜에 모든 샘플링 날짜에 컨트롤보다 아플라톡신의 상당히 낮은 수준을 나타냈다. RNAi의 라인 288-74 대부분의 샘플링 날짜에 아플라톡신의 상당히 낮은 수준을 보였다. Tukey에의 테스트의 중요성의 수준은 그림 7의 그래픽에 별표로 표시됩니다.
g7.jpg "/>
. 그림 7. 아플라톡신은 Aspergillus flavus를 함께 배양 (24, 48, 72 및 96 시간) 후 반 땅콩 자엽에서 B 1, B 2 제어 : 288-9 비 형질 전환 라인의 씨앗; 의 RNAi : RNAi의-288-72 및 RNAi의-288-74, RNAi의 p5XCAPD에 대한 유전자 변형 종자 다섯 아플라톡신 합성 유전자를 침묵. (A) 성숙 종자에 아플라톡신 B 1 (갈색) (B) 아플라톡신 B (2) 성숙 종자; 미숙 씨 (노란색)와 미숙 씨의 (D) 아플라톡신 B 2 (C) 아플라톡신 B 1. 중복 된 생물학적 시료의 해당 표준 오차 막대 (T)와 평균 값이 표시됩니다. 통계적으로 유의 한 차이 Tukey에의 테스트 * : 0.05 ≤ P, ** : P ≤ 0.01, *** : P ≤ 0.001.S : //www.jove.com/files/ftp_upload/53398/53398fig7large.jpg "대상 ="_ 빈 ">이 그림의 더 큰 버전을 보려면 여기를 클릭하십시오.
전반적으로, RNAi의-288-72 실험 (24-96 시간 배양), 아플라톡신 B 2에서 94 % -100 % 감소, 제어에 비해 아플라톡신 B 1의 90 % -100 %의 감소를 통해 보여 주었다. RNAi의-288-74는 아플라톡신 B 1, 그림 7의 아플라톡신 B 2, 60 % -100 % 감소에서 63 % -100 %의 감소를 보였다.
RNAi의 인서트의 표현의 실시간 PCR 검출을 위해 사용 된 프라이머는도 1에 도시된다. 땅콩 종자 자엽은 자연 방어를 제거하고 가장 노출 영역에서의 RNAi의 잠재적 효과를 검출 할 수 있도록, 배아없이 분석했다 곰팡이 침입, 자엽. C T 위에 4 배는 RNAi의의 발현이 삽입 RT_5X_1 설정 프라이머로 라인 288-74의 미성숙 자엽 (노란색)에서 검출되었다 = 1 음성 대조군의 임계 값과 RT_5X_2 설정 프라이머로는 24 시간 배양에 모든 19 배 임계 값을 초과했습니다. 땅콩 변환에 사용 된 5 개의 연속 유전자 단편 중 적어도 세 가지가, 5027, 7223 및 7228는 (aflep가, AFL이는 / aflJ 및 AFLC / pksA, 각각이) RT-PCR로 검출되었다 (그림 1, 6). RNAi의 삽입의 발현은 24 시간에, 또는 48 시간 배양, 그림 6 (아래)에서 성숙 또는 미성숙 자엽에 성숙한 자엽에서 검출되지 않았다.
(1)| 땅콩 라인 | 샘플링 시간 | RT-PCR에 대한 샘플 | 아플라톡신 분석을위한 샘플 (접종) | 엔씨의 암갈색 | ||||
| (비 접종) | ||||||||
| 의원 1 | 담당자 2 | 의원 3 | 의원 1 | 담당자 2 | 의원 3 | |||
| RNAi의 (노란색) | 24 시간 | 1 | 1 | 1 | 1 | 1 | 1 | 4.5 |
| 48 시간 | 1 | 1 | 1 | 1 | 1 | |||
| 72 시간 | 1 | 1 | 1 | 1 | 1 | 1 | ||
| 제어 | 24 시간 | 1 | 1 | 1 | 1 | 1 | 1 | 4.5 |
| (황색) | 48 시간 | 1 | 1 | 1 | 1 | 1 | 1 | |
| 72 시간 | 1 | 1 | 1 | 1 | 1 | 1 | ||
표 2. 작은 샘플 설정의 예는 RNAi의 땅콩 종자의 유전자 발현 및 아플라톡신 축적을 분석합니다. 완전한 표현의 분석과 하나의 성숙 그룹에 대한 아플라톡신 (즉., 노란색), 세 샘플링 시간 (24, 48 및 72 시간)로 , 중으로, 4.5 씨앗을 요구, 테이블의 각 번호를 하나의 절반 자엽을 나타냅니다.
토론
식물 숙주 식물에서 곰팡이 독소 축적의 RNAi를 매개 제어의 가능성을 보여주는 더 출판물이없는, 곰팡이 병원균의 유전자의 침묵하지만, 27,43를 입증되었다의 RNAi는 매개. 잎은 지하 포드의 곰팡이 감염시 증상을 보여주지로 땅콩 이러한 연구에 대한 하나의 제한 요소는 개별 공장의 노 아플라톡신 축적 표현형을 평가하는 방법의 부족이었다. 또한, 아플라톡신의하지-정규 분포를 축적하고, 화학 분석 (15, 16)에 대한 큰 시료에 대한 필요성은 단일 공장에 잠재적 인 RNAi의 효과의 정량화를 방해했다. 여기에 제시된 방법은 세중 세 24 시간 간격 샘플링 (표 1, 그림 7)를 수행하기 위해 다섯 가지 씨앗을 사용하여 72 시간 실험으로 구성되어 있습니다. 종자 (100)보다 작지 G를 필요 전형적인 아플라톡신 분석에 비해, 우리의 방법은 individua 특히 적합처음 두 개 또는 세 개 이하 포드를 생산하지 땅콩 식물 L 트랜스 제닉 이벤트.
아플라톡신 합성 RNA를 매개 침묵은 유 전적으로 아스 페르 길 루스 flavus를하고 A를 변환하여 증명되었다 parasiticus. aflR는 A.에서 아플라톡신 생산 주 레귤레이터이므로 flavus를하고 A. parasiticus 44, 45, 그것은 식물에서 RNA를 매개 침묵에 대한 흥미로운 대상이됩니다. 그러나 aflR 유전 변이는 아스 페르 길 루스 종 (46) 사이에 도시되어 있고, 공장에서 생산 숙주 RNAi의 신호와 일치하는 완전한 서열이 존재하지 않는 경우 그 유전자 변이체 사일런 벗어날 수있다. 따라서, aflR 벡터 p5XCAPD에 침묵의 목표 중 하나였다,하지만 하나가 아니었다. A. 도입 aflR 유전자 역전 반복 flavus를하고 A. 변환에 의해 parasiticus는없이 또는 최소한의 제품을 침묵 결과아플라톡신 (47)의 이온 (맥도날드 등., 2005b). 또한, 침묵 aflD 유전자는 A에서 최대 98 %로 아플라톡신 생산을 방지 flavus를하고 A. 직접 변환 48 parasiticus. 우리의 시스템의 성공 확률을 높이기 위해, 땅콩은 A에서 아플라톡신 생산에 관여 다섯 유전자의 반전 반복 조각으로 변형되었다 flavus를. 여기가 아플라톡신 합성 경로에 여러 유전자를 침묵의 대상 p5XCAPD, 90 % 아플라톡신 B 1, B 2 -100 % 낮은 수준을 사용하는 것은 라인 288-72 달성 한 것으로 나타났다 및 60-100% 낮은 수준에 축적된다 반 자엽은 A. 접종 하였다 제어에 비해 라인 2백88에서 74 사이 flavus를, (7)은 4도. 가장 중요한 것은,이 방법은 파라 메트릭 statis을 적용하여 288-74 제어 대 실험을하는 동안 라인 288-72에 의해 아플라톡신 축적에 통계적으로 유의 한 차이를 발견틱,도 7. 작은 샘플 크기를 고려할 때, 이러한 실험은 높은 해상도, 높은 성능과보다 3 배 높은 민감도를 5 배를 갖는다 UPLC 분석 하였다 아플라톡신을 검출하는 강력한 방법을 사용하기위한 필요성을 강조하는 것이 중요 HPLC 49.
RNAi의의 발현 삽입 288-74 만 24 시간 배양에 미숙 한 자엽 (노란색)에서 검출되었다. RNAi의 삽입이 24 시간에서 288-74의 성숙 자엽에 RT-PCR에 의해 감지되지 않았거나 48 시간, 그림 6에 어떤 성숙 그룹에.이 같은 현상은 다른 RNAi의 유전자 변형 땅콩 라인에서 관찰되었다 (아리아, RS 2015 미 출판), 보통 RNAi의 성적 증명서 만 24 시간에서 미성숙 자엽에서 검출되었다한다. RNA 시료 cDNA 합성 전에 DNase를 처리하여, 데이터는 액틴 발현 수준과 관찰 DNA 오염의 증거로 표준화했다. DNA가 샘플에 출석해야한다, 그것은해야뿐만 아니라 48 시간 샘플에서 검출되지만 일관 즉 그렇지 않았다되었다. 35S 프로모터의 제어하에 발현이 항상 일정하지 않다; 그것은 환경 조건 (50), 조직 및 발달 단계 (51, 52)의 유형에 의해 영향을받을 수있다. 동시에, RNA 간섭의 통로에, mRNA의 감쇠율과의 siRNA 감쇠율이 크게 변할 수있다 (53). 이는 RNA 간섭 메카니즘에 의해 mRNA를 분해성 48 시간 배양에서 mRNA의 검출을 방지 할 수도 있다는 것이 가능하다. 48 시간에서 표현의 부재가 낮은 35S 발기인 중심의 전사 때문이든, 또는 다이 서 (Dicer)에 의해 dsRNA를 빠르게 분해에 대답 일이다. 따라서, 높은 처리량 시퀀싱에 의해 작은 RNA를 검출은 RNAi의 54를 통해 발생하는 프로세스에 대한 더 나은 통찰력을 줄 것이다 이 실험에서. 그러나, RNA 침묵은 주로 photosynt에서 체관부를 통해 체계적으로 확산 이후55 (이 경우 땅콩 씨앗) 싱크를 자당에, 아플라톡신 합성의 침묵이 삽입 RNAi의 로컬 표현하지 않고 씨앗에서 발생할 수있는 소스를 싫어. 많은 연구는 씨앗 아플라톡신 축적을 방지 할 필요가 작은 간섭 RNA는 (siRNA의)의 임계 레벨을 결정하기 위해 수행 될 남아있다. 이 RNAi의 양, mRNA 발현이 (도 6)를 구성한다는 사실을 강조하는 것이 중요하고, 아플라톡신의 B 1, B 2 (도 7)의 축적은 VS 미성숙 (황색)에 대한 다른 결과를 나타내었다. 성숙 (갈색) 자엽. 땅콩 식물 즉, 그들은 수확 성숙 포드의 범위, 그림 2에 제시, 불확실한 성장이있다. 또한, 다른 성숙 그룹의 씨앗, 그들의 화학 성분에 차이가 예., 2.4 %의 미성숙 종자에서 자당, 1.9 %에 같은 현장 조건 56,57에서 성숙한 종자. 따라서, 실제 efficienc을 이해아플라톡신 RNA 축적의 중재 제어 Y, 별도로 성숙 그룹을 분석하는 것이 중요하다.
땅콩 종자의 자연 방어 제조 화합물 및 종자 및 환경 조건 58-61의 만기에 따라 상대적인 양의 다양성으로 변화 phytoalexins의 제조이며, 자엽 (62)에 비해 배아에서 특히 높다. 배아는 (게시되지 않은 아리아스의 RS) 핵산, 자엽보다 모두 DNA와 RNA의 상당히 높은 농도를 가지고있다. 땅콩 씨앗이 성숙, 자신의 생리 및 화학 성분의 변화 (63)를 발생한다. 진균의 미생물 발육 저지 활동 (64)와 땅콩 종피의 형태로 응축 된 탄닌의 페놀 항산화 제; 타닌과 페놀 성 화합물의 함량은 성숙 (65)와 함께 증가로이 또한 35 검정색에 노란색 성숙 단계를 반영 mesocarp 색상에 분명하다. T의 따라서, 존재이들 항균성 부여 ESTA 또는 배아 실험은 진균 성장을 제한하기 때문에, 따라서, 그들은 제거하고, RNAi의 사일런 싱 효과를 과대 있었다. 또한, 종피 및 배아를 제거하여 배아 phytoalexins 더 많은 RNA 함량을 가질 것이다 운반 절반 떡잎 같이 분석 변동의 소스를 제한하는 데 도움이.
성숙 그룹과이 실험에서 종피와 배아를 제거하여 분석뿐만 아니라, 그것은 몇 가지 더 관찰을 지적하는 것이 중요하다 :) 결과는 최대 96 시간 보육에 대해 표시 되더라도을, 그것은 더 이상 72 이상을 사용하지 않는 것이 좋습니다 씨앗이 96 시간에 의해 분해되는 때 시간은 일관된 결과를 얻기 위해; 및 b) 같은 씨앗에서 반 자엽이 무작위로 샘플링하지만, 완벽하게 독립적 인 샘플을 구성하지 않는 반면, RT-PCR 및 형질 전환 이벤트 내에서 아플라톡신 축적 씨 사이에 최소한의 변화를 보여 주었다. 또한, 정확한 진균 포자는 접종 부피 카운트2 μL의 S 및 측면 상에 떨어지는 회피 자엽의 절단면에 포자 애플리케이션 포자 발아 식물 조직에 노출되어 있는지 확인하는 것이 중요하다. 접시에 물 / 한천은 그림 4 (하단)의 마지막 프레임과 같이 (V / W), 부드러운 한천은 포자의 유출을 야기 1.5 %에 있어야합니다. ; 특정 형질 전환 이벤트에서 제한의 유용성을 시드 경우, 샘플링 대신 세중 획득 유사한 결과 (즉, 그림 7) 중복으로 수행 할 수 있습니다 그러나, 세중의 샘플은 표준 오차를 줄이는 데 도움이됩니다. 이 방법의 유일한 제한은 아플라톡신 검출 / 정량 고감도 시스템 (UPLC)을 필요로하지만, 동시에이 덜 민감한 방법으로 아플라톡신 검출되지 않아야의 RNAi 효과를 과대 평가의 가능성을 감소시킨다.
결론적으로,이 방법은 처음을 연구하는 방법을 제공하는 신뢰할아플라톡신의 제어에서의 RNAi의 효과. 이하 일주에 전체 자르기 시즌에서 실험 시간을 단축,이 방법은 대단히 완화 및 / 또는 아플라톡신의 제거를 향해 RNAi의-땅콩 / 아스 페르 길 루스 pathosystem에 대한 연구를 가속화 할 것이다.
공개
The authors have nothing to disclose or any conflicts of interest.
감사의 말
This work received the financial support of USDA-ARS CRIS project 6604-21000-004-00D, CRIS project 6604-42000-008-00D, and USAID Feed-the-Future program Agreement number 58-0210-3-012. We thank Valerie Orner, LaTanya Johnson, Joseph Powell and Kathy Gray for their technical assistance. Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the US Department of Agriculture.
자료
| Name | Company | Catalog Number | Comments |
| Primers, oligonucleotides | DNA Technologies, Coralville, IA, USA | n/a | |
| Dneasy Plant Mini Kit | Qiagen, Valencia, CA | 69106 | |
| Czapek Dox agar medium | Oxoid, by Thermo Fisher Scientific, Waltham, MA | CM0095 | |
| Agar | Thermo Fisher Scientific, Waltham, MA | BP 1423 | |
| Freezer -80 °C | n/a | n/a | |
| Aluminum Oxide, Al2O3 | Fisher Scientific | A941 | |
| SPE Reservoirs 1.5 ml | Grace Davison Discovery Scientific | 210011 | |
| Frits for 1.5 ml SPE reservoir | Grace Davison Discovery Scientific | 211401 | |
| Autosampler vials | Waters Corporation, Milford, MA | 186005221 | |
| Waters Acquity Ultra-Performance Liquid-Chromatography (UPLC) instrument; UPLC-H-Class Quaternary Solvent Manager; UPLC Sample Manager; UPLC Fluorescent detector (FLR); UPLC BEH C18 2.1 mm x 50 mm, 1.7 mm column | Waters Corporation, Milford, MA | ||
| Finnigan LCQ Advantage MAX ion trap mass spectrometer, with Xcalibur version 1.4 software | Thermo Electron Corp., San Jose, CA | ||
| Aflatoxin standards, B1, B2, G1 and G2 | Sigma-Aldrich, St. Louis, MO | A6636; A9887; A0138; A0263 | |
| Systat Software 12.2 | SYSTAT Software Inc., Point Richmond, CA | ||
| Trizol reagent | Invitrogen, CA | 15596-018 | |
| SuperScript III First Strand Synthesis Super Mix | Invitrogen, CA | 11752-050 | |
| ABI 7500 Real-Time PCR | Lifetechnologies, Grand Island, NY | 4406984 | |
| Luria Broth-Miller | Fisher Scientific | R453642 | |
| pENTR1A | Invitrogen, CA | A10462 | |
| LR Clonase II enzyme mix | Invitrogen, CA | 11791-020 | |
| T4 DNA Ligase | NEB Biolabs | M0202L | |
| Gelrite | Sigma-Aldrich, St. Louis, MO | G1919 | |
| Acetosyringone | Sigma-Aldrich, St. Louis, MO | D134406 | |
| QIAcube robot workstation | Qiagen, Valencia, CA | 9001292 | |
| Antibiotics: kanamycin, cefotaxime, gentamicin; streptomycin | Goldbio, St. Louis, MO | cef.: C-104-25; kan: K-120-5; gent.: G-400-1; strep.: S-150-50 | |
| Platinum Taq DNA Polymerase High Fidelity | Invitrogen, CA | 11304-029 |
참고문헌
- Williams, J. H., et al. Human aflatoxicosis in developing countries: a review of toxicology, exposure, potential health consequences, and interventions. The American Journal of Clinical Nutrition. 80, 1106-1122 (2004).
- American Association for Cancer Research: AACR. An evaluation of chemicals and industrial processes associated with cancer in humans based on human and animal data: IARC Monographs Volumes 1 to 20. Cancer Research. 40, 1-12 (1980).
- Turner, P. C. The molecular epidemiology of chronic aflatoxin driven impaired child growth. Scientifica. , (2013).
- Rasooly, R., Hernlem, B., He, X., Friedman, M. Non-linear relationships between aflatoxin B1 levels and the biological response of monkey kidney vero cells. Toxins (Basel). 5, 1447-1461 (2013).
- Gong, Y. Y., et al. Determinants of aflatoxin exposure in young children from Benin and Togo, West Africa: the critical role of weaning. International Journal of Epidemiology. 32, 556-562 (2003).
- Eaton, D. L., Groopman, J. D. The toxicology of aflatoxins: human health, veterinary, and agricultural significance. , Academic Press. (1994).
- Murugavel, K. G., et al. Prevalence of aflatoxin B1 in liver biopsies of proven hepatocellular carcinoma in India determined by an in-house immunoperoxidase test. Journal of Medical Microbiology. 56, 1455-1459 (2007).
- Wang, J. S., et al. Hepatocellular carcinoma and aflatoxin exposure in Zhuqing Village, Fusui County, People's Republic of China. Cancer Epidemiology, Biomarkers & Prevention. 10, American Association for Cancer Research. 143-146 (2001).
- Azziz-Baumgartner, E., et al. Case-control study of an acute aflatoxicosis outbreak, Kenya, 2004. Environmental Health Perspectives. 113, 1779-1783 (2005).
- Lye, M. S., Ghazali, A. A., Mohan, J., Alwin, N., Nair, R. C. An outbreak of acute hepatic encephalopathy due to severe aflatoxicosis in Malaysia. American Journal of Tropical Medicine and Hygiene. 53, 68-72 (1995).
- Villers, P. Aflatoxins and safe storage. Frontiers in Microbiology. 5, 158(2014).
- Kensler, T. W., Roebuck, B. D., Wogan, G. N., Groopman, J. D. Aflatoxin: A 50-year odyssey of mechanistic and translational toxicology. Toxicological Sciences. 120, S28-S48 (2011).
- Dorner, J. W., Cole, R. J., Wicklow, D. T. Aflatoxin reduction in corn through field application of competitive fungi. Journal of Food Protection. 62, 650-656 (1999).
- Cotty, P. J., Bhatnagar, D. Variability among atoxigenic Aspergillus flavus strains in ability to prevent aflatoxin contamination and production of aflatoxin biosynthetic-pathway enzymes. Applied and Environmental Microbiology. 60, 2248-2251 (1994).
- Whitaker, T. B. Standardisation of mycotoxin sampling procedures: an urgent necessity. Food Control. 14, 233-237 (2003).
- Whitaker, T. B., Dorner, J. W., Giesbrecht, F. G., Slate, A. B. Variability among aflatoxin test results on runner peanuts harvested from small field plots. Peanut Science. 31, 59-63 (2004).
- Fire, A., et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 391, 806-811 (1998).
- Rafael, D., et al. EMT blockage strategies: Targeting Akt dependent mechanisms for breast cancer metastatic behaviour modulation. Current Gene Therapy. , (2015).
- Li, G., Chang, H., Zhai, Y. P., Xu, W. Targeted silencing of inhibitors of apoptosis proteins with siRNAs: a potential anti-cancer strategy for hepatocellular carcinoma. Asian Pacific. Journal of Cancer Prevention: APJCP. 14, 4943-4952 (2013).
- Koldehoff, M. Targeting bcr-abl transcripts with siRNAs in an imatinib-resistant chronic myeloid leukemia patient: challenges and future directions. Methods in Molecular Biology. 1218, 277-292 (2015).
- Zhang, J., et al. Pest control. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science. 347, 991-994 (2015).
- Ajjappala, H., Chung, H. Y., Sim, J. S., Choi, I., Hahn, B. S. Disruption of prefoldin-2 protein synthesis in root-knot nematodes via host-mediated gene silencing efficiently reduces nematode numbers and thus protects plants. Planta. 241, 773-787 (2015).
- Jose, A. M., Hunter, C. P. Transport of sequence-specific RNA interference information between cells. Annual Review of Genetics. 41, 305-330 (2007).
- Vazquez, F., Hohn, T. Biogenesis and biological activity of secondary siRNAs in plants. Scientifica. , Hindawi Publishing Corporation. (2013).
- Tinoco, M. L. P., Dias, B. B. A., Dall'Astta, R. C., Pamphile, J. A., Aragao, F. J. L. In vivo trans-specific gene silencing in fungal cells by in planta expression of a double-stranded RNA. BMC Biology. 8, (2010).
- Govindarajulu, M., Epstein, L., Wroblewski, T., Michelmore, R. W. Host-induced gene silencing inhibits the biotrophic pathogen causing downy mildew of lettuce. Plant Biotechnology Journal. , (2014).
- Yin, C., Jurgenson, J. E., Hulbert, S. H. Development of a host-induced RNAi system in the wheat stripe rust fungus Puccinia striiformis f. sp. tritici. Molecular Plant-Microbe Interactions. 24, 554-561 (2011).
- Ghag, S. B., Shekhawat, U. K., Ganapathi, T. R. Host-induced post-transcriptional hairpin RNA-mediated gene silencing of vital fungal genes confers efficient resistance against Fusarium. wilt in banana. Plant Biotechnology Journal. 12, 541-553 (2014).
- Filichkin, S. A., et al. Efficiency of gene silencing repeats vs. transitive RNAi in Arabidopsis: direct inverted vectors. Plant Biotechnology Journal. 5, 615-626 (2007).
- Sciaky, D., Montoya, A. L., Chilton, M. D. Fingerprints of Agrobacterium Ti Plasmids. Plasmid. 1, 238-253 (1978).
- Clark, D. J., Maaloe, O. DNA Replication and Division Cycle in Escherichia coli. Journal of Molecular Biology. 23, 99-112 (1967).
- Murashige, T., Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plantarum. 15, 473-497 (1962).
- Srinivasan, T., Kumar, K. R. R., Kirti, P. B. Establishment of efficient and rapid regeneration system for some diploid wild species of Arachis. Plant Cell Tissue and Organ Culture. 101, 303-309 (2010).
- Gomes, A. L. V., et al. Single-tube nested PCR using immobilized internal primers for the identification of dengue virus serotypes. Journal of Virology Methods. 145, 76-79 (2007).
- Williams, E. J., Drexler, J. S. A non-destructive method for determining peanut pod maturity. Peanut Science. 8, 134-141 (1981).
- Sobolev, V. S., Dorner, J. W. Cleanup procedure for determination of aflatoxins in major agricultural commodities by liquid chromatography. Journal of AOAC International. 85, 642-645 (2002).
- Empower Software, Getting Started Guide. , Waters Corporation. Milford, MA. Available from: http://sites.chem.colostate.edu/diverdi/C431/experiments/high%20pressure%20liquid%20chromatography/references/Empower%20getting%20started%2071500031203rA.pdf (2002).
- Biselli, S., Hartig, L., Wegner, H., Hummert, C. Analysis of Fusarium. toxins using LC-MS-MS: Application to various food and feed matrices. LC GC North America. 23, 404-413 (2005).
- Arias, R. S., Sobolev, V. S., Orner, V. A., Dang, P. M., Lamb, M. C. Potential involvement of Aspergillus flavus laccases in peanut invasion at low water potential. Plant Pathology. 63, 353-363 (2014).
- Dang, P. M., Chen, C. Y., Holbrook, C. C. Evaluation of five peanut (Arachis hypogaea) genotypes to identify drought responsive mechanisms utilising candidate-gene approach. Functional Plant Biology. 40, 1323-1333 (2013).
- Schmittgen, T. D., Livak, K. J. Analyzing real-time PCR data by the comparative C-T method. Nature Protocols. 3, 1101-1108 (2008).
- Amaike, S., Keller, N. P. Aspergillus flavus. Annual Review of Phytopathology. 49, 107-133 (2011).
- Nowara, D., et al. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell. 22, 3130-3141 (2010).
- Woloshuk, C. P., et al. Molecular characterization of aflR, a regulatory locus for aflatoxin biosynthesis. Applied and Environmental Microbiology. 60, 2408-2414 (1994).
- Price, M. S., et al. The aflatoxin pathway regulator AflR induces gene transcription inside and outside of the aflatoxin biosynthetic cluster. FEMS Microbiology Letters. 255, 275-279 (2006).
- Ehrlich, K. C., Montalbano, B. G., Cotty, P. J. Sequence comparison of aflR from different Aspergillus. species provides evidence for variability in regulation of aflatoxin production. Fungal Genetics and Biology. 38, 63-74 (2003).
- McDonald, T., Brown, D., Keller, N. P., Hammond, T. M. RNA silencing of mycotoxin production in Aspergillus and Fusarium species. Molecular Plant Microbe Interactions. 18, 539-545 (2005).
- Abdel-Hadi, A. M., Caley, D. P., Carter, D. R., Magan, N. Control of aflatoxin production of Aspergillus flavus. and Aspergillus parasiticus. using RNA silencing technology by targeting aflD. (nor-1) gene. Toxins (Basel). 3, 647-659 (2011).
- Swartz, M. E. Ultra performance liquid chromatography (UPLC): An introduction: Separation Science Redefined. LCGC North America. , Suppl ement 8. 8-14 (2005).
- Maghuly, F., Khan, M. A., Fernandez, E. B., Druart, P., Watillon, B., Laimer, M. Stress regulated expression of the GUS-marker gene (uidA) under the control of plant calmodulin and viral 35S promoters in a model fruit tree rootstock: Prunus incisa x serrula. Journal of Biotechnology. 135, 105-116 (2008).
- de Mesa, M. C., Santiago-Doménech, N., Pliego-Alfaro, F., Quesada, M. A., Mercado, J. A. The CaMV 35S promoter is highly active on floral organs and pollen of transgenic strawberry plants. Plant Cell Reports. 23, 32-38 (2004).
- Sunilkumar, G., Mohr, L., Lopata-Finch, E., Emani, C., Rathore, K. S. Developmental and tissue-specific expression of CaMV 35S promoter in cotton as revealed by GFP. Plant Molecular Biology. 50, 463-474 (2002).
- Groenenboom, M. A. C., Maree, A. F. M., Hogeweg, P. The RNA silencing pathway: The bits and pieces that matter. PLoS Computational Biology. 1, 155-165 (2005).
- Zhao, D., Song, G. Q. High-throughput sequencing as an effective approach in profiling small RNAs derived from a hairpin RNA expression vector in woody plants. Plant Science: an International Journal of Experimental Plant Biology. 228, 39-47 (2014).
- Kamthan, A., Chauduri, A., Kamthan, M., Datta, A. Small RNAs in plants: recent development and application for crop improvement. Frontiers in Plant Science. 6, 208(2015).
- Manda, A., Bodapati, P. N., Rachaputi, N. C., Wright, G., Fukai, S. Aflatoxins and their relationship with sugars in peanut (Arachis hypogaea L). 4th International Crop Science Congress, 2004, , Available from: http://www.cropscience.org.au/icsc2004/poster/5/1/3/625_manda.htm (2004).
- Uppala, S. S. Factors affecting pre-harvest aflatoxin contamination of peanut (Arachis hypogaea L). , Auburn University. (2011).
- Sobolev, V. S. Localized production of phytoalexins by peanut (Arachis hypogaea) kernels in response to invasion by Aspergillus species. Journal of Agricultural and Food Chemistry. 56, 1949-1954 (2008).
- Sobolev, V. S., Guo, B. Z., Holbrook, C. C., Lynch, R. E. Interrelationship of phytoalexin production and disease resistance in selected peanut genotypes. Journal of Agricultural and Food Chemistry. 55, 2195-2200 (2007).
- Sobolev, V. S., Neff, S. A., Gloer, J. B. New stilbenoids from peanut (Arachis hypogaea) seeds challenged by an Aspergillus caelatus strain. Journal of Agricultural and Food Chemistry. 57, 62-68 (2009).
- Dorner, J. W., Cole, R. J., Sanders, T. H., Blankenship, P. D. Interrelationship of kernel water activity, soil temperature, maturity, and phytoalexin production in preharvest aflatoxin contamination of drought-stressed peanuts. Mycopathologia. 105, 117-128 (1989).
- Sobolev, V. S. Production of phytoalexins in peanut (Arachis hypogaea) seed elicited by selected microorganisms. Journal of Agricultural and Food Chemistry. 61, 1850-1858 (2013).
- Basha, S. M. M., Cherry, J. P., Young, C. T. Changes in free amino acids, carbohydrates, and proteins of maturing seeds from various peanut (Arachis hypogaea L.) cultivars. Cereal Chemistry. 53, 586-596 (1976).
- Lansden, J. A. Aflatoxin inhibition and fungistasis by peanut tannins. Peanut Science. 9, 17-20 (1982).
- Yen, G. C., Duh, P. D., Tsai, C. L. Relationships between antioxidant activity and maturity of peanut hulls. Journal of Agricultural and Food Chemistry. 41, 67-70 (1993).
재인쇄 및 허가
JoVE'article의 텍스트 или 그림을 다시 사용하시려면 허가 살펴보기
허가 살펴보기This article has been published
Video Coming Soon
Copyright © 2025 MyJoVE Corporation. 판권 소유