
Method Article
РНК-интерференции-опосредованного управления афлатоксинов в арахисовом: метод анализа микотоксинов, производство и трансгенных выражение в Арахис /
В этой статье
Резюме
Мы демонстрируем метод для анализа афлатоксинов и экспрессии трансгена в семенах арахиса, которые содержат РНК-интерференции сигналов для глушителей генов афлатоксин-синтеза в гриба Aspergillus Flavus. RNAi-опосредованной контроль микотоксинов в растениях не сообщалось ранее.
Аннотация
Продовольственная и сельскохозяйственная организация Объединенных Наций оценивает, что 25% из продовольственных культур в мире заражены афлатоксинов. Это представляет 100 миллионов тонн продовольствия разрушается или направляются на потребление нечеловеческого каждый год. Афлатоксины мощные канцерогены, как правило, накопленные грибов Aspergillus Flavus и А. parasiticus в крупах, орехах, корнеплодах и других сельскохозяйственных продуктов. Подавление пяти генов афлатоксин-синтеза РНК интерференции (РНК-интерференции) в арахиса растений используется для управления накопление афлатоксина после прививки с А. Flavus. Ранее ни один метод не существовал, чтобы проанализировать эффективность РНК-интерференции в отдельных арахиса трансгенных событий, так как они, как правило, производят несколько семян и традиционных методов крупных полевых экспериментов в рамках афлатоксина-благоприятные условия были не вариант. В поле, вероятность нахождения естественно загрязненных семян часто 1/100 до 1/1,000. Кроме того, содержание афлатоксинов не распределены равномерно. Наш способ использует несколько трансгенных семян на событии, с небольшими кусочками, обработанных для ПЦР в реальном времени (RT-PCR) или небольшой РНК последовательности, и для анализа накопления афлатоксина методом жидкостной хроматографии ультра-производительности (UPLC). RNAi-экспрессирующих арахиса линии 288-72 288-74 и, показали до 100% снижения (p≤0.01) в афлатоксина В 1 и В 2 по сравнению с контролем, что накопленный до 14000 нг. Г -1 афлатоксина В1, когда засевают aflatoxigenic А. Flavus. Для справки, общий максимальный объем афлатоксинов допустимых для потребления человеком в Соединенных Штатах составляет 20 нг. Г -1. Этот протокол описывает применение РНК-интерференции-опосредованного контроля афлатоксинов в трансгенных семян и методы ее оценки арахиса. Мы считаем, что его применение в разведении арахиса и других культур принесет быстрое продвижение в этой важной области науки, Медицина и питание человека, и внесет значительный вклад в международные усилия, чтобы контролировать афлатоксины и потенциально другие микотоксины в основных продовольственных культур.
Введение
Примерно 4,5 миллиарда человек хронически подвергаются афлатоксинов 1, наиболее мощных канцерогенов, известных в природе 2. Эти микотоксины загрязняют 25% продовольственных культур в мире, в том числе 3 кукурузы, маниоки, риса, орехов, злаков и специй. 4. Афлатоксины причина задержки роста у детей 5, ослабляют иммунную систему 6, присутствуют в 58% гепатоцеллюлярной карциномы в-человека биопсии 7,8, и убить сотни людей во время периодических вспышек Афлатоксикоз 9,10. Афлатоксины поликетида полученных микотоксинов обычно производимые Aspergillus Flavus и А. parasiticus; афлатоксины В 1 и В 2 получают путем А. Flavus, в то время как А. parasiticus также производит G 1 и G 2. Химическая структура этих соединений и хроматограмме, показывая их разделение по UPLC показаны на рисунке 1.

Рисунок 1. Афлатоксины и РНК-интерференции вставить Топ:. Химическая структура (слева) и пример хроматограммы (справа) из четырех наиболее распространенных поликетида полученных афлатоксинов: В 1, В 2, С 1 и С 2, производится Aspergillus parasiticus, A . Flavus производит В 1 и В 2 Bottom: Схема генных фрагментов в РНК-интерференции построить p5XCAPD для арахиса трансформации, цифры под стрелы ген-фрагмент инвентарные номера в Flavus генома Aspergillus;. PIV2: картофель интрон; BP: пар оснований; RT_5X_1 и RT_5X_2:. Real-Time ПЦР праймеров сайты Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}
Экономические потери в экспорте из-за афлатоксинов в арахисе в одиночку превышать $ 450 млн долларов США, если рассчитывается на основе 4 нг. Г -1 предел афлатоксина разрешено для потребления человеком в Европейском союзе 11. Афлатоксины были известны в течение 60 лет 12; Однако, хотя многие сельскохозяйственные практики были разработаны, чтобы смягчить их влияние, в том числе применения других грибковых штаммов 13,14, либо последовательный способ управления не существует, и устойчивые сорта растений не доступны. Тестирование зародышевой плазмы растений на устойчивость к афлатоксинов особенно трудно, потому что даже при благоприятных условиях для патогена вторжения, микотоксинов накопление непредсказуемо и не соответствует нормальному распределению. Таким образом, эксперименты, как правило, требуют больших площадей посадки, сотни семян и нескольких образцов 100-1,700 г, чтобы уменьшить изменчивость 15,16 данных.
РНК-интерференция былаобнаружили в 1998 году 17; и преимущества "глушителей" В настоящее время изучаются в ряде новых приложений, например., в человеческих терапий против метастатическим раком молочной железы 18, рака печени 19, миелолейкоза 20, и в защите растений от насекомых и нематод 21 22. В растениях, сигналы помех РНК может путешествовать клетки к клетке, с малых интерферирующих РНК (миРНК) и высокой молекулярной массы РНК отвечать за системной посттранскрипционном гена глушителей 23,24, даже внутри грибковых патогенов, которые находятся в тесном контакте с растением-хозяином 25. Эффективность РНК-интерференции на завод-опосредованной молчания грибковых патогенов генов был описан в нескольких pathosystems растений, для них, визуальный осмотр симптомов в надземные части растения (листья) позволил количественного заболевания, т.е. оомицет Bremia в салат 26 , Puccinia в пшенице 27 и Fusarium в банан 28. Гораздо сложнее оценить RNAi эффективность для контроля микотоксинов в растениях, в частности афлатоксины в арахисе, как листья не показывают никаких признаков инфекции, органы вторглись (семена) находятся под несколькими дюймами почвы, возникновение инфекции является непредсказуемым, и только химический Анализ может определить наличие афлатоксинов. Кроме того, каждый трансгенной событие арахиса обычно производит несколько семян (4-6 на растение); Поэтому традиционные испытания на нет-афлатоксина накопления признака в больших полевых участках, прочного весь обрезки сезонов, и, используя сотни семян не представляется возможным. Способ описан здесь анализировать менее одной недели, РНК-интерференции семена арахиса для присутствия трансгена и для не-афлатоксина накопления признака, используя только несколько семян.
протокол
1. Молекулярная строительство и арахиса Преобразование
- Смешайте фрагменты ДНК из пяти A. Flavus гены, AFL2G_07223 (AFLS или aflJ), AFL2G_07224 (AFLR), AFL2G_07228 (aflC / pksA / pksL1), AFL2G_07731 (pes1) и AFL2G_05027 (афлатоксин отток насос, aflep). Для этого используйте следующие праймеры и ultramers: DIR-1, краткосрочный Dir1-R DIR-2-перевернутая, короткого Dir2-R, DirAll-Nco-RV, и DirAll-BamEco-FW, таблица 1.
- Сделать DIR-1 двойную нить на 5 циклов ПЦР (реакция 25 мкл, 95 ° C 2 мин, а затем 5 циклов при 94 ° С 45 сек, 55 ° С 30 сек, 68 ° С 15 сек) с использованием ДНК-полимеразы в соответствии с производителем инструкции и грунтовки короткого Dir1-R, чтобы оставить 3 'навеса CCCGT. Повторите эти шаги, чтобы сделать DIR-2-перевернутую двойную нить с использованием праймера Короткое DIR2-R, чтобы оставить 3 'навеса ACGGG комплементарную DIR-1.
- Перевязывать двух 199 б.п. fragmeНТС с Т4 ДНК-лигазы в соответствии с инструкциями изготовителя. ПЦР-амплификации полученный фрагмент 393 п.н., как указано в 1.1.1, используя праймеры DirAll-CACC-FW и DirAll-NCO-RV (Таблица 1), и клонировать продукт с использованием стандартных методик в pENTR1A сделать плазмиды P2 + 4ENTR.
- Рекомбинируют P2 + 4ENTR в pCAPD 29 (NCBI Присоединение: KC176455.1) с помощью LR Clonase II смеси ферментов в соответствии с инструкциями завода-изготовителя, чтобы сделать плазмиды p5XCAPD, и превратить его в кишечной палочки DH5 & alpha с использованием стандартных методов с последующим частичным последовательности. Примечание: В комплект RNAi вставка показано в таблице 1.
- Transform Agrobacterium штамм C58C1 30 плазмидой p5XCAPD, как сообщалось ранее 30, и использовать полученный бактерии превратить арахиса растений следующим образом:
- Расти 30 ° С Agrobacterium, несущие p5XCAPD, использовать для этого, 50 мл LB-бульона с добавлением 500мкг мл -1 стрептомицин, 25 мкг мл -1 гентамицин, 10 мкг мл -1 канамицин, и не трясти культуры на 250 оборотов в минуту до достижения OD 260 1.
- Урожай Agrobacterium клетки центрифугированием (6000 XG) в течение 10 мин, ресуспендируют в 50 мл среды АБ минимальной 31 100 мкМ ацетосирингона в течение 1 ч, и места в бактериальной суспензии эксплантов из 10-14-дневных проростков Exp27-1516, бегун типа арахиса линии разведения. Промокните насухо эксплантов на 3 мм промокательной бумаги после 30 мин, и разместить их на съемки индукции среды (SIM) [MS соли 32, 3% сахарозы, 20 мкМ бензиламинопурина (БАТ), 10 мкМ тидиазурон (ТДЗ), рН 5,8, 0,3 % геллановая резинки] без антибиотиков в темноте в течение трех дней.
- У выбор тканей и регенерации, как сообщалось ранее 33. Перемещение ткани в SIM (500 мкмоль цефотаксим и 100 мкм) канамицин для формирования побегов, с би-еженедельно переводов в течение 2 месяцев. Затем месторасширение побеги на стрелять удлинения среды (SEM) [5 мкМ БАП, 1 мкМ гибберелловой кислота (ГК 3)], два раза в неделю в течение нескольких месяцев.
- Место отдельных побегов, 2 см в размерах, в корневой-индукционной среде (RIM) [1/2 MS, 1,5% сахарозы, 5 мкМ α-нафталин-уксусной кислоты (НУК), 2,5 мкМ индол-масл ную кислоту (IBA)], затем акклиматизироваться рассаду и передавать их в теплицу.
2. Идентификация арахиса растений, несущих RNAi, чтобы заставить замолчать афлатоксин синтеза генов
- Использование растений мини комплект в рабочей станции робота с 200 мкл элюции в соответствии с инструкциями завода-изготовителя для извлечения ДНК из молодых листьев арахиса растений, которые подлежат процессе трансформации (как описано выше) с РНК-интерференции построить p5XCAPD (рисунок 1), который имеет, как позвоночника плазмиды pCAPD 29 для генной глушителей.
- Экран образцы ДНК от одного-трубки, вложенной ПЦР (СТН-ПЦР), как описано previменно 34 для обнаружения селективного маркера, NPTII и РНК-интерференции вставить из p5XCAPD. Клонально распространяться от порезов (3-4 узлов) ПЦР положительных растений производить достаточное семена для тестирования в первом поколении.
- Используйте четыре 2-кратного разведения ДНК (50-100 нг мкл -1 перед разбавлением) во всех реакций СТН-ПЦР. Для NPTII, использовать внешние праймеры PCAPD 5714F: 5'-AGGCTATTCGGCTATGACTG-3 'и PCAPD 6446R: 5'-CGTCAAGAAGGCGATAGAAG-3' и внутренние праймеры PCAPD 5730F: 5'-ACTGGGCACAACAGACAATC-3 'и PCAPD 6249R: 5'-ATATTCGGCAAGCAGGCATC- 3 '.
- Для обнаружения RNAi вставить в STN-ПЦР-реакций внешних праймеров используют 35S-PDSFw: 5'-CCTAACAGAACTCGCCGTAA-3 'и DirAll-NCO-Rv: 5'-ATGCCATGGGGTTATTGGGTGCAGAATGG-3', и внутренние праймеры Probe_5027_Fw: 5'-gtatttgtgaccatgtttctg-3 'и Probe_7228_Rv: 5'-GGACGGATAGTAAACTGCGG-3'.
- Урожай арахиса стручки от STN-ПЦР положительные растений и кондитер TROL растений, выращенных в тех же условиях. Важным шагом: Убедитесь, что контроль растения, выращенные в тех же условиях и того же сезона, как РНК-интерференции растений. Вода взрыв стручки с шайбой давления при низкой интенсивности или очистить вручную удалить внеплодник, определить цвет мезокарпий путем размещения стручки на погашения платы и отдельных стручков в группах (желтый, оранжевый, коричневый и черный) 35 (рис 2).

Рисунок 2. Подготовка арахиса стручки для анализа. Слева: Различные размеры арахиса найдено во время уборки урожая, как арахис неопределенный рост растений; Центр: размещение арахис в металлическом корзину для удаления под давлением воды внеплодник; справа: погашения группы по мезокарпий цвета на арахисовое профильной доски (желтый, оранжевый, коричневый и черный).98fig2large.jpg "целевых =" _blank "> Нажмите здесь, чтобы посмотреть большую версию этой фигуры.
3. Экспериментальная установка
- Удалить корпусов, процесс желтый и коричневый семена отдельно. Рассчитать количество семян для использования в эксперименте, согласно таблице 2. Обратите внимание, что как минимум три семенных штук (1 семя штучных = половина семядольных) в арахисовом линии и даты отбора проб требуется для выполнения статистического анализа и уменьшить стандартную ошибку.
| имя | Последовательность | ||
| DIR-1 | 5' GCCAGCTCAAAAGTGCGATGCACCAAGGAGAAACCGGCCTGTGCT CGGTGTATCGAACGTGGTCTTGCCTGTCAATACATGGTCGTATTTG TGACCATGTTTCTGGTGGCATTGGACCGTCTTGTCATCTCTACAGCC ATTCCCCAGATCACGGACGAATCCCCTGCATCTACGCGCACGCATC ACTTGGGGTACCCGT-3 " | ||
| Краткосрочные Dir1-R | 5'-Фос-TACCCCAAGTGATGCGTGCGCG-3 ' | ||
| DIR-2-инвертированный | 5'GGTTATTGGGTGCAGAATGGTAAACCACCCAACAGTACGCGAAATG TCAATTCCAGAGTCCCAAACCTCCCTACCGTGGCCTGGACGGATAG TAAACTGCGGAGCTTGGGAACAAAATCCGCTGTCTGATCGCCGAAG AGAAAGAGTTGCCTTGATTGAGCCGCATCGAGGACAGGTTGTGTTG CTGTTGATAGACGGG-3 " | ||
| Краткосрочные Dir2-R | 5'-Фос-CTATCAACAGCAACACAACC-3 ' | ||
| DirAll-CACC-FW | 5'-CACCGCCAGCTCAAAAGTGCGATGC-3 " | ||
| DirAll-Nco-RV | 5'-ATGCCATGGGGTTATTGGGTGCAGAATGG-3 ' | ||
| DirAll-BamEco-FW | 5'-ATGGGATCCGAATTCGCCAGCTCAAAAGTGCGATGC-3 ' | ||
| Полный РНК-интерференции вставка | 5'GCCAGCTCAAAAGTGCGATGCACCAAGGAGAAACCGGCCTGTGCT CGGTGTATCGAACGTGGTCTTGCCTGTCAATACATGGTC / GTATTTG TGACCATGTTTCTGGTGGCATTGGACCGTCTTGTCATCTCTACAGCC ATTCCCCAGATCACGGACGAAT / CCCCTGCATCTACGCGCACGCAT CACTTGGGGTACCCGTCTATCAACAGCAACACAACCTGTCCTCGAT GCG / GCTCAATCAAGGCAACTCTTTCTCTTCGGCGATCAGACAGCG GATTTTGTTCCCAAGCTCCGCAGTTTACTATCCGTCCA / GGCCACGG TAGGGAGGTTTGGGACTCTGGAATTGACATTTCGCGTACTGTTGGG TGGTTTACCATTCTGCACCCAATAACC-3 " | ||
. Таблица 1. Олигонуклеотиды и ultramers, используемые для построения построить p5XCAPD Phos RNAi: фосфорилируется 5 'конец; "/" Отделяет фрагменты генов пять использоваться; Полное вставки РНК-интерференции: последовательность используется в качестве 2 инвертированных повторов, чтобы сформировать p5XCAPD.
- Поместите целые семена арахиса в один слой так, что они охватывают дно стерильной стакан. Добавить 75% раствора этанол / вода (о / о), чтобы покрыть семена, а затем добавить равный объем того же раствора. Выдержите при комнатной температуре в течение 30 сек, а затем промыть стерильной деионизированной воды (ТБО).
- Добавить 2% гипохлорита в химический стакан, содержащий этанол обрабатывают семена следующим образом: добавить достаточно 2% раствор гипохлорита для покрытия семян, а затем добавить равный объем того же раствора и инкубируют в течение 5 мин. Важным шагом: тщательно промыть три раза с объемом ТБО, эквивалентной 5 раз объем гипохлорита используется (рисунок 3).

Рисунок 3. Настройка экспериментальный. Вверху: стерилизации поверхности семян арахиса до и после гипохлорита, удаление семенных оболочек (Testa); Средний: удаление эмбриона и тесного зрения эмбриона, а затем вырезать семядолей в два раза, а нижняя половина семядоли в стерильной дистиллированной воде, промокательной на стерильную абсорбента бумага, вмятины на воде агар, и размещения половиной семядолей (разрезанной стороной вверх) на поверхности агара. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}
- Позволить поверхность стерилизуют семена впитывать погружен в ТБО в течение 2 ч. Место семена на стерильную чашку Петри, удалить семенных оболочек щипцами, отделить семядоли, и скальпелем удалить эмбрионов. Заметка:Эмбрионы могут быть отброшены или используется для регенерации новых заводов.
- Разрежьте каждый семядолей в половине, используя скальпель. Чтобы избежать обезвоживания, не держать вырезать семенные штук в стерильной воде, пока все семена обрабатываются.
- Были подготовлены чашки Петри, содержащие агар стерильной водой (1,5% агар / воды; вес / объем), по одному для каждого трех семенных штук. Сделать небольшие вмятины в агар с использованием щипцов, кратко промокните излишки воды семенных штук на стерильных бумажных полотенец, а затем поместить семена штук (вырезать лицо вверх) на водных / пластин агара.
- Из свежей культуры aflatoxigenic Aspergillus Flavus NRRL 3357 в агаровой среде Чапека, выращенных при 25 ° С в течение 10 дней, приготовить суспензию 50000 спор на мкл ТБО, подсчитанных с помощью гемоцитометра.
- Поместите 2 мкл суспензии спор на срезанных поверхностей каждого наполовину семядолей кусок избегая сток по бокам, чтобы убедиться, что споры получить воздействию семян ткани, таит RNAi (рисунок 4).

Рисунок 4. Прививка и инкубации для анализа афлатоксина. Топ: Половина семядолей, прививка с суспензией спор, и Aspergillus Flavus роста мицелия на половину семядолей после 24 ч инкубации Внизу: слева. Инкубацию 48 часов на 1,5% агара; центр: инкубационный в течение 72 ч на 1,5% агара; справа:. пример некорректного экспериментальной установки, инкубирование в течение 72 ч на 0,5% агара Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}
- Не Инкубируйте чашки Петри, содержащие инокулированных и не засеянные половиной семядоли при 30 ° С в темноте до тех пор, выборки.
4. Отбор проб для анализа и афлатоксина экспрессии генов
- Соберите образцы втройные на 24, 48 и 72 ч (по желанию) 96 ч инкубации, как для ОТ-ПЦР и анализа афлатоксина, каждый репликации быть одна часть (половина семядолей). Выберите выборку образцов из различных пластин на каждую дату отбора проб. Использование ткани, мягко удалить избыток агара и грибковые споры, прежде чем делать заказ семян штук в флаконах или пробирках.
- Для анализа афлатоксина, место каждой повторности (один кусок) в 4 мл стеклянный флакон с завинчивающейся пробкой и хранить при температуре -80 ° C. Примечание: Принимая во внимание химическую стабильность афлатоксинов, образцы могут храниться в этом состоянии в течение нескольких месяцев (в текущих экспериментов обычно 1-2 месяцев).
- Для RT-PCR месте каждой повторности (одна часть) в уже подготовлен 2 мл шлифования пробирки, содержащие два борта из нержавеющей стали (2,5 мм в диаметре) и три циркония бусы (2 мм в диаметре).
- Сразу заморозить (предпочтительно в жидком азоте) всех образцов, и держать их при температуре -80 ° С до обработки.
5. Афлатоксин анализ отдельных Half Сotyledon шт
- Используйте А. Flavus прививку образцы для анализа. Принести образцы до комнатной температуры в течение 30 мин, добавляют четыре тома (обычно 2-3 мл; вес / объем) метанола (вести учет для дальнейших расчетах), закрыть крышки, и инкубируют O / N (~ 16 часов) в темно без перемешивания.
- Поставьте фритты в сопрягая 1,5 мл пропилена миниколонку, добавить 200 мг основную Al 2 O 3 и довершение с другой шведской как описано ранее 36; Затем поместите Ультра-высокоэффективной жидкостной Chomatographer (UPLC) AutoSampler флакон под колонкой достаточно близко, чтобы избежать возможного испарения элюата.
- В одноразового стеклянную пробирку (не использовать пластик), поместить 0,5 мл метанольного экстракта, полученного на стадии 4.1, 0.5 добавить мл ацетонитрила, перемешивают с помощью пипетки и применять 0,5 мл смеси в колонну, полученного на стадии 4.2. Разрешить элюирование в автосамплера флакон под действием силы тяжести (не давить). Элюирование обычно занимает 2-4 мин, рядом тысе флакон сразу с помощью совместимого крышку UPLC с перегородками. Держите флаконов при температуре окружающей среды и анализировать их в тот же день на UPLC.
- Для разделения афлатоксинов, место AutoSampler флаконы, содержащие образцы элюаты и AutoSampler флаконов со стандартами афлатоксина (В 1 и В 2 при использовании A. Flavus) в прибора UPLC оборудованной с соответствующим UPLC четвертичного растворителя Manager, UPLC Образец Manager, UPLC Люминесцентная детектор, и C 18 2.1 мм х 50 мм, 1,7 мкм колонка.
- Использование изократический подвижной фазы, состоящей из воды / МеОН / CH 3 CN (64:23:13, объем / объем / объем) смеси при скорости потока 0,30 мл мин -1. Получить хроматограммы обеспечивая стабилизированную базовой разделение для точного расчета концентрации афлатоксина, в соответствии с инструкциями изготовителя 37.
- Подтвердите личность афлатоксинов путем получения их данные масс-спектрального и сравнивая их с опубликованными данными 38. Используйте Iна ловушки масс-спектрометра оснащен интерфейсом ESI и соответствующего программного обеспечения в соответствии с инструкциями изготовителя.
- Определение концентрации афлатоксинов со ссылкой на калибровочных кривых, полученных путем введения различных количеств соответствующих коммерческих стандартов афлатоксинов В 1, В 2, С 1 и С 2, как предложено производителем UPLC и определяется с помощью программного обеспечения 37.
- Место семена штук уже экстрагировали метанолом, в отдельных стеклянных флаконах для O / N (~ 16 ч) лиофилизации, чтобы определить их сухой вес. Затем рассчитать концентрацию афлатоксина в нг. Г -1 сухого веса семян кусок.
- Для анализа данных, конвертировать результаты афлатоксина войти (нг. Г -1 +1), с последующим тестом Тьюки для средних сравнений.
6. Экспрессия гена, ОТ-ПЦР Обработка образцов
- Возьмите 2 мл шлифовальные трубки содержатING образцы из -80 ° C морозильнике, и сразу же (без размораживания) растереть их в шаровой мельнице гомогенизаторе при 3100 оборотах в минуту в течение 40 сек, затем переходите к экстракции РНК с помощью Trizol в соответствии с инструкциями изготовителя.
- Готовят кДНК из каждого образца с использованием 1 мкг РНК и равное количество олиго дТ и случайных гексамеров, чтобы в разведении 1: 8 кДНК и 2 мкл используют на реакцию ОТ-ПЦР (как описано ранее 39).
- Для обнаружения экспрессии РНК-интерференции использование вставки праймеров: RT_5X_1_105F: 5'GGTGGCATTGGACCGTCTTG-3 ', RT_5X_1_232R: 5'-CGCATCGAGGACAGGTTGTG-3'; и RT_5X_2_95F: 5'-CCATGTTTCTGGTGGCATTG-3 ', RT_5X_2_229R: 5'-ATCGAGGACAGGTTGTGTTG-3'.
- Для обнаружения экспрессии селектируемого маркерного NPTII, использование праймеров: RT_NPTII_1_6871F: 5'-CTCGCTCGATGCGATGTTTC-3 ', RT_NPTII_1_7004R: 5'-GCAGGATCTCCTGTCATCTC-3'. Используйте ген уборка актина по стандартизации, и праймеров: актин-Fw: CACATGCCATCCTTCGATTG; Актин-RV: CCAAGGCAACATATGCAAGCT 40.
- Анализ результатов от Дельта-дельта метода С Т 41 стандартизирована для выражения актина. Представлять результаты как кратное увеличение по сравнению с контролем.
Результаты
Плазмиды p5XCAPD было сделано как производное pCAPD 29, и использовали его для преобразования арахиса растений; этот вектор несет инвертированные повторы пяти небольших фрагментов, каждый 70-80 б.п., генов афлатоксин-синтеза А. Flavus разделенные интрона (рисунок 1). Фрагменты AFL2G_07224 (AFLR), AFL2G_07223 (AFLS или aflJ), AFL2G_05027 (афлатоксина оттока насоса, aflep), AFL2G_07228 (aflC / pksA / pksL1), и AFL2G_07731 (pes1) были использованы для конструкции, цифры на рисунке 1 соответствует , Flavus генома аннотации в Broad Institute, Кембридж, штат Массачусетс, и литературы 42. В общей сложности 99 линий арахиса регенерируют после прохождения процесса трансформации, 50 были ПЦР положительный для NPTII обнаруженного STN-ПЦР, и 33 линии были ПЦР положительные и производства семян. Только семь ПЦР положительные линии клонально размножают и проходят проверку на PresenСпособ т к накоплению афлатоксина, все семь показали между 60% и 100% меньше, чем накопления афлатоксина контролем. Здесь мы показываем результаты двух из этих семи линий. Как отдельные трансгенные события обычно производят несколько семян, был разработан метод, чтобы использовать минимальное количество семян в то же время быть в состоянии сделать параметрический статистический анализ. Блок-схема подготовки пробы и экспериментальной установки показана на рисунке 5 и в таблице 1. Хотя первое поколение трансгенных семян, как правило, гемизиготной, ожидается, что клетка-к-клетке и системной движения малых интерферирующих РНК (киРНК) генерируется через РНК-интерференции должны придать афлатоксин-синтез молчание по всему растению.

Рисунок 5. Схема блок-схема метода для анализа эффективности RNAi замолчать Aspergillus гены афлатоксин-синтеза в семенах арахиса. Графическое представление потока рабочей при обработке образцов арахиса для экспрессии генов или анализ афлатоксина. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
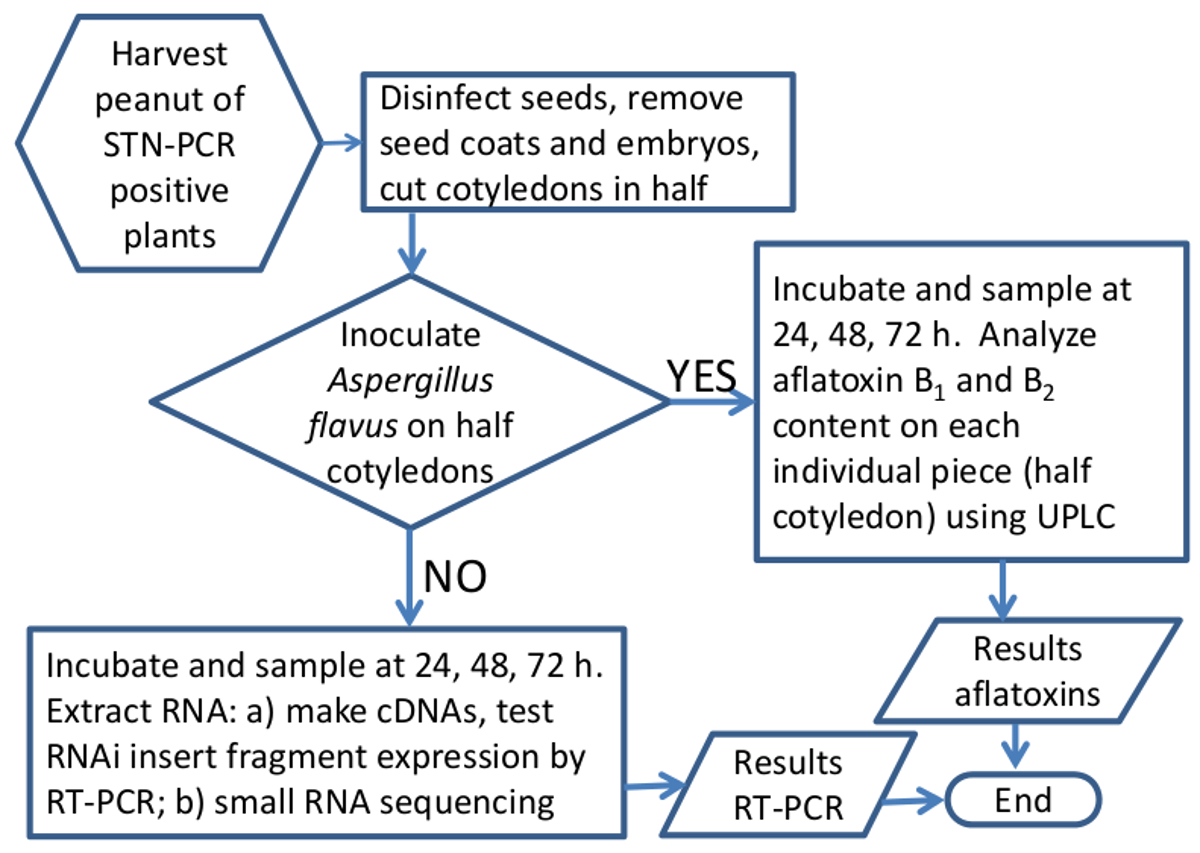{kind=link}
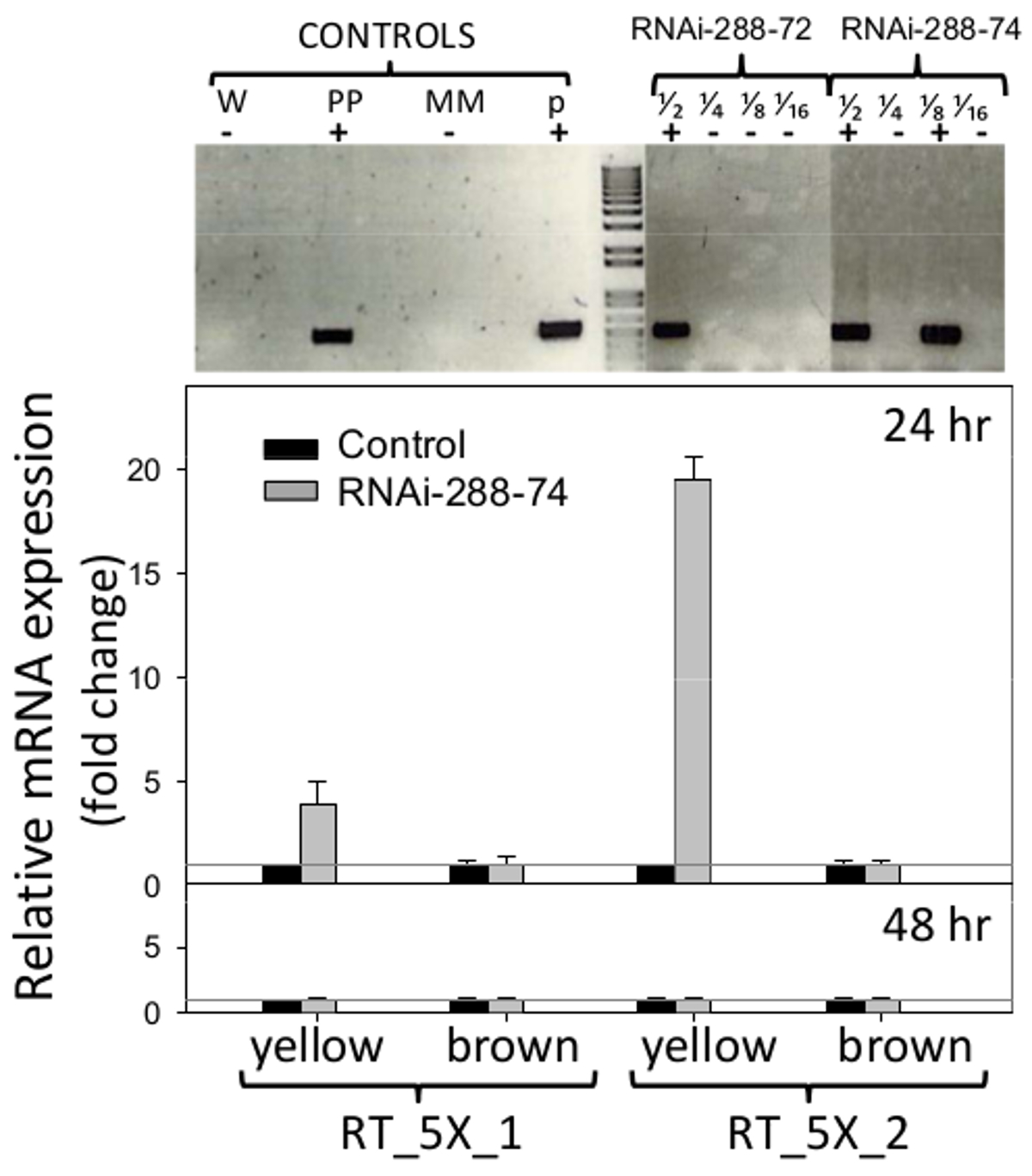
RNAi арахиса линии 288-72 288-74 и показал наличие селективного производителя NPTII, когда проходят проверку STN-ПЦР, гель разделы показаны на рис 6 (сверху), оригинальные фотографии доступны от авторов по запросу. Плазмиду pCAPD не использовали в качестве контроля трансформации, поскольку он кодирует инвертированных повторов двух генов CMR (устойчивости к хлорамфениколу) и БДКК (токсин) неизвестного воздействия на растения. ПЦР отрицательный арахисовое линии 288-9 и другие, которые прошли через процесс регенерации и выращивали в тех же условиях, что и РНК-интерференции линий, были использованы в качестве отрицательного конTrol.

. Рисунок 6. Обнаружение трансгенов и в режиме реального времени выражение РНК-интерференции вставка Топ: Однотрубная, обнаружение Вложенные ПЦР трансгенных линий арахиса РНК-интерференции 288-72 и 288-74 РНК-интерференции, положительных установок. Управление: W (вода), ПП (положительный завод), ММ (мастер микс), р (плазмиды p5XCAPD); фракции 1/2 1/4 1/8 1/16 представляют 2-кратного разведения ДНК Внизу:. ПЦР в реальном времени обнаружение экспрессии РНК-интерференции вставки (наборы праймеров: RT_5X_1, RT_5X_2, как показано на рисунке 1, на незрелых ( желтый) и пожилые (коричневый) семядоли трансгенных линий на 24 и 48hr инкубации; серая линия: C T = 1. Гистограммы представляют средства и стандартные бары ошибках (Т) трех биологических образцовс трех технических повторяет. Относительная количественная оценка РНК-интерференции вставкой нормализовалось по отношению к гену домашнего хозяйства актина в качестве внутреннего контроля и сравнительного выражения кратным трансгена, рассчитанной, как описано в 5.2.2 и 5.3. Пожалуйста, нажмите здесь, чтобы посмотреть большую версию этой фигуры.
{kind=link}
Чтобы проверить эффективность завода-хост РНК-интерференции-опосредованного потенциального контроля накопления афлатоксина, свежеубранных конидий Aspergillus Flavus NRRL 3357 были применены на поверхности среза половиной семядолей, из которых эмбрионы и теста были удалены (рис 3, 4). А. Flavus NRRL 3357, для которых геном был последовательности и стала основой для проектирования p5XCAPD, был любезно предоставлен доктором Рога в USDA-ARS-NPRL. Полученную грибковых вторжения половины семядолей после 24, 48 и 72 ч при 30 ° С в шоу п на рисунке 4. Образцы инокулированных половиной семядолей были собраны в 24, 48, 72, 96 ч инкубации и анализировали на главной четыре афлатоксинов В 1, В 2, G 1 и G 2 с помощью UPLC и подтвердили методом ЖХ-МС ; Результаты показаны на рисунке 6. концентрации Афлатоксин были определены с использованием опубликованного метода с изменениями 36. В 96 ч инкубации, семядоли начинают разрушаться из-за грибковой инфекции. РНК-интерференция линии 288-72 показали значительно более низкие уровни афлатоксинов, чем контроль на всех выборки дат в незрелых семядолей и в большинстве даты отбора проб в зрелых. РНК-интерференция линии 288-74 показали значительно более низкие уровни афлатоксинов в большинстве даты отбора проб. Уровни значимости теста Тьюки указаны звездочками в графике на рис 7.
g7.jpg "/>
. Рисунок 7. Афлатоксины В 1 и В 2 в половине арахиса семядолей после инкубации (24, 48, 72 и 96 часов) с Aspergillus Flavus управления: семена 288-9 без трансгенной линии; РНК-интерференция: семена РНК-интерференции-288-72 и 288-74-РНК-интерференции, трансгенных для РНК-интерференции p5XCAPD, чтобы заставить замолчать афлатоксин пять генов-синтеза. (А) афлатоксин В1 в зрелых семян (коричневый); (B), афлатоксин B 2 зрелые семена; (С) афлатоксин В1 в незрелых семян (желтый) и (D), афлатоксин В 2 в незрелых семян. Представлены средние значения с соответствующими стандартными баров ошибках (Т) дубликатов биологических образцов. Статистически значимые различия Тьюки тест *: р ≤ 0,05, ** Р ≤ 0,01, ***: р ≤ 0,001.с: //www.jove.com/files/ftp_upload/53398/53398fig7large.jpg "целевых =" _blank "> Нажмите здесь, чтобы посмотреть большую версию этой фигуры.
В целом, RNAi-288-72 показали в течение всего эксперимента (24 до 96 ч инкубации), 94% снижение афлатоксина В 2 в -100%, и 90% снижение на -100% в афлатоксина В1 по сравнению с контролем. РНК-интерференции 288-74 показали 63% снижение в -100% в афлатоксина B 2 и 60% -100% снижения в афлатоксина B 1, рисунке 7.
Праймеры, используемые в режиме реального времени обнаружения ПЦР экспрессии вставки РНК-интерференции показаны на рисунке 1. Арахис-семян семядоли были проанализированы без эмбрионов, чтобы удалить их естественные защитные и быть в состоянии обнаружить потенциальный эффект РНК-интерференции на площади, наиболее подверженных грибковые вторжения, семядоли. Экспрессия РНК-интерференции вставка обнаружен в незрелых семядолей (желтый) линии 288-74 от набора праймеров RT_5X_1 был четыре раза за C T = 1 порог отрицательного контроля, и набор праймеров RT_5X_2 был 19 раз выше порога, все при 24 ч инкубации. По крайней мере, три из пяти последовательных генных фрагментов, используемых в преобразовании арахисовое, 5027, 7223 и 7228 (aflep, AFLS / aflJ и aflC / pksA, соответственно) были обнаружены методом ОТ-ПЦР (Рисунок 1, 6). Выражение РНК-интерференции вставкой не был обнаружен в зрелых семядолей на 24 часов, или на зрелых или незрелых семядолей на 48 ч инкубации, рис 6 (нижний).
1| Арахис линия | Время выборки | Образцы для ОТ-ПЦР | Образцы для анализа афлатоксина (прививку) | Nхариус семян | ||||
| (не инокулированный) | ||||||||
| Вес 1 | Вес 2 | Вес 3 | Вес 1 | Вес 2 | Вес 3 | |||
| РНК-интерференция (желтый) | 24 ч | 1 | 1 | 1 | 1 | 1 | 1 | 4.5 |
| 48 ч | 1 | 1 | 1 | 1 | 1 | |||
| 72 ч | 1 | 1 | 1 | 1 | 1 | 1 | ||
| Контроль | 24 ч | 1 | 1 | 1 | 1 | 1 | 1 | 4.5 |
| (желтый) | 48 ч | 1 | 1 | 1 | 1 | 1 | 1 | |
| 72 ч | 1 | 1 | 1 | 1 | 1 | 1 | ||
Таблица 2. Пример установки маленького образца для анализа экспрессии генов и накопление афлатоксина в RNAi семян арахиса. Полный анализ экспрессии и афлатоксины для одной группы зрелости (т.е.., Желтый), три раза выборки (24, 48 и 72 ч) в трех экземплярах, потребует 4,5 семена, каждый номер один в таблице представляет половину семядолей.
Обсуждение
Завод-хост RNAi-обеспечиваемого молчания генов в патогенных грибов было продемонстрировано 27,43, однако, нет публикаций, показывающие возможность RNAi-опосредованной контроля микотоксинов накопления в растениях. Один ограничивающим фактором для этих исследований в арахиса было отсутствие метода оценки не-афлатоксина накопления фенотип в отдельных растений, а листья не показывают симптомов при грибковой инфекции подземных контейнеров. Кроме того, не-нормально распределена накопления афлатоксинов, и потребность в больших выборках для химического анализа 15,16 препятствовали количественно потенциального эффекта РНК-интерференции на одном заводе. Метод, представленный здесь, состоит из 72 ч экспериментов с использованием пяти семена, чтобы выполнить три 24 ч интервал выборки в трех экземплярах (таблица 1, рисунок 7). По сравнению с типичной анализа афлатоксина, который требует не менее 100 г семян, наш метод особенно подходит для индивидамил трансгенные события арахиса растений, которые изначально не производят не более двух или трех стручков.
РНК-опосредованного молчание синтеза афлатоксина была продемонстрирована генетически трансформации Aspergillus Flavus и A. parasiticus. Так AFLR является основным регулятором производства афлатоксина в А. Flavus и А. parasiticus 44,45, он становится интересным объектом для РНК-опосредованного молчания в растениях. Тем не менее, генетические вариации в AFLR было показано среди Aspergillus видов 46, и эти генетические варианты могли избежать, если заглушить нет идеально совпадающей последовательности с сигналом RNAi, полученного в растения-хозяина. Таким образом, AFLR был одним из мишеней для глушителей в векторном p5XCAPD, но не был единственным. Инвертированных повторов гена AFLR, внесенные в А. Flavus и А. parasiticus путем преобразования привели к глушителей и минимальным или нет продуктионный афлатоксинов 47 (McDonald и др., 2005b). Кроме того, глушителей AFLD ген предотвратить производство афлатоксина до 98% в А. Flavus и А. parasiticus в прямом преобразовании 48. Для повышения вероятности успеха в нашей системе, арахисовое трансформировали инвертированный повтор фрагментов пяти генов, участвующих в производстве афлатоксина в А. Flavus. Здесь показано, что использование p5XCAPD, что цели для глушителей несколько генов в синтез афлатоксина пути, 90% -100% более низкие уровни афлатоксина B 1 и B 2 были достигнуты в соответствии 288-72 и 60-100% более низкие уровни, накопленный в Линия 288-74 по сравнению с контролем, когда половина семядоли были засеянной А. Flavus, цифры 4, 7. Самое главное, этот метод обнаружено статистически значимых различий в накоплении афлатоксина от линий 288-72 288-74, против контроля на протяжении всего эксперимента с применением параметрического STATISтики, Рисунок 7. Учитывая небольшой размер выборки, важно подчеркнуть необходимость использования мощного метода для обнаружения афлатоксины, эти эксперименты были проанализированы с помощью UPLC который имеет высокое разрешение, в пять раз более высокую производительность и в три раза более высокую чувствительность, чем ВЭЖХ 49.
Экспрессия РНК-интерференции вставки в 288-74 был обнаружен только в незрелых семядолей (желтый) в 24 ч инкубации. Вставка РНК-интерференции не обнаруженные RT-PCR на зрелых семядолей 288-74 на 24 часов, или на любом погашения группы в 48 ч, на рисунке 6. Это же явление наблюдалось и в других РНК-интерференции трансгенных линий арахиса (Arias, RS, 2015 неопубликованные), где обычно RNAi стенограммы были обнаружены только на незрелых семядолей на 24 часов. Образцы РНК обрабатывали ДНКазой перед синтеза кДНК, данные были нормированы на уровень экспрессии актина и без признаков контаминации ДНК наблюдалось. Должен ДНК были присутствовать в образцах, он долженбыли обнаружены в образцах ч 48, а также, но последовательно, что было не так. Выражение под контролем 35S промотора не всегда равномерное; это может повлиять на условия окружающей среды 50, типа ткани и стадии развития 51,52. В то же время, в пути РНК-интерференции, скорость распада мРНК и скорость распада миРНК может значительно варьироваться в зависимости 53. Вполне возможно, что быстрое ухудшение мРНК по механизму РНК-интерференции могло бы предотвратить обнаружение мРНК на 48 ч инкубации. Будь отсутствие экспрессии на 48 часов было связано с низким 35S-промотора приводом транскрипции, или быстрой деградации дцРНК по Dicer остается без ответа. Таким образом, обнаружение малых РНК высокой пропускной последовательности даст лучшее представление о процессах, происходящих через РНК-интерференции 54 В этих экспериментах. Однако, поскольку РНК молчание распространяется системно, в основном, через флоэмы от photosyntненавижу источники сахарозу раковины (в этом случае арахиса семян) 55, замалчивание афлатоксина-синтеза может произойти в семенах без местного выражения RNAi вставки. Много исследований еще предстоит сделать, чтобы определить пороговый уровень малых интерферирующих РНК (миРНК), необходимых для предотвращения накопления афлатоксина в семенах. Важно подчеркнуть тот факт, что оба, экспрессия мРНК в РНК-интерференции построить (рисунок 6), и накопление афлатоксин B 1 и B 2 (рис 7) показали разные результаты для незрелых (желтый) против. зрелая (коричневый) семядоли. Арахис растения имеют неопределенный рост, то есть, они представляют во время сбора урожая в диапазоне от погашения стручки, рис 2. Кроме того, семена из разных групп зрелости отличаются по своему химическому составу., Например, 2,4% сахарозы в незрелых семян, а 1,9% в зрелые семена под одной полевых условиях. 56,57 Таким образом, чтобы понять фактическое efficiencу РНК-опосредованного контроля накопления афлатоксина, важно, чтобы проанализировать группы отдельно зрелости.
Естественный защита семян арахиса является производство фитоалексинов, которая меняется в многообразии соединений, полученных и их относительных величин, в зависимости от зрелости семян и условий окружающей среды 58-61, и это особенно выше у эмбрионов по сравнению с семядолей 62. Эмбрионы также значительно более высокие концентрации нуклеиновых кислот, ДНК и РНК, чем семядолей (Arias RS, неопубликованных). Как семена арахиса созревают, изменения в их физиологии и химического состава происходит 63. Фенольные антиоксиданты в форме арахиса теста конденсированных дубильных веществ с фунгистатическим деятельности; 64 это очевидно в мезокарпий цвета, который отражает погашения этапы, желтые черный 35, а его содержание дубильных веществ и фенольных соединений увеличивается с зрелости 65. Таким образом, наличие тТелепрограм или эмбрионов в эксперименте, с учетом их антимикробные свойства, мог бы ограничен рост грибков и, следовательно, трудно переоценить эффект RNAi молчания, следовательно, они были удалены. Кроме того, удаление кожуры и эмбрионов помогает ограничить источники вариации в анализе, как половина семядолей, который несет зародыш будет больше и больше фитоалексинов содержание РНК.
В дополнение к анализу зрелости групп и удаление семенной кожуры и зародыша в этих экспериментах, важно отметить, несколько наблюдений: а) если результаты приведены в течение 96 ч инкубации, рекомендуется использовать не более чем на 72 ч, чтобы получить достоверные результаты, так как семена получают разрушается 96 часов; и б), тогда как половина семядоли из того же семени, хотя пробы случайным образом, не являются идеально независимых выборок, ОТ-ПЦР и накопление афлатоксина в трансгенных событий показал минимальную вариацию между семенами. Кроме того, рассчитывать точное грибковые споры, объем посевнойс 2 мкл, и применение спор на поверхности среза семядолей избегая капает по бокам важны, чтобы убедиться, проросшие споры подвергаются ткани растений. Вода / агар на пластинах должны быть на уровне 1,5% (вес / объем), более мягкий агар вызывает сток спор, как показано на последнем кадре, показанном на фиг.4 (внизу). Если семена наличие от конкретного трансгенного случае ограничивается, выборка может быть сделано в двух экземплярах вместо тройных получения аналогичными результатами (например, рисунок 7); Однако три образца поможет уменьшить стандартную ошибку. Единственное ограничение этого метода является то, что он требует очень чувствительная система (UPLC) для обнаружения афлатоксин / количественного, но в то же время это уменьшает вероятность переоценить влияние РНК-интерференции должны афлатоксины не обнаружено менее чувствительных методов.
В заключение, этот метод предлагает впервые надежный подход для изученияЭффект РНК-интерференции в контроле афлатоксинов. Сокращение времени для эксперимента из всей посевной сезон до менее чем за одну неделю, этот метод будет чрезвычайно ускорить исследования по РНК-интерференции арахиса / Aspergillus pathosystem к смягчению и / или устранению афлатоксинов.
Раскрытие информации
The authors have nothing to disclose or any conflicts of interest.
Благодарности
This work received the financial support of USDA-ARS CRIS project 6604-21000-004-00D, CRIS project 6604-42000-008-00D, and USAID Feed-the-Future program Agreement number 58-0210-3-012. We thank Valerie Orner, LaTanya Johnson, Joseph Powell and Kathy Gray for their technical assistance. Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the US Department of Agriculture.
Материалы
| Name | Company | Catalog Number | Comments |
| Primers, oligonucleotides | DNA Technologies, Coralville, IA, USA | n/a | |
| Dneasy Plant Mini Kit | Qiagen, Valencia, CA | 69106 | |
| Czapek Dox agar medium | Oxoid, by Thermo Fisher Scientific, Waltham, MA | CM0095 | |
| Agar | Thermo Fisher Scientific, Waltham, MA | BP 1423 | |
| Freezer -80 °C | n/a | n/a | |
| Aluminum Oxide, Al2O3 | Fisher Scientific | A941 | |
| SPE Reservoirs 1.5 ml | Grace Davison Discovery Scientific | 210011 | |
| Frits for 1.5 ml SPE reservoir | Grace Davison Discovery Scientific | 211401 | |
| Autosampler vials | Waters Corporation, Milford, MA | 186005221 | |
| Waters Acquity Ultra-Performance Liquid-Chromatography (UPLC) instrument; UPLC-H-Class Quaternary Solvent Manager; UPLC Sample Manager; UPLC Fluorescent detector (FLR); UPLC BEH C18 2.1 mm x 50 mm, 1.7 mm column | Waters Corporation, Milford, MA | ||
| Finnigan LCQ Advantage MAX ion trap mass spectrometer, with Xcalibur version 1.4 software | Thermo Electron Corp., San Jose, CA | ||
| Aflatoxin standards, B1, B2, G1 and G2 | Sigma-Aldrich, St. Louis, MO | A6636; A9887; A0138; A0263 | |
| Systat Software 12.2 | SYSTAT Software Inc., Point Richmond, CA | ||
| Trizol reagent | Invitrogen, CA | 15596-018 | |
| SuperScript III First Strand Synthesis Super Mix | Invitrogen, CA | 11752-050 | |
| ABI 7500 Real-Time PCR | Lifetechnologies, Grand Island, NY | 4406984 | |
| Luria Broth-Miller | Fisher Scientific | R453642 | |
| pENTR1A | Invitrogen, CA | A10462 | |
| LR Clonase II enzyme mix | Invitrogen, CA | 11791-020 | |
| T4 DNA Ligase | NEB Biolabs | M0202L | |
| Gelrite | Sigma-Aldrich, St. Louis, MO | G1919 | |
| Acetosyringone | Sigma-Aldrich, St. Louis, MO | D134406 | |
| QIAcube robot workstation | Qiagen, Valencia, CA | 9001292 | |
| Antibiotics: kanamycin, cefotaxime, gentamicin; streptomycin | Goldbio, St. Louis, MO | cef.: C-104-25; kan: K-120-5; gent.: G-400-1; strep.: S-150-50 | |
| Platinum Taq DNA Polymerase High Fidelity | Invitrogen, CA | 11304-029 |
Ссылки
- Williams, J. H., et al. Human aflatoxicosis in developing countries: a review of toxicology, exposure, potential health consequences, and interventions. The American Journal of Clinical Nutrition. 80, 1106-1122 (2004).
- American Association for Cancer Research: AACR. An evaluation of chemicals and industrial processes associated with cancer in humans based on human and animal data: IARC Monographs Volumes 1 to 20. Cancer Research. 40, 1-12 (1980).
- Turner, P. C. The molecular epidemiology of chronic aflatoxin driven impaired child growth. Scientifica. , (2013).
- Rasooly, R., Hernlem, B., He, X., Friedman, M. Non-linear relationships between aflatoxin B1 levels and the biological response of monkey kidney vero cells. Toxins (Basel). 5, 1447-1461 (2013).
- Gong, Y. Y., et al. Determinants of aflatoxin exposure in young children from Benin and Togo, West Africa: the critical role of weaning. International Journal of Epidemiology. 32, 556-562 (2003).
- Eaton, D. L., Groopman, J. D. The toxicology of aflatoxins: human health, veterinary, and agricultural significance. , Academic Press. (1994).
- Murugavel, K. G., et al. Prevalence of aflatoxin B1 in liver biopsies of proven hepatocellular carcinoma in India determined by an in-house immunoperoxidase test. Journal of Medical Microbiology. 56, 1455-1459 (2007).
- Wang, J. S., et al. Hepatocellular carcinoma and aflatoxin exposure in Zhuqing Village, Fusui County, People's Republic of China. Cancer Epidemiology, Biomarkers & Prevention. 10, American Association for Cancer Research. 143-146 (2001).
- Azziz-Baumgartner, E., et al. Case-control study of an acute aflatoxicosis outbreak, Kenya, 2004. Environmental Health Perspectives. 113, 1779-1783 (2005).
- Lye, M. S., Ghazali, A. A., Mohan, J., Alwin, N., Nair, R. C. An outbreak of acute hepatic encephalopathy due to severe aflatoxicosis in Malaysia. American Journal of Tropical Medicine and Hygiene. 53, 68-72 (1995).
- Villers, P. Aflatoxins and safe storage. Frontiers in Microbiology. 5, 158(2014).
- Kensler, T. W., Roebuck, B. D., Wogan, G. N., Groopman, J. D. Aflatoxin: A 50-year odyssey of mechanistic and translational toxicology. Toxicological Sciences. 120, S28-S48 (2011).
- Dorner, J. W., Cole, R. J., Wicklow, D. T. Aflatoxin reduction in corn through field application of competitive fungi. Journal of Food Protection. 62, 650-656 (1999).
- Cotty, P. J., Bhatnagar, D. Variability among atoxigenic Aspergillus flavus strains in ability to prevent aflatoxin contamination and production of aflatoxin biosynthetic-pathway enzymes. Applied and Environmental Microbiology. 60, 2248-2251 (1994).
- Whitaker, T. B. Standardisation of mycotoxin sampling procedures: an urgent necessity. Food Control. 14, 233-237 (2003).
- Whitaker, T. B., Dorner, J. W., Giesbrecht, F. G., Slate, A. B. Variability among aflatoxin test results on runner peanuts harvested from small field plots. Peanut Science. 31, 59-63 (2004).
- Fire, A., et al. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature. 391, 806-811 (1998).
- Rafael, D., et al. EMT blockage strategies: Targeting Akt dependent mechanisms for breast cancer metastatic behaviour modulation. Current Gene Therapy. , (2015).
- Li, G., Chang, H., Zhai, Y. P., Xu, W. Targeted silencing of inhibitors of apoptosis proteins with siRNAs: a potential anti-cancer strategy for hepatocellular carcinoma. Asian Pacific. Journal of Cancer Prevention: APJCP. 14, 4943-4952 (2013).
- Koldehoff, M. Targeting bcr-abl transcripts with siRNAs in an imatinib-resistant chronic myeloid leukemia patient: challenges and future directions. Methods in Molecular Biology. 1218, 277-292 (2015).
- Zhang, J., et al. Pest control. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science. 347, 991-994 (2015).
- Ajjappala, H., Chung, H. Y., Sim, J. S., Choi, I., Hahn, B. S. Disruption of prefoldin-2 protein synthesis in root-knot nematodes via host-mediated gene silencing efficiently reduces nematode numbers and thus protects plants. Planta. 241, 773-787 (2015).
- Jose, A. M., Hunter, C. P. Transport of sequence-specific RNA interference information between cells. Annual Review of Genetics. 41, 305-330 (2007).
- Vazquez, F., Hohn, T. Biogenesis and biological activity of secondary siRNAs in plants. Scientifica. , Hindawi Publishing Corporation. (2013).
- Tinoco, M. L. P., Dias, B. B. A., Dall'Astta, R. C., Pamphile, J. A., Aragao, F. J. L. In vivo trans-specific gene silencing in fungal cells by in planta expression of a double-stranded RNA. BMC Biology. 8, (2010).
- Govindarajulu, M., Epstein, L., Wroblewski, T., Michelmore, R. W. Host-induced gene silencing inhibits the biotrophic pathogen causing downy mildew of lettuce. Plant Biotechnology Journal. , (2014).
- Yin, C., Jurgenson, J. E., Hulbert, S. H. Development of a host-induced RNAi system in the wheat stripe rust fungus Puccinia striiformis f. sp. tritici. Molecular Plant-Microbe Interactions. 24, 554-561 (2011).
- Ghag, S. B., Shekhawat, U. K., Ganapathi, T. R. Host-induced post-transcriptional hairpin RNA-mediated gene silencing of vital fungal genes confers efficient resistance against Fusarium. wilt in banana. Plant Biotechnology Journal. 12, 541-553 (2014).
- Filichkin, S. A., et al. Efficiency of gene silencing repeats vs. transitive RNAi in Arabidopsis: direct inverted vectors. Plant Biotechnology Journal. 5, 615-626 (2007).
- Sciaky, D., Montoya, A. L., Chilton, M. D. Fingerprints of Agrobacterium Ti Plasmids. Plasmid. 1, 238-253 (1978).
- Clark, D. J., Maaloe, O. DNA Replication and Division Cycle in Escherichia coli. Journal of Molecular Biology. 23, 99-112 (1967).
- Murashige, T., Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plantarum. 15, 473-497 (1962).
- Srinivasan, T., Kumar, K. R. R., Kirti, P. B. Establishment of efficient and rapid regeneration system for some diploid wild species of Arachis. Plant Cell Tissue and Organ Culture. 101, 303-309 (2010).
- Gomes, A. L. V., et al. Single-tube nested PCR using immobilized internal primers for the identification of dengue virus serotypes. Journal of Virology Methods. 145, 76-79 (2007).
- Williams, E. J., Drexler, J. S. A non-destructive method for determining peanut pod maturity. Peanut Science. 8, 134-141 (1981).
- Sobolev, V. S., Dorner, J. W. Cleanup procedure for determination of aflatoxins in major agricultural commodities by liquid chromatography. Journal of AOAC International. 85, 642-645 (2002).
- Empower Software, Getting Started Guide. , Waters Corporation. Milford, MA. Available from: http://sites.chem.colostate.edu/diverdi/C431/experiments/high%20pressure%20liquid%20chromatography/references/Empower%20getting%20started%2071500031203rA.pdf (2002).
- Biselli, S., Hartig, L., Wegner, H., Hummert, C. Analysis of Fusarium. toxins using LC-MS-MS: Application to various food and feed matrices. LC GC North America. 23, 404-413 (2005).
- Arias, R. S., Sobolev, V. S., Orner, V. A., Dang, P. M., Lamb, M. C. Potential involvement of Aspergillus flavus laccases in peanut invasion at low water potential. Plant Pathology. 63, 353-363 (2014).
- Dang, P. M., Chen, C. Y., Holbrook, C. C. Evaluation of five peanut (Arachis hypogaea) genotypes to identify drought responsive mechanisms utilising candidate-gene approach. Functional Plant Biology. 40, 1323-1333 (2013).
- Schmittgen, T. D., Livak, K. J. Analyzing real-time PCR data by the comparative C-T method. Nature Protocols. 3, 1101-1108 (2008).
- Amaike, S., Keller, N. P. Aspergillus flavus. Annual Review of Phytopathology. 49, 107-133 (2011).
- Nowara, D., et al. HIGS: Host-induced gene silencing in the obligate biotrophic fungal pathogen Blumeria graminis. Plant Cell. 22, 3130-3141 (2010).
- Woloshuk, C. P., et al. Molecular characterization of aflR, a regulatory locus for aflatoxin biosynthesis. Applied and Environmental Microbiology. 60, 2408-2414 (1994).
- Price, M. S., et al. The aflatoxin pathway regulator AflR induces gene transcription inside and outside of the aflatoxin biosynthetic cluster. FEMS Microbiology Letters. 255, 275-279 (2006).
- Ehrlich, K. C., Montalbano, B. G., Cotty, P. J. Sequence comparison of aflR from different Aspergillus. species provides evidence for variability in regulation of aflatoxin production. Fungal Genetics and Biology. 38, 63-74 (2003).
- McDonald, T., Brown, D., Keller, N. P., Hammond, T. M. RNA silencing of mycotoxin production in Aspergillus and Fusarium species. Molecular Plant Microbe Interactions. 18, 539-545 (2005).
- Abdel-Hadi, A. M., Caley, D. P., Carter, D. R., Magan, N. Control of aflatoxin production of Aspergillus flavus. and Aspergillus parasiticus. using RNA silencing technology by targeting aflD. (nor-1) gene. Toxins (Basel). 3, 647-659 (2011).
- Swartz, M. E. Ultra performance liquid chromatography (UPLC): An introduction: Separation Science Redefined. LCGC North America. , Suppl ement 8. 8-14 (2005).
- Maghuly, F., Khan, M. A., Fernandez, E. B., Druart, P., Watillon, B., Laimer, M. Stress regulated expression of the GUS-marker gene (uidA) under the control of plant calmodulin and viral 35S promoters in a model fruit tree rootstock: Prunus incisa x serrula. Journal of Biotechnology. 135, 105-116 (2008).
- de Mesa, M. C., Santiago-Doménech, N., Pliego-Alfaro, F., Quesada, M. A., Mercado, J. A. The CaMV 35S promoter is highly active on floral organs and pollen of transgenic strawberry plants. Plant Cell Reports. 23, 32-38 (2004).
- Sunilkumar, G., Mohr, L., Lopata-Finch, E., Emani, C., Rathore, K. S. Developmental and tissue-specific expression of CaMV 35S promoter in cotton as revealed by GFP. Plant Molecular Biology. 50, 463-474 (2002).
- Groenenboom, M. A. C., Maree, A. F. M., Hogeweg, P. The RNA silencing pathway: The bits and pieces that matter. PLoS Computational Biology. 1, 155-165 (2005).
- Zhao, D., Song, G. Q. High-throughput sequencing as an effective approach in profiling small RNAs derived from a hairpin RNA expression vector in woody plants. Plant Science: an International Journal of Experimental Plant Biology. 228, 39-47 (2014).
- Kamthan, A., Chauduri, A., Kamthan, M., Datta, A. Small RNAs in plants: recent development and application for crop improvement. Frontiers in Plant Science. 6, 208(2015).
- Manda, A., Bodapati, P. N., Rachaputi, N. C., Wright, G., Fukai, S. Aflatoxins and their relationship with sugars in peanut (Arachis hypogaea L). 4th International Crop Science Congress, 2004, , Available from: http://www.cropscience.org.au/icsc2004/poster/5/1/3/625_manda.htm (2004).
- Uppala, S. S. Factors affecting pre-harvest aflatoxin contamination of peanut (Arachis hypogaea L). , Auburn University. (2011).
- Sobolev, V. S. Localized production of phytoalexins by peanut (Arachis hypogaea) kernels in response to invasion by Aspergillus species. Journal of Agricultural and Food Chemistry. 56, 1949-1954 (2008).
- Sobolev, V. S., Guo, B. Z., Holbrook, C. C., Lynch, R. E. Interrelationship of phytoalexin production and disease resistance in selected peanut genotypes. Journal of Agricultural and Food Chemistry. 55, 2195-2200 (2007).
- Sobolev, V. S., Neff, S. A., Gloer, J. B. New stilbenoids from peanut (Arachis hypogaea) seeds challenged by an Aspergillus caelatus strain. Journal of Agricultural and Food Chemistry. 57, 62-68 (2009).
- Dorner, J. W., Cole, R. J., Sanders, T. H., Blankenship, P. D. Interrelationship of kernel water activity, soil temperature, maturity, and phytoalexin production in preharvest aflatoxin contamination of drought-stressed peanuts. Mycopathologia. 105, 117-128 (1989).
- Sobolev, V. S. Production of phytoalexins in peanut (Arachis hypogaea) seed elicited by selected microorganisms. Journal of Agricultural and Food Chemistry. 61, 1850-1858 (2013).
- Basha, S. M. M., Cherry, J. P., Young, C. T. Changes in free amino acids, carbohydrates, and proteins of maturing seeds from various peanut (Arachis hypogaea L.) cultivars. Cereal Chemistry. 53, 586-596 (1976).
- Lansden, J. A. Aflatoxin inhibition and fungistasis by peanut tannins. Peanut Science. 9, 17-20 (1982).
- Yen, G. C., Duh, P. D., Tsai, C. L. Relationships between antioxidant activity and maturity of peanut hulls. Journal of Agricultural and Food Chemistry. 41, 67-70 (1993).
Перепечатки и разрешения
Запросить разрешение на использование текста или рисунков этого JoVE статьи
Запросить разрешениеСмотреть дополнительные статьи
This article has been published
Video Coming Soon
Авторские права © 2025 MyJoVE Corporation. Все права защищены