
Method Article
利用相扑融合策略生成国产、未加标签的杭丁顿蛋白 Exon1 单体和纤维
摘要
在这里, 我们提出了一个健壮和优化的协议, 以生产毫克数量的本地, 无标签单体和纤维的 exon1 的杭丁顿蛋白蛋白 (Httex1) 的基础上的瞬态融合的小泛素相关修饰剂 (相扑)。
摘要
亨廷顿氏病 (hd) 是一种遗传性的致命神经退行性疾病, 由 CAG 扩张 (≥36) 在 HD 基因的第一个外显子, 导致杭丁顿蛋白蛋白 (Htt) 或 N 末端片段的表达, 其扩大 polyglutamine (polyQ) 伸展。
杭丁顿蛋白蛋白 (Httex1) 的 exon1 是最小的 Htt 片段, 它概括了细胞和动物模型中 HD 的许多特征, 是 Htt 最广泛研究的片段之一。Httex1 的小尺寸使得实验更适合使用标准和高分辨率技术与较长的碎片或全长 Htt 的生物物理特性进行比较。然而, 随着 polyQ 含量 (≥42) 的增加, 突变体 Httex1 (mHttex1) 的高聚集倾向使得开发高效的表达和纯化系统难以产生足够数量的蛋白质并使其易于科学家从不同的学科, 没有使用融合蛋白或其他策略, 改变了本机序列的蛋白质。本文提出了一种基于小泛素相关修饰剂 (相扑) 的瞬态融合技术, 对国产、无标签 Httex1 毫克量的生产进行了稳健而优化的方法。该策略的简单性和效率将消除使用非本机序列 Httex1 的需要, 从而使研究人员更容易获得这种蛋白质, 并提高了不同实验室的实验重现性。我们相信, 这些进展也将促进今后的研究, 旨在阐明 Htt 的结构功能关系, 以及开发新的诊断工具和治疗方法, 以治疗或减慢 HD 的进展。
引言
Htt 是一种 348 kDa 蛋白, 并被牵连在几个生理功能1。当 Htt 包含一个扩展的 polyQ 区域超过36残余在它的 N 末端, 它导致 HD2,3。HD 病理特征为细胞包裹体在纹状物和皮层, 导致神经元死亡和萎缩的受影响的组织4,5。从 HD 患者死后的大脑中发现了一些含有 polyQ 重复道的 n-端 Htt 片段, 被认为是由杭丁顿蛋白蛋白6的蛋白质水解处理产生的。最近的研究表明, Httex1 也可以形成由于异常的 mRNA 剪接。Httex1 含有病理 polyQ 突变, 其在动物中的过度表达可以重述 hd7的许多关键特征, 从而突显出这个片段在 hd 病理和疾病进展6中可能发挥的核心作用, 8,9。
由于突变体 Httex1 (mHttex1) 的高聚集倾向与扩张的 polyQ, 大多数现有的表达系统是基于 Httex1 与蛋白质的瞬态融合 (如谷胱甘肽 s-转移酶 (GST), thioredoxin (TRX) 或麦芽糖结合蛋白 (MBP) 和/或多肽 (聚组氨酸) 差异改善其表达, 稳定性, 纯化和/或溶解度10,11,12,13,14 ,15,16,17,18,19,20,21,22,23 ,24,25,26,27,28。融合伙伴与 Httex1 链接到一个短序列, 其中包含一个蛋白酶的裂解部位, 如胰蛋白酶, 烟草蚀刻病毒 (TEV) 蛋白酶或 PreScission, 以允许分裂和释放 Httex1 之前开始聚合或净化。这些方法的缺点包括可能留下额外的残余由于非无影无踪分裂和创造被截断的片断由于 miscleavage 在 Httex1 序列之内, 除了异质性由于不完全分裂 (见 Vieweg等人就这一方法的优点和局限性进行更深入的讨论)10.为了解决这些限制, 我们最近开发了一种表达式策略, 它利用Synechocystis sp的瞬态 N 终端融合, 首次启用无标记本机 Httex1 的生成。(Ssp)DnaB 蛋白质内含子到 Httex110。虽然蛋白质内含子是无影无踪和特异的 , 并产生镁数量的蛋白质 , 它仍然有两个缺点 , 可以降低产量 : 即在表达过程中可能发生的蛋白质内含子的过早裂 , 和分裂发生的事实几个小时, 这可能导致蛋白质的损失由于聚集, 特别是为 Httex1 与扩大的 polyQ 重复。
为了解决这些限制, 并完善我们的国产, 无标签 Httex1 的生产策略, 我们开发了一个新的表达系统的基础上的瞬态融合的相扑, 更确切的酵母同源 Smt3 Httex1。相扑系统在重组蛋白生产中的应用首次发表于 2004年29, 其中相扑融合蛋白的表达率和溶解度提高。相扑标记可以通过泛素, 如蛋白特异蛋白酶 1 (ULP1), 这并不需要一个识别网站, 但承认相扑的三重结构, 并实际上消除了 miscleavage30的可能性。此外 , ULP1 - mediated 是快速和无影无踪 , 并没有留下额外的残留物后面。与自催化蛋白质内含子10所观察到的融合标记的过早劈裂是由外部蛋白酶的要求完全避免的。虽然相扑战略目前广泛用于重组蛋白生产31,32,33, 我们证明, 它是特别有用的产生一个内在无序,聚集性容易, amyloidogenic 蛋白如 Httex1。我们认为, 我们的相扑融合方法的简单性、效率和鲁棒性将使来自不同学科的研究人员更容易接触到无标记的 Httex1, 并消除了在体外使用非本机序列的需要 Httex1.这是一个重要的进步, 将促进未来的研究, 以阐明结构-功能关系的 Httex1。
该协议描述了 Httex1 从细菌培养的 12 L 的纯化, 但该协议可以很容易地适应较小或更大规模的生产。该协议描述了野生型 Httex1 (wtHttex1) 的生产, polyQ 重复长度低于 (23Q) 和突变 Httex1 (mHttex1), polyQ 重复长度超过 (43Q) 致病阈值 (36Q)。
研究方案
1. 重组 Httex1 23Q 和43Q 的表达
- 准备所需的缓冲区和解决方案。准备1000x 氨苄西林 (安培, 100 毫克/毫升) 库存解决方案, 过滤器 (0.2 µm), 整除和存储在-20 摄氏度。准备溶源性汤 (磅) 培养基 (25 克磅米勒每1升2O), 高压釜。准备 1 M 异丙基ß-1 thiogalactopyranoside (IPTG) 库存溶液, 过滤器 (0.2 µm), 整除和贮存在-20 摄氏度。
- 用 pTWIN1 载体转化化学能力大肠杆菌B ER 2566, 包含人类 Httex1 融合到 N 终端 His6-SUMO 标记与热休克方法34。
注:大肠杆菌BL21 DE3 菌株也被使用。然而, 在这种情况下, 观察到了截断的增加量。 - 将200毫升的 LB 培养基与1x 安培在1升圆锥烧瓶中添加一个单一的殖民地, 从琼脂板与不育的吸管提示。孵化的文化在30摄氏度和 180 rpm 20 小时 (隔夜) 在细菌孵化器。
- 用不育的吸管取1毫升的培养基样本。测量的光学密度在600毫微米 (OD600) 的样品与一次性塑料试管和光度计 (尊敬测量范围在0.1 和1之间, 稀释与 LB 介质, 如果需要)。计算在 3 L 文化中, 将导致起始 OD600为0.05 的 preculture 的数量 (与 od600 = 3 的 preculture, 这意味着50毫升)。
- 接种四种培养 (每3升的 LB 培养基与1x 安培在 5 L 烧瓶), 通过增加计算量的 preculture 与不育的吸管。孵化的文化在37摄氏度和 180 rpm 的细菌孵化器。
- 每30分钟, 取一个1毫升的培养基样本, 用不育的吸管。用一次性塑料试管和光度计测量样品的外径600 。当 OD600达到 0.1 (典型地在 1-2 h 以后), 设置细菌孵化器的温度到14°c 并且在冷却期间继续孵化。每30分钟, 取一个1毫升的培养基样本, 用不育的吸管。用一次性塑料试管和光度计测量样品的外径600 。
注: 冷却文化的时间可能因使用的孵化器而异, 因此开始降温的时间可能需要根据使用的孵化器类型进行调整。然而, 改变温度梯度只会对产量有很小的影响, 因为相扑融合蛋白看起来相当稳定。 - 当 OD600达到 0.3-0.4 (通常是在1-2 小时之后), 取一个预先感应样品的文化为 SDS 页分析的过度表达。计算在考马斯染色的 SDS 页上给出可比较数量的细胞和良好信号的样本大小:10 井凝胶: 容量 = 0.2 毫升/OD600;用一半做15井凝胶。
- 对于细菌培养与 OD600= 0.4, 采取500µL。用不育的吸管来计算细菌培养量。向下旋转样品 (18000 x g, 4 °c, 2 分钟) 并丢弃上清。保持颗粒在-20 °c, 直到准备用于分析 (步骤 1.11)。
- 通过吹打1.2 毫升1米 IPTG 库存溶液诱导蛋白质表达, 每3升培养溶液 (最终浓度0.4 毫米)。继续孵化的文化在14°c 16 小时 (隔夜)。
注: 在添加 IPTG 时, 温度通常会达到摄氏20摄氏度, 这取决于孵化器的性能。 - 按照步骤1.7 中描述的步骤, 在对过度表达进行 SDS 页分析的情况下, 采取文化的后诱导样本。
- 通过离心 1 L 管 (3993 x g, 4 °c, 10 分钟) 来收获细胞。丢弃上清液, 将细胞颗粒放在冰上, 直接进行纯化。
- 用 SDS 分析35、36页的过度表达。并用重悬在20µL 的运行缓冲器和20µL 的2x 加载染料的前、后感应样品。加热样品5分钟在95摄氏度在一个热块和负载20µL 在15% 凝胶, 而仍然热。根据制造商的指示, 在 180 v 上用考马斯染料染色凝胶90分钟。将结果与图 1C中的代表结果进行比较。
注: 该协议可以在这里停止, 细胞颗粒可以冻结和储存在-80 °c 几个星期。为获得最佳效果, 建议使用新鲜的细菌颗粒, 避免冷冻。冻融可能导致细胞的裂解和 Httex1 的降解。这可能会降低蛋白质的产量和质量。
2. 固定化金属亲和层析 (IMAC) 对其6-相扑 Httex1 融合蛋白的细胞裂解纯化
- 在玻璃瓶 (50 毫米三 (羟甲基)-aminomethan (三), 500 毫米氯化钠, 15 毫米咪唑) 上准备 2 L 的缓冲 a。在玻璃瓶中准备1升的缓冲 B (50 毫米三, 500 毫米氯化钠, 500 毫米咪唑 pH 7.4)。溶解盐后, 用10氮盐调节 pH 值, 用瓶顶过滤器 (0.65 µm) 将溶液过滤成新鲜瓶子。准备一个 1000x phenylmethylsulfonyl 氟 (PMSF, 0.3 米) 库存解决方案, 整除到100µL 和存储在-20 摄氏度。
注: 该协议的目的是使完成所有步骤, 从裂解的细菌小球的反相高效液相色谱 (RP) 纯化和冻干在8-9 小时内。为了限制聚合和蛋白质分解, 建议在不暂停的情况下快速工作, 并在4摄氏度或冰上执行所有步骤。 - 添加100µL 的 PMSF 库存溶液和五片 (1 每30毫升的最终体积) 蛋白酶抑制剂到100毫升的预冷缓冲器 a. 将细菌颗粒添加到缓冲液中, 用磁搅拌杆搅拌并通过吹打上下无线来融汇悬浮液。无菌10毫升吸管 (~ 30 分钟)。
- 将细菌悬浮在50毫升一次性塑料管中整除数40毫升。油脂实验每个整除在水/冰批为细胞裂解 (70% 振幅, 总超声波时间5分钟, 间隔三十年代超声波, 三十年代暂停)。
注意: 在超声波步骤中, 样品不加热是很重要的。建议在冰浴中加入一些水, 以改善超声波期间的散热。如果使用不同的仪器, 超声波过程可能必须被适应。其他裂解方法, 如法国媒体或 microfluidizer 应该工作, 并可能有利于避免加热样品和蛋白质聚集。这些设备没有在我们的实验室, 我们取得了良好的结果, 我们的超声波协议。 - 取50µL 的裂解液为月桂酸钠的聚丙烯酰胺凝胶电泳 (SDS 页) 分析。离心样品 (18000 x g, 4 °c, 2 分钟) 和吸管的可溶性分数在一个新的试管。用吸管并用重悬50µL 的不溶性分数。将样品放在冰上直到 SDS 页分析 (步骤 2.6)。
- 离心法澄清裂解液 (39191 x g, 4 °c, 60 分钟)。
- 在离心步骤中, 通过 SDS 页分析细胞裂解步骤。将50µL 的2x 负载染料分别添加到该裂解物的可溶性和不溶性馏分中。热量为5分钟在95摄氏度和负载2µL 在15% 凝胶, 而仍然热。根据制造商的指示, 在 180 v 上用考马斯染料染色凝胶90分钟。将结果与图 1C中的代表结果进行比较。
- 过滤上清 (0.45 µm, 注射器过滤器)。抽取20µL 的过滤上清液样本进行 SDS 页分析 (步骤 2.11)。
注: 一般情况下, 可获得90至100毫升的澄清和过滤上清液。通常, 3 注射器过滤器是充足的。如果过滤很繁琐, 尽量增加离心速度和/或时间。 - 用固定化的金属亲和层析 (IMAC) 在快速性能的液相色谱 (FPLC) 系统上, 将 His6-SUMO Httex1 融合蛋白从澄清的细菌裂解物中分离出来, 在4°c37。
- 将澄清的裂解液填入 superloop, 并载入镍 NTA 柱上 (剥离, 根据制造商的手册进行清洗和重新加载, 建议使用以前的空白运行), 2 毫升/分. 通过10列容积 (CV, 200 毫升) 的缓冲 a 在10毫升/分钟洗涤了未绑定的蛋白质。
- 洗脱融合蛋白的 2.5 CV (50 毫升) 100% 的缓冲 B 在2毫升/分钟. 使用一小部分大小的50毫升的装载和洗涤和5毫升洗脱。将结果与图 1D中的代表结果进行比较。
- 对 SDS 页分析 (20 µL) 中的每一个分数取样, 并根据 IMAC 色谱的峰值将含有融合蛋白的分数进行池。添加 (2 s, 3 s)-14-双 (sulfanyl) 丁烷-23 二醇 (德勤) 和 l-半胱氨酸 (最终浓度100毫米每) 作为粉末, 并通过轻轻翻转管溶解。
注: 在我们的经验, 融合蛋白的纯度在不同的分数是可比的。为了防止高浓度分数的聚集, 应在 IMAC 后快速汇集纯化融合蛋白的组分。此外, 建议直接进行相扑标记的裂解和高效液相色谱纯化。如有必要, 可在此处停止该协议。熔融蛋白的稀释溶液被冷冻在液氮中, 在-80 摄氏度内保存, 在解冻后提纯, 不显著降低产量。将融合蛋白的稀释溶液贮存在4摄氏度的24小时内, 也能得到类似的结果。 - 通过 SDS 页面对 IMAC 进行分析。在每个样品上添加20µL 的载染料。在15% 凝胶上加载2µL 的粗材料 (2.7)、未绑定分数、洗涤分数和洗脱峰的每一个部分。根据制造商的指示, 在 180 v 上用考马斯染料染色凝胶90分钟。将结果与图 1D中的代表结果进行比较。
3. 他的6-相扑标记和 HPLC 纯化的解理
注意: 三氟乙酸酸 (TFA) 是一种挥发性液体, 可以引起严重烧伤, 所以要小心处理。在通风罩内进行所有处理, 并佩戴适当的个人防护设备 (即一次性丁腈橡胶手套、安全眼镜和实验室大衣)。
- 在一个5升瓶, 添加5毫升的 TFA 与一个塑料注射器到5升水 (溶剂 a: H2O, 0.1% (TFA)。用塑料注射器将2.5 毫升的 TFA 加入乙腈的2.5 升瓶 (溶剂乙: 乙腈, 0.1% TFA)。
- 按照制造商的建议准备高效液相色谱系统。执行空白运行以确保列干净。
- 在添加 ULP1 前, 取100µL 的融合蛋白样本, 以监测 UPLC 的裂解反应 (步骤 3.5)。
- 将20毫升的融合蛋白转移到新的50毫升管中, 加入0.4 毫升的 ULP1 溶液, 在冰上孵化。把剩下的融合蛋白放在冰上。
注: 他标记的催化片段403-621 的泛素样特异蛋白酶 1 (这里称为 "ULP1") 被用来切割相扑标记。融合蛋白比劈裂 Httex1 更稳定。建议不要将整个批次的相扑标记切割。相反, 继续整除数的大小, 可以直接和完全适用于 HPLC 柱。 - 每10分钟, 取样100µL 的裂解反应, 以监测进展的超效液相色谱 (UPLC)。离心机样品 (18000 rpm, 4 °c, 2 分钟) 和分析2µL 的清液由 UPLC (梯度从10% 到90% 溶剂 B 在 A 0.25 到3分钟, 10% B 为1分钟, 参考制造商的指示为仪器使用)。比较在添加 ULP1 前和样品后所获得的图谱。将结果与图 2B中的代表结果进行比较。
- 一旦相扑结束 (融合蛋白的峰值已经消失在 UPLC 色谱, 并完全转化为新出现的相扑和 Httex1 峰), 筛选的样本与注射器过滤器 (0.22 µm)。
注: 相扑劈裂通常是非常快的 (10-20 分钟在4°c) 因此 UPLC 分析与运行时间4分钟是一个有价值的工具监测反应。在高效液相色谱纯化前对样品进行过滤, 主要是提高柱体寿命的预防措施。样品不应变混浊。 - 反相色谱法纯化过滤样品 (溶剂 a 中25-35% 溶剂 B 的梯度, 40 分钟15毫升/分钟 (0-10 分钟: 5%; 10-12.5 分钟: 5 至 25%; 12.5-52.5 分钟:25 至 35%; 52.5-57.5 分钟35至 95%; 有关仪器使用的说明书, 请参阅制造商的说明)。将结果与图 2C中的代表结果进行比较。
注: Httex1 和他的6-相扑分离井由反相色谱法。然而, 在峰值的开始和结束时, 可能会有少量的截断 Httex1。收集小的分数, 以获得最大数量的纯材料。
注意: 在处理低温液体时, 应使用适当的安全设备 (如实验室外套、绝缘手套和面罩)。 - 电喷雾电离质谱法分析 HPLC 法 (ESI-MS, autosampler, 注入10µL, 流量0.6 毫升/分钟, 溶剂: 20% B 在 A, 没有列, 参考制造商的指示仪器使用) 和 UPLC (梯度从10% 到90%溶剂 B 在 A 为0.25 到3分钟, 10% B 为1分钟, 参考制造商的指示为仪器使用)。50毫升塑料管中类似纯度的池馏分, 在液氮和 lyophilize 中冷冻。将冻干蛋白重调成2毫升塑料管, 贮存在-20 摄氏度。
- 用 UPLC、ESI-MS 和 SDS 页表征纯化材料。将100µg 冻干 Httex1 在1.5 毫升管中的8µL 中溶解, 在封闭试管中的室温下孵化20分钟。小心地用氮气或氩气将 TFA 在通风罩下蒸发。使用低压氮气/氩气, 以避免损失的样品。
- 将100µL 中的蛋白质分解为2o, 用 UPLC 和5µL 分析2µL, 如步骤3.8。将20µL 的蛋白质溶液与20µL 的2x 负载染料混合。
- 通过 SDS 页面分析1µg 到10µg 的数量。根据制造商的指示, 在 180 v 上用考马斯染料染色凝胶90分钟。将结果与图 2D中的代表结果进行比较。
4. Httex1 蛋白的分类和 Resolubilization
注意: TFA 是一种挥发性液体, 可以引起严重烧伤, 所以要小心处理。在通风罩内进行所有处理, 并佩戴适当的个人防护设备 (即一次性丁腈橡胶手套、安全眼镜和实验室大衣)。
- 在2毫升管中制备10毫升的 Dulbecco 磷酸盐缓冲盐水 (DPBS) (137 毫米氯化钠, 2.7 毫米氯化钾, 10 毫米 Na4HPO2, 2毫米.在每次使用前通过0.2 µm 过滤器筛选 DPBS 解决方案。
- 将150µg 冻干 Httex1 在1.5 毫升管中的12µL 中溶解, 在封闭试管中的室温下孵化20分钟。小心地用氮气或氩气将 TFA 在通风罩下蒸发。使用低压氮气/氩气, 以避免损失的样品38。
注意: 一般情况下, 使用4µL TFA 溶解和分解50µg 的蛋白质。本程序将在试管内壁上创建一层蛋白质膜。为了防止在以下步骤中立即聚合 Httex1, 请使用预冷却的缓冲, 保持蛋白质始终放在冰上, 避免高浓度。 - 将分类蛋白分解为1毫升预冷 DPBS, 并将 pH 值调整为 7.2-7.4 与1米氢氧化钠。过滤蛋白溶液通过 100 kDa 离心过滤器成1.5 毫升塑料管 (20000 x g, 4 °c, 20 分钟)。
注: 计算的 Httex1 的理论浓度高于预期的最终浓度, 以解释可能的损失。过滤步骤是必要的, 以消除任何聚集, 可能已形成在溶解的蛋白质。 - 采用基于氨基酸分析 (214) 的 UPLC 标定曲线确定 Httex1 浓度, 并将蛋白质的2µg 送往氨基酸分析, 以验证浓度10。计算需要添加的 DPBS 量, 以获得3µM Httex1 的浓度。
- 将所计算的 DPBS 量添加到试管中, 将蛋白质稀释至3µM。将管放在冰上直到聚合协议启动。
注意: Httex1 43Q 不应存储在解决方案中。总是根据上面的协议准备一个新的蛋白质溶液。Httex1 蛋白最好储存在20°c 的冻干粉中。
5. 利用 UPLC 和圆二色谱 (CD) 光谱技术监测 Httex1 43Q 的聚集动力学, 并通过透射电镜 (TEM) 对骨料进行表征
- 为 TEM 制备甲酸铀溶液, 如前所报告的39。
- 通过在 DPBS 37 °c 中孵化3µM 溶液, 开始聚合 Httex1 43Q (使用在分类协议中所述的1毫升溶液)。
注: Httex1 的聚集可以根据实验的需要和目的进行更高浓度的聚合。 - 用 UPLC 在指示的时间点 (0、1、2、4、6、8、12、24、48和 120 h) 量化可溶性蛋白质的数量。要做到这一点, 采取整除35µL 和去除不溶性骨料离心 (20000 x g, 4 °c, 20 分钟)。将上清的4µL 注入 UPLC。利用仪器软件40计算峰值面积变化的可溶性单体比例。将结果与图 3A中的代表结果进行比较。
- 用0和 48 h 的 CD 光谱学表征二级结构的变化。采取整除100µL 和测量椭圆 (1 毫米石英试管, 195 毫微米到250毫微米, 20 °c, 数据点每0.2 毫微米, 速度10毫微米/分钟, 数字积分时间 2 s, 带宽1.0 毫微米)。获得6光谱的样品和平均和平滑使用一个二项式过滤器, 卷积宽度为99。绘制光谱作为平均残留摩尔椭圆 (θ)41.将结果与图 3B中的代表结果进行比较。
- 用 TEM 表征了骨料的结构和形态特征。将3µL 的蛋白质溶液放在 Formvar/碳包覆的200目上, 辉光放电铜网格为1分钟. 用 15 ul 水清洗网格两次, 一次与 15 ul 0.7% (w/v) 甲酸铀和染色三十年代与15瓦特/v 铀酰甲酸。对网格进行 TEM 分析。将结果与图 3C中的代表结果进行比较。
结果
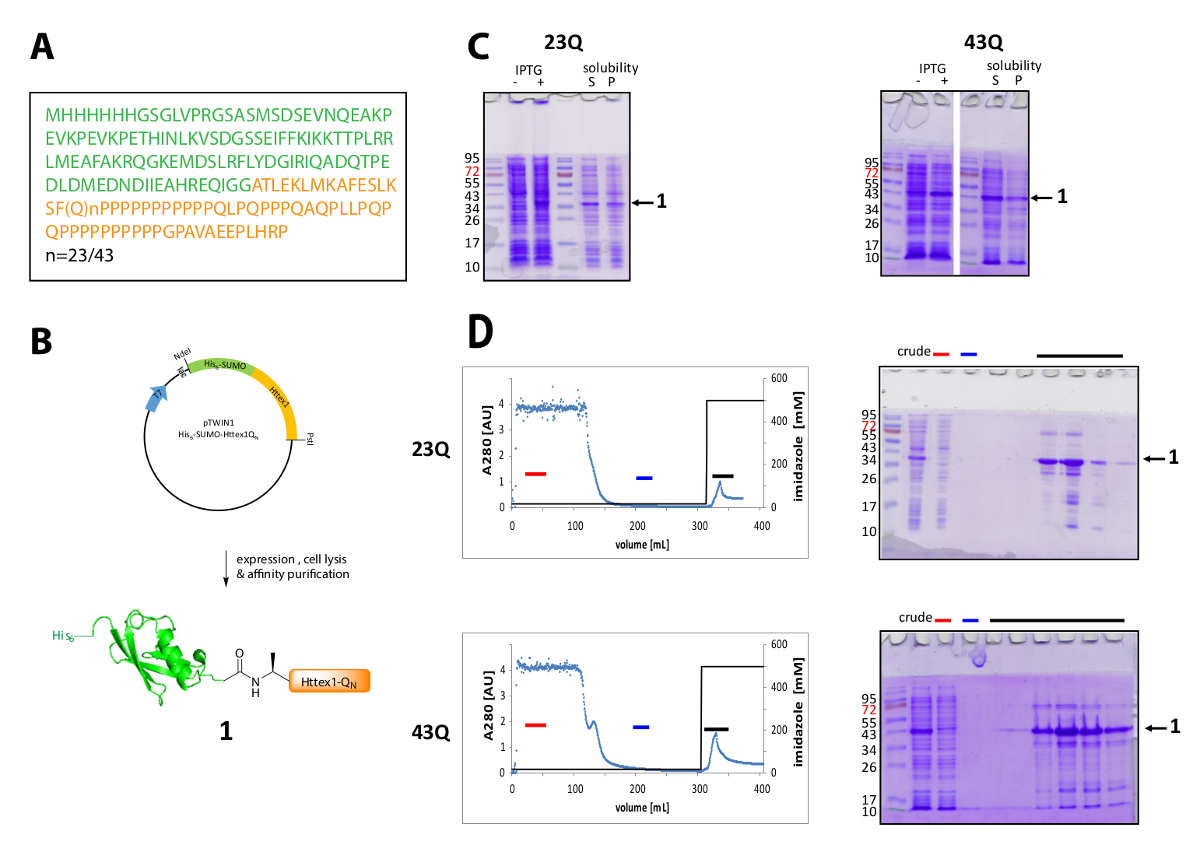
Httex1 在大肠杆菌中表达, 他的6相扑标记为 N 终端。图 1总结了融合蛋白的表达和纯化的代表性结果。Httex1 的序列由 Htt 的残留物2-90 组成, 从 Ala2 开始, 因为 Met1在体内42完全被劈裂。氨基酸的编号是指23Q 变种, 表达融合蛋白的完整序列如图 1所示。该质粒将在不久的将来存入 Addgene, 供社会人士分享。图 1B显示了质粒和表达的融合蛋白的示意图。His6-SUMO Httex1 表示在中等水平 (图 1C) 和大多数融合蛋白是存在的可溶性分数后溶解, 两者为23Q 和43Q 变种。根据分子量, 融合蛋白迁移比预期高。这部分是由于相扑的强烈褶皱, 但主要是由于 Httex1 的异常序列组成, 主要含有谷氨酰胺和脯氨酸残留物。wildtype (23Q) 和突变体 (43Q) 融合蛋白均可通过 IMAC (图 1D) 富集至80% 纯度, Httex1 和大样本的表达水平比较低, 可以解释共纯化宿主蛋白的存在性。应用于该列的卷。
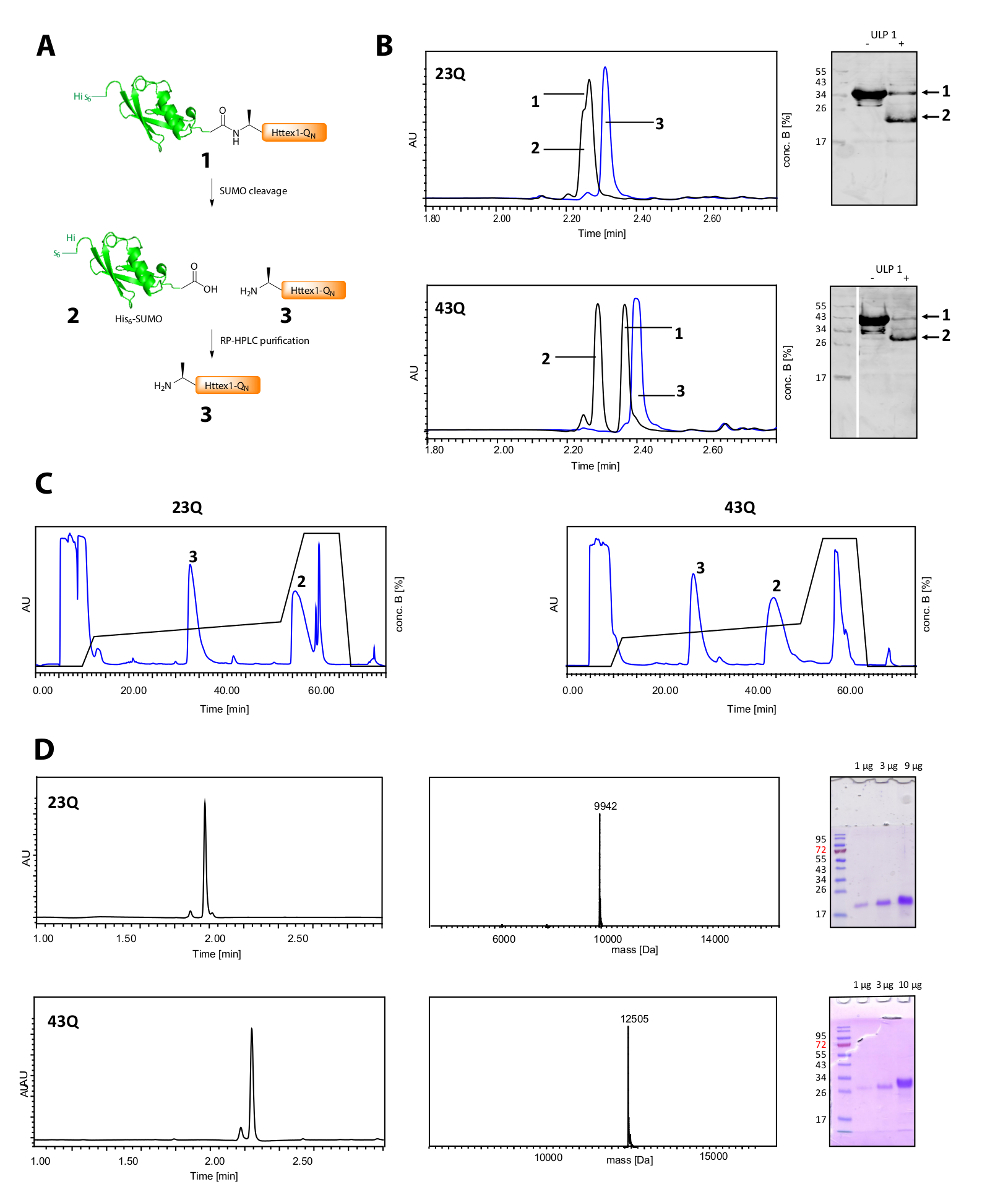
在图 2A中, 他的6相扑标记的裂解和 Httex1 的纯化。UPLC 是一个有效的工具, 以监测他的6相扑标签 (图 2B) 的分裂。融合蛋白的原始峰值被消耗, 两个新的和良好分离的峰值对应他的6相扑标记和 Httex1 出现。劈裂反应在10-20 分钟内完成。西方印迹 (WB) 的速度太慢, 无法有效地监测劈裂反应, 但它已被列入图中以供参考, 并证明了相扑卵裂的完整性。Httex1 23Q 和43Q 都可以从他的6相扑标记的反相色谱 (图 2C), 并获得了高纯度, 如 UPLC, MS 和 SDS 页分析 (图 2D)。
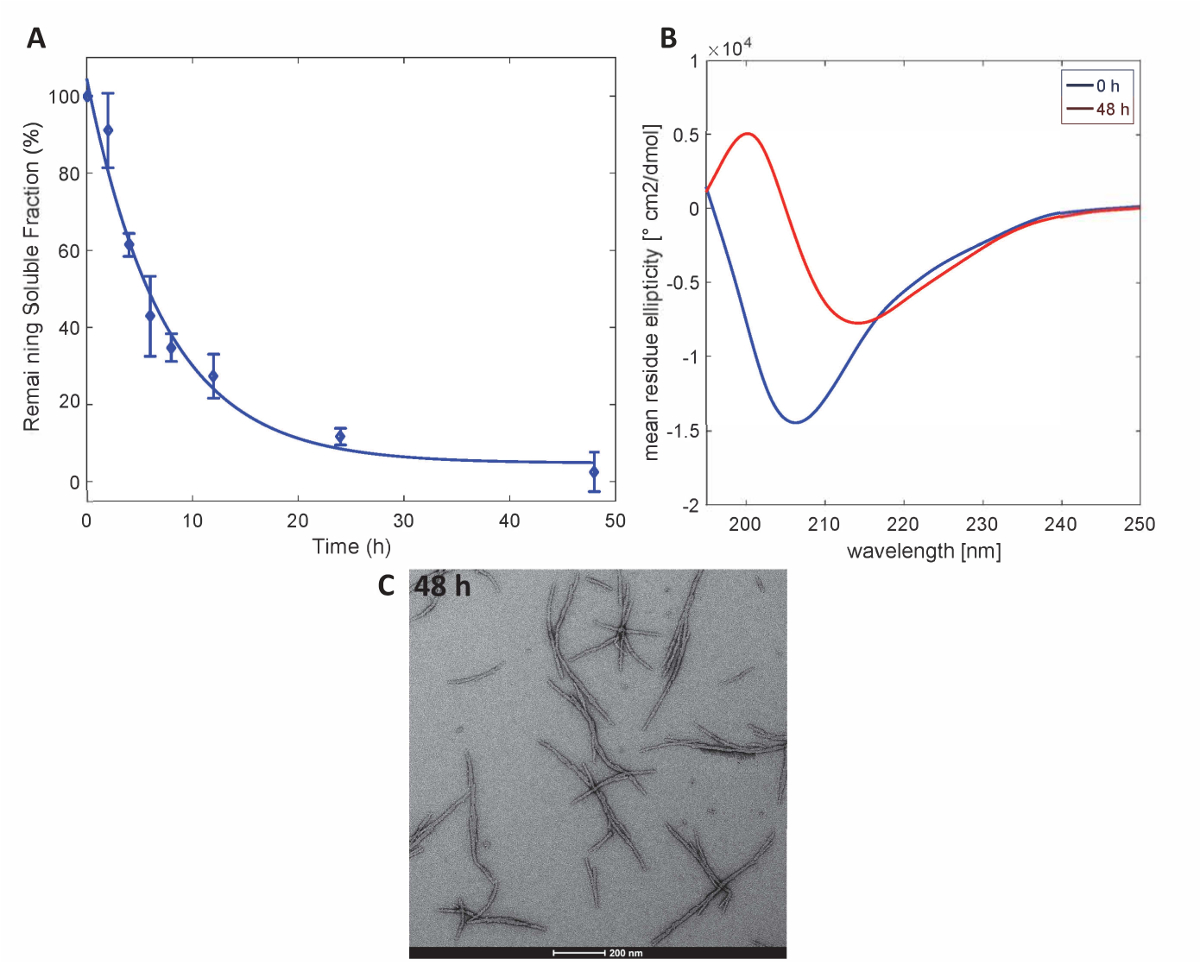
为了说明该方法制备的 Httex1 蛋白保留了 Httex1 的预期聚集特性, 我们用沉淀法对37°c 突变 Httex1 的 fibrillization 动力学进行了评估, 并通过 CD 对二次结构的变化进行了监测。光谱学, 并以透射电镜表征骨料的形态。在图 3a中, mHttex1 纤维形成的聚集动力学具有代表性的数据集, 由沉淀法确定。随着时间的推移, 可溶性 Httex1 43Q 的损失, 由于纤维的形成被 UPLC 量化。在48小时的孵化后, 我们观察到可溶性蛋白的完全耗尽。此外, 我们通过 CD 光谱学确定了蛋白质的二级结构 (图 3B)。Httex1 43Q 从非结构化 (λ最小205 毫微米) 转移到主要β表丰富构象 (λ极小215 毫微米) 在48小时的孵化之后。这一结构变化伴随着长纤维骨料的形成, 经 TEM 观察可观测到48小时 (图 3C)。

图 1.6-相扑 Httex1 融合蛋白的表达及纯化.
(A) 他的6-SUMO-Httex1-QN融合构造的氨基酸序列 (他的6-相扑在绿色和 Httex1-QN在橙色);(B)融合蛋白的表达和纯化示意图;(C) 在裂解后融合蛋白的表达和溶解度的 SDS 页分析;(D) 融合蛋白的 IMAC 纯化和分数的分析的色谱 (红色条: 未绑定分数, 蓝色条: 洗涤分数, 黑色条: 含洗脱峰的分数);请单击此处查看此图的较大版本.
{kind=link}

图 2.他的6相扑标记和纯化无标记 Httex1-QN蛋白的裂解。
(A) 示意图概述;(B) 对相扑标记与 ULP1 的 UPLC 的分析 (蓝色: 在添加 ULP1 之前; 黑色:20 分钟 (23Q), 分别为10分钟 (43Q) 后添加 ULP1) 和世界银行 (MAB5492 1:2000, 次级山羊抗鼠抗体 1:5000);(C) 制备反相色谱法纯化 Httex1;D: UPLC、SDS 页和 ESI-MS 对纯化 Httex1 的分析;预期分子量分别为 9943 da (23Q) 和 12506 da (43Q)。请单击此处查看此图的较大版本.
{kind=link}

图 3.Httex1-43Q 的聚集: (A) 基于 UPLC 的沉淀试验。(B) 二级结构的 CD 光谱为0小时和 48 h. (C) 在 48 h (鳞片条为 200 nm) 的聚合体的 TEM 显微照片。请单击此处查看此图的较大版本.
{kind=link}
讨论
在本议定书中, 我们概述了获取含有23或43谷氨酰胺残留物的本机、未加标签 Httex1 毫克数量的有效程序。这是通过表达 Httex1 作为一个 C 终端融合到一个他的6相扑标记, 这是用来分离融合蛋白从细胞裂解物的 IMAC, 并在 HPLC 纯化之前的 Httex1。虽然相扑战略已经用于生产的其他几个蛋白质, 我们的方法表明, 独特的特性相扑也可以用来产生内在紊乱, 聚集容易, amyloidogenic 蛋白, 以前证明是极难处理和生产43,44。我们提出了一种简单、易于使用的协议, 可与生成 "行为良好" 的蛋白质的协议相媲美。相扑融合 solubilizes 和稳定 Httex1 在表达和 IMAC 净化步骤。标记的过早的分裂, 如观察与蛋白质内含子战略10和汇集不再是问题。
内在紊乱的蛋白质特别容易降解。虽然 N17 地区的 n-末端退化不是使用此协议的问题, 但截断在珠三角 Httex1 可能发生。由于截断蛋白与 Httex1 在疏水性、电荷和大小等方面非常相似, 因此用色谱法去除它们是很有挑战性的, 因此最好首先防止它们的形成。严格遵守该议定书, 总是在冰上工作, 使用足够数量的蛋白酶抑制剂, 将有助于保持观察到的截断水平非常低。在 Httex1 的 C 端应用融合标记可以很容易地去除珠三角的截断, 因为截断后的蛋白质也会失去亲和标记。但是, 如果需要维护本机序列, 则此选项不能作为 Httex1 以脯氨酸结束而应用, 而在我们最了解的情况下, 没有已知能诱导无影无踪和有效的脯氨酸后裂解的 C 终端融合标记。
该协议最关键的部分是处理的 Httex1 释放后, 相扑标记的 ULP1。蛋白质应立即通过反相色谱法纯化。幸运的是, 这是一种高效快速的反应, 通常是在4摄氏度的10-20 分钟内完成的。相比之下, 蛋白质内含子策略需要几个小时才能完全解切蛋白质内含子, 因此需要在不完全的解理和开始聚集之间进行权衡, 以最大限度地提高产量。突变 Httex1 需要快速的检查, 因为它将开始聚集在相对较高浓度存在的裂解反应, 而23Q 变种是稳定的较长的时间。在反相色谱纯化过程中, 相扑的另一优点是明显的: 当Ssp DnaB 蛋白质内含子是疏水性和强烈的柱状, 相扑是更亲水和 elutes 完全从 C4 反相柱。虽然商业 ULP1 成本相当高, 但在以前发布的协议29中, 这种蛋白质可以很容易地产生高产。
在使用 Httex1 之前应用分类协议的关键重要性是不够强调的。冻干 polyQ 蛋白如 Httex1 是稳定的, 可以储存长时间, 但不能完全溶于水和缓冲。预制寡聚物或纤维的存在对蛋白质45的聚集动力学和生物物理特性有显著的影响。此处描述的分类协议允许对蛋白质进行分类, 去除预制骨料, 并从冻干样品中生成单体 Httex1 溶液。观察了 Httex1 与蛋白质内含子的相似聚集动力学和纤维形态学。
与以前生产 Httex1 的方法相比, 这里描述的相扑策略提供了一些优势, 并扩大了可能的研究范围, 以研究这种蛋白质在健康和疾病中的结构和功能特性。SUMO-Httex1 融合蛋白易于处理, 可在常温下冷冻保存或保存在溶液中24小时, 而自由 mHttex1 会快速聚合。稳定性和高溶解度的 SUMO-Httex1 融合蛋白提供了更大的灵活性, 以操纵蛋白质和/或引入酶和化学改性到 mHttex1, 否则将无法在卵裂后。这包括引入翻译后的修改, 显影, 自旋标签, 生物素标签等.这里提出的进展应 1) 促进今后的研究, 以阐明 Httex1 的结构功能关系;2) 生成新的工具, 以调查 Htt 聚集和病理传播;3) 能够开发新的化验, 以识别稳定突变体 Httex1 的分子, 防止其聚集;4) 鼓励来自其他领域的科学家来研究这种蛋白质, 并加入我们的探索, 寻找亨廷顿病的治疗方法。
披露声明
作者声明他们与本文的内容没有利益冲突。
致谢
这项工作的经费主要来自 CHDI 基金会和瑞士国家科学基金会的赠款。我们感谢索菲 Vieweg 博士在开发这个新的表达系统和 Lashuel 小组的其他成员时进行了有益的讨论, 以分享他们在这个表达系统中的经验以及他们的投入和宝贵的反馈。我们还感谢奥利弗 Hantschel 教授提供 ULP1 质粒。作者感谢约翰 b. 华纳博士和 Senthil 博士 Thangaraj 对手稿的批判性审查
材料
| Name | Company | Catalog Number | Comments |
| Uranyl formate (UO2(CHO2)2) | EMS | 22450 | |
| Formvar/carbon 200 mesh, Cu 50 grids | EMS | FCF200-Cu-50 | |
| High Precision Cell made of Quartz SUPRASIL 1 mm light path from Hellma Analytics | HellmaAnalytics | 110-1-40 | |
| Buffer Substance Dulbecco's (PBS w/o Ca and Mg) ancinne ref 47302 (RT) SERVA | Witech | SVA4730203 | |
| Ampicillin | AxonLab | A0839.0100 | |
| Luria Broth (Miller's LB Broth) | Chemie Brunschwig | 1551.05 | |
| Isopropyl β-D-1-thiogalactopyranoside (IPTG) | AxonLab | A1008.0025 | |
| E. coli B ER2566 | NEB | NEB# E4130 | |
| Imidazole | Sigma | 56750-500G | |
| cOmplete Protease Inhibitor Cocktail | Roche | 4693116001 | |
| Anti-Huntingtin Antibody, a.a. 1-82 | Merck Millipore Corporation | MAB5492 | |
| IRDye 680RD Goat anti-Mouse IgG (H + L) | Licor | 925-68070 | |
| PMSF | AxonLab | A0999.0005 | |
| HisPrep 16/10 column | GE Healthcare | 28936551 | |
| C4 HPLC column | Phenomenex | 00G-4168.P0 | 10 µm C4 300 Å, LC Column 250 x 21.2 mm, Phenomenex, 19x10 mm guard column, not temperature jacketed |
| Acetonitrile HPLC | MachereyNagel | C2502 | |
| Filtre seringue Filtropur S 0,45 ul sans prefiltre sterile | Sarstedt AG | 83.1826 | |
| Spectrophotometer semi-micro cuvette | Reactolab S.A. | 2534 | |
| Superloop, 1/16" fittings (ÄKTAdesign), 50 ml | GE Healthcare | 18111382 | |
| Trifluoroacetic acid | Sigma | 302031 | |
| GREINER Tubes fo FPLC 16 x 100 mm, cap. 12.0 ml | Greiner Bio-One | 7.160 102 | |
| 100 kD Microcon fast flow filters | Merck Millipore Corporation | MRCF0R100 | |
| Vibra-cell VCX130 ultrasonic liquid processor | Sonics | ||
| Äkta 900 equipped with a fraction collector | GE Healthcare | ||
| Jasco J-815 Circular Dichroism | Jasco | ||
| Waters UPLC system | Waters | C8 BEH acquity 2.1x100 mm 1.7 micron column , preheated column (40 °C), flow rate of 0.6 mL/min, injection volume of 4 μL | |
| waters HPLC system | Waters | 2489 UV detector and 2535 quaternary gradient module, 20 mL loop in a FlexInject housing | |
| ESI-MS: Finnigan LTQ | Thermo Fisher Scientific | ||
| lyophylizer instrument | FreeZone 2.5 Plus | ||
| Tecnai Spirit BioTWIN | FEI | electron microscope equipped with a LaB6 gun and a 4K x 4K FEI Eagle CCD camera (FEI) and operated at 80 kV | |
| 37 °C shaking incubator | Infors HT multitron Standard | ||
| Biophotometer plus | Eppendorf |
参考文献
- Saudou, F., Humbert, S. The Biology of Huntingtin. Neuron. 89 (5), 910-926 (2016).
- MacDonald, M. E., Gines, S., Gusella, J. F., Wheeler, V. C. Huntington's disease. Neuromolecular Medicine. 4 (1-2), 7-20 (2003).
- Li, S., Li, X. J. Multiple pathways contribute to the pathogenesis of Huntington disease. Molecular Neurodegeneration. 1, 19 (2006).
- DiFiglia, M. Aggregation of Huntingtin in Neuronal Intranuclear Inclusions and Dystrophic Neurites in Brain. Science. 277 (5334), 1990-1993 (1997).
- Atwal, R. S., et al. Huntingtin has a membrane association signal that can modulate huntingtin aggregation, nuclear entry and toxicity. Human Molecular Genetics. 16 (21), 2600-2615 (2007).
- Sathasivam, K., et al. Aberrant splicing of HTT generates the pathogenic exon 1 protein in Huntington disease. Proceedings of the National Academy of Sciences U S A. 110 (6), 2366-2370 (2013).
- Mangiarini, L., et al. Exon 1 of the HD gene with an expanded CAG repeat is sufficient to cause a progressive neurological phenotype in transgenic mice. Cell. 87 (3), 493-506 (1996).
- El-Daher, M. T., et al. Huntingtin proteolysis releases non-polyQ fragments that cause toxicity through dynamin 1 dysregulation. EMBO Journal. 34 (17), 2255-2271 (2015).
- Lunkes, A., et al. Proteases acting on mutant Huntingtin generate cleaved products that differentially build up cytoplasmic and nuclear inclusions. Molecular Cell. 10 (2), 259-269 (2002).
- Vieweg, S., Ansaloni, A., Wang, Z. M., Warner, J. B., Lashuel, H. A. An Intein-based Strategy for the Production of Tag-free Huntingtin Exon 1 Proteins Enables New Insights into the Polyglutamine Dependence of Httex1 Aggregation and Fibril Formation. Journal of Biological Chemistry. 291 (23), 12074-12086 (2016).
- Georgalis, Y., et al. Huntingtin aggregation monitored by dynamic light scattering. Proceedings of the National Academy of Sciences U S A. 95 (11), 6118-6121 (1998).
- Scherzinger, E., et al. Huntingtin-encoded polyglutamine expansions form amyloid-like protein aggregates in vitro and in vivo. Cell. 90 (3), 549-558 (1997).
- Scherzinger, E., et al. Self-assembly of polyglutamine-containing huntingtin fragments into amyloid-like fibrils: implications for Huntington's disease pathology. Proceedings of the National Academy of Sciences U S A. 96 (8), 4604-4609 (1999).
- Muchowski, P. J., et al. Hsp70 and hsp40 chaperones can inhibit self-assembly of polyglutamine proteins into amyloid-like fibrils. Proceedings of the National Academy of Sciences U S A. 97 (14), 7841-7846 (2000).
- Heiser, V., et al. Inhibition of huntingtin fibrillogenesis by specific antibodies and small molecules: implications for Huntington's disease therapy. Proceedings of the National Academy of Sciences U S A. 97 (12), 6739-6744 (2000).
- Bennett, E. J., Bence, N. F., Jayakumar, R., Kopito, R. R. Global impairment of the ubiquitin-proteasome system by nuclear or cytoplasmic protein aggregates precedes inclusion body formation. Molecular Cell. 17 (3), 351-365 (2005).
- Tam, S., et al. The chaperonin TRiC blocks a huntingtin sequence element that promotes the conformational switch to aggregation. Nature Structural & Molecular Biology. 16 (12), 1279-1285 (2009).
- Nekooki-Machida, Y., et al. Distinct conformations of in vitro and in vivo amyloids of huntingtin-exon1 show different cytotoxicity. Proceedings of the National Academy of Sciences U S A. 106 (24), 9679-9684 (2009).
- Wacker, J. L., Zareie, M. H., Fong, H., Sarikaya, M., Muchowski, P. J. Hsp70 and Hsp40 attenuate formation of spherical and annular polyglutamine oligomers by partitioning monomer. Nature Structural & Molecular Biology. 11 (12), 1215-1222 (2004).
- Legleiter, J., et al. Monoclonal antibodies recognize distinct conformational epitopes formed by polyglutamine in a mutant huntingtin fragment. Journal of Biological Chemistry. 284 (32), 21647-21658 (2009).
- Legleiter, J., et al. Mutant huntingtin fragments form oligomers in a polyglutamine length-dependent manner in vitro and in vivo. Journal of Biological Chemistry. 285 (19), 14777-14790 (2010).
- Nucifora, L. G., et al. Identification of novel potentially toxic oligomers formed in vitro. from mammalian-derived expanded huntingtin exon-1 protein. Journal of Biological Chemistry. 287 (19), 16017-16028 (2012).
- Dahlgren, P. R., et al. Atomic force microscopy analysis of the Huntington protein nanofibril formation. Nanomedicine. 1 (1), 52-57 (2005).
- Poirier, M. A., et al. Huntingtin spheroids and protofibrils as precursors in polyglutamine fibrilization. Journal of Biological Chemistry. 277 (43), 41032-41037 (2002).
- Duim, W. C., Chen, B., Frydman, J., Moerner, W. E. Sub-diffraction imaging of huntingtin protein aggregates by fluorescence blink-microscopy and atomic force microscopy. Chemphyschem. 12 (13), 2387-2390 (2011).
- Pieri, L., Madiona, K., Bousset, L., Melki, R. Fibrillar alpha-synuclein and huntingtin exon 1 assemblies are toxic to the cells. Biophysical Journal. 102 (12), 2894-2905 (2012).
- Monsellier, E., Redeker, V., Ruiz-Arlandis, G., Bousset, L., Melki, R. Molecular interaction between the chaperone Hsc70 and the N-terminal flank of huntingtin exon 1 modulates aggregation. Journal of Biological Chemistry. 290 (5), 2560-2576 (2015).
- Isas, J. M., Langen, R., Siemer, A. B. Solid-State Nuclear Magnetic Resonance on the Static and Dynamic Domains of Huntingtin Exon-1 Fibrils. Biochemistry. 54 (25), 3942-3949 (2015).
- Malakhov, M. P., et al. SUMO fusions and SUMO-specific protease for efficient expression and purification of proteins. Journal of Structural Function Genomics. 5 (1-2), 75-86 (2004).
- Mossessova, E., Lima, C. D. Ulp1-SUMO crystal structure and genetic analysis reveal conserved interactions and a regulatory element essential for cell growth in yeast. Molecular Cell. 5 (5), 865-876 (2000).
- Kumari, S., Pal, R. K., Gupta, R., Goel, M. High Resolution X-ray Diffraction Dataset for Bacillus licheniformis Gamma Glutamyl Transpeptidase-acivicin complex: SUMO-Tag Renders High Expression and Solubility. Protein Journakl. 36 (1), 7-16 (2017).
- Zhang, J., Sun, A., Dong, Y., Wei, D. Recombinant Production and Characterization of SAC, the Core Domain of Par-4, by SUMO Fusion System. Applied Biochemistry and Biotechnology. , (2017).
- Reif, A., et al. Semisynthesis of biologically active glycoforms of the human cytokine interleukin 6. Angewandte Chemie International Edition English. 53 (45), 12125-12131 (2014).
- Froger, A., Hall, J. E. Transformation of plasmid DNA into E. coli using the heat shock method. Journal of Visualized Experiments. (6), 253 (2007).
- Smith, B. J. SDS Polyacrylamide Gel Electrophoresis of Proteins. Methods in Molecular Biology. 1, 41-55 (1984).
- Lawrence, A. M., Besir, H. U. Staining of proteins in gels with Coomassie G-250 without organic solvent and acetic acid. Journal of Visualized Experiments. (30), (2009).
- Block, H., et al. Immobilized-metal affinity chromatography (IMAC): a review. Methods in Enzymology. 463, 439-473 (2009).
- Chen, S. M., Wetzel, R. Solubilization and disaggregation of polyglutamine peptides. Protein Science. 10 (4), 887-891 (2001).
- Booth, D. S., Avila-Sakar, A., Cheng, Y. Visualizing proteins and macromolecular complexes by negative stain EM: from grid preparation to image acquisition. Journal of Visualized Experiments. (58), (2011).
- O'Nuallain, B., et al. Kinetics and thermodynamics of amyloid assembly using a high-performance liquid chromatography-based sedimentation assay. Amyloid, Prions, and Other Protein Aggregates, Pt C. 413, 34-74 (2006).
- Greenfield, N. J. Analysis of circular dichroism data. Methods in Enzymology. 383, 282-317 (2004).
- Aiken, C. T., et al. Phosphorylation of threonine 3: implications for Huntingtin aggregation and neurotoxicity. Journal of Biological Chemistry. 284 (43), 29427-29436 (2009).
- Satakarni, M., Curtis, R. Production of recombinant peptides as fusions with SUMO. Protein Expression and Purification. 78 (2), 113-119 (2011).
- Davies, H. A., Wilkinson, M. C., Gibson, R. P., Middleton, D. A. Expression and purification of the aortic amyloid polypeptide medin. Protein Expression and Purification. 98, 32-37 (2014).
- Chen, S., Wetzel, R. Solubilization and disaggregation of polyglutamine peptides. Protein Science. 10 (4), 887-891 (2001).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。