
Method Article
斑马鱼光学杯形态形成的四维成像
摘要
本协议描述了斑马鱼早期眼睛发育的托托标记和多维成像方法。我们使用激光扫描共聚焦显微镜描述标签、嵌入和四维(4D)成像,以及优化数据集获取以解剖光学杯形态发生机制的考虑因素。
摘要
视觉系统功能要求建立精确的组织和器官结构。在脊椎动物眼睛中,结构缺陷是视力受损的常见原因,但眼形态机制仍不为人所知。胚胎眼的基本组织在整个脊椎动物中得到保护,因此斑马鱼胚胎的活成像已成为在正常病理条件下直接观察眼睛发育的有力途径。动态细胞过程,包括运动,形态,相互作用,分裂和死亡可以在胚胎中可视化。我们开发了亚细胞结构的均匀标记方法和斑马鱼早期眼睛发育的延时共聚焦显微镜。该协议概述了将封顶mRNA注射到1细胞斑马鱼胚胎、在光学囊泡阶段(受精后12小时)安装胚胎,以及在同一成像会话中对光学杯形态进行多维超时成像的方法,以便在同一成像会话中连续获得多个数据集。这种方法可产生可用于各种目的的数据,包括细胞跟踪、体积测量、三维 (3D) 渲染和可视化。我们的方法使我们能够精确定位细胞和分子机制,推动光学杯的发展,在野生类型和遗传突变条件下。这些方法可以直接被其他群体使用,也可以被改编为斑马鱼眼睛发育的许多其他方面。
引言
脊椎动物眼睛的发育始于未来大脑神经质的视囊的出现或回避。然后,光学囊泡会经历一系列组织形状的变化,拉长,然后侵入以产生光学杯。在光学杯中,神经视网膜和视网膜色素上皮,都来自神经蛋白,包裹新生的透镜,这是从表面外皮衍生的。整个过程需要一系列复杂的细胞和组织运动和分子信号,在神经细胞群、细胞皮质和间质细胞群之间协调。这些初始事件建立了眼睛的基本结构,眼睛发育的后期步骤,包括虹膜和角膜的形成,是早期组织的基础。早期眼睛发育和形态发育的破坏是人类许多视觉损伤状况的基础,包括眼科、微噬菌体和结肠瘤。解锁控制光学杯形态形成的细胞和分子机制对于进一步了解视觉系统的发展以及当这些过程出错时产生的病理条件至关重要。
我们对脊椎动物眼睛发育和形态生成的理解,从大量的工作中浮现出来,从经典的组织学研究到各种模型生物体的胚胎学和遗传方法,包括小鼠、小鸡、青蛙和鱼1、2、3、4、5。虽然这一系列工作建立了分子机制来调节早期眼睛发育,但从历史上看,对眼睛的形态形成存在一种不了解:其3D结构的出现。这些发现大部分来自离散时间点的成像分割胚胎。虽然这足以提供2维的组织形态视图,但形态生成是一个动态的3D过程。为了确定组织形状如何随着时间推移而变化,单个细胞的行为方式,以及这些行为如何促成 3D 组织形状的变化,需要采用不同的方法。
解决这一重大知识差距的一个解决方案是活成像,它使细胞和组织能够实时动态观察器官的形状。不幸的是,由于胚胎发育的限制,这在许多模型生物体中是不容易可行的。例如,小鼠和小鸡胚胎(在子宫内或蛋壳内发育)不容易获得,许多活胚胎在光学上不透明,导致大量光散射,并限制了图像的获取深度。斑马鱼,与外部发展和透明的胚胎,提供了一个独特的机会,进行活成像的眼睛形态6,7,8,9,10,11,12,13,14,15,16,17,18,19。此外,还提供充足的转基因和突变系,以及产生新的转基因和突变体的工具20,21,22,23,24。此外,光杯形态在斑马鱼中迅速发生,在12小时的时间范围内(施肥后12-24小时,hpf),使整个过程的成像可行。
荧光蛋白家族的扩展和优化加速了活成像工作,使亚细胞结构具有基因编码的重要标记,以及显微镜方法的改进和创新。这里描述的协议使用激光扫描共聚焦显微镜,而不是其他目前的方法成像斑马鱼胚胎生成,包括旋转盘共焦显微镜,选择性平面照明显微镜(SPIM及其变种),和其他更专业的显微镜方法。对于正在发育的斑马鱼眼睛,我们发现旋转盘共聚焦显微镜不足以在组织中进行更深入的成像。虽然 SPIM 拥有非常快的成像时间,并且使用越来越广泛,但处理用于可视化和分析的大型数据集仍然是一项挑战。相比之下,激光扫描共聚焦显微镜很容易获得,尤其是对于缺乏组装光学硬件专业知识的个人。我们希望激光扫描共聚焦显微镜的广泛可用性将使我们的协议对许多实验室有用。
在这里,我们描述了我们捕捉光学杯形态形成的4D数据集的方法 ,用于对 胚胎进行膜和色质的标记,以及用于图像采集的激光扫描共聚焦显微镜( 图1中化)。这里使用的荧光蛋白(EGFP-CAAX 和 H2A)。F/Z-mCherry) 被选中提供单细胞分辨率的组织标记。我们使用此协议生成的数据集进行各种图像分析和可视化功能。如果需要其他子细胞结构,则此协议可以轻松调整。等离子膜标签被选作细胞形状的可视化:我们使用EGFP-CAAX,其中最后21个H-ras氨基酸,作为预制信号序列,融合到EGFP13的C终点站。其他以荧光蛋白为靶点的血浆膜(如跨膜融合或肌酸化)也可能同样有效。为了标记核,我们选择了H2A。F/Z-mCherry,其中麦切里被融合到一个 histone 蛋白质13。这确保了细胞分裂,包括线粒体主轴方向,很容易可视化。
使用任何实时成像方法,必须考虑在提高成像速度和最大化信号与噪声比、轴向分辨率和样品保存之间权衡。我们优化了我们的方法,以最大限度地提高图像质量和胚胎数量,可以在一次运行中成像。通常,目标是在同源突变胚胎中成像光学杯形态,在视杯形态形成开始时,这种变异胚胎可能与野生类型在表型上没有区别,而异质基因的后代(25%的胚胎是所需的基因型)。通过优化,然后多路复用图像采集,捕获同源突变胚胎数据集的可能性增加。
时间分辨率(或卷数据(Z 堆栈)的获取频率是时间拉大成像的一个关键方面。根据此类数据集的目的,对速度有不同的要求。最初,此协议是针对手动 4D 单元格跟踪开发的。跟踪统一标记的组织内的单个细胞需要足够高的时间分辨率,以提供信心,任何一个细胞正在持续跟踪随着时间的推移。我们发现,斑马鱼光学杯形态的Z堆必须至少每3.1分钟获得一次超过12小时:在这里,我们优化了我们在激光扫描共聚焦显微镜上的采集,这样我们每2.5分钟就可以获得4-5个胚胎的Z堆栈。
建立 Z 步大小是协议优化的关键步骤:对于 3D 渲染和可视化,同位素数据是理想的,其中 Z 步大小等于 XY 像素尺寸。实际上,由于成像时间和光模糊的限制,很难通过实时样品获得此类超时数据。因此,确定足够的 Z 步大小对于实验的渲染和可视化需求非常重要,具体来说,需要什么 X:Y:Z voxel 比率来最大化轴向信息,同时保持速度并防止光出血。对于此协议,使用 40 倍长工作距离浸水目标确定的 voxel 比率为 1:1:3.5(0.6 μm x 0.6 μm x 2.1 μm Z 步尺寸)。当获得 130-140 μm 的 Z 深度时,这会产生具有适当时间分辨率和少量光模糊的体积数据。
如上所述,此协议是专门为斑马鱼光学杯形态的4D成像,使用胚胎在托托标记等离子体膜和染色质,以及激光扫描连续显微镜。下面的协议可以很容易地适应各种实验和需求。首先,对于子细胞结构,可以成像存在活细胞标记的任何结构。接下来,虽然这里的重点是光学杯形态生成,但定时成像可以适应眼部发育的其他阶段,例如神经发生25、26、27、28、29、30、31、32、33。对于成像后期发展,人们可能需要考虑胚胎固定(自发肌肉活动开始约24马力),色素运动(开始出现约24马力),组织大小(眼睛在神经生成过程中体积显著增长),和成像速度(应根据兴趣过程的速度进行调整)。所有这些考虑都可以很容易地加以管理。协议相当灵活:除了具体协议的细节,有一般原则,将帮助那些有兴趣现场成像眼睛发育的其他方面。
研究方案
这里描述的所有使用斑马鱼的方法都包含在Kristen Kwan博士的动物协议"斑马鱼视觉系统发展的细胞和分子机制"中,并得到了犹他大学机构动物护理和使用委员会(IACUC)的批准。
1. 盖住RNA合成
- 生成体外转录的DNA模板。
- 通过在反应混合物的 100 μL 体积中消化 10 微克的 DNA,将 DNA 模板线性化。典型的反应组合如下:使用 3 μL 的酶和 10 μL 的缓冲器消化 10 μg DNA,将反应体积与水一起达到 100 μL。
注:一个典型的模板消化是PCS2载体,例如,PCS2-EGFP-CAAX或PCS2FA-H2A.F/Z-mcherry的膜和色度素标签在这里描述。在这种情况下,质粒DNAs是每个消化使用酶诺蒂。用户应谨慎对待明星活动:在这种情况下,建议使用高保真度酶并提供商业服务。 - 在 37 °C 的夜间孵育反应,以确保 DNA 被消化到完成。
- 按照制造商的协议使用 PCR 净化套件清理限制摘要。将DNA与30微升ddH2O.一起,将线性DNA存储在-20°C,并根据需要使用。
注:消化的DNA量和脱氧核糖核酸量产生约0.3微克/μL;这足以满足 +5 轮体外转录, 每轮使用 +2 μg 的 Dna 作为模板。
- 通过在反应混合物的 100 μL 体积中消化 10 微克的 DNA,将 DNA 模板线性化。典型的反应组合如下:使用 3 μL 的酶和 10 μL 的缓冲器消化 10 μg DNA,将反应体积与水一起达到 100 μL。
- 使用体外转录套件进行体外转录封顶 RNA。对于 PC2 模板(如此处所述),请使用 SP6 套件。
- 组装体外转录反应,如下: 文摘 2 μg DNA 模板或高达 6 μL 与 2 μL 的 10 倍反应缓冲器, 10 μL 的 2x 核糖核苷酸混合, 和 2 μL 的 10 倍酶混合.
- 在 37 °C 下孵育 2-4 小时或更长时间(更长的时间将导致更高的产量)。如果需要,1微升的酶混合可以在潜伏期的中途补充。
- 通过添加 1 μL 的无 Rnase DNase 并在 37 °C 下孵育 15 分钟来消化 DNA 模板。
- 按照制造商的协议(参见 材料表),使用 RNA 净化套件净化封顶 RNA。Elute 与 100 微升的无 Rnase H2O 。
注:本应用不需要添加β-甲醇。虽然此协议中描述的方法很简单,但替代试剂也可用于净化 RNA。 - 沉淀RNA。
- 将 10 μL 的 3 M 无 RNase NaOAc 和 2.5 卷 (+275 μL) 的无冰冷无 Rnase 100% EtOH 添加到无冰冷 RNA 中。
- 将反应在 -20 °C 下孵化 15-30 分钟。在 4 °C 高速旋转 15 分钟,以颗粒 RNA。
注:颗粒应在管壁上可见。在此步骤中,请注意离心机中的管子是如何对齐的,以便预测颗粒在旋转结束时的位置。 - 用注射器小心地取出 EtOH,小心避免加热、过度干燥或脱落颗粒。在 20 微升的无 Rnase H2O 中重新使用颗粒。
- 检查RNA以确保合成成功。使用 1 μL 检测光谱仪上的浓度,在 1% 的玫瑰凝胶上运行 0.5 μL,以检查一个或两个离散频段,而不是低分子量涂片。体外转录反应的产量应为 ±1 μg/μL。
- 阿里库特和存储RNA在-20°C或-80°C。 直到注射前,RNA 才稀释。
2. 1细胞斑马鱼胚胎的微注射
注:每RNA注射200-300 pg,在整个光学杯形态形成过程中获得色素和细胞膜的无所不在的表达。准备 5-10 μL 的 RNA 稀释,并加载 2.5-5 μL/needle;计划额外的,如果针头断裂。
- 提前准备注射。
- 准备注射模具盘,以促进1细胞胚胎的对齐和定向进行微注射。在E3(标准胚胎介质)中融化2%的玫瑰,倒入培养皿中。小心地将注塑模具(参见 材料表)漂浮在热气糖的顶部,以在气精凝固时产生气垫槽的印记。一旦阿加罗斯凝固,取出注塑模具。
注:注:注塑盘可使用数月:覆盖在 E3 中,在不使用时存储在 4 °C。 - 拉微注射针。将玻璃毛细血管拉到长锥形器上,使用拔针机制作微注射针。对机器进行特定于所用毛细血管类型的编程。对于 1.0 x 0.78 mm 玻色硅酸盐毛细血管使用以下程序: 热 = 546 °C, 拉 + 130, 速度 = 70, 时间 = 90 (图 2F).将针头存放在培养皿中,并用建模粘土固定。
注意:拉力配方会有所不同,具体取决于机器和灯丝,并且应始终校准新细丝。每轮拉力产生两针(图2G):这样做几次,以产生足够的针头,以防意外中断和未来的实验。 - 稀释封顶RNA进行注射。使用无 RNase H 2 O 和 1 μL 的苯酚红色,将 EGFP-CAAX 和 H2A-mCherry RNA 的浓度稀释至 200-300 ng/μL 浓度,总体积为 5 μL(苯酚红色最终浓度为 0.1%)。轻拂混合物并短暂旋转以收集总体积。注射前将稀释的RNA留在冰上。
- 准备注射模具盘,以促进1细胞胚胎的对齐和定向进行微注射。在E3(标准胚胎介质)中融化2%的玫瑰,倒入培养皿中。小心地将注塑模具(参见 材料表)漂浮在热气糖的顶部,以在气精凝固时产生气垫槽的印记。一旦阿加罗斯凝固,取出注塑模具。
- 注射1细胞胚胎
- 一旦鱼开始繁殖,允许15-20分钟,以确保鸡蛋受精。在此期间,使用 P10 和 P10 微型装载机尖端,用 2.5-5 μL 的 RNA 稀释回针头。
- 使用笔切射射体,使用目镜灯片校准针头(微米校准滑梯也足够),以测量液滴的体积(球体体积 = (4/3) * pi * 半径+3)。调整注射时间和压力,使注射体积为1nL(图2I)。
- 用滚筒/转移移液器,小心地将鸡蛋装入注塑模具(图2J)。如果有帮助,使用钳子滚动胚胎,使单个细胞可见,无论是在注射之前还是在注射期间。用户倾向于使用微操纵器。
- 在1细胞阶段注射胚胎,瞄准细胞而不是蛋黄(图2M)。这将确保发育中的胚胎的统一标记。
- 将胚胎提升到所需的阶段(在12马力之前,根据标准分期34)。注射的胚胎将有一个延迟的发展,所以提高他们在一个稍微高一点的温度(29.0-29.5°C),以弥补时间。下午,检查胚胎并取出那些已经死亡,以保持离合器的健康。
3. 安装光学囊泡阶段斑马鱼胚胎进行时垂成像
- 安装前,在E3中准备1.6%的低熔化气。如果计划进行多个成像实验,准备 ±20 mL 的低熔融玫瑰,并在室温下储存。在胚胎安装当天,熔化在安装前的管子中产生新鲜的阿利库特(1-5 mL),该管子可以放置在 42 °C 的热块中。
- 安装前使用荧光显微镜成功注射和荧光整体亮度的屏幕胚胎。理想的样品将具有很强的EGFP和麦片荧光,并处于正确的发育阶段(图3B-B')。
- 使用荧光显微镜筛选胚胎。选择能强烈表达EGFP和麦切里荧光的胚胎进行安装。
- 选择11马力的胚胎。数一数,以适当分阶段胚胎34:在 11 hpf 应该有 3 个烟草, 到 12 hpf 有 6 个闷闷不乐。
注:通过在 12 hpf(11 hpf)之前安装胚胎,当时间延误开始时,样本将以 12 hpf 的适中分级。
- 安装前将胚胎脱钩。手动使用细钳或化学使用假肢(2毫克/毫升)。对于如此年轻的胚胎,在涂有阿加涂层的盘中进行脱毛和胚胎不能接触空气-水接口是至关重要的。
- 使用玻璃滚筒移液器,吸出胚胎并弹出尽可能多的E3,使胚胎位于玻璃巴斯德移液器的尖端。
- 将胚胎从热块中滴入藻子管中。让它沉入阿加罗斯几秒钟,然后先吸一些阿加罗斯,然后吸出胚胎,确保胚胎保持在移液器的尖端(图3C -C')。
- 将玻璃底盘置于解剖范围下进行成像。将胚胎弹出,并放入玻璃底盘中的玫瑰水滴中。使用钳子快速定向,使胚胎向下(玻璃底部的头顶)。让玫瑰水滴硬化(图3D - D')。这一步骤必须迅速而谨慎地完成,因为现阶段的胚胎是脆弱的。受损的胚胎无法在时间拉伤成像过程中存活下来。
注:重要的是要为这个实验始终定向所有胚胎。执行时间拉时时时,胚胎必须适合指定的视野,同时保持额外的空间,使光学囊泡成长。最佳定位是将所有胚胎的前后轴"垂直"对齐,或沿12点和6点钟在钟面上对齐(图3G)。 - 安装在10-12个胚胎之间,这是图像的两倍多,因此,一旦对共聚焦进行评估,就可以选择最好的样本(图3E-E')。 样品将根据年龄、荧光标签的均匀性和安装方向的精度进行评估。
- 安装胚胎后,移液器更阿加罗斯完全覆盖盘底,从而将所有单独的玫瑰液滴包裹在一个大的玫瑰盘中(图3F-F')。 足够的玫瑰将确保单个胚胎液滴或整个盘不会从盘底抬起并浮出视线。
- 一旦硬化,将 agarose 与 E3 覆盖,以便在延时成像实验期间保持样品水分(这取决于建筑物的湿度和环境中的安全问题)。
注:对于这些特定的定时时成像实验来说,三卡因是没有必要的,因为这些胚胎足够年轻:然而,如果需要的话,如果与胚胎的较旧阶段配合,三卡因可以添加到玫瑰本身,并覆盖在媒体上。 - 在纸上,绘制胚胎地图,以便在时间延误期间提供方便参考。这对于以后将该位置与每个样本的基因型关联至关重要。
4. 使用激光扫描共聚焦显微镜进行多个位置共聚焦时间拉伸
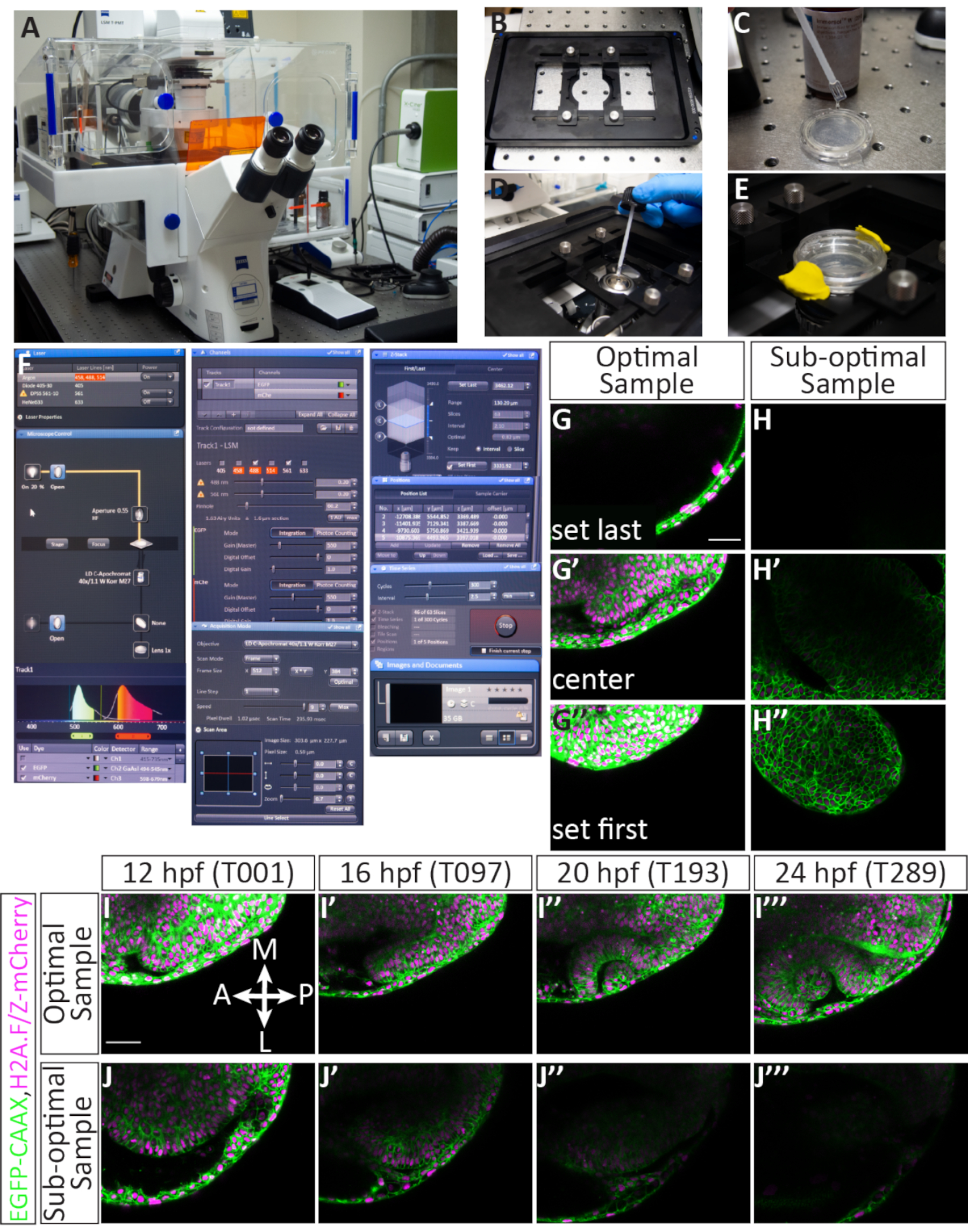
注:此超时成像协议设计用于配备制造商软件的激光扫描共聚焦显微镜(参见 材料表)。该系统配备了一个Piezo Z级设备,允许快速获取Z堆栈。本协议中使用的激光线为 488 nm Argon 离子激光器和 DPSS 561 nm 激光。561 nm 激光非常适合成像 mCherry 荧光:它离 mCherry 激发 (587 nm) 的峰值足够近,而且动力充足。
- 设置焦。
- 在主机上供电并登录台式计算机。
- 安装 Piezo Z 级插入件(图 4B),然后启动采集软件。
- 在 "获取 "模式下的软件中,使用下拉菜单(图 4F)打开488和 561 激光。激光需要几分钟来预热,因此此步骤提前执行以节省时间。
注:获取参数如下:检查Z 堆栈、时间系列和位置框。将帧大小设置为 512 (X) x 384 (Y),扫描速度至 9,扫描模式设置为双向,缩放到 0.7,平孔设置为 60.2(1.63 Airy 单元 == 1.6 μm 部分),Z 间隔设置为 2.1 μm。这将产生 303.6 μm x 227.7 μm 的图像大小,像素大小为 0.59 μm。应检查488和561激光器并分配到同一轨道:EGFP 检测范围为 494-545 nm,mCherry 检测范围为 598-679(图 4F)。 - 随意涂上玻璃盘底部的浸入介质,与水的折射指数相匹配,小心避免气泡(图4C)。选择40倍水的目标,并应用一小滴浸入介质到目标(图4D)。固定在舞台插入的玻璃底盘,应用E3保持胚胎湿润过夜,然后使用建模粘土密封在盘子上的塑料盖(图4E)。提高与菜接触的目的。
- 筛选样品并准备时间拉时。
- 从 "获取 "选项卡切换到 "定位 "选项卡,然后单击灯泡符号以打开传输的光。使用传输的光,用操纵杆定位第一个样品:通过眼睛,首先将样品移入光束中,然后使用目镜到中心,并聚焦在光学囊泡上。
- 返回到 收购 选项卡。确保检查 Z 堆栈、 时间系列和 位置 框,并预热激光以供使用。
- 将 帧大小 设置为 512 (X) x 384 (Y), 将扫描速度 设置为 9,将 扫描模式 设置为双向,并将 缩放 设置为 0.7。
- 在 通道 标题下,首先将 488 和 561 激光分配到 Power 2.0 和 Gain 550(稍后可以调整)。将 针孔 设置为 60.2(1.63 通风单元 +1.6 μm 部分)。
- 开始使用 连续 按钮进行扫描。通过精细的对焦旋钮,将样品定位在 Z 轴中,并将 X-Y 轴放置在框架中。停止扫描。
- 在 "位置 "标题下,单击 "添加" 以保存第一个示例上的 XYZ 信息(图 4F)。这样做只是为了使样品被定时。选择样品,以获得强荧光和最佳安装。
- 切换到 Locate 选项卡,移动到下一个示例,并重复步骤 4.2.5-4.2.6,选择选择示例。
- 完成示例位置,分配 z 范围,然后开始时间拉时。一旦选择所有样本(可在 2.5 分钟的总时间间隔内对 4-5 个样本进行成像),并保存位置,请浏览每个样本并在查看帧内调整 XYZ 位置。
- 突出显示第一个位置,然后单击 "移动到"。在持续扫描时,将光学囊泡排列在帧内。相对于光学囊泡和大脑,在前部和相向区域留出充足的空间。
- 接下来,通过选择 "设置第一 "和" 设置最后 "来分配第一个和最后一个 Z 切片,同时通过具有精细对焦旋钮的 Z 方向。保持 63 的切片总数,因为这将适应光学杯的生长。在光学囊泡的腹腔侧提供额外的空间,以留出生长空间。
- 设置第一个和最后一个 Z 切片后,单击 C 按钮以移动到 Z 堆栈的中心。调整激光功率和两种激光的增益:如果可能,将激光功率保持在 5 以下,并在必要时使用更高的增益。停止扫描。点击" 更新"更新位置信息。
- 单击位置 2 移动到下 一个位置。每个分配的职位重复步骤 4.3.1-4.3.3。激光功率和增益应设置为所有样品都有足够的照明。
- 位置信息和激光设置完成后,分配时间系列设置。在 时间系列 标题下,将 时间间隔 分配为 2.5 分钟, 将周期 分配为 300(图 4F)。计算周期数,当光学杯开发完成时,时间段会超过 24 hpf 阶段。
- 要开始时间拉时,请单击 "开始"。监控第一个完整周期:使用定时器确保一个完整周期(成像所有位置)小于 2.5 分钟,并验证每个位置看起来正确。显微镜独立运行一夜,无需监控。
注:虽然主室温度得到控制,但室温为 20-25 °C。 随着设备的运行和激光扫描,舞台上的温度似乎接近28.5°C,因为胚胎发育以这个预期的速度进行,如视觉上与形态地标的测定。 - 有了这些设置,时间会持续 12.5 小时。一旦时间拉时完成,请保存文件。它将是大(+40 GB),并可以分为个别位置后,它保存使用收购软件。
结果
荧光RNA的注射使亚细胞标记
给细胞的血浆膜和组织素贴上标签的RNA被注射,以捕捉光杯形态形成背后的单个细胞形态和运动。图2演示了在1细胞阶段微射斑马鱼胚胎过程的每一步。简言之,斑马鱼交配(图2E),胚胎被收集并装入注射模具(图2J)。微注射针被反填充(图2H),胚胎直接注射到单细胞(图2K-M),这是获得统一标签所必需的(图2M,图4I-I',电影1)。除了统一标签外,还可以观察到强健的荧光,并不受干扰地进行开发(图 3B-B)。
有效的安装对于 12-24 hpf 的超时成像 OCM 至关重要
对于长时间发生的图像光学杯形态生成,选择理想的胚胎并在嵌入时正确定位每个样本至关重要。图3显示了成功安装11马力夫胚胎的详细情况。注射胚胎首先筛选出足够的荧光(图3B',B',F',F'')。单个胚胎浸入阿加罗斯(图3C,C'),并安装在单滴玫瑰(图3D,D')。所有样品都安装在一个集体的玫瑰盘中(图3E-F)。当胚胎正确、荧光充足、安装充分(图3G)时,样本将停留在显像框架中,以便对整个器官进行成像(图4G-G',I-I',电影1)。如果不能完成其中任何步骤,将导致安装的胚胎不是进行时间拉时成像的最佳样本。旋转程度的微小变化将对胚胎在时间拉时期间的生长方向产生显著影响。图3显示的亚最佳旋转,包括过度旋转的胚胎(图3I),这将导致更后的推移,以及一个胚胎,这是未旋转(图3J),其中只有最前组织可以观察到。除了在安装过程中进行适当的旋转外,安装与框架方向有关的样品更有可能产生成功的减速(图 4G-G',I-I',电影 1)。最佳样品是垂直定位在盘面上的样品,前后轴与时钟面上的12点和6点保持一致(图3G)。次优样品是那些不定向于此轴的样本,如水平或对角线的样本(图3H)。随着时间的推进,这些样品会从成像框架中生长出来,不能用于进一步分析(图4H-H',J-J')。
获取适合未来分析的时限数据集
准确注射、提高和安装的胚胎将产生成功的共聚焦成像样品。图4显示了一个最佳的时展样本:甚至有荧光,样本是12马力。用于成像的 z 堆栈被分配,使第一个切片只是与光学囊泡的正交,而最后一个切片在 12 hpf 光学囊泡的腹腔边界之外留下几片(图 4G-G')。这种对腹腔侧的偏置为心室方向的组织生长提供了多余的空间。这些因素,当满足,导致一个最佳的时限(图4I-我'',电影1)。次优样品具有马赛克或昏暗的荧光,已经发展超过 12 hpf(当光学杯变形正在进行中),或在 Z 堆栈或 XY 框架(图 4H-H')中定位不良) 。当不符合这些标准时,时间拉时将不再捕获组织发展的整个3D体积,不能用于进一步分析(图4J-J')。

图1:斑马鱼光学杯形态形成的4D超时成像工作流。 (A) 荧光标记感兴趣的亚细胞结构,合成适当的封顶mRNA。(B) 在1细胞阶段将mRNA微射入斑马鱼胚胎。(C) 胚胎在光学囊泡阶段、受精后 11 小时或 3 个软骨阶段以圆顶或头朝下安装,用于倒置共聚焦显微镜。(D) 多个胚胎可以在单个时间段的成像过程中按顺序成像。 请单击此处查看此图的较大版本。
{kind=link}

图2:微射1细胞斑马鱼胚胎的视觉指南。(A)注射包(顺时针):中型盒,可用一次性亚硝酸盐手套容纳整个套件:含有微注射针的菜:钳子;矿物油的一块,切成方形的准膜:微载提示;冰上稀释的RNA;阿加罗斯注射模具;标记;滚筒/转移移液器;和 P10 移液器。(B) 微喷射设置:与外部压缩气体源相连的皮科喷油器旁边的解剖显微镜。脚踏板和微操纵器连接到笔画喷油器。显微操纵器配有针架,位于显微镜台上方。(C) 含有六排凹槽的阿加罗斯注塑模具,一侧为 90 度角,另一侧为 45 度角。(D) 笔克喷油器前面板。压力显示屏读取 7.8 PSI:这可以调整与旋钮标记P注射。下面,有按钮手动触发压力注射,改变注射时间,或清除,填充,或持有液体在针。注射时间设置为 08(相当于 8 x 10msec)。输出软管连接到 P输出,并连接到注射针。压力计源开关设置为 P注射,同时设置注射压力,当未进行注射时,可以切换到 P平衡以调整对针头施加的基底压力。P余额监管机构紧邻 P注入监管机构。电源按钮已亮起,以指示机器已打开。(E) 成年斑马鱼在小型养殖池中交配,其中含有一个网状鱼缸,将成年鱼与受精卵分离,便于收集。降低水位以模拟浅水条件,并移除水箱分隔器以启动交配行为。(F) 微管(针)拉拔前面板:包含显示用于拉针的具体条件的显示屏。压力设置为 P = 500,然后是程序编辑的最后日期和时间、程序编号、热度 = 546、PULL = 130、VEL = 70 和时间 = 90。(G) 一个玻璃毛细管插入微管(针)拉拔器并固定到位。每个编程拉运行结果在两个长锥形针(红色箭头)。(H) RNA稀释被反载到针中:酚红色染料使溶液的可视化变得简单。(I) 在打破针尖后,使用解剖范围上的眼镜网度测量RNA的波卢斯。对于此立体显微镜,总放大倍数为 30 倍 6 哈希标记等于 1 nL 体积。(J) 受精卵被装入阿加罗斯注射模具中,并沿模具的槽使用滚筒移液器分发。(K) 含有单细胞阶段胚胎的注射模具位于显微镜下,胚胎按顺序注射。(L) 注射针插入每个胚胎,瞄准单个细胞。(M) 一旦进入细胞,脚踏板用于触发调节的压力量,并将1nL的RNA释放到胚胎细胞中(由苯酚红色可视化)。秤杆:I = 0.1 毫米;L, M = 0.5 毫米。请点击这里查看此图的较大版本。
{kind=link}

图3:安装胚胎进行成像的视觉指南。(A)安装设置(从左到右):一个热块编程为42°C,低熔化的加热管:滚筒移液器;一对钳子;含有脱钩胚胎的阿加罗斯涂层盘;和与金属卤化物荧光灯相连的解剖立体显微镜。(B, B ', B' )注射EGFP-CAAX mRNA和mCherry-H2A mRNA的11马力胚胎,分别在亮场、GFP荧光和麦切里荧光下显示。(C, C' )装有阿加罗斯的玻璃巴氏巴氏管和坐在尖端的胚胎;C'是放大视图。(D, D' )一个胚胎安装在一滴低熔化的玻璃底盘上,从显微镜的阶段和显微镜的目镜中都能看到。(E,E')12个胚胎安装在玻璃底盘上,从显微镜的阶段和显微镜的目镜中都能看到。(F, F ', F ', F '' )所有安装的胚胎都覆盖着一层完整的玫瑰,以填充盘底,从显微镜的阶段和明亮的领域显示的显微镜目镜看到:F''GFP荧光;和F '''mcherry荧光。(G) 12 hpf 的最佳样品以 12 点和 6 点为准,同时在同一平面上安装双光学囊泡,垂直定位。(H) 次优样品:虽然安装的圆角和光学囊泡是彼此在平面上,但此样品以对角线轴为导向,随着开发的进行,将从帧大小中生长出来。(一) 次优样品:此样品是过度旋转的,而不是背安装,因此,光学囊泡前部不会在计时时捕获。(J) 次优样品:此样品旋转不足,不是背架,因此不会在时间段内捕获光学囊泡的后部。点线表示每个光学囊泡。秤杆: B = 0.1 毫米;D' = 1.5 毫米;E', F' = 2.5 毫米;G = 0.1 毫米。请单击此处查看此图的较大版本。
{kind=link}

图4: 设置多个位置时间拉时和潜在结果。 (一个) 激光扫描聚焦显微镜。(B) 皮耶佐 Z 阶段插入。(C) 含有嵌入在阿加罗斯中的胚胎的玻璃底盘底部涂有浸入介质,与水的折射指数相匹配。(D) 浸入介质滴放在 40x W 目标(长工作距离)上。(E) 菜是固定到位与舞台插入, E3 被添加在玫瑰层的顶部, 和盖子用建模粘土固定.(F) 采集软件设置:所需的激光线从下拉菜单打开。对于注射EGFP-CAAXRNA和H2A-mCherryRNA的胚胎,打开Argon和DPSS 561-10激光。红色亮点表示阿贡激光正在升温。为了在目标上定位样品,通过显微镜控制面板打开传输的光。EGFP 和 mCherry 都分配到轨道 1(用于同步成像),并设置每个探测器的范围。在这里,EGFP 范围设置为 494-545 nm,mCherry 范围设置为 598-679 nm。下 渠道 菜单,激光电源可以设置。激光加热后,功率可增加到 5.0,增益(主)可调整。针孔设置为 60.2,等于 1.63 通风单元或 1.6 μm 部分。下 收购 菜单,帧大小设置为 512 x 384,扫描速度为 9.0,平均为双向,变焦设置为 0.7。下 Z 堆栈 菜单,Z 轴中的第一个和最后一个位置设置,而共焦正在扫描。间隔(步数大小)设置为 2.1 μm。在选择第一个和最后一个 Z 位置时,通常保持 63 片的标准数:这适应了眼睛的整体生长。选定了"使用皮耶佐"选项。下 位置 菜单中列出每个选定的位置,包括分配的号码和 X、Y 和 Z 位置。还有其他按钮来控制图像中的胚胎数量(单独的位置),包括添加、更新或删除。下 时间系列 菜单,周期设置为 300(将获取的 Z 堆栈数量),间隔(时间步骤)设置为 2.5 分钟。设置完成后,通过按下启动启动时间安排。该软件将显示 Z 堆栈的切片、周期和当前扫描的位置。当延时完成时,通过单击软盘图标来保存文件。 图片和文档 面板。(G-J) 细胞膜 (green) 标记有 EGFP-CAAX 和细胞核 (色度素, magenta) 标有 H2a - mcherryRNA。(G,G',G'') 为最佳安装的样品设置第一个和最后一个 Z 切片的示例。第一个 Z 切片(在正点侧)以牺牲正点异皮为代价,而最后一个 Z 切片(在腹腔侧)远远低于光学囊泡,以适应其在腹腔方向的生长。(H,H',H'') 设置次优示例的第一个和最后一个 Z 切片示例。样品有微弱的mCherry荧光,它是对角线安装的,因此开发中的光学杯不太可能在时间延长期间留在框架中。(我,我,我,我') 最佳样品的时限示例。Z 堆栈中间的单片图像从整个 63 切片卷中取自 4 个时间点(T 值)。布尔,大脑;神经视网膜;RPE,视网膜色素上皮;勒,镜头(J, J ', J ', J '' ') 从次优示例中抽取的减时示例。在这种情况下,样品移出Z平面,只捕获了前光学杯的斜部分。秤杆: G, I = 50 μm。 请单击此处查看此图的较大版本。
{kind=link}
电影1:最佳野生类型胚胎中光学杯形态发生的时间拉时。 带有EGFP-CAAX(等离子膜、 绿色)和H2A标签的野生胚胎的面食视图。F/Z-米切里(色红素, 品红色)。时间段为 12-24 hpf,包含整个 4D 数据集中的单个共焦部分。时间间隔为 z 堆栈之间的 2.5 分钟,以每秒 22.5 帧的速度显示。请点击这里下载这部电影。
讨论
在这里,我们描述了一个协议 ,在托托 标签和4D定时成像的光学杯形态。我们逐步通过生成覆盖RNA编码荧光蛋白的过程,以标记不同的亚细胞隔间;注射斑马鱼1细胞胚胎;将11个 hpf 胚胎嵌入阿加罗斯进行多路复用成像;并在光学杯形态发生期间获取多个胚胎的 4D 数据集 (12/24 hpf)。
这些信息密集型数据集可以回答无数问题。4D 数据可以通过多种方式进行可视化和定量分析。虽然超出了本协议的范围,但我们在这里包括了我们的一些目标和标准应用程序,作为可以完成的事物类型的示例。当然,定量图像分析方法正在不断开发中,并且可以使用商用和定制工具。如果以前没有使用过此类方法,则应准备通过一些优化来确保已获得的数据集足以用于选择定量分析方法。
由于文件的大小,可视化和定量评估 4D 数据集可能具有挑战性。采集软件可用于将数据集分离到单个胚胎中,ImageJ/Fiji 可用于将共聚焦文件从商业格式转换为更标准的 tif 堆栈,其中每个时间点都保存为单独的文件。这将减少文件大小并标准化文件格式。每个时间点的单个光学部分可以使用 ImageJ/Fiji 将 2D (XY) 定时器组装为 2D,从而实现数据的快速 2D 可视化和评估。电影1就是一个例子:一个光学部分随着时间的推移组装为图4I'I'中显示的最佳样本的时限。从那里,对于3D和4D可视化,我们通常使用FloRender35,36,这是免费的,但有特定的显卡要求。使用 FluoRender,您可以在任何轴中旋转 3D 渲染数据,随着时间推移在 4D 中可视化数据集,在任何平面上生成切口,并将旋转和可视化保存为电影或系列 tif 图像。
在定量分析方面,有许多问题需要回答。我们开发了内部软件,以帮助我们自己的特定目标,了解细胞行为背后的光杯变形。我们的程序,长跟踪器,使用核信号作为位置的代理来跟踪细胞13。通过这些数据,我们可以确定细胞移动的时间、地点和方式:细胞移动的速度和移动程度;以及细胞分裂的频率。除了我们自己的软件,还有多个商业可用和定制的选项,用于 4D 单元格跟踪。我们还开发了程序LongAxis进行细胞分割和量化细胞形状和组织内神经视网膜37。但是,输入到 LongAxis 的数据集是单个 Z 堆栈,以高分辨率拍摄。一个持续的挑战是生成具有足够高分辨率的超时数据集,以便对细胞进行自信分割,并推断其形态。另一种选择是马赛克(稀疏)标签使用光转换荧光,如Kaede直接可视化细胞形状,因为我们和其他人已经进行了在发展的眼睛8,11,13。这简化了细胞分割问题,并且可以通过 FluoRender 中的 3D 渲染轻松量化细胞形状和大小。
此协议的每一步都专门针对我们的目的进行了优化。此协议的特殊性导致几个限制:协议,如书面,没有优化成像斑马鱼眼睛发育的其他方面(如视网膜神经生成),其他眼睛结构,其他发育阶段,或其他亚细胞隔间。嵌入的方向、成像速度和标签都旨在让我们回答我们的生物学问题。例如,为了成像视网膜神经生成,此处描述的胚胎的做家定向可能不允许可视化感兴趣的特定特征,而且成像速度可能不合适。协议的许多方面可以根据个人的具体兴趣和目标,根据各种需求进行调整和修改。首先,使用RNA注射使标签过程非常灵活。荧光蛋白融合结构可用于标记感兴趣的亚细胞结构,RNA注入量可多种多样,以优化标签。根据我们与光转换荧光素Kaede的工作,RNA注射似乎支持一阵翻译,是超过12马力11,13。RNA注射产生的高浓度荧光蛋白表达可以对抗光出血,但这种方法不支持持续表达感兴趣的荧光标签。如果需要持续表达,例如在后期成像胚胎时,转基因线是一种选择,在斑马鱼中构建新线条是直截了当的。
其次,协议可以适应后期的发展阶段。由于色素可能阻碍后期成像,胚胎可以用苯甲酸酯(PTU)来抑制色素的形成,或者可以使用基因突变体进行色素合成。为了防止胚胎抽搐,三卡因可以添加到阿加罗斯和胚胎介质覆盖溶液中,并且可以根据需要调整百分比的糖。随着眼睛的生长,可能需要改变安装方向:在这里,我们安装胚胎多管,但在后期阶段,它可能更有意义,横向或前部安装,这取决于兴趣的结构。由于不同的过程发生在不同的空间和时间尺度上,因此还可以优化图像采集的 Z 步和时间步骤。这些功能实际上只能根据每个实验室的需求进行经验确定。
最后,这个协议是专门为激光扫描共聚焦显微镜开发的,胚胎嵌入在眼睛发育早期相对较高的高比例的玫瑰中。如果在眼睛发育过程中在不同时间或不同位置进行成像,则此协议需要适应感兴趣的过程。目前有许多不同的成像方法是可能的,更多的是由光学工程师开发的。每种方法都带来了自己的一系列挑战,从嵌入和安装胚胎进行成像的不同方式,到不同的文件大小和格式。我们在介绍中概述了指导优化过程的考虑因素,其中最大空间和时间分辨率与光模糊、光毒性和巨大的文件大小相平衡。我们希望,这些一般原则,除了上述实际信息外,将有助于其他国家建立时间拉长成像方法,以研究眼睛发育中的许多开放性问题。
披露声明
作者没有什么可透露的。
致谢
我们感谢关实验室的过去和现在的成员就本议定书的时效方法和讨论开展工作。这项工作得到了国家卫生研究院(R01 EY025378 和 R01 安永025780至KMK的支持;F31 EY030758 至 SL;和 T32 GM007464 到 MAC)。
材料
| Name | Company | Catalog Number | Comments |
| Delta TPG Dish 0.17mm clear | Bioptechs | 0420041500C | coverglass bottom for optical compatibility |
| Disposable Pasteur Pipettes | VWR | 14672-608 | overall length 14.6 cm (53/4") |
| Dumont #5, #55, or #5SF Forceps | Fine Science Tools | 11254-20 | For style #5: straight tip, 0.05 x 0.01 mm tip, inox, 11 cm long |
| Flaming/Brown Micropipette Puller | Sutter | P-97 | |
| Immersol W | Carl Zeiss Microscopy | 4449690000000 | immersion fluid for water-emission objectives, halogen-free, low fluorescence |
| Microinjection Mold | Adaptive Science Tools | TU-1 | Six ramps, one 90 degree and one 45 degree beveled side. |
| Microloader Tips | Eppendorf | 930001007 | Microloader, tip for filling glass microcapillaries, 0.5 – 20 µL |
| mMessage mMachine SP6 Transcription Kit | Invitrogen | AM1340 | contains SP6 Enzyme Mix, 10X Reaction Buffer, 2XNTP⁄CAP Solution, GTP Solution, pTRI-Xef, TURBO DNase, Ammonium Acetate Stop Solution, Lithium Chloride Precipitation Solution, and Gel Loading Buffer and are all stored at –20°C. Nuclease-free Water may be stored at any temperature. |
| Nikon SMZ645 Stereo Microscope | Nikon | SMZ645 | used for injecting embryos (see Fig. 2B) |
| NotI-HF enzyme | NEB | R3189S | comes with Gel Loading Dye, Purple (6X) |
| NuSieve GTG Agarose | Lonza | 50081 | fine resolution, low melt agarose |
| Objective LD "C-Apochromat" 40x/1.1 W Korr M27 | Carl Zeiss Microscopy | 421867-9970-000 | |
| Olympus SZX16 Stereo Microscope | Olympus | SZX2-ZB16 | used for screening, embedding embryos (see Fig. 3A) |
| Phenol red solution | Sigma-Aldrich | P0290 | 0.5% in DPBS, sterile filtered, endotoxin tested, cell culture tested |
| Pico-liter Microinjector | Harvard Apparatus | PLI-100 | Femtoliter to microliter injections; digital readouts for injection count, time, and pressure; contains 5 pressures including inject, balance, clear, fill, and hold |
| Pipette Pump Pipettor | SP Bel-Art | F37898 | fits a disposable pasteur pipette, contains thumbwheel on side for aspirating or dispensing and plunger for quick emptying |
| Premium Thin Wall Borosilicate Glass Capillaries | Warner Instruments | G100T-4 | 1.0 x 0.78 mm |
| QIAquick PCR Purification Kit | Qiagen | 28106 | contains QIAquick Spin Columns, Buffers, Collection Tubes (2 ml) |
| RNase-free H2O | Invitrogen | AM9906 | DNase-Free, Molecular Biology Grade, RNase-Free |
| RNeasy Mini Kit | Qiagen | 74104 | contains RNeasy Mini Spin Columns, Collection Tubes (1.5 ml and 2 ml), RNase-free Reagents and Buffers |
| Stage Attachment Z PIEZO WSB 500 | Carl Zeiss Microscopy | 432339-9000-000 | |
| Stage Insert Z PIEZO WSB Universal | Carl Zeiss Microscopy | 432339-9030-000 | |
| Zeiss LSM 880 with Airyscan | Carl Zeiss Microscopy | N/A | inverted Axio Observer microscope |
| Zen Black 2.3 SP1 software | Carl Zeiss Microscopy | N/A | Zeiss Efficient Navigation (ZEN) controls all light microscope systems from Zeiss |
参考文献
- Adler, R., Canto-Soler, M. V. Molecular mechanisms of optic vesicle development: complexities, ambiguities and controversies. Developmental Biology. 305 (1), 1-13 (2007).
- Chow, R. L., Lang, R. A. Early eye development in vertebrates. Annual Review of Cell and Developmental Biology. 17, 255-296 (2001).
- Fuhrmann, S. Eye morphogenesis and patterning of the optic vesicle. Current Topics in Developmental Biology. 93, 61-84 (2010).
- Martinez-Morales, J. R., Wittbrodt, J. Shaping the vertebrate eye. Current Opinion in Genetics and Development. 19 (5), 511-517 (2009).
- Yang, X. J. Roles of cell-extrinsic growth factors in vertebrate eye pattern formation and retinogenesis. Seminars in Cell & Developmental Biology. 15 (1), 91-103 (2004).
- Bogdanovic, O., et al. Numb/Numbl-Opo antagonism controls retinal epithelium morphogenesis by regulating integrin endocytosis. Developmental Cell. 23 (4), 782-795 (2012).
- Bryan, C. D., Casey, M. A., Pfeiffer, R. L., Jones, B. W., Kwan, K. M. Optic cup morphogenesis requires neural crest-mediated basement membrane assembly. Development. 147 (4), (2020).
- Bryan, C. D., Chien, C. B., Kwan, K. M. Loss of laminin alpha 1 results in multiple structural defects and divergent effects on adhesion during vertebrate optic cup morphogenesis. Developmental Biology. 416 (2), 324-337 (2016).
- Cavodeassi, F., Ivanovitch, K., Wilson, S. W. Eph/Ephrin signalling maintains eye field segregation from adjacent neural plate territories during forebrain morphogenesis. Development. 140 (20), 4193-4202 (2013).
- England, S. J., Blanchard, G. B., Mahadevan, L., Adams, R. J. A dynamic fate map of the forebrain shows how vertebrate eyes form and explains two causes of cyclopia. Development. 133 (23), 4613-4617 (2006).
- Gordon, H. B., et al. Hedgehog signaling regulates cell motility and optic fissure and stalk formation during vertebrate eye morphogenesis. Development. 145 (22), (2018).
- Ivanovitch, K., Cavodeassi, F., Wilson, S. W. Precocious acquisition of neuroepithelial character in the eye field underlies the onset of eye morphogenesis. Developmental Cell. 27 (3), 293-305 (2013).
- Kwan, K. M., et al. A complex choreography of cell movements shapes the vertebrate eye. Development. 139 (2), 359-372 (2012).
- Martinez-Morales, J. R., et al. Ojoplano-mediated basal constriction is essential for optic cup morphogenesis. Development. 136 (13), 2165-2175 (2009).
- Miesfeld, J. B., et al. Yap and Taz regulate retinal pigment epithelial cell fate. Development. 142 (17), 3021-3032 (2015).
- Nicolas-Perez, M., et al. Analysis of cellular behavior and cytoskeletal dynamics reveal a constriction mechanism driving optic cup morphogenesis. eLife. 5, (2016).
- Picker, A., et al. Dynamic coupling of pattern formation and morphogenesis in the developing vertebrate retina. PLoS Biology. 7 (10), e1000214 (2009).
- Rembold, M., Loosli, F., Adams, R. J., Wittbrodt, J. Individual cell migration serves as the driving force for optic vesicle evagination. Science. 313 (5790), 1130-1134 (2006).
- Sidhaye, J., Norden, C. Concerted action of neuroepithelial basal shrinkage and active epithelial migration ensures efficient optic cup morphogenesis. eLife. 6, (2017).
- Driever, W., et al. A genetic screen for mutations affecting embryogenesis in zebrafish. Development. 123, 37-46 (1996).
- Haffter, P., et al. The identification of genes with unique and essential functions in the development of the zebrafish, Danio rerio. Development. 123, 1-36 (1996).
- Nusslein-Volhard, C. The zebrafish issue of Development. Development. 139 (22), 4099-4103 (2012).
- Hoshijima, K., et al. Highly efficient CRISPR-Cas9-based methods for generating deletion mutations and F0 embryos that lack gene function in zebrafish. Developmental Cell. 51 (5), 645-657 (2019).
- Kwan, K. M., et al. The Tol2kit: a multisite gateway-based construction kit for Tol2 transposon transgenesis constructs. Developmental Dynamics. 236 (11), 3088-3099 (2007).
- Almeida, A. D., et al. Spectrum of Fates: a new approach to the study of the developing zebrafish retina. Development. 141 (9), 1971-1980 (2014).
- Baye, L. M., Link, B. A. Interkinetic nuclear migration and the selection of neurogenic cell divisions during vertebrate retinogenesis. The Journal of Neuroscience. 27 (38), 10143-10152 (2007).
- Clark, B. S., et al. Loss of Llgl1 in retinal neuroepithelia reveals links between apical domain size, Notch activity and neurogenesis. Development. 139 (9), 1599-1610 (2012).
- Das, T., Payer, B., Cayouette, M., Harris, W. A. In vivo time-lapse imaging of cell divisions during neurogenesis in the developing zebrafish retina. Neuron. 37 (4), 597-609 (2003).
- Kay, J. N., et al. Transient requirement for ganglion cells during assembly of retinal synaptic layers. Development. 131 (6), 1331-1342 (2004).
- Norden, C., Young, S., Link, B. A., Harris, W. A. Actomyosin is the main driver of interkinetic nuclear migration in the retina. Cell. 138 (6), 1195-1208 (2009).
- Suzuki, S. C., et al. Cone photoreceptor types in zebrafish are generated by symmetric terminal divisions of dedicated precursors. Proceedings of the National Academy of Sciences of the United States of America. 110 (37), 15109-15114 (2013).
- Wan, Y., et al. The ciliary marginal zone of the zebrafish retina: clonal and time-lapse analysis of a continuously growing tissue. Development. 143 (7), 1099-1107 (2016).
- Yoshimatsu, T., et al. Presynaptic partner selection during retinal circuit reassembly varies with timing of neuronal regeneration in vivo. Nature Communications. 7, 10590 (2016).
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., Schilling, T. F. Stages of embryonic development of the zebrafish. Developmental Dynamics. 203 (3), 253-310 (1995).
- Wan, Y., Otsuna, H., Chien, C. B., Hansen, C. An interactive visualization tool for multi-channel confocal microscopy data in neurobiology research. IEEE Transactions on Visualization and Computer Graphics. 15 (6), 1489-1496 (2009).
- Wan, Y., Otsuna, H., Chien, C. B., Hansen, C. FluoRender: an application of 2D image space methods for 3D and 4D confocal microscopy data visualization in neurobiology research. IEEE Pacific Visualization Symposium. , 201-208 (2012).
- Carney, K. R., Bryan, C. D., Gordon, H. B., Kwan, K. M. LongAxis: a MATLAB-based program for 3D quantitative analysis of epithelial cell shape and orientation. Developmental Biology. 458 (1), 1-11 (2020).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。