
Method Article
Zebra Balığı Optik Kupası Morfogenezinin 4 Boyutlu Görüntülenmesi
Bu Makalede
Özet
Bu protokol, zebra balığının erken göz gelişiminin toto etiketlemesi ve çok boyutlu görüntülenmesi için bir yaklaşımı açıklar. Lazer tarama konfokal mikroskopisi kullanarak etiketleme, gömme ve dört boyutlu (4D) görüntülemeyi ve optik bardak morfogenezinin diseksiyon mekanizmaları için veri kümelerinin alımını optimize etmek için dikkat edilmesi gereken noktaları açıklıyoruz.
Özet
Görsel sistem fonksiyonu hassas doku ve organ yapılarının kurulmasını gerektirir. Omurgalı gözde yapısal defektler görme bozukluğunun yaygın bir nedenidir, ancak göz morfogenez mekanizmaları hala yezifez anlaşılmaktadır. Embriyonik gözün temel organizasyonu omurgalılar boyunca korunur, böylece zebra balığı embriyolarının canlı görüntülenmesi, normal ve patolojik koşullar altında göz gelişimini gerçek zamanlı olarak doğrudan gözlemlemek için güçlü bir yaklaşım haline gelmiştir. Embriyoda hareketler, morfolojiler, etkileşimler, bölünme ve ölüm gibi dinamik hücre süreçleri görselleştirilebilir. Zebra balıklarında erken göz gelişiminin hücre altı yapılarının ve timelapse konfokal mikroskopisinin tek tip etiketlenerek etiketlendirilme yöntemleri geliştirdik. Bu protokol, 1 hücreli zebra balığı embriyosuna enjeksiyon için kapaklı mRNA oluşturma, embriyoları optik vezikül aşamasında monte etme (döllenmeden sonra ~ 12 saat, hpf) ve optik bardak morfogenezinin lazer tarama konfokal mikroskopta çok boyutlu timelapse görüntülemesini gerçekleştirme yöntemini özetlemektedir, böylece aynı görüntüleme oturumunda ardışık olarak birden fazla veri kümesi elde edilir. Böyle bir yaklaşım, hücre izleme, hacim ölçümleri, üç boyutlu (3D) işleme ve görselleştirme dahil olmak üzere çeşitli amaçlar için kullanılabilecek veriler sağlar. Yaklaşımlarımız, hem vahşi tip hem de genetik mutant koşullarında optik bardak gelişimini yönlendiren hücresel ve moleküler mekanizmaları tam olarak belirlememizi sağlar. Bu yöntemler doğrudan diğer gruplar tarafından kullanılabilir veya zebra balığı göz gelişiminin birçok ek yönünü görselleştirmek için uyarlanabilir.
Giriş
Omurgalı göz gelişimi, potansiyel beyin nöroepithelium'dan optik veziklitlerin ortaya çıkması veya buharlaşması ile başlar. Optik veziklinler daha sonra bir dizi doku şekli değişikliğine uğrar, uzaır ve daha sonra optik kabı oluşturmak için invaginasyona uğrar. Optik kapta, her ikisi de nöroepitheliumdan türetilen nöral retina ve retina pigment epitel, yüzey ektoderminden elde edilen nascent lensi sarar. Tüm süreç, nöroepithelium, ektoderm ve mezenkimal hücre popülasyonları arasında koordine edilen karmaşık bir dizi hücre ve doku hareketi ve moleküler sinyalizasyon gerektirir. Bu ilk olaylar gözün temel yapısını oluşturur ve iris ve kornea oluşumu da dahil olmak üzere daha sonraki göz gelişimi adımları erken organizasyon üzerine detaylandırmalardır. Erken göz gelişimi ve morfogenezdeki aksaklıklar, anoftalmi, mikroftalmi ve kolobom da dahil olmak üzere insanlarda çok sayıda görme bozukluğu koşullarının altında yatandır. Optik bardak morfogenezini yöneten hücresel ve moleküler mekanizmaların kilidini açmak, görsel sistem gelişimini ve bu süreçler ters gittiğinde ortaya çıkan patolojik koşulları daha iyi anlamak için çok önemlidir.
Omurgalı göz gelişimi ve morfogenez anlayışımız, klasik histolojik çalışmalardan embriyolojik ve genetik yaklaşımlara kadar uzanan muazzam miktarda çalışmadan, fare, civciv, kurbağa ve balık1, 2,3,4,5dahil olmak üzere çeşitli model organizmalarda ortayaçıkmıştır. Bu çalışma gövdesi erken göz gelişimini düzenleyen moleküler mekanizmalar oluştururken, tarihsel olarak gözün morfogenezinin zayıf bir şekilde anlaşılması vardır: 3D yapısının ortaya çıkması. Bu bulguların büyük kısmı, ayrık zaman noktalarındaki bölümlenmiş embriyoların görüntülenmesinden gelmiştir. Bu, doku morfolojisinin 2 boyutlu bir görünümünü sağlamak için yeterli olmakla birlikte, morfogenez dinamik, 3D bir süreçtir. Dokunun şeklinin zaman içinde 3 boyutta nasıl değiştiğini, tek hücrelerin nasıl davrandığını ve bu davranışların 3D doku şeklindeki değişikliklere nasıl katkıda bulunduğunu belirlemek için farklı yaklaşımlar gereklidir.
Bilgideki bu önemli boşluğu gidermek için bir çözüm, organ şekillendikçe hücrelerin ve dokuların gerçek zamanlı olarak dinamik olarak gözlemlenmesini sağlayan canlı görüntülemedir. Ne yazık ki, embriyonik gelişimin kısıtlamaları nedeniyle birçok model organizmada bu kolayca mümkün değildir. Örneğin, fare ve civciv embriyolarına (rahimde veya bir yumurta kabuğu içinde gelişen) kolayca erişilmez ve birçok canlı embriyo optik olarak şeffaf değildir, bu da önemli ışık saçılımına neden olur ve görüntülerin elde edilebileceği derinliği sınırlar. Zebra balığı, dış gelişim ve şeffaf embriyolar ile göz morfogenezinin canlı görüntülenmesini gerçekleştirmek için benzersiz bir fırsat sağlar 6,7,8,9,10,11,12, 13,14,15,16,17,18,19. Geniş transgenik ve mutant hatların yanı sıra yeni transgenikler ve mutantlar üretmek için araçlar da mevcuttur20,21,22,23,24. Ayrıca, optik bardak morfogenez zebra balıklarında 12 saatlik bir zaman diliminde (döllenmeden 12-24 saat sonra, hpf) hızla ortaya çıkar ve tüm sürecin görüntülenmesini mümkün hale getirir.
Canlı görüntüleme çalışmaları, hücre altı yapıların genetik olarak kodlanmış hayati etiketlemesine izin veren floresan protein ailesinin genişletilmesi ve optimizasyonu ile hızlandırılmış, ayrıca mikroskopi yöntemlerinde iyileştirmeler ve yenilikler gerçekleştirilmiştir. Burada açıklanan protokolde, dönen disk konfokal mikroskopisi, seçici düzlem aydınlatma mikroskopisi (SPIM ve varyantları) ve diğer daha özel mikroskopi yöntemleri de dahil olmak üzere zebra balığı embriyogenezinin görüntülenmesine yönelik diğer güncel yaklaşımlar yerine lazer tarama konfokal mikroskopisi sunusu kullanmaktadır. Gelişen zebra balığı gözü için, dönen disk konfokal mikroskopinin dokuda daha derin görüntüleme için yeterli olmadığını gördük. SPIM son derece hızlı bir görüntüleme süresine sahip olmasına ve daha yaygın olarak kullanılmaya başlamasına rağmen, görselleştirme ve analiz için büyük veri kümelerini işlemek zor olmaya devam etmektedir. Buna karşılık, lazer tarama konfokal mikroskopiye, özellikle optik donanımın montajında uzmanlık eksikliği olan bireyler için kolayca erişilebilir. Lazer tarama konfokal mikroskopisinin geniş mevcudiyetinin protokolümüzü birçok laboratuvar için yararlı hale getireceğini umuyoruz.
Burada, optik bardak morfogenezinin 4D veri kümelerini yakalama, embriyonun membranlar ve kromatin için toto etiketlemesini ve görüntü alımı için lazer tarama konfokal mikroskobu (Şekil 1'deşematize edilmiş) yakalama yöntemimizi açıklıyoruz. Burada kullanılan floresan proteinler (EGFP-CAAX ve H2A. F/Z-mCherry) tek hücre çözünürlüğü ile doku etiketlemesi sağlamak için seçildi. Çeşitli görüntü analizi ve görselleştirme işlevleri için bu protokolle oluşturulan veri kümelerini kullanıyoruz. Diğer hücre altı yapılar isteniyorsa bu protokol kolayca uyarlanabilir. Hücre şeklinin görselleştirilmesi için plazma membran etiketlemesi seçildi: Bir prenilasyon sinyal dizisi olarak hizmet veren H-ras'ın son 21 amino asidinin EGFP13'ünC-terminusuna kaynaştığı EGFP-CAAX kullanıyoruz. Diğer plazma membran hedefli floresan proteinlerin (örneğin transmembran füzyonu veya miristosilat) da işe yaraması muhtemeldir. Çekirdekleri işaretlemek için H2A'yı seçtik. F/Z-mCherry, mCherry bir histone proteinine kaynaştırılır13. Bu, mitotik mil yönlendirmesi de dahil olmak üzere hücre bölünmesinin kolayca görselleştirilmesini sağlar.
Herhangi bir canlı görüntüleme yaklaşımıyla, sinyal-gürültü oranını, eksenel çözünürlüğü ve numune korumasını en üst düzeye çıkarırken görüntüleme hızını artırmak arasındaki takasları göz önünde bulundurmak gerekir. Tek bir koşuda görüntülenebilen embriyoların sayısını ve görüntü kalitesini en üst düzeye çıkarmak için yöntemlerimizi optimize ettik. Genellikle, amaç, optik bardak morfogizinin başlangıcında vahşi tipten fenotipik olarak ayırt edilemeyen homozigot mutant embriyosunda optik kupa morfogenezini ve heterozipöz bir incrossun yavrularını (embriyoların% 25'i istenen genotiptir) imgelemektir. Görüntü alımını optimize ederek ve daha sonra çoklayıcı olarak, homozijik mutant embriyonun veri kümesini yakalama olasılığı artar.
Zamansal çözünürlük veya hacim verilerinin (Z yığınları) ne sıklıklarda elde edildiği, timelapse görüntülemenin önemli bir yönüdür. Bu tür veri kümelerinin amacına bağlı olarak, hız için farklı gereksinimler vardır. Başlangıçta bu protokol manuel 4D hücre takibi için geliştirilmiştir. Tek tek hücrelerin düzgün etiketlenmiş bir doku içinde izlenmesi, herhangi bir hücrenin zaman içinde sürekli olarak izlendiğinden emin olmak için yeterince yüksek zamansal çözünürlük gerektirir. Zebra balığı optik bardak morfogenezinin Z-yığınlarının 12 saat boyunca en az 3,1 dakikada bir alınması gerektiğini bulduk; Burada, her 2,5 dakikada bir 4-5 embriyonun Z yığınlarını elde edebileceğimiz bir lazer tarama konfokal mikroskop üzerinde kazanımımızı optimize ettik.
Z adım boyutunu belirlemek protokol optimizasyonunda çok önemli bir adımdı: 3D işleme ve görselleştirme için izotropik veriler idealdir ve bu da Z adım boyutunun XY piksel boyutuna eşit olduğu idealdir. Gerçekte, görüntüleme süresi ve fotobleaching ile kısıtlamalar göz önüne alındığında, canlı örneklerle bu tür timelapse verilerini elde etmek son derece zordur. Bu nedenle, yeterli Z-adım boyutunun belirlenmesi, deneyin oluşturulması ve görselleştirilmesi ihtiyaçları ve özellikle, hızı korurken ve fotobleachingi önlerken eksenel bilgileri en üst düzeye çıkarmak için hangi X:Y:Z voksel oranının gerekli olduğu için önemlidir. Bu protokol için, belirlenen voksel oranı 1:1:3.5 idi (0.6 μm x 0.6 μm x 2.1 μm Z-step boyutu 40x uzun çalışma mesafesi su daldırma hedefi kullanılarak). 130-140 μm'lik bir Z derinliği elde ederken, bu uygun zamansal çözünürlük ve az fotobleaching ile hacim verileri verir.
Yukarıda tartışıldığı gibi, bu protokol zebra balığı optik bardak morfogenezinin 4D görüntülemesi, plazma membran ve kromatin için etiketlenmiş totodaki embriyolar ve lazer tarama konfokal mikroskobu için özeldir. Aşağıdaki protokol çeşitli deneyler ve ihtiyaçlar için kolayca uyarlanabilir. İlk olarak, hücre altı yapılarla ilgili olarak, canlı bir hücre işaretleyicisi bulunan herhangi bir yapı görüntülenebilir. Daha sonra, buradaki odak sadece optik bardak morfogenezine odaklansa da, timelapse görüntüleme göz gelişiminin diğer aşamaları için uyarlanabilir, örneğin, nörogenez25 , 26,27,28,29,30,31,32,33. Daha sonra gelişmeyi görüntülemek için embriyo immobilizasyonunu (spontan kas aktivitesi 24 hpf civarında başladığında), pigmentasyonu (24 hpf civarında ortaya çıkmaya başlar), doku boyutunu (göz nörogenez sırasında hacim olarak önemli ölçüde büyür) ve görüntüleme hızını (ilgi sürecinin hızına bağlı olarak ayarlanmalıdır) göz önünde bulundurmak gerekebilir. Tüm bu hususlar kolayca yönetilebilir. Protokol oldukça esnektir; buradaki özel protokolün ayrıntılarına ek olarak, göz gelişiminin diğer yönlerini canlı görüntülemeye ilgi duyanlara yardımcı olacak genel ilkeler vardır.
Protokol
Zebra balığı kullanılarak burada açıklanan tüm yöntemler Dr. Kristen Kwan'ın "Zebra Balıklarında Görsel Sistem Gelişiminin Hücresel ve Moleküler Mekanizmaları" hayvan protokolü kapsamındadır ve Utah Üniversitesi Kurumsal Hayvan Bakım ve Kullanım Komitesi (IACUC) tarafından onaylanmıştır.
1. Kapaklı RNA Sentezi
- Tüp bebek transkripsiyon için DNA şablonunu oluşturun.
- Reaksiyon karışımının 100 μL hacminde 10 μg DNA sindirerek DNA şablonunun doğrusallaştırının. Tipik bir reaksiyon aşağıdaki gibi monte edilir: 3 μL enzim ve 10 μL tampon kullanarak 10 μg DNA'yı sindirin, reaksiyon hacmini su ile 100 μL'ye getirin.
NOT: Özet için tipik bir şablon, burada açıklanan membran ve kromatin etiketlemesi için pCS2-EGFP-CAAX veya pCS2FA-H2A.F/Z-mCherry gibi bir pCS2 vektörüdür. Bu durumda, plazmid DNA'ların her biri NotI enzimi kullanılarak sindirilir. Kullanıcılar yıldız etkinliğine karşı dikkatli olmalıdır; Bu durumda, yüksek doğrulukta bir enzim önerilir ve ticari olarak kullanılabilir. - DNA'nın tamamlanması için sindirilmesini sağlamak için reaksiyonun bir gecede 37 °C'de kuluçkaya yatırılması.
- Üreticinin protokolünü izleyerek bir PCR temizleme kiti kullanarak kısıtlama özetini temizleyin. DNA'yı 30 μL ddH2O ile kurulayın Doğrusallaştırılmış DNA'yı -20 °C'de saklayın ve gerektiğinde kullanın.
NOT: Sindirilen DNA miktarı ve elüsyon hacmi yaklaşık 0,3 μg/μL verim; bu, şablon olarak ~2 μg DNA kullanılarak her turda ~ 5 tur in vitro transkripsiyon için yeterlidir.
- Reaksiyon karışımının 100 μL hacminde 10 μg DNA sindirerek DNA şablonunun doğrusallaştırının. Tipik bir reaksiyon aşağıdaki gibi monte edilir: 3 μL enzim ve 10 μL tampon kullanarak 10 μg DNA'yı sindirin, reaksiyon hacmini su ile 100 μL'ye getirin.
- In vitro transkripsiyon kiti kullanarak in vitro transkript kapaklı RNA. PCS2 şablonları için (burada açıklandığı gibi), bir SP6 kiti kullanın.
- İn vitro transkripsiyon reaksiyonunu aşağıdaki gibi birleştirin: 2 μg DNA şablonunu veya 2 μL 10x Reaksiyon Tamponu, 10 μL 2x Ribonükleotid Karışımı ve 2 μL 10x Enzim Karışımı ile 6 μL'ye kadar sindirin.
- 2-4 saat veya daha uzun süre 37 °C'de kuluçkaya yatırın (daha uzun süreler daha fazla verime yol açacaktır). İstenirse, kuluçka süresinin yarısında 1 μL Enzim Karışımı desteklenebilir.
- 1 μL RNase içermeyen DNase ekleyerek ve 37 °C'de 15 dakika kuluçkaya yatırarak DNA şablonunun sindirilmesi.
- Üreticinin protokolünü izleyen bir RNA arıtma kiti kullanarak kapaklı RNA'yı arındırın (bkz. Malzeme Tablosu). 100 μL RNase içermeyen H2O ile elute.
NOT: Bu uygulama için β-mercaptoethanol eklenmesi gerekli değildir. Bu protokolde açıklanan yöntem basit olsa da, RNA'yı arındırmak için alternatif reaktifler de kullanılabilir. - RNA'yı çökelt.
- Eluted RNA'ya 10 μL 3 M RNase içermeyen NaOAc ve 2,5 cilt (~275 μL) buz gibi RNase içermeyen %100 EtOH ekleyin.
- Reaksiyonun 15-30 dakika boyunca -20 °C'de kuluçkaya yatması. RNA'yı peletmek için 4 °C'de yüksek hızda 15 dakika döndürün.
NOT: Pelet tüpün duvarına karşı görünür olmalıdır. Peletin dönüşün sonunda nerede olması gerektiğini tahmin etmek için tüpün bu adımda santrifüjde nasıl hizalandığına dikkat etmek yararlıdır. - EtOH'u bir şırınna ile dikkatlice çıkarın, peletin ısınmasını, aşırı kurumasını veya yerinden çıkarılmasını önlemeye dikkat edin. Peleti 20 μL RNase içermeyen H2O'da yeniden boşaltın.
- Sentezin başarılı olduğundan emin olmak için RNA'yı kontrol edin. Spektrofotometredeki konsantrasyonu test etmek için 1 μL kullanın ve düşük moleküler ağırlıklı bir smear yerine bir veya iki ayrı bant olup olmadığını kontrol etmek için% 1 agarose jel üzerinde 0,5 μL çalıştırın. In vitro transkripsiyon reaksiyonunun verimi ~1 μg/μL olmalıdır.
- Aliquot ve RNA'yı -20 °C veya -80 °C'de saklayın. RNA enjeksiyonlardan hemen öncesine kadar seyreltilmez.
2. 1 hücreli zebra balığı embriyolarının mikroenjeksiyon
NOT: Optik bardak morfogenez boyunca kromatin ve hücre zarlarının her yerde ekspresyonini elde etmek için RNA başına 200-300 pg enjekte edin. RNA seyreltmenin 5-10 μL'sini hazırlayın ve 2,5-5 μL / iğne yükleyin; iğnenin kırılması durumunda ekstra plan yapın.
- Enjeksiyonlar için önceden hazırlık.
- Mikroenjeksiyon için 1 hücreli embriyoların hizalamasını ve yönünü kolaylaştırmak için bir enjeksiyon kalıp kabı hazırlayın. E3'te (standart embriyo ortamı) % 2 agarose eritin ve bir Petri kabına dökün. Katılaştıkça agarosedaki olukların izini oluşturmak için enjeksiyon kalıbını (bkz. Malzeme Tablosu)sıcak agarose'un üzerine dikkatlice yüzdürün. Agarose katılaştıktan sonra enjeksiyon kalıbını çıkarın.
NOT: Enjeksiyon kalıp yemekleri birkaç ay boyunca kullanılabilir; E3'te örtün ve kullanılmadığında 4 °C'de saklayın. - Mikroenjeksiyon iğnelerini çekin. Bir iğne çekme makinesi kullanarak mikroenjeksiyon iğneleri yapmak için cam kılcal damarları uzun bir konik içine çekin. Makineyi kullanılan kılcal damarların türüne göre programla. 1.0 x 0.78 mm borosilikat kılcal damarlar için aşağıdaki programı kullanın: ısı = 546 °C, çekme = 130, hız = 70, zaman = 90 (Şekil 2F). İğneleri bir Petri kabında saklayın ve modelleme kili ile sabitleyin.
NOT: Çekme tarifi, makineye ve filamente bağlı olarak değişecektir ve yeni filamentler her zaman kalibre edilmelidir. Her çekme turu iki iğne üretir (Şekil 2G); kazara kırılmalar ve gelecekteki deneyler durumunda yeterli iğne üretmek için bunu birkaç kez yapın. - Enjeksiyon için kapaklı RNA'yı seyreltin. Toplam 5 μL hacimde RNase içermeyen H 2 O ve 1 μL fenol kırmızısı kullanarak hemEGFP-CA'yı hem de H2A-mCherry RNA'ları 200-300 ng/μL konsantrasyona seyreltin (son fenol kırmızı konsantrasyonu% 0,1'dir). Karışımı hafifçe vurun ve toplam hacmi toplamak için kısa bir süre aşağı çevirin. Enjekte etmeden önce seyreltilmiş RNA'yı buzda tutun.
- Mikroenjeksiyon için 1 hücreli embriyoların hizalamasını ve yönünü kolaylaştırmak için bir enjeksiyon kalıp kabı hazırlayın. E3'te (standart embriyo ortamı) % 2 agarose eritin ve bir Petri kabına dökün. Katılaştıkça agarosedaki olukların izini oluşturmak için enjeksiyon kalıbını (bkz. Malzeme Tablosu)sıcak agarose'un üzerine dikkatlice yüzdürün. Agarose katılaştıktan sonra enjeksiyon kalıbını çıkarın.
- 1 hücreli embriyoların enjeksiyonu
- Balıklar üremeye başladıktan sonra, yumurtaların döllenmesini sağlamak için ~ 15-20 dakika izin verin. Bu süre zarfında, iğneyi bir P10 ve P10 mikro yükleyici uçları kullanarak RNA seyreltmenin 2,5-5 μL'si ile geri yükleyin.
- Bir pikoinjektör kullanarak, damlacığın hacmini ölçmek için bir göz merceği retikülünü (mikrometre kalibrasyon kaydırağı da yeterlidir) kullanarak iğneyi kalibre edin (kürenin hacmi = (4/3) * pi * radius^3). Enjeksiyon süresini ve basıncını enjeksiyon hacmi 1 nL olacak şekilde ayarlayın (Şekil 2I).
- Bir silindir / transfer pipet ile, enjeksiyon kalıbına dikkatlice yumurta yükleyin (Şekil 2J). Yararlıysa, embriyoları tek hücrenin enjeksiyondan önce veya enjeksiyon sırasında görülebilecek şekilde yuvarlamak için asalar kullanın. Bir mikromanipülatör kullanmak kullanıcının tercihidir.
- Embriyoları 1 hücreli aşamada enjekte edin, yumurta sarısını değil hücreyi hedef alarak(Şekil 2M). Bu, gelişmekte olan embriyonun tek tip etiketlemesini sağlayacaktır.
- Embriyoları istenen aşamaya yükseltin (standart evreleme34'egöre 12 hpf'den önce). Enjekte edilen embriyolar gecikmiş bir gelişime sahip olacaktır, bu nedenle telafi süresi için biraz daha yüksek bir sıcaklıkta (29.0-29.5 ° C) yükseltin. Öğleden sonra, embriyoları kontrol edin ve debriyajın sağlığını korumak için ölü olanları çıkarın.
3. Timelapse görüntüleme için optik veziklin evre zebra balığı embriyolarının montajı
- Montajdan önce, E3'te% 1.6 düşük eriyik agarose hazırlayın. Birden fazla görüntüleme deneyi planlıyorsanız, ~20 mL düşük eriyik agarose hazırlayın ve oda sıcaklığında saklayın. Embriyo montaj gününde, monte edilmeden önce 42 ° C ısı bloğuna yerleştirilebilen bir tüpte taze bir aliquot (1-5 mL) oluşturmak için bunu eritin.
- Embriyoları başarılı enjeksiyon ve floresan genel parlaklığı için monte edilmeden önce floresan mikroskobu kullanarak tarayın. İdeal bir örnek güçlü EGFP ve mCherry floresansına sahip olacak ve doğru gelişim aşamasında olacaktır(Şekil 3B - B'').
- Embriyoları floresan mikroskop kullanarak tarayın. Montaj için hem EGFP hem de mCherry floresanlarını güçlü bir şekilde ifade eden embriyoları seçin.
- 11 hpf olan embriyoları seçin. Embriyoları düzgün bir şekilde sahneleyene kadar sayma somitleri34; at 11 hpf 3 somites olmalıdır ve tarafından 12 hpf 6 somites vardır.
NOT: Embriyolar 12 hpf'den önce monte edilerek (11 hpf'de), numuneler zaman dilimi başladığında uygun şekilde 12 hpf'de sahnelenecektir.
- Embriyoları montajdan önce kaynatır. Bunu ince toksalarla manuel olarak veya pronaz (2 mg/mL) kullanarak kimyasal olarak yapın. Bu kadar genç embriyolarda, aforizyonun agar kaplı bir tabakta yapılması ve embriyoların hava-su arayüzüne temas etmemeleri esastır.
- Cam silindir pipet kullanarak, bir embriyoyu emin ve embriyonun camın ucuna oturması için mümkün olduğunca fazla E3 fırlatın Pasteur pipeti.
- Embriyoyu ısı bloğundan agarose tüpüne bırakın. Birkaç saniye boyunca agarose içine batmasına izin verin, sonra önce biraz agarose ve sonra embriyoyu emerek embriyonun pipet ucunda kaldığından emin olun (Şekil 3C - C').
- Görüntüleme için cam tabanlı bir kabı bir diseksiyon dürbünü altına yerleştirin. Embriyoyu ve agaroseyu cam tabanlı tabaktaki agarose damlacıklarına fırlatın. Embriyonun dorsal-down (cam altta başın üstü) olacak şekilde çok hızlı bir şekilde yönlendirmek için tokmaklar kullanın. Agarose damlacık sertleşmesine izin verin (Şekil 3D - D'). Bu aşamada embriyolar kırılgan olduğu için bu adım hızlı ama dikkatli bir şekilde yapılmalıdır. Hasarlı embriyolar timelapse görüntüleme sürecinde hayatta kalamaz.
NOT: Bu deney için tüm embriyoları tutarlı bir şekilde yönlendirmek önemlidir. Timelapse yapılırken, embriyolar optik vezikliğin büyümesi için ek alan korurken atanan görüş alanına sığmalıdır. En uygun yönelim, tüm embriyoların ön-arka eksenini "dikey" veya saat yüzünde saat 12 ve 6 boyunca hizalamaktır (Şekil 3G). - 10-12 embriyo arasında monte edin, bu da görüntülenecek olanın iki katından fazladır, böylece en iyi örnekler konfokal üzerinde değerlendirildikten sonra seçilebilir (Şekil 3E - E'). Örnekler yaş, floresan etiketlemenin homojenliği ve montaj yönünün hassasiyeti açısından değerlendirilecektir.
- Embriyoları monte ettikten sonra, pipet yemeğin altını tamamen örtmek için daha agarose, böylece tüm ayrı agarose damlacıklarını tek bir büyük agarose diske kaplar (Şekil 3F - F'). Yeterli miktarda agarose, bireysel embriyo damlacıklarının veya tüm diskin çanağın dibinden kalkmamasını ve gözden uzak yüzmemesini sağlayacaktır.
- Sertleştikten sonra, numuneleri timelapse görüntüleme deneyi süresince nemli tutmak için Agarose'u E3 ile kaplayın (binanın nemine ve çevreye bağlı bir endişe).
NOT: Trikain bu spesifik timelapse görüntüleme deneyleri için gerekli değildir, çünkü bu embriyolar yeterince gençtir; bununla birlikte, istenirse ve embriyoların daha eski aşamalarıyla çalışıyorsanız, agarose'un kendisine trikain eklenebilir ve medyada kaplanabilir. - Kağıt üzerinde, zaman çizer kurulumu sırasında kolay referans için embriyoların bir haritasını çizin. Bu, pozisyonu daha sonra her numunenin genotipiyle ilişkilendirmek için çok önemlidir.
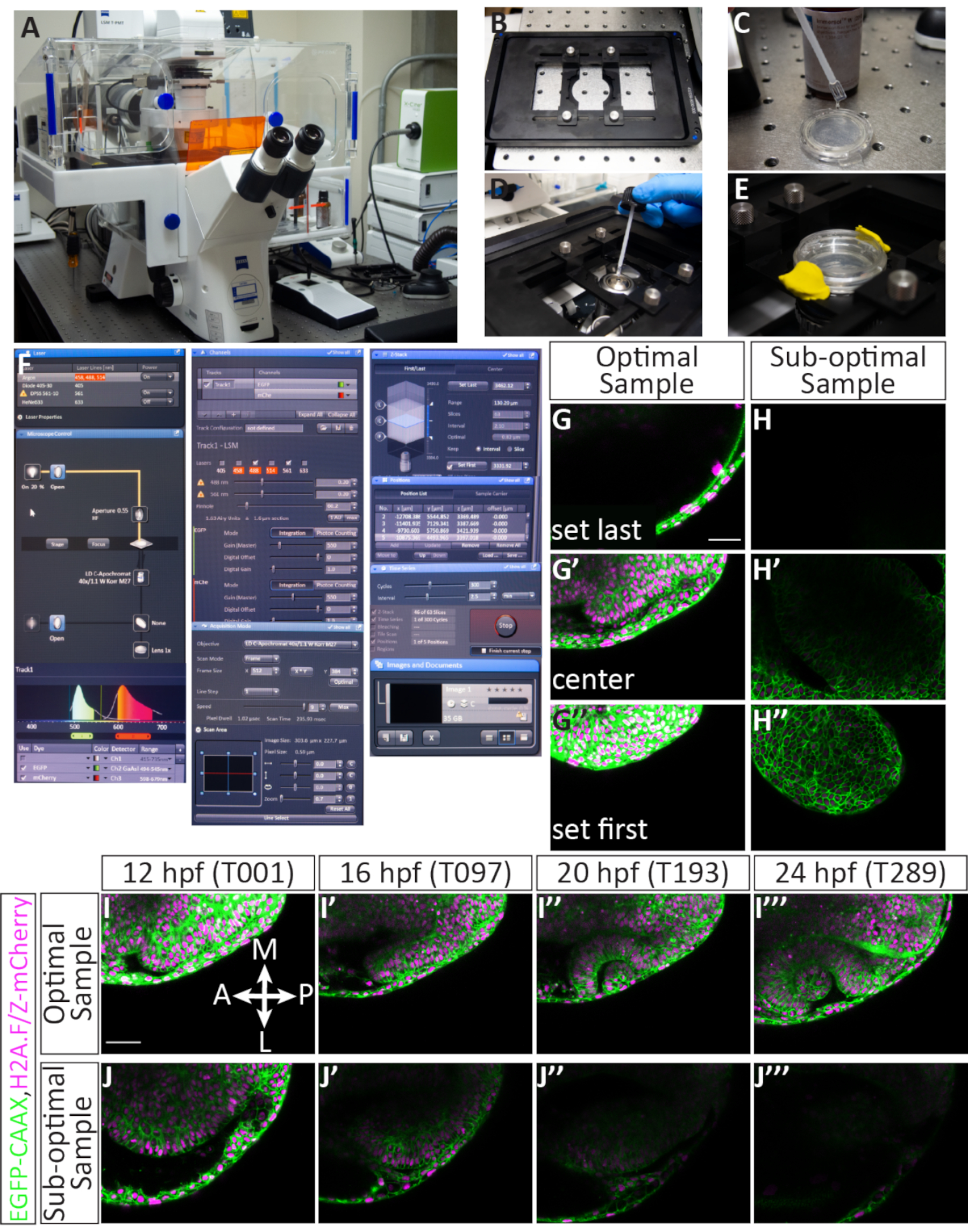
4. Lazer taramalı konfokal mikroskop ile birden fazla konum konfokal timelapse
NOT: Bu timelapse görüntüleme protokolü, üreticinin yazılımı ile donatılmış bir lazer tarama konfokal mikroskobu ile kullanılmak üzere tasarlanmıştır (bkz. Malzeme Tablosu). Bu sistem, Z yığınlarının hızlı bir şekilde edinimine izin veren piezo Z-stage bir cihazla donatılmıştır. Bu protokolde kullanılan lazer hatları 488 nm Argon iyon lazer ve DPSS 561 nm lazerdir. 561 nm lazer, mCherry floroforunu görüntülemek için çok uygundur: mCherry uyarlamanın (587 nm) zirvesine yeterince yakındır ve iyi güçlendirilmiştir.
- Konfokal'ı kurun.
- Konfokal üzerinde güç ve masaüstü bilgisayarda oturum açın.
- Piezo Z-stage kesici ucu (Şekil 4B) yükleyin, ardından satın alma yazılımını başlatın.
- Edinme modundaki yazılımda, açılır menüyü ( Şekil 4F ) kullanarak 488 ve561lazerleri açın. Lazerlerin ısınması birkaç dakika sürer, bu nedenle bu adım zaman kazanmak için erken gerçekleştirilir.
NOT: Edinme parametreleri aşağıdaki gibidir: Z yığını, Zaman Serisive Konum kutularını işaretleyin. Çerçeve Boyutunu 512 (X) x 384 (Y), Tarama Hızını 9'a, Tarama Modunu çift yönlüye, Yakınlaştırma modunu 0,7'ye, Pinhole'u 60,2'ye (1,63 Havadar Birim ~= 1,6 μm bölüm) ve Z aralığını 2,1 μm'ye ayarlayın. Bu, 303,6 μm x 227,7 μm görüntü boyutu ve 0,59 μm piksel boyutu sağlayacaktır. 488 ve 561 lazerler kontrol edilmeli ve aynı piste atanmalıdır; EGFP algılama aralığı 494-545 nm ve mCherry algılama aralığı 598-679(Şekil 4F). - Cam kabın altını, hava kabarcıklarından kaçınmaya dikkat ederek, suyun kırılma indeksine uyan daldırma ortamı ile serbest bir şekilde kaplayın (Şekil 4C). 40x su hedefini seçin ve hedefe küçük bir daldırma ortamı damlası uygulayın (Şekil 4D). Cam tabanlı kabı sahne kesici ucuna sabitleyin, embriyoları bir gecede nemli tutmak için E3 uygulayın, ardından plastik kapağı kabın üzerine kapatmak için modelleme kili kullanın (Şekil 4E). Yemekle temas etmek için amacı yükseltin.
- Örnekleri tarayın ve timelapse için hazırlanın.
- Alma sekmesinden Bul sekmesine geçin ve iletilen ışığı açmak için ampul sembolüne tıklayın. İletilen ışıkla, joystick ile ilk örneği bulun: gözle, önce numuneyi ışık huzmesine taşıyın, ardından göz merceği merkeze kullanın ve optik bir vezikliğe odaklanın.
- Edinme sekmesine dönün. Z yığını, Zaman Serisive Konumlar kutularının işaretli olduğundan ve lazerlerin kullanım için ısındığından emin olun.
- Çerçeve Boyutunu 512 (X) x 384 (Y), Tarama Hızını 9, Tarama Modunu çift yönlü olarak ayarlayın ve Yakınlaştırma'yı 0,7 olarak ayarlayın.
- Kanallar başlığı altında, Power 2.0 ve Gain 550'ye 488 ve 561 lazerleri atayarak başlayın (bu daha sonra ayarlanabilir). Pim deliğini 60,2 olarak ayarlayın (1,63 Havadar Ünite ~= 1,6 μm kesit).
- Sürekli düğmesini kullanarak taramaya başlayın. İnce odak düğmesiyle, numuneyi Z ekseninde bulun ve X-Y eksenini çerçeveye yerleştirin. Taramayı durdurun.
- Konumlar başlığı altında, XYZ bilgilerini ilk örneğe kaydetmek için Ekle'ye tıklayın (Şekil 4F). Bunu yalnızca örneklerin zamanlanmış olması için yapın. Güçlü floresanları ve optimum montajları için numuneleri seçin.
- Bul sekmesine geçin, bir sonraki örneğe geçin ve örnekleri seçme konusunda seçici davranarak 4.2.5-4.2.6 adımlarını yineleyin.
- Örnek konumları sonlandırın, z aralığı atayın ve sonra zamanlamaya başlayın. Tüm örnekler seçildikten sonra (2,5 dakikalık toplam zaman aralığında 4-5 örnek görüntülemek mümkündür) ve konumlar kaydedildikten sonra, her numuneyi gözden geçirin ve görüntüleme çerçevesindeki XYZ konumunu ayarlayın.
- İlk konumu vurgulayın ve Taşı 'yıtıklatın. Sürekli tarama yaparken, optik vezikliti çerçeve içinde hizala. Ön ve distal bölgelerde optik veziklin ve beyne göre geniş alan bırakın.
- Ardından, ince odak düğmesiyle Z yönünde ilerlerken İlk ve Son Ayarla'yı seçerek ilk ve son Z dilimlerini atayın. Optik kabın büyümesini karşılayacağından, toplam dilim sayısını ~63 olarak koruyun. Büyüme için yer açmak için optik veziklinin ventral tarafında ekstra yer sağlayın.
- İlk ve son Z dilimleri ayarlandıktan sonra, Z yığınının ortasına gitmek için C düğmesine tıklayın. Her iki lazer için lazer gücünü ve kazancınızı ayarlayın; mümkünse lazer gücünü 5'in altında tutun ve gerekirse daha yüksek bir kazanç kullanın. Taramayı durdurun. Güncelleştir 'i tıklatarak konum bilgilerini güncelleştirin.
- Konum 2'ye tıklayarak bir sonraki konuma geçin. Atanan her pozisyon için 4.3.1-4.3.3 adımlarını yineleyin. Lazer gücü ve kazancı, tüm numuneler yeterince aydınlatılacak şekilde ayarlanmalıdır.
- Hem konum bilgileri hem de lazer ayarları tamamlandıktan sonra Zaman Serisi ayarlarını atayın. Zaman Serisi başlığı altında, Zaman Aralığı'nı 2,5 dk'ya ve Döngüleri 300'e atayın ( Şekil4F). Döngü sayısı, optik fincan geliştirme tamamlandığında zaman diliminin 24 hpf aşamasını geçmesi için hesaplanır.
- Zamanlamaya başlamak için Başlat'ı tıklatın. İlk tam döngüyü izleyin: bir tam döngünün (tüm konumları görüntüleme) 2,5 dakikadan az olduğundan emin olmak için bir zamanlayıcı kullanın ve her konumun doğru göründüğünü doğrulayın. Mikroskop bir gecede bağımsız olarak çalışır ve izlenmesi gerekmez.
NOT: Konfokal oda sıcaklık kontrollü olmasına rağmen, oda sıcaklığı 20-25 °C'dir. Ekipmanın çalışması ve lazerlerin taranmasıyla, sahnedeki sıcaklık 28,5 ° C'ye daha yakın görünüyor, çünkü embriyonik gelişim, morfolojik yer işaretleriyle görsel olarak tahmin edildiği gibi beklenen hızda ilerler. - Bu ayarlarla, timelapse 12.5 saat sürecektir. Zaman çakışması tamamlandıktan sonra dosyayı kaydedin. Büyük (~40 GB) olacak ve satın alma yazılımı kullanılarak kaydedildikten sonra bireysel pozisyonlara ayrılabilir.
Sonuçlar
Floresan RNA'ların enjeksiyonu hücre altı etiketlemeyi sağlar
Hücrenin plazma zarını ve histonlarını etiketleyen RNA'lar, optik bardak morfogenezinin altında yatan bireysel hücre morfolojilerini ve hareketlerini yakalamak için enjekte edilir. Şekil 2, zebra balığı embriyolarının 1 hücreli aşamada mikroenjecting sürecinin her adımını göstermektedir. Kısaca, zebra balıkları çiftleşir (Şekil 2E) ve embriyolar toplanır ve enjeksiyon kalıbına yüklenir (Şekil 2J). Bir mikroenjeksiyon iğnesi doldurulur (Şekil 2H) ve embriyolar tek kişilik etiketleme elde etmek için gerekli olan tek hücreye (Şekil 2K-M) doğrudan enjekte edilir (Şekil 2M, Şekil 4I-I''', Film1). Tek tip etiketlemeye ek olarak, sağlam floresan gözlenir ve gelişim bozulmadan ilerler (Şekil 3B-B'').
Etkili montaj, 12 - 24 hpf'den timelapse görüntüleme OCM için gereklidir
Uzun bir süre boyunca ortaya çıkan görüntü optik fincan morfogenezine göre, hem ideal embriyoları seçmek hem de her numuneyi gömerken uygun şekilde konumlandırmak önemlidir. Şekil 3, 11 hpf embriyonun başarılı bir şekilde montesi için ayrıntılı bir hesap göstermektedir. Enjekte edilen embriyolar ilk olarak yeterli floresan için taranır(Şekil 3B',B'',F'',F'''). Bireysel embriyolar agaroseya daldırilir (Şekil 3C,C') ve tek bir agarose damlasına dorsally monte edilir (Şekil 3D,D'). Tüm örnekler agarose'un kolektif bir diskine monte edilir (Şekil 3E-F'). Embriyolar doğru, yeterince floresan ve yeterince monte edildiğinde (Şekil 3G), örnekler tüm organın görüntülenmesini sağlayan zaman akışı sırasında görüntüleme çerçevesinde kalacaktır (Şekil 4G-G'',I-I''', Film1). Bu adımlardan herhangi birinin gerçekleştirilememesi, timelapse görüntüleme için en uygun örnek olmayan monte edilmiş bir embriyo ile sonuçlanacaktır. Dönme derecesindeki en ufak bir değişiklik, zamanlama sırasında embriyo büyümesinin yönü üzerinde dramatik bir etkiye sahip olacaktır. Şekil 3'te,daha arka bir zaman ile sonuçlanacak aşırı döndürülmüş bir embriyo (Şekil 3I) ve sadece en ön dokunun gözlemlenebildiği az döndürülmüş bir embriyo (Şekil 3J) dahil olmak üzere en uygun altı rotasyonlar gösterilmiştir. Montaj sırasında uygun döndürmeye ek olarak, çerçevenin yönüne göre monte edilen örneklerin başarılı bir zaman gölgesi verme olasılığı daha yüksektir(Şekil 4G-G'', I-I''', Film1). Optimal numuneler, bir saat kadrağı üzerinde saat 12 ve 6 ile aynı hizada ön-arka eksen ile çanak üzerinde dikey olarak yönlendirilmiş olanlardır (Şekil 3G). Alt optimal örnekler, yatay veya çapraz olanlar gibi bu eksende yönlendirmeyen numunelerdir (Şekil 3H). Bu örnekler zamanlamaya devam ettikçe görüntüleme çerçevesinden çıkacak ve daha fazla analiz için kullanılamaz (Şekil 4H-H'', J-J''').
Gelecekteki analizler için uygun bir timelapse veri kümesi edinme
Doğru enjekte edilen, yükseltilen ve monte edilen embriyolar başarılı konfokal görüntülenmiş örnekler sağlayacaktır. Şekil 4, timelapse için en uygun örneği göstermektedir: floresan bile vardır ve örnek 12 hpf'dir. Görüntüleme için z yığını, ilk dilimin optik veziküle sadece dorsal olacak şekilde atanır, son dilim ise 12 hpf optik vezikül(Şekil 4G-G'') ventral sınırının ötesinde birkaç dilim bırakır. Ventral tarafa yönelik bu önyargı, ventral yönde doku büyümesi için fazla alan sağlar. Bu faktörler, karşılandığında, en uygun timelapse ile sonuçlanır (Şekil 4I-I''', Film 1). En uygun alt örnek mozaik veya loş floresan vardır, zaten geçmiş 12 hpf geliştirmiştir (optik fincan morfogenez devam ederken) veya Z-yığınında veya XY çerçevesinde zayıf bir şekilde konumlandırılmıştır (Şekil 4H-H''). Bu kriterler karşılanmadığında, timelapse artık geliştikçe dokunun tüm 3D hacmini yakalayamaz ve daha fazla analiz için kullanılamaz (Şekil 4J-J''').

Şekil 1: Zebra balığı optik bardak morfogenezinin 4D timelapse görüntüleme iş akışı. (A) İlgi çekici hücre altı yapıları floresan olarak etiketlemek için, uygun kapaklı mRNA'ları sentezlemek. (B) Mikroinject mRNA'ları 1 hücreli aşamada zebra balığı embriyolarına sokun. (C) Embriyolar, optik veziklin aşamasında, ~11 saat döllenme sonrası veya 3 somit aşamasında ters konfokal mikroskop için dorsally veya head-down monte edilir. (D) Tek bir timelapse görüntüleme seansı sırasında birden fazla embriyo ardışık olarak görüntülenebilir. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 2: 1 hücreli zebra balığı embriyolarının mikroenjecting için görsel bir rehber. (A) Bir enjeksiyon kitine dahil edilmesi gereken öğeler (saat yönünde): tüm kiti tek kullanımlık nitril eldivenlerle tutabilen orta büyüklükteki kutu; mikroenjeksiyon iğneleri içeren bir tabak; kümesler; bir aliquot mineral yağ ve parafilm karelerini kesmek; mikro yükleme ipuçları; buz üzerinde seyreltilmiş RNA; agarose enjeksiyon kalıbı; işaretleyici; silindir/transfer pipet; ve P10 pipet. (B) Mikroenjeksiyon kurulumu: harici bir sıkıştırılmış gaz kaynağına bağlı bir piko-enjektörün yanındaki diseksiyon mikroskobu. Pico enjektörüne bir ayak pedalı ve mikromanipülatör takılır. Mikromanipülatör bir iğne tutucu ile donatılmıştır ve mikroskop aşamasının üzerinde yerleştirilmiştir. (C) Bir tarafta 90 derecelik açıya ve diğer tarafta 45 derecelik açıya sahip altı sıra oluk içeren bir agarose enjeksiyon kalıbı. (D) Pico enjektör ön paneli. Basınç göstergesi 7.8 PSI okur; bu, Penjekteedilen düğme ile ayarlanabilir. Altında, enjekte etmek için basıncı manuel olarak tetiklemek, enjeksiyon süresini değiştirmek veya sıvıyı iğnede temizlemek, doldurmak veya tutmak için düğmeler vardır. Ekleme süresi 08 olarak ayarlanır (8 x 10 milise eşdeğerdir). Pçıkışına bir çıkış hortumu bağlanır ve enjeksiyon iğnesine bağlanır. Basınç ölçer kaynak anahtarı enjeksiyon basıncı ayarlanırken Penjeksiyonuna ayarlanır ve enjeksiyonlar yapılmazken iğneye uygulanan bazal basıncı ayarlamak için Pdengesine geçilebilir. Pdengesi regülatörü, Penjekte regülatörün yanındadır. Güç düğmesi, makinenin açık olduğunu belirtmek için yanar. (E) Yetişkin zebra balıkları, yetişkin balıkları döllenmiş yumurtalardan ayıran ve kolay toplanmasına izin veren ağ tabanlı yuvalanmış bir tank içeren küçük üreme tanklarında çiftleştirilir. Sığ su koşullarını taklit etmek için su seviyesi düşürülür ve çiftleşme davranışını başlatmak için tank bölücüler çıkarılır. (F) Mikropipette (iğne) çekme ön paneli: iğneleri çekmek için kullanılan özel koşulları gösteren bir ekran içerir. Basınç ayarı P = 500'dür ve ardından programın düzenlendiği son tarih ve saat, program numarası, HEAT = 546, PULL = 130, VEL = 70 ve TIME = 90'dır. (G) Mikropipette (iğne) çekirdiciye bir cam kılcal damar yerleştirilir ve yerine sabitlendirilir. Her programlanmış çekme çalıştırması iki uzun konik iğne (kırmızı ok ucu) ile sonuçlanır. (H) RNA seyreltme bir iğneye geri yüklenir; fenol kırmızı boyası çözeltinin kolay görselleştirilmesini sağlar. (I) İğnenin ucu kırıldıktan sonra, RNA'nın bolusları diseksiyon dürbününüz üzerinde bir göz merceği retikül kullanılarak ölçülür. Bu stereomikroskop için, 30x toplam büyütmede 6 karma işaret 1 nL ses seviyesine eşittir. (J) Döllenmiş yumurtalar agarose enjeksiyon kalıbına yüklenir ve bir makaralı pipet kullanılarak kalıbın olukları boyunca dağıtılır. (K) Tek hücreli evre embriyoları içeren enjeksiyon kalıbı mikroskop altına yerleştirilir ve embriyolar ardışık olarak enjekte edilir. (L) Enjeksiyon iğnesi tek hücreyi hedef alan her embriyoya yerleştirilir. (M) Hücreye girdikten sonra, ayak pedalı düzenlenmiş miktarda basıncı tetiklemek ve embriyonun hücresine 1 nL RNA salınmasını tetiklemek için kullanılır (fenol kırmızısı ile görselleştirildiği gibi). Ölçek çubukları: I = 0,1 mm; L, M = 0,5 mm. Bu rakamın daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 3: Görüntüleme için embriyoların montajı için görsel bir kılavuz. (A) Montaj kurulumu (soldan sağa): 42 ° C'ye programlanmış bir ısı bloğu, düşük erimiş agarose ısıtma tüpleri; bir silindir pipet; bir çift kümes çarık; dekonsiyone embriyolar içeren agarose kaplı bir tabak; ve metal halide floresan lambaya bağlı bir diseksiyon stereomikroskop. (B,B',B'') EGFP-CAAX mRNA ve mCherry-H2A mRNA enjekte edilen 11 hpf embriyo, sırasıyla brightfield, GFP floresan ve mCherry floresan altında gösterilmiştir. (C,C') Agarose içeren bir cam pastör pipet ve ucunda oturan bir embriyo; C' büyütülmüş bir görünümdür. (D,D') Hem mikroskop aşamasından hem de mikroskop göz merceğinden görülen cam tabanlı bir kabın üzerine düşük erimiş agarose damlacıklarına monte edilmiş bir embriyo. (E,E') Cam tabanlı bir kabın üzerine düşük erimiş agarose'un bireysel damlacıklarına monte edilmiş, hem mikroskop aşamasından hem de mikroskop göz merceğinden görülen 12 embriyo. (F,F',F'',F''') Tüm monte edilmiş embriyolar, hem mikroskop aşamasından hem de parlak alanda gösterilen mikroskop göz merceğinden görülen, kabın altını doldurmak için tam bir agarose tabakası ile kaplanır; F'' GFP floresan; ve F''' mCherry floresan. (G) 12 hpf'de optimum bir örnek dorsally monte edilir ve aynı düzlemde her iki optik vezikül ile saat 12 ve 6 doğrultusunda dikey olarak yönlendirilir. (H) Alt optimal bir örnek: monte edilmiş dorsally ve optik vesicles birbirleriyle düzlemde olmasına rağmen, bu örnek çapraz bir eksende yönlendirilir ve geliştirme ilerledikçe çerçeve boyutunun dışına çıkar. (I) Alt optimal bir örnek: bu örnek aşırı döndürülür ve dorsal bir montaj değildir, sonuç olarak optik vezikliğin ön kısmı zamanlapsta yakalanmaz. (J) En uygun alt örnek: Bu örnek az döndürülür ve dorsal bir montaj değildir, sonuç olarak optik vezikliğin arka kısmı zamanlapsta yakalanmaz. Noktalı çizgiler her optik veziklikleri gösterir. Ölçek çubukları: B = 0,1 mm; D' = 1,5 mm; E', F' = 2,5 mm; G = 0.1 mm. Bu rakamın daha büyük bir versiyonunu görüntülemek için lütfen buraya tıklayın.
{kind=link}

Şekil 4: Birden fazla pozisyon zamanlayıcısı ve olası sonuçlar ayarlama. (A) Lazer tarama konfokal mikroskop. (B) Piezo Z-sahne kesici ucu. (C) Agarose'a gömülü embriyolar içeren bir cam dip kabının tabanı, suyun kırılma indeksiyle eşleşen daldırma ortamında kaplanır. (D) 40x W hedefine (uzun çalışma mesafesi) daldırma ortamı damlacık yerleştirilir. (E) Tabak, sahne kesici ucu ile yerinde tutulur, agarose tabakasının üzerine E3 eklenir ve kapak modelleme kili ile sabitlenerek sabitlanır. (F) Edinme yazılımı ayarları: Açılan menüden istenen lazer çizgileri açılır. EGFP-CAAX RNA ve H2A-mCherry RNA ile enjekte edilen embriyolar için Argon ve DPSS 561-10 lazerleri açılır. Kırmızı vurgu Argon lazerinin ısındığını gösteriyor. Numuneyi hedefin üzerinde bulmak için, iletilen ışık mikroskop kontrol panelinden açılır. Hem EGFP hem de mCherry Parça 1'e (eşzamanlı görüntüleme için) atanır ve her dedektörün aralığı ayarlanır. Burada, EGFP aralığı 494-545 nm ve mCherry aralığı 598-679 nm olarak ayarlanır. Altında Kanal menü, lazer gücü ayarlanabilir. Lazerler ısındıktan sonra, güç 5.0'a kadar artırılabilir ve kazanç (ana) ayarlanabilir. Pim deliği 1,63 Havadar Üniteye veya 1,6 μm bölüme eşit olan 60,2 olarak ayarlanır. Altında Edinim menü, çerçeve boyutu 512 x 384, tarama hızı 9.0, ortalama çift yönlü ve yakınlaştırma 0.7 olarak ayarlanır. Altında Z-Yığını menüsünde, konfokal tarama yapılırken Z eksenindeki ilk ve son konum ayarlanır. Aralık (adım boyutu) 2,1 μm olarak ayarlanır. İlk ve son Z pozisyonunu seçerken, genellikle standart 63 dilim sayısı korunur; bu, gözün genel büyümesini barındırır. "Piezo kullan" seçeneği seçilir. Altında Pozisyon menüsünde, seçilen her konum atanan bir sayı ve X, Y ve Z konumu dahil olmak üzere listelenir. Ekleme, güncelleme veya kaldırma dahil olmak üzere görüntüye embriyo sayısını (ayrı pozisyonlar) kontrol etmek için ek düğmeler vardır. Altında Zaman Serisi menüde, döngü 300 (elde edilecek Z yığınlarının sayısı) ve aralık (zaman adımı) 2,5 dakika olarak ayarlanır. Ayarlar tamamlandıktan sonra, start tuşuna basılarak timelapse alma işlemi başlatılır. Yazılım, Z yığınının dilimini, döngüsünü ve şu anda tarama yapan konumu gösterecektir. Zaman faraşı tamamlandığında, dosya Resimler ve Belgeler panel. (G-J) Hücre zarları (green) EGFP-CAAX ve hücre çekirdeği (kromatin, magenta) H2A-mCherry RNA'ları ile etiketlenmiştir. (G,G',G'') En iyi şekilde monte edilmiş bir örnek için ilk ve son Z dilimini ayarlama örneği. İlk Z dilimi (sırt tarafında) dorsal ektoderm pahasına gelirken, son Z dilimi (ventral tarafta) ventral yönde büyümesini karşılamak için optik vezikliğin çok altındadır. (H,H',H'') En uygun alt numunenin ilk ve son Z dilimini ayarlama örneği. Örnek zayıf mCherry floresan vardır ve çapraz monte edilir, böylece gelişmekte olan optik fincanın zaman akışı boyunca çerçevede kalması olası değildir. (Ben, ben,','') En uygun örnekten bir timelapse örneği. Z yığınının ortasından tek dilimli görüntüler, 63 dilim hacminin tamamından 4 zaman noktasında (T değeri) alınır. Br, beyin; NR, nöral retina; RPE, retina pigmentli epitel; Le, lens. (J,J',J'',J''') Yetersiz örnekten zaman gölgesi örneği. Bu durumda, örnek Z düzleminin dışına taşındı ve ön optik kabın sadece eğik bir kısmı yakalandı. Ölçek çubukları: G, I = 50 μm. Bu rakamın daha büyük bir sürümünü görüntülemek için lütfen buraya tıklayın.
{kind=link}
Film 1: Optimal bir vahşi tip embriyoda optik bardak morfogenezinin timelapse. EGFP-CAAX (plazma zarları, yeşil)ve H2A ile etiketlenmiş vahşi tip embriyonun dorsal görünümü. F/Z-mCherry (kromatin, macenta). Timelapse ~12-24 hpf'dir ve tüm 4D veri kümesinden tek bir konfokal bölüm içerir. Zaman aralığı z yığınları arasında 2,5 dk'dır ve saniyede 22,5 kare olarak gösterilir. Bu filmi indirmek için lütfen tıklayınız.
Tartışmalar
Burada, optik kupa morfogenezinin toto etiketleme ve 4D timelapse görüntüleme için bir protokol açıklıyoruz. Farklı hücre altı bölmeleri işaretlemek için kaplanmış RNA kodlama floresan proteinleri üretme sürecinde adım atıyoruz; zebra balığına 1 hücreli embriyo enjekte etmek; çoklamalı görüntüleme için agarose içine 11 hpf embriyo gömmek; ve optik kupa morfogenez (12-24 hpf) süresince birden fazla embriyonun 4D veri kümelerinin eldei.
Bu bilgi yoğun veri kümeleriyle sayısız soru yanıtlanabilir. 4D veriler çeşitli şekillerde görselleştirilebilir ve nicel olarak analiz edilebilir. Bu protokolün kapsamı dışında olsa da, gerçekleştirilebilecek şeylerin türlerine örnek olarak bazı hedeflerimizi ve standart uygulamalarımızı buraya dahil ediyoruz. Tabii ki, nicel görüntü analizi yöntemleri sürekli geliştirilmektedir ve hem ticari olarak mevcut hem de özel olarak üretilmiş araçlar kullanılabilir. Daha önce bu tür yöntemler kullanılmadıysa, edinilen veri kümelerinin tercih edilen nicel analiz yaklaşımı için yeterli olduğundan emin olmak için bazı iyileştirmeler üzerinde çalışmaya hazırlanmalıdır.
Dosyaların boyutu nedeniyle 4D veri kümelerini görselleştirmek ve nicel olarak değerlendirmek zor olabilir. Satın alma yazılımı, veri kümelerini tek tek embriyolara ayırmak için kullanılabilir ve ImageJ / Fiji, konfokal dosyaları ticari formatlardan her zaman noktasının ayrı bir dosya olarak kaydedildiği daha standart tif yığınlarına dönüştürmek için kullanılabilir. Bu, dosya boyutlarını azaltır ve dosya biçimlerini standartlaştırır. Her zaman noktasından tek tek optik bölümler ImageJ/Fiji kullanılarak 2D (XY) timelapse olarak monte edilebilir ve verilerin hızlı 2D görselleştirilmesini ve değerlendirilmesini sağlar. Film 1 tam olarak bunun bir örneğidir: Şekil 4I–I''''de gösterilen en uygun numunenin zaman farzı olarak monte edilen zaman içinde tek bir optik bölüm. Oradan, 3D ve 4D görselleştirme için, genellikle fluoRender35,36, serbestçe kullanılabilir, ancak belirli grafik kartı gereksinimlerine sahiptir. FluoRender kullanarak, 3B işlenmiş verileri herhangi bir eksende döndürebilir, veri kümesini zaman içinde 4D olarak görselleştirebilir, herhangi bir düzlemde kesitler oluşturabilir ve döndürmeleri ve görselleştirmeleri bir film veya dizi tif görüntü olarak kaydedebilir.
Nicel analiz açısından cevaplanması gereken çok sayıda soru vardır. Optik bardak morfogenezinin altında bulunan hücre davranışlarını anlama konusunda kendi özel hedeflerimize yardımcı olmak için şirket içinde yazılım geliştirdik. Programımız, LongTracker, nükleer sinyali hücreleri izlemek için bir proxy olarak kullanır13. Bu verilerle hücrelerin ne zaman, nerede ve nasıl hareket ederek hareket ederek hareket ettirdiğini belirleyebiliriz; hücrelerin ne kadar hızlı ve ne kadar uzağa hareket etmesi; ve hücrelerin ne sıklık tadılacağını. Kendi yazılımımıza ek olarak, 4D hücre takibi için birden fazla ticari ve özel olarak oluşturulmuş seçenek vardır. Biz de hücre segmentasyonunu gerçekleştirmek ve nöral retina içinde hücre şekli ve organizasyonuni ölçmek için LongAxis programını geliştirdik37. Ancak, LongAxis'e giren veri kümeleri, yüksek çözünürlükte alınan tek Z yığınlarıdır. Kalıcı bir zorluk, hücrelerin güvenle bölünebileceği ve morfolojilerinin tahmin edilebileceği kadar yüksek çözünürlüğe sahip zamanlanmış veri kümeleri oluşturuyor. Alternatiflerden biri, biz ve diğerlerinin gelişmekte olan göz 8,11,13'tegerçekleştirdiğimiz gibi, hücre şeklinindoğrudan görselleştirilmesi için Kaede gibi foto-dönüşümlü bir florofor kullanarak mozaik (seyrek) etiketlemedir. Bu, hücre segmentasyon sorununu basitleştirir ve fluoRender'da 3D işleme yoluyla hücre şekli ve boyutu kolayca ölçülebilir.
Bu protokolün her adımı özellikle amaçlarımız için optimize edildi. Bu protokolün özgüllüğü çeşitli sınırlamalarla sonuçlanır: protokol, yazıldığı gibi, zebra balığı göz gelişiminin diğer yönlerini (retinal nörogenez gibi), diğer göz yapılarını, diğer gelişim aşamalarını veya diğer hücre altı bölmeleri görüntülemek için optimize edilmemiştir. Gömmenin yönü, görüntüleme hızı ve etiketlerin hepsi biyolojik sorularımızı cevaplamamızı sağlamak için tasarlanmıştır. Örneğin retinal nörogenezin görüntülenmesi için, embriyonun burada açıklandığı gibi dorsal yönelimi, ilgi çekici belirli özelliklerin görselleştirilmesine izin vermeyebilir ve görüntüleme hızı uygun olmayabilir. Protokolün birçok yönü, kişinin özel ilgi alanlarına ve hedeflerine bağlı olarak çeşitli ihtiyaçlar için uyarlanabilir ve değiştirilebilir. İlk olarak, RNA enjeksiyonlarının kullanılması etiketleme işlemini çok esnek hale getirir. Floresan protein füzyon yapıları, ilgi çekici hücre altı yapıları etiketlemek için kullanılabilir ve etiketlemeyi optimize etmek için RNA enjeksiyon miktarı değiştirilebilir. Fotokonverter florofor Kaede ile yaptığımız çalışmalara dayanarak, RNA enjeksiyonları 12 hpf 11,13ile sona ermiş bir çeviri patlamasını destekliyor gibi görünüyor. RNA enjeksiyonundan yüksek düzeyde floresan protein ifadesi fotobleaching ile mücadele edebilir, ancak böyle bir yaklaşım floresan ilgi etiketinin sürekli ifadesini desteklemez. Embriyoların daha sonraki aşamalarda görüntülenmesi gibi sürekli ifade gerekliyse, transgenik çizgiler bir seçenektir ve zebra balıklarında yeni çizgiler oluşturmak basittir24.
Daha sonra, protokol geliştirmenin sonraki aşamaları için uyarlanabilir. Pigment daha sonraki aşamalarda görüntülemeyi engelleyebileceğinden, embriyolar pigment oluşumunu inhibe etmek için fenilthiourea (PTU) ile tedavi edilebilir veya pigment sentezi için genetik mutantlar kullanılabilir. Embriyo seğirmesini önlemek için agarose ve embriyo ortam kaplama çözeltilerine trikain eklenebilir ve yüzde agarose gerektiği gibi ayarlanabilir. Göz büyüdükçe, montaj yönünü değiştirmek gerekebilir; burada, embriyoları dorsally monte ediyoruz, ancak daha sonraki aşamalarda, ilginin yapısına bağlı olarak yanal veya ön olarak monte etmek daha mantıklı olabilir. Farklı uzamsal ve zamansal ölçeklerde farklı işlemler gerçekleştiğinden, görüntü alımının Z adımını ve zaman adımını da optimize edebilirsiniz. Bu özellikler gerçekten sadece her laboratuvarın ihtiyaçları için ampirik olarak belirlenebilir.
Son olarak, bu protokol özellikle göz gelişiminin erken bir aşamasında nispeten yüksek bir agarose yüzdesi içine gömülü embriyolarla lazer tarama konfokal mikroskobu için geliştirilmiştir. Göz gelişimi sırasında farklı zamanlarda veya farklı yerlerde görüntüleme varsa, bu protokolün ilgi sürecine uyarlanmış olması gerekecektir. Optik mühendisleri tarafından daha fazlası geliştirilmekte olan birçok farklı görüntüleme yaklaşımı şu anda mümkündür. Her yaklaşım, görüntüleme için embriyoları gömmenin ve monte etmenin farklı yollarından farklı dosya boyutlarına ve formatlarına kadar kendi zorluklarını getirir. Maksimum uzamsal ve zamansal çözünürlüğün fotobleaching, fototoksiklik ve muazzam dosya boyutlarıyla dengelenmiş olduğu optimizasyon sürecine rehberlik etmek için girişte dikkat edilmesi gereken hususları özetliyoruz. Bu genel ilkelerin, yukarıda açıklanan pratik bilgilere ek olarak, göz gelişimindeki birçok açık soruyu incelemek için timelapse görüntüleme yaklaşımlarının oluşturulmasında başkalarına yardımcı olacağını umuyoruz.
Açıklamalar
Yazarların açıklayacak bir şeyi yok.
Teşekkürler
Kwan Lab'ın geçmiş ve şimdiki üyelerine, bu protokolün timelapse yaklaşımları ve tartışmaları üzerine çalıştıkları için minnettarız. Bu çalışma NIH (R01 EY025378 ve R01 EY025780 to KMK; F31 EY030758 için SL; ve T32 GM007464 'den MAC'e).
Malzemeler
| Name | Company | Catalog Number | Comments |
| Delta TPG Dish 0.17mm clear | Bioptechs | 0420041500C | coverglass bottom for optical compatibility |
| Disposable Pasteur Pipettes | VWR | 14672-608 | overall length 14.6 cm (53/4") |
| Dumont #5, #55, or #5SF Forceps | Fine Science Tools | 11254-20 | For style #5: straight tip, 0.05 x 0.01 mm tip, inox, 11 cm long |
| Flaming/Brown Micropipette Puller | Sutter | P-97 | |
| Immersol W | Carl Zeiss Microscopy | 4449690000000 | immersion fluid for water-emission objectives, halogen-free, low fluorescence |
| Microinjection Mold | Adaptive Science Tools | TU-1 | Six ramps, one 90 degree and one 45 degree beveled side. |
| Microloader Tips | Eppendorf | 930001007 | Microloader, tip for filling glass microcapillaries, 0.5 – 20 µL |
| mMessage mMachine SP6 Transcription Kit | Invitrogen | AM1340 | contains SP6 Enzyme Mix, 10X Reaction Buffer, 2XNTP⁄CAP Solution, GTP Solution, pTRI-Xef, TURBO DNase, Ammonium Acetate Stop Solution, Lithium Chloride Precipitation Solution, and Gel Loading Buffer and are all stored at –20°C. Nuclease-free Water may be stored at any temperature. |
| Nikon SMZ645 Stereo Microscope | Nikon | SMZ645 | used for injecting embryos (see Fig. 2B) |
| NotI-HF enzyme | NEB | R3189S | comes with Gel Loading Dye, Purple (6X) |
| NuSieve GTG Agarose | Lonza | 50081 | fine resolution, low melt agarose |
| Objective LD "C-Apochromat" 40x/1.1 W Korr M27 | Carl Zeiss Microscopy | 421867-9970-000 | |
| Olympus SZX16 Stereo Microscope | Olympus | SZX2-ZB16 | used for screening, embedding embryos (see Fig. 3A) |
| Phenol red solution | Sigma-Aldrich | P0290 | 0.5% in DPBS, sterile filtered, endotoxin tested, cell culture tested |
| Pico-liter Microinjector | Harvard Apparatus | PLI-100 | Femtoliter to microliter injections; digital readouts for injection count, time, and pressure; contains 5 pressures including inject, balance, clear, fill, and hold |
| Pipette Pump Pipettor | SP Bel-Art | F37898 | fits a disposable pasteur pipette, contains thumbwheel on side for aspirating or dispensing and plunger for quick emptying |
| Premium Thin Wall Borosilicate Glass Capillaries | Warner Instruments | G100T-4 | 1.0 x 0.78 mm |
| QIAquick PCR Purification Kit | Qiagen | 28106 | contains QIAquick Spin Columns, Buffers, Collection Tubes (2 ml) |
| RNase-free H2O | Invitrogen | AM9906 | DNase-Free, Molecular Biology Grade, RNase-Free |
| RNeasy Mini Kit | Qiagen | 74104 | contains RNeasy Mini Spin Columns, Collection Tubes (1.5 ml and 2 ml), RNase-free Reagents and Buffers |
| Stage Attachment Z PIEZO WSB 500 | Carl Zeiss Microscopy | 432339-9000-000 | |
| Stage Insert Z PIEZO WSB Universal | Carl Zeiss Microscopy | 432339-9030-000 | |
| Zeiss LSM 880 with Airyscan | Carl Zeiss Microscopy | N/A | inverted Axio Observer microscope |
| Zen Black 2.3 SP1 software | Carl Zeiss Microscopy | N/A | Zeiss Efficient Navigation (ZEN) controls all light microscope systems from Zeiss |
Referanslar
- Adler, R., Canto-Soler, M. V. Molecular mechanisms of optic vesicle development: complexities, ambiguities and controversies. Developmental Biology. 305 (1), 1-13 (2007).
- Chow, R. L., Lang, R. A. Early eye development in vertebrates. Annual Review of Cell and Developmental Biology. 17, 255-296 (2001).
- Fuhrmann, S. Eye morphogenesis and patterning of the optic vesicle. Current Topics in Developmental Biology. 93, 61-84 (2010).
- Martinez-Morales, J. R., Wittbrodt, J. Shaping the vertebrate eye. Current Opinion in Genetics and Development. 19 (5), 511-517 (2009).
- Yang, X. J. Roles of cell-extrinsic growth factors in vertebrate eye pattern formation and retinogenesis. Seminars in Cell & Developmental Biology. 15 (1), 91-103 (2004).
- Bogdanovic, O., et al. Numb/Numbl-Opo antagonism controls retinal epithelium morphogenesis by regulating integrin endocytosis. Developmental Cell. 23 (4), 782-795 (2012).
- Bryan, C. D., Casey, M. A., Pfeiffer, R. L., Jones, B. W., Kwan, K. M. Optic cup morphogenesis requires neural crest-mediated basement membrane assembly. Development. 147 (4), (2020).
- Bryan, C. D., Chien, C. B., Kwan, K. M. Loss of laminin alpha 1 results in multiple structural defects and divergent effects on adhesion during vertebrate optic cup morphogenesis. Developmental Biology. 416 (2), 324-337 (2016).
- Cavodeassi, F., Ivanovitch, K., Wilson, S. W. Eph/Ephrin signalling maintains eye field segregation from adjacent neural plate territories during forebrain morphogenesis. Development. 140 (20), 4193-4202 (2013).
- England, S. J., Blanchard, G. B., Mahadevan, L., Adams, R. J. A dynamic fate map of the forebrain shows how vertebrate eyes form and explains two causes of cyclopia. Development. 133 (23), 4613-4617 (2006).
- Gordon, H. B., et al. Hedgehog signaling regulates cell motility and optic fissure and stalk formation during vertebrate eye morphogenesis. Development. 145 (22), (2018).
- Ivanovitch, K., Cavodeassi, F., Wilson, S. W. Precocious acquisition of neuroepithelial character in the eye field underlies the onset of eye morphogenesis. Developmental Cell. 27 (3), 293-305 (2013).
- Kwan, K. M., et al. A complex choreography of cell movements shapes the vertebrate eye. Development. 139 (2), 359-372 (2012).
- Martinez-Morales, J. R., et al. Ojoplano-mediated basal constriction is essential for optic cup morphogenesis. Development. 136 (13), 2165-2175 (2009).
- Miesfeld, J. B., et al. Yap and Taz regulate retinal pigment epithelial cell fate. Development. 142 (17), 3021-3032 (2015).
- Nicolas-Perez, M., et al. Analysis of cellular behavior and cytoskeletal dynamics reveal a constriction mechanism driving optic cup morphogenesis. eLife. 5, (2016).
- Picker, A., et al. Dynamic coupling of pattern formation and morphogenesis in the developing vertebrate retina. PLoS Biology. 7 (10), e1000214 (2009).
- Rembold, M., Loosli, F., Adams, R. J., Wittbrodt, J. Individual cell migration serves as the driving force for optic vesicle evagination. Science. 313 (5790), 1130-1134 (2006).
- Sidhaye, J., Norden, C. Concerted action of neuroepithelial basal shrinkage and active epithelial migration ensures efficient optic cup morphogenesis. eLife. 6, (2017).
- Driever, W., et al. A genetic screen for mutations affecting embryogenesis in zebrafish. Development. 123, 37-46 (1996).
- Haffter, P., et al. The identification of genes with unique and essential functions in the development of the zebrafish, Danio rerio. Development. 123, 1-36 (1996).
- Nusslein-Volhard, C. The zebrafish issue of Development. Development. 139 (22), 4099-4103 (2012).
- Hoshijima, K., et al. Highly efficient CRISPR-Cas9-based methods for generating deletion mutations and F0 embryos that lack gene function in zebrafish. Developmental Cell. 51 (5), 645-657 (2019).
- Kwan, K. M., et al. The Tol2kit: a multisite gateway-based construction kit for Tol2 transposon transgenesis constructs. Developmental Dynamics. 236 (11), 3088-3099 (2007).
- Almeida, A. D., et al. Spectrum of Fates: a new approach to the study of the developing zebrafish retina. Development. 141 (9), 1971-1980 (2014).
- Baye, L. M., Link, B. A. Interkinetic nuclear migration and the selection of neurogenic cell divisions during vertebrate retinogenesis. The Journal of Neuroscience. 27 (38), 10143-10152 (2007).
- Clark, B. S., et al. Loss of Llgl1 in retinal neuroepithelia reveals links between apical domain size, Notch activity and neurogenesis. Development. 139 (9), 1599-1610 (2012).
- Das, T., Payer, B., Cayouette, M., Harris, W. A. In vivo time-lapse imaging of cell divisions during neurogenesis in the developing zebrafish retina. Neuron. 37 (4), 597-609 (2003).
- Kay, J. N., et al. Transient requirement for ganglion cells during assembly of retinal synaptic layers. Development. 131 (6), 1331-1342 (2004).
- Norden, C., Young, S., Link, B. A., Harris, W. A. Actomyosin is the main driver of interkinetic nuclear migration in the retina. Cell. 138 (6), 1195-1208 (2009).
- Suzuki, S. C., et al. Cone photoreceptor types in zebrafish are generated by symmetric terminal divisions of dedicated precursors. Proceedings of the National Academy of Sciences of the United States of America. 110 (37), 15109-15114 (2013).
- Wan, Y., et al. The ciliary marginal zone of the zebrafish retina: clonal and time-lapse analysis of a continuously growing tissue. Development. 143 (7), 1099-1107 (2016).
- Yoshimatsu, T., et al. Presynaptic partner selection during retinal circuit reassembly varies with timing of neuronal regeneration in vivo. Nature Communications. 7, 10590 (2016).
- Kimmel, C. B., Ballard, W. W., Kimmel, S. R., Ullmann, B., Schilling, T. F. Stages of embryonic development of the zebrafish. Developmental Dynamics. 203 (3), 253-310 (1995).
- Wan, Y., Otsuna, H., Chien, C. B., Hansen, C. An interactive visualization tool for multi-channel confocal microscopy data in neurobiology research. IEEE Transactions on Visualization and Computer Graphics. 15 (6), 1489-1496 (2009).
- Wan, Y., Otsuna, H., Chien, C. B., Hansen, C. FluoRender: an application of 2D image space methods for 3D and 4D confocal microscopy data visualization in neurobiology research. IEEE Pacific Visualization Symposium. , 201-208 (2012).
- Carney, K. R., Bryan, C. D., Gordon, H. B., Kwan, K. M. LongAxis: a MATLAB-based program for 3D quantitative analysis of epithelial cell shape and orientation. Developmental Biology. 458 (1), 1-11 (2020).
Yeniden Basımlar ve İzinler
Bu JoVE makalesinin metnini veya resimlerini yeniden kullanma izni talebi
Izin talebiThis article has been published
Video Coming Soon
JoVE Hakkında
Telif Hakkı © 2020 MyJove Corporation. Tüm hakları saklıdır