
Method Article
从真核细胞 中快速体内 固定和分离转化复合物
* 这些作者具有相同的贡献
摘要
我们提出了一种技术,可以快速稳定活酵母和哺乳动物细胞中具有甲醛交联的转化(蛋白质生物合成)复合物。该方法能够解剖瞬态中间体和动态RNA:蛋白质相互作用。交联复合物可用于多种下游应用,例如基于深度测序的分析方法、显微镜和质谱分析。
摘要
涉及信使(m)RNA的快速再分配和mRNA翻译的改变的快速反应与细胞的持续稳态调整有关。这些调整对于真核细胞的生存能力和"损伤控制"在营养和盐度水平,温度以及各种化学和辐射应激的波动期间至关重要。由于RNA水平反应的高度动态性,以及许多RNA:RNA和RNA:蛋白质中间体的不稳定性,因此只有使用有限数量的方法才能获得细胞质RNA状态的有意义的快照。转录组范围的、基于RNA-seq的核糖体分析型实验是控制翻译的最翔实的数据来源之一。然而,缺乏均匀的RNA和RNA:蛋白质中间体稳定会导致不同的偏差,特别是在快节奏的细胞反应途径中。在本文中,我们提供了适用于不同通透性的真核细胞的快速固定的详细方案,以帮助RNA和RNA:蛋白中间体的稳定。我们进一步提供了基于稳定RNA:蛋白质复合物与核糖体和多(核糖)体质组分共沉淀的分离示例。分离的稳定材料随后可以用作核糖体分析型实验的一部分,例如在翻译复合物分析测序(TCP-seq)方法及其衍生物中。TCP-seq风格方法的多功能性现在已经通过各种生物体和细胞类型的应用得到了证明。稳定的配合物还可以另外进行亲和纯化并使用电子显微镜成像,分离成不同的聚(核糖)体质组分并进行RNA测序,因为交联逆转很容易。因此,基于速冻和甲醛固定的方法,然后是基于沉降或其他类型的RNA:蛋白质复合物富集的方法,对于研究活细胞中快速RNA:蛋白质复合物动力学的更精细细节特别感兴趣。
引言
生物体在其生命周期中受到细胞内和细胞外动态变化的影响,这需要快速反应以维持体内平衡并确保生存。为了适应环境,真核细胞通过基因表达控制来调整其新陈代谢。基因表达控制可以在转录和/或翻译过程中进行;翻译反应通常发生得更快1,2,3,4.例如,翻译变化通常在压力发作后1-30分钟内出现,而转录水平的改变在压力暴露后数小时后出现3,4,5。由于细胞质中信使(m)RNA分子的持续可用性,翻译输出的改变可以更快地实现。相反,在转录水平上,必须合成新的mRNA分子,并且在真核生物中,从细胞核中处理和输出,在响应时间2,4,6,7,8中产生广泛的延迟。
对应激的急性转化反应通常表现为翻译输出总体下降,细胞存活所必需的蛋白质有选择性上调1,3,4,9.由于该过程的高能量消耗,降低蛋白质产量被认为是至关重要的3,7.为了促进选择性抑制和上调,一系列复杂的调节机制为转化反应服务。可以在翻译的所有阶段进行调节:多肽生物合成的启动,伸长,终止和核糖体回收10,11,12,13,但在初始阶段表现最强烈5,7,9,10,13.在起始过程中,小核糖体亚基(SSU)在真核起始因子(eIFs)的辅助下,与mRNA的5'未翻译区域(UTR)结合并扫描,直到识别起始密码子2,5,6,8,11,12,13.监管机制通常针对影响附着、扫描和开始密码子识别的 eIF。例如,起始因子eIF2,一种有助于招募引发剂Met-tRNA的基本翻译因子。iMet 到SSU,通常在压力条件下的真核生物中靶向4,6,11.在酵母中,该因子的磷酸化可以在营养剥夺和渗透应激下诱导1,4,11,14,15,在哺乳动物细胞中,氨基酸饥饿、内质网 (ER) 应激、紫外线应激、病毒感染和氧水平改变可能引发这种反应8,9,11.特异性mRNA翻译的快速上调在哺乳动物细胞对缺氧的反应中是显而易见的,其表现出缺氧诱导因子(HIFs)生物合成的全局快速翻译抑制和选择性上调。HIF是转录因子,然后在DNA转录水平上引发长期的细胞重编程8,9,16.在热应激下,在酵母中观察到类似的反应,热休克蛋白(HSP)的快速翻译表达,随后是延迟的转录水平反应17,18.除了营养剥夺和热休克外,还研究了酵母在不同氧气下转化反应8,19盐度5,磷酸盐,硫磺20,21 和氮气22,23 水平。这项研究对酵母的工业用途具有广泛的意义,例如烘焙和发酵24,25.转化反应也可能有助于进一步了解神经退行性疾病和心脏病等疾病,这些疾病的特征是细胞内应激,如氧化应激。总体而言,转化反应是基因表达控制不可或缺的一部分,有助于快速适应真核生物体中广泛的应激条件。
为了研究翻译响应,需要提供翻译环境最小扭曲快照的方法。多体分析是用于研究跨mRNA翻译的经典方法,涉及通过蔗糖梯度26,27通过超速离心分离mRNA的多(核)体质组分。该方法可用于探索单个mRNA的翻译水平(使用逆转录和聚合酶链反应等检测方法,RT-PCR26),或在全球范围内与高通量技术(微阵列或RNA-seq28,29)结合使用。一种更进化的方法是核糖体分析,它允许在全基因组尺度上研究沿着mRNA分子的细长核糖体的位置,以及推断跨转录组的翻译效率和利用主要和替代起始位点30,31。核糖体分析涉及由核糖体存在的mRNA片段的分离和测序。核糖体分析为许多条件下的翻译动力学提供了相当多的见解,包括缺氧应激,热休克和氧化应激31,32。该技术已适用于多种源材料类型,包括酵母和哺乳动物细胞。
虽然多体和核糖体分析是扩展翻译研究能力的基础,但翻译过程包括各种翻译中间体和复合物,这些方法难以捕获11,13。另一个局限性源于缺乏研究快速反应类型的能力,因为转化复合物要么通过添加特异性翻译抑制剂(抗生素)在体内稳定,导致某些核糖体分布伪影,要么在细胞特异性裂解时离体(抗生素)或非特异性(高盐或镁离子),导致短寿命或不太稳定的中间体的剥夺33, 34,35.
甲醛广泛用于交联核酸和蛋白质,例如染色质免疫沉淀(ChIP)和交联免疫沉淀(CLIP)研究。其小尺寸和优异的细胞通透性允许在36中快速的体内作用。基于甲醛的快速交联,核糖体分析方法已经扩展为翻译复合物谱测序(TCP-seq)10,36,37,38,39,40。TCP-seq,首先在酵母中开发,允许捕获所有翻译中间体,包括扫描或终止后SSU复合物和多个核糖体构型37,38,41,42。该方法已在若干研究10、38、39、41、42中使用,其中一些使用翻译抑制剂和甲醛交联的组合方法来促进翻译的停滞。该技术的进一步修改版本,选择性TCP-seq39,最近已被用于包括交联复合物的免疫纯化,扩大了TCP-seq应用的范围。甲醛交联的快速、高效和可逆特性使这些方法适用于研究瞬态mRNA:翻译复合相互作用,特别是在高度动态的翻译水平响应途径的背景下。
在这里,我们详细介绍了 体内 甲醛交联的过程,以达到综合转化复合物稳定和分离的目的。我们为酵母细胞和哺乳动物细胞提供了细微差别的单独方案(图1)。我们进一步概述了随后使用交联稳定材料的示例(图1),例如使用免疫印迹(免疫印迹)进行共纯化的蛋白质因子检测,免疫辅助纯化(或"免疫沉淀";IP)和富集含有特定目标因子的翻译复合物,电子显微镜和RNA测序。

图1:典型实验设置概述的示意图。 转化复合物 体内 甲醛稳定化的主要步骤被描述为流程图,并辅以有关关键必要仪器的信息。本文概述了交联材料的潜在下游应用,包括已成功使用但未在本方案中直接涵盖的示例,例如RNA的SPRI珠纯化,RNA测序和质谱分析。 请点击此处查看此图的放大版本。
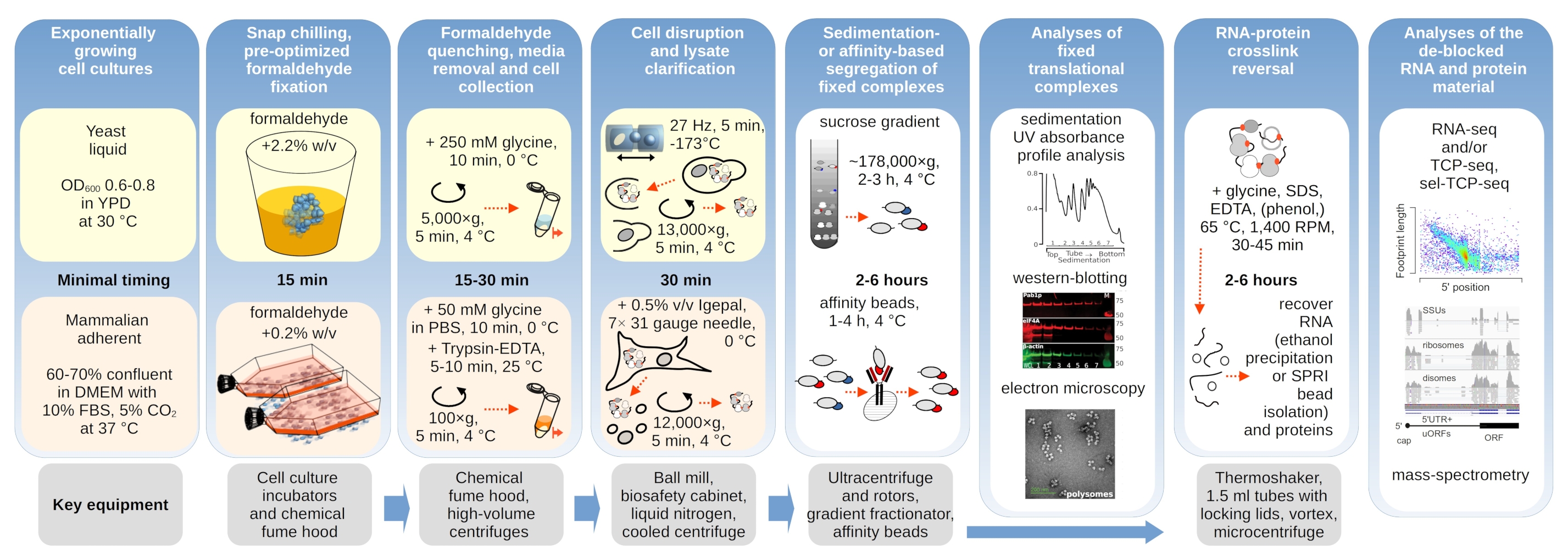{kind=link}
研究方案
1. 酵母细胞方案
- 酵母细胞培养和固定
注意:细胞固定和收获从10,38经过修改。- 在所需条件下(1%w / v酵母提取物,2%w / v,葡萄糖,40mg / L硫酸腺嘌呤(30°C用作示例)中设置1 L酵母细胞培养物(以野生型(WT)在600nm处起始光密度不超过0.05AU(OD 600)的轨道振荡器中培养1 L酵母细胞(以野生型(WT)为例)实验)。
- 设置具有兼容转子和离心机瓶的制备离心机,用于造粒酵母细胞的液体悬浮培养物。对于葡萄糖饥饿实验,一旦达到600nm处的光密度为0.6-0.8 AU(OD600),使用在30°C,5,000×g下短暂离心1分钟,就沉淀细胞。
注意:记录生长细胞的OD,如果指数生长阶段感兴趣,则让细胞生长,直到OD600 达到0.6-0.8 AU。 - 立即将沉淀重悬于含有不加入或低(0.25%w / v)的温(30°C)YP培养基中,并在轨道振荡器-培养箱中在30°C下再孵育培养物10分钟。
注:介质成分可能会影响后续的交联效率。该协议仅使用YPD进行测试。在进行饥饿实验时,坚持时机并最大限度地减少程序之间的延迟至关重要。 - 一旦细胞准备就绪,在通风橱内设置一个冰柜,里面有一个装有250克干净碎水冰的烧杯。确保在烟罩内可接触到 25 mL 条带和新购买的甲醇稳定 37% w/v 甲醛溶液。将1升培养物倒入装有25%w / v碎水冰的烧杯中。
注意:除非另有说明,否则在整个后续操作中将细胞保持在冰上,直到细胞被冷冻。 - 加入75 mL 37%w / v甲醛溶液至2.2%w / v的最终浓度,并强烈搅拌混合物直至冰融化。
- 冰融化后,设置一个计时器10分钟。
注意:坚持推荐的时间和温度方案,以获得可重复的固定结果。 - 孵育10分钟后,将培养物转移到预冷的离心机瓶中,并通过在4°C,5,000×g下离心5分钟来沉淀细胞。 当旋转开始时,将50 mL管预冷,并将新鲜制备的缓冲液A(含有甘氨酸以中和任何剩余的甲醛)放在冰上。
注:有关确切的缓冲液成分,请参阅提供的表格。 - 离心后,将离心管放在冰上,沉淀面与冰接触。将管子带入通风橱,并将上清液丢弃到甲醛废物容器中。
- 使用25 mL条带将所有管中的细胞沉淀重悬于20mL缓冲液A中,并转移到50mL管中。
注意:这种洗涤对于避免不可重复的交联至关重要,并且缓冲液的添加不得超过细胞收获后的20分钟。 - 用缓冲液A使体积达到40mL,并通过在4°C,5,000×g下离心5分钟收集洗涤的细胞。
- 弃去上清液并将细胞沉淀重悬于40mL缓冲液A1(不含甘氨酸的缓冲液A)中以除去任何甘氨酸污染。
- 通过在4°C,5,000×g下离心5分钟再次沉淀细胞。
- 用缓冲液A1重复洗涤一次。弃去上清液并将细胞沉淀放在冰上。用沉淀称量管(湿细胞质量应为每1L细胞培养物〜1g)。
- 酵母细胞破碎和细胞质基质收集
- 用液氮填充衬有铝箔的聚苯乙烯泡沫盒,深度约为3厘米。将 50 mL 试管直立放入包装盒中。
- 通过移液和涡旋10s将沉淀(约1g湿细胞质量)重悬于550μL缓冲液A2中。加入10μL的40 U / μL RNase抑制剂并再次涡旋10秒。
注意:处理液氮时,请穿戴适当的防护装备,如隔热手套。确保任何用于容纳液氮的容器不会泄漏,并且内部的管架不会漂浮或侧落。在通风良好的区域工作,以避免氧气耗尽。 - 使用1 mL移液器,将细胞悬浮液滴入含有液氮的50 mL管中。
注意:滴落必须缓慢而小心地进行,以避免液滴聚集。在引入新液滴之前,确保液滴冻结。 - 将带有冷冻细胞悬浮液滴的50mL管转移到室温,并等待液氮完全蒸发。用盖子密封管并将细胞沉淀储存在-80°C或立即进一步进行。
注意:在密封管子之前,请确保液氮完全蒸发。密封管中残留的液氮会导致危险的压力积聚。 - 为了准备下一步,在干冰上预冷却1.5 mL无核酸酶管和10 mL不锈钢研磨罐。
- 使用干净,无菌的刮刀将冷冻的细胞悬浮液滴转移到罐中。
注意:确保研磨罐密封牢固。 - 将研磨罐浸入液氮中1分钟,确保液相保持在液结下方。设置一个27 Hz的冷冻混合器研磨机进行搅拌1分钟。
注意:即使样品只需要一个研磨罐进行处理,也要始终将研磨罐与同一型号的另一个研磨罐进行平衡。 - 在混合器研磨机中以27 Hz搅拌密封的研磨罐1分钟。
- 像以前一样在液氮中重新冷却研磨罐,并在混合器研磨机中以27 Hz进一步摇动1分钟。
- 将罐子与1.5 mL无核酸酶管一起转移到装有干冰的冰盒中。使用小钢刮刀,将所得粉末状样品转移到〜100mg等分试样的管中,并将管储存在-80°C。
注意:建议每个实验使用约600mg的样品,包括多体沉降谱分析,将细胞质基质分离成翻译和非翻译的组分,以及在RNase消化后将翻译的组分进一步分离成SSU,核糖体和二组分。
- 从细胞质基质的非翻译部分分离固定(多)核糖体复合物
注:先前确定的程序10,38通常遵循基于其与(多)核糖体共沉降来富集翻译的RNA。这里介绍了一种更精细的方法来分离翻译和非翻译的细胞质基质组分,从而无需沉淀并随后重新溶解材料。- 使用冻融法43在薄壁超速离心管(5 mL,13 x 51 mm)中制备2.5 mL线性10%-20%w / v蔗糖梯度,使用缓冲液B。
注意:冻融法是通过连续添加和冷冻缓冲蔗糖层来执行的,其浓度彼此线性递减。有关详细信息,请参阅补充表 1。 - 为了形成不连续的50%w / v蔗糖垫,在线性梯度解冻和稳定后,使用1 mL注射器将缓冲液B中的0.5mL 50%蔗糖缓慢分配到管的底部,该注射器连接到19 G x 1.5"针头或类似/合适尺寸的玻璃毛细管。在分配之前,小心而缓慢地将针尖或毛细管从预先形成的蔗糖梯度的顶部驱动到底部,避免任何干扰,直到它到达管底部。
注:有关缓冲液B制备的说明,请参见补充表1。 - 通过去除顶部部分或在缓冲液B中分层更多的10 w / v蔗糖并保持冰冷或4°C,仔细平衡梯度。
注意:需要底部50%蔗糖层的不连续梯度来收集沉降率较高的材料,而不会将其沉淀在管壁上。 - 在室温下解冻〜100mg冷冻细胞粉末样品并立即置于冰上。通过移液在150μL缓冲液A2中混合,将RNase抑制剂加入1 U / μL并通过涡旋混合(避免过度发泡并与气相混合)混合10秒。
注意:除非另有说明,否则在将材料放在冰上的同时继续所有操作。 - 通过将管在4°C,13,000×g下离心5分钟来沉淀细胞碎片,并在新的1.5mL低蛋白结合管中回收澄清的上清液(〜150μL)。
- 将所得的澄清混合物加载到步骤1.3.3的不连续蔗糖梯度管上,并仔细平衡它们。
- 在4°C下将管子在中等体积的摆动桶转子中超速离心,平均重力为287,980 x g(k 因子49)1小时30分钟。
注意:这些条件已经过预先优化(使用超速离心梯度紫外吸光度痕量分析),以将游离(非(多)核糖体)SSU和LSU(大核糖体亚基)保留在梯度的顶部(10%-20%蔗糖)部分,同时浓缩底部(50%)蔗糖垫中的(多)核糖体部分而不造粒材料。 - 使用配备19 G x 1.5"针头的新型无菌1 mL注射器收集翻译的细胞质基质组分。将 5 mL 梯度放在稳定的支架上,确保试管底部可见。
- 从试管的顶部,将针头直接插入梯度的底部(不刺穿试管),轻轻地,不产生任何气泡,取出含有翻译的RNA池的底部溶液的0.5mL。
注意:确保在冷室中执行此步骤,并牢固地握住管子。建议在单次上冲程运动中绘制整个0.5 mL,以避免梯度的干扰。 - 通过超速离心运行时蔗糖梯度的吸光度读数,确认(多)核糖体的存在以及所得混合物中SSU,LSU和较轻馏分的耗尽。
- 使用具有10 kDa截止再生纤维素膜的微量浓缩装置中的超滤将上一步收集的翻译RNA池浓缩至100μL。
注意:用0.5 mL缓冲液1预洗微浓缩装置的膜(见图2a),并使用制造商推荐的旋转条件(g)。 - 用缓冲液1进一步稀释上一步中的材料五次(加入400μL),并浓缩回200μL,以允许更小的体积以及部分去除蔗糖。
注意:建议将所得混合物在-80°C下储存长达6个月,并用作"总翻译RNA"RNA-seq文库构建的输入材料,或TCP-seq文库构建的RNase消化步骤。"非翻译"细胞质基质组分可以使用类似的程序从梯度顶部回收并储存在-80°C。
- 使用冻融法43在薄壁超速离心管(5 mL,13 x 51 mm)中制备2.5 mL线性10%-20%w / v蔗糖梯度,使用缓冲液B。
- RNase消化固定(多)核糖体复合物并将消化物质分离成小核糖体亚基(SSU),单核糖体(核糖体,RS)和双核糖体(二组,DS)级分
注意:该过程通常遵循前面描述的方法10,38但是采用了改进的梯度类型,分离时间,加速度和RNase消化条件,以在所有三个分离的馏分上实现最佳分辨率。- 使用步骤1.3.1中描述的冻融方法43,用缓冲液1在13 mL薄壁聚丙烯管中制备精心平衡的12.5 mL线性10%-40%w / v蔗糖梯度,14 x 89 mm,并注意其中。
- 在室温下解冻并立即将样品转移到冰上或取步骤1.3.12中浓缩和蔗糖耗尽的翻译细胞质组分。
注意:除非另有说明,否则在冰上继续所有程序。 - 通过在23°C下混合4.5U 大肠杆菌 RNase I每1 OD260 单位的馏分30分钟来消化翻译的细胞质基质组分。 通过移液能够使RNase I失活的RNase抑制剂立即加入并混合至0.25 U / μL到混合物中,以使RNase I失活。
注意:使用能够抑制RNase I的RNase抑制剂,通过使用AU260 =(260nm处的吸光度标准化为相当于1cm光程x以μL为单位的裂解物体积)/ 1,000来获得AU260。 - 立即将样品转移到冰上。
注意:坚持推荐的消化条件并仔细测量添加的RNase I的量至关重要。这里提到的RNase I单元被定义为在37°C下30分钟内从小鼠肝脏RNA产生1μg酸溶性物质所需的酶量。如果酶储备物过于浓缩,建议用缓冲液1稀释,以避免移液非常小体积的溶液。 - 将反应混合物加载到步骤1.4.1的10%-40%w / v蔗糖梯度上。
注:使用最终体积,每个梯度为150-300 μL。每次纯化至少需要两个梯度。使用不同输入体积的材料(较低的AU 260,10-11 AU260,用于DS,相对较高的AU260,13-14AU260,用于SSU或RS)以实现最佳分离。 - 在中等体积的摆动桶形转子中,在4°C下超速离心管,平均重力为178,305 x g(k 因子143.9)3小时30分钟。
注意:如果需要备用平衡管,请使用含样品管平衡其质量和质量分布。使用备用蔗糖梯度覆盖,其缓冲液量相当于样品覆盖量,而不是蔗糖浓度均匀的试管。 - 在超速离心离心旋转完成前至少30分钟设置梯度分馏器装置,包括将0.2μm过滤的重追逐溶液(例如,此处使用的去离子水中含有60%蔗糖)填充到排量泵中。
注意:建议使用去离子水去除分馏器的管路和管道,然后在运行前后在去离子水中加入1%-2%SDS溶液,去离子水,最后在去离子水溶液中加入80%乙醇。 - 调整吸光度读数基线,首先用去离子水填充系统并根据制造商的建议将光学器件归零,然后使用备用的卸载的14 x 89 mm蔗糖梯度补偿基线,该缓冲液由与样品管相同的缓冲液(例如,缓冲液1)制成。
注:使用与样品读数相同的位移速度进行调整,例如1.5 mL/min。 - 通过精确计算溶液首先进入检测器的光路到首先出现在馏分收集器输出端之间的时间,测量位移系统死体积。
注意:推荐速度为 1.5 mL/min 时,可在室温下进行分馏。建议立即将收集到的馏分转移到冰上。 - 使用 254 nm 的活吸光度读数、1.5 mL/min 置换速度以及基于样品的预期沉降位置和吸光度曲线的在线馏分检测进行分馏。使用与之前测量的死体积相对应的时间延迟的收集管切换。
- 分离与SSU,RS和DS复合物的位置和迁移率相对应的级分,并将它们收集到新的低蛋白结合1.5mL微量离心管中;立即将分离的馏分转移到冰上,如果不立即进一步处理,则将其冷冻。
注意:建议立即将收集的馏分在干冰或液氮中闪冻,并在-80°C或以下的温度下储存长达6个月。
- 核糖体复合物的脱交联和RNA的分离以构建RNA-seq文库
- 为了去嵌段/逆转交联并将RNA从相关蛋白质中分离出来,将大约一半的整个蔗糖梯度级分转移到带有盖子安全/锁定装置的新型低核酸结合无核酸酶聚丙烯1.5mL微量离心管(每管350μL)中。
- 用40μL100%停止溶液(10%SDS w / v和100mM EDTA),4μL1 M Tris-HCl pH 2在25°C(至10mM)下补充混合物,1.6μL2.5M甘氨酸(至10mM)和去离子无核酸酶水,以获得最终体积为400μL。
- 通过移液混合试管的内容物,并在室温下转移试管。
- 向每个管中加入等体积的酸性苯酚:氯仿:异戊醇125:24:1(pH 4.0-5.0)混合物。使用设置为最大速度的涡旋混合器用力摇动混合物2分钟。
注意:苯酚和氯仿具有腐蚀性和毒性。避免与液体发生物理接触,并在通风良好的区域或通风橱下工作。使用苯酚或氯仿时,请务必使用手套、实验服和护目镜或面罩。 - 将管子放入热塑轮中,在65°C,1,400 rpm下连续摇动30分钟。
- 通过在室温下以12,000×g离心混合物10分钟来促进相聚集。
- 收集上层水相并将其转移到新鲜的低核酸结合1.5mL管中。
注:为避免交叉污染,请勿尝试完全回收水相。合理的回收量为300-350μL。 - 用0.1体积的3M乙酸钠(25°C时pH 5),20μg糖原(使用5μg/ μL储备)和2.5体积的无水乙醇补充收集的水相。通过涡旋管1分钟小心地混合溶液。
- 通过将样品在-20°C下孵育至少2小时(推荐过夜)来沉淀RNA。
- 将管加热到室温并通过涡旋混合。
注意:试管的预热和随后在室温下离心(无需强制冷却)有助于减少盐和苯酚的共沉淀和残留。如果按照所述进行并使用足够纯的乙醇,这些条件不应导致材料损失或RNA收集效率低下。 - 通过在室温下以12,000×g离心管30分钟来沉淀RNA沉淀。
- 弃去上清液并用80%v / v乙醇洗涤沉淀两次,每次通过在室温下以12,000×g离心10分钟收集。
- 通过打开管盖并将打开的管放入设置为45°C的干块加热器中10分钟来干燥RNA沉淀。将所得的干燥沉淀溶解在20μL的1x HE缓冲液中。
- 使用紫外吸光度光谱测量估计所得的RNA浓度。
注意:RNA片段长度和总量可以使用变性凝胶电泳进一步评估,例如在基于荧光的自动毛细管凝胶电泳装置中。
- 通过标记的eIF和选择性SSU富集的蛋白质印迹分析对SSU进行选择性共免疫纯化
注意:使用步骤1.4.11中约15 AU(260nm)的消化和沉降分离的SSU馏分,使用磁性IgG磁珠进行亲和纯化。保存约5%的SSU分数作为输入控制(输入分数,I)。eIF4A标记 (TIF1-TAP;使用串联亲和纯化标签)酵母菌株,这也使得通过使用抗TAP抗体探测TAP标签来检测eIF4A成为可能。- 将100μL磁性IgG微球悬浮液(每15 AU(260nm)裂解物或馏分使用1mg微球)转移到新的低蛋白结合1.5mL管中;使用磁性架收集珠子并吸出它们。
- 使用1 mL缓冲液1洗涤磁珠两次,方法是使用移液和收集的顺序重悬,并使用磁性支架收集。
- 洗涤后,收集并倾倒珠子,同时将它们放在磁性架上。
- 将SSU馏分加入洗涤的珠中,并将混合物在4°C下旋转,在约20rpm的环形混合器中孵育4小时。
- 在4°C下使用磁性架收集磁珠并保存上清液(流通分数,FT)。
- 用补充有4mM DTT的缓冲液1在4°C下洗涤珠子两次,每次在循环混合器中旋转10分钟,并在磁性架上收集和倾倒珠子。保存洗涤液(W1 和 W2 分数)。
- 对于分析应用(如蛋白质印迹),通过加入LDS(十二烷基硫酸锂)聚丙烯酰胺凝胶电泳(PAGE)样品缓冲液,pH值为8.5至1x,DTT至2mM,在变性和还原条件下洗脱结合材料。
- 在热块中将混合物在95°C下加热5分钟以完成洗脱。
- 使用磁架收集微珠,并在新鲜的低蛋白结合1.5mL微量离心管中回收变性洗脱液(E级分)。
- 使用上一步中的E级分立即运行变性十二烷基硫酸钠(SDS)PAGE,或将E级分储存在-20°C。
注意:对于任何后续应用,对于TAP标签富集的转化复合物的制备性收集,请使用采用烟草蚀刻病毒(TEV)蛋白酶的替代洗脱方法。有关更多详细信息,请参阅补充表 1。 - 为了浓缩稀释的FT,W1和W2馏分,通过加入3倍体积的冰冷丙酮来沉淀其物质。将样品 - 丙酮混合物在-20°C孵育3小时。
- 通过在4°C下以13,000×g离心管10分钟来沉淀沉淀。
- 弃去上清液,并在室温下在打开的管中风干沉淀30分钟。
- 将沉淀溶解在7μL的1x LDS上样缓冲液中,并补充2mM DTT。将样品在设置为95°C的热块中加热5分钟。
- 将所有I,FT,W1,W2和E样品加载到4%-12%w / v丙烯酰胺梯度,双-Tris聚丙烯酰胺变性凝胶上。使用1x MES SDS(2-[N-mopholino]乙烷磺酸,十二烷基硫酸钠)在80 V下电泳缓冲液运行凝胶,直到蛋白质标记物(10-250 kDa)解析良好并且铅染料到达凝胶底部。
注意:建议在凝胶上加载WCL(全细胞裂解物)(2-10μg)的连续稀释液作为对照。可能需要多次尝试才能在馏分材料上实现凝胶的可比负载。 - 按照蛋白质印迹设备制造商的建议,通过湿转方法在100 V下在冷室中将凝胶的蛋白质含量转移到聚偏二氟乙烯(PVDF)膜上1小时。
- 在恒定振荡下,使用适当的封闭缓冲液(基于磷酸盐缓冲液)在室温下阻断膜1小时。
- 按照制造商的抗体稀释说明,在冷室中用封闭缓冲液(PBS)稀释的抗体(1:1,000稀释)在膜上孵育过夜,用抗TAP抗体探测膜,用于检测标记的eIF4A蛋白,抗Pab1p抗体或抗β肌动蛋白抗体(或任何其他理想的靶标)。
注意:1:1,000抗体稀释液是一个很好的起点。 - 用1x磷酸盐缓冲盐水洗涤膜三次,每次0.2%v / v吐温20(PBST)10分钟。
- 按照制造商的说明,通过在室温下在环混合体中孵育1小时,用荧光标记的二抗探查膜。
注意:1:20,000抗体稀释是一个很好的起点。 - 用1x PBST洗涤膜三次,每次10分钟。用去离子水短暂冲洗膜,然后用无水甲醇冲洗。根据制造商的说明,在荧光成像系统中干燥并可视化膜。
注意:其他蛋白质的染色可以通过使用具有匹配不同荧光通道的染料的二抗来实现(例如,在eIF4A-TAP 与 此处使用的β-肌动蛋白对中),通过顺序染色或剥离并染色相同的膜或从装载有重复级分图案的凝胶中切割膜,并用各自的抗体单独探测每块(如此处使用的Pab1p示例所示)。
2. 哺乳动物细胞方案
- 哺乳动物细胞培养和固定
- 在2个T-175烧瓶中,在Dulbecco的改性鹰培养基和37°C和5%v / v二氧化碳下将HEK293细胞培养至60%-70%汇合,10%v / v胎牛血清。
注:将55 mL商用FBS加入500 mL商用DMEM中,含有L-谷氨酰胺,酚红和碳酸氢钠,但不含HEPES或丙酮酸钠,从而制成完整的培养基。在70%汇合时,每个T-175烧瓶的细胞计数应在1.7-2.0 x 107的范围内。 - 在所需固定时间前至少3小时,用精确30 mL预热的完整培养基替换T-175烧瓶的培养基,并更换细胞培养箱中的烧瓶。
注意:确保将新鲜培养基移液到烧瓶的另一侧,以达到细胞单层,以避免细胞脱离。尝试尽快进行介质交换,引入最小的气体和温度平衡干扰。 - 更换细胞培养基后,准备固定所需的缓冲液和化学品。通过将10.2毫升2.5米甘氨酸储备液加入500毫升DPBS瓶中并混合,用50 mM甘氨酸制备Dulbecco的磷酸盐缓冲盐水(DPBS)。
- 按照步骤2.1.1准备一瓶补充有10%FBS的DMEM,用于非无菌条件和100毫升0.25%胰蛋白酶-EDTA等分试样。采购另一瓶用氯化钙(CaCl2)和氯化镁(MgCl2)预先配制的商用DPBS。
注意:溶液可以在4°C下储存长达2周。 - 准备一个装满碎水冰的冰盒,使T-175烧瓶可以均匀地放在顶部,并与准备好的缓冲液一起保持在通风橱中,也可以放在冰上。
注意:由于翻译对任何环境变化的快速反应,必须最小化从培养箱中取出细胞烧瓶和加入甲醛溶液之间的所有时间。 - 为了使细胞快速冷却,请从培养箱中取出T-175烧瓶,并将其牢固地压在冰上,以确保最大的表面接触。在化学通风橱内,将烧瓶倾斜到其侧面,以便培养基聚集在与细胞相反的一侧。将168μL37%w / v甲醛直接移入混合培养基中(最终浓度为0.2%w / v)。通过轻轻地来回摇动烧瓶,关闭烧瓶并将其重新定位在冰上,确保其水平并且细胞均匀覆盖,立即混合。
注意:甲醛是一种有害物质,具有潜在的长期不良反应,对呼吸系统和皮肤也有刺激性。它只能在合适的化学通风橱中处理。甲醛容器在通风橱外时必须始终密封。
注意:确保将甲醛直接添加到细胞培养基中,而不是添加到烧瓶壁中。步骤2.1.6应少于1分钟。 - 将烧瓶在冰上再孵育10分钟。将培养基倒入适当的废物容器中,通过与细胞相对的烧瓶侧。
- 使用条带,在30 mL Dulbecco的磷酸盐缓冲盐水中移液,不含钙和镁离子,另外含有50 mM甘氨酸,轻轻地放在与细胞相反的一侧。通过摇动烧瓶混合;将烧瓶放回水平位置,并在冰上再孵育10分钟。
- 通过与细胞相反的烧瓶侧倒出溶液,轻轻加入7mL标准0.25%w / v胰蛋白酶-EDTA溶液以分离和重悬细胞。将烧瓶在室温下孵育5-10分钟。
注意:确保胰蛋白酶-EDTA溶液均匀覆盖所有细胞。使用周期性的轻柔倾斜和摇摆来促进细胞脱离。 - 垂直放置烧瓶,并使用条带通过轻轻地从烧瓶壁上清洗任何剩余的细胞来收集分离的细胞。将悬浮液转移到冰上的50 mL管中。
注意:固定细胞会变得更加脆弱;不要用力移液或超过将细胞从烧瓶壁上分离所需的量程。 - 立即用20mL完整培养基(具有10%FBS的非无菌冰冷培养基)补充收集的细胞悬浮液,并通过轻轻翻转管子混合。
注意:加入完整的细胞培养基(包括10%FBS)以中和胰蛋白酶,防止进一步损伤细胞膜和细胞崩解。 - 通过将管以100×g离心5分钟和4°C来沉淀细胞。 细胞沉淀必须清晰可见。
- 倒出培养基,将细胞沉淀轻轻重悬于10 mL冰冷的DPBS中,其中含有Ca2 +,Mg2 +,不含甘氨酸。
- 重复步骤 2.1.12。
- 倒出洗涤缓冲液,将细胞沉淀轻轻重悬于800μL冰冷的DPBS中,其中含有Ca2 +,Mg2 +,不含甘氨酸,在冰上。将重悬的细胞转移到新的低蛋白结合1.5mL微量离心管中。
- 将管在100×g和4°C下离心3分钟。 使用1 mL移液器小心地丢弃上清液。在此阶段,细胞沉淀可以在-80°C下冷冻或进入细胞裂解步骤。
注意:冷冻细胞沉淀可以在-80°C下储存长达1年。我们发现细胞沉淀冷冻有助于随后的裂解,即使没有计划长期储存,也建议冷冻。
- 在2个T-175烧瓶中,在Dulbecco的改性鹰培养基和37°C和5%v / v二氧化碳下将HEK293细胞培养至60%-70%汇合,10%v / v胎牛血清。
- 哺乳动物细胞破碎和细胞质基质收集
- 在生物安全柜中,加入300μL基于非离子,非变性洗涤剂的裂解缓冲液和7μL的40 U / μL RNase抑制剂。使用 1 mL 吸头移液充分混合。
- 小心地将25 G针头连接到1-3 mL注射器上,并使用至少7次缓慢向上进气和快速向下排气冲程大力移取混合物。
- 将注射器和针头丢弃到尖锐的料仓中,并使用装有31 G针头的0.3 mL注射器重复该过程。
- 将注射器和针头丢弃到尖锐的垃圾箱中。将管在4°C,12,000×g下离心5分钟以沉淀细胞碎片。
- 将上清液转移到新的低蛋白结合1.5mL微量离心管中。将细胞碎片(用于控制目的)和所得澄清的细胞裂解物在-80°C下储存。
注:当两个T-175烧瓶组合并遵循推荐体积时,裂解物的光密度范围在25-30 AU260 之间。裂解物和细胞碎片可以在-80°C下储存长达1年。
- 从细胞质基质的非翻译部分分离固定(多)核糖体复合物
- 在13 mL薄壁聚丙烯管中制备线性15%-45%w / v蔗糖梯度,14×89 mm,使用冻融方法,通常如酵母方案的步骤1.3.1中所述,但使用缓冲液2(图2a)。
注意:在分馏前一天晚上,在4°C的冷室中解冻梯度过夜。 - 将150-250(最大300)μL细胞裂解物从前一步2.2.5加载到平衡梯度上。将剩余的裂解物储存在-80°C并用于控制目的。
注:这里提供了一个基于沉降的分离成多体、核糖体和"游离"SSU级分的例子。有关替代方法,请参阅提供的补充表1。 - 在中等体积的摆动桶形转子中,在4°C下超速离心管,平均重力178,305 x g(k 因子143.9)1小时45分钟。
- 在离心完成前30分钟,设置梯度分馏器并基线,如酵母方案步骤1.4.7-1.4.9中所述。
- 分馏梯度通常如酵母协议步骤1.4.10-1.4.11中所述。
注意:此步骤将分离多体、核糖体和"游离"SSU 级分。多体组分可用于多体分析实验。 - 立即将收集的馏分转移到冰上,如果没有进一步处理,请在-80°C下储存长达6个月。
注意:如果馏分收集管更换与在线馏分鉴定和分离同步,我们建议使用最多800 μL级分(在1.5 mL/min下,每馏分的收集时间为32秒)。如果在不使用在线吸光度读数的情况下进行分馏,则建议使用250-500μL级分(每馏分10-20秒,1.5 mL / min)。分离后,这些级分可用于免疫纯化、电子显微镜检查、变性 PAGE 和立即进行蛋白质印迹,或进行交联逆转以进行后续的 RNA 和/或蛋白质组学分析。
- 在13 mL薄壁聚丙烯管中制备线性15%-45%w / v蔗糖梯度,14×89 mm,使用冻融方法,通常如酵母方案的步骤1.3.1中所述,但使用缓冲液2(图2a)。
结果
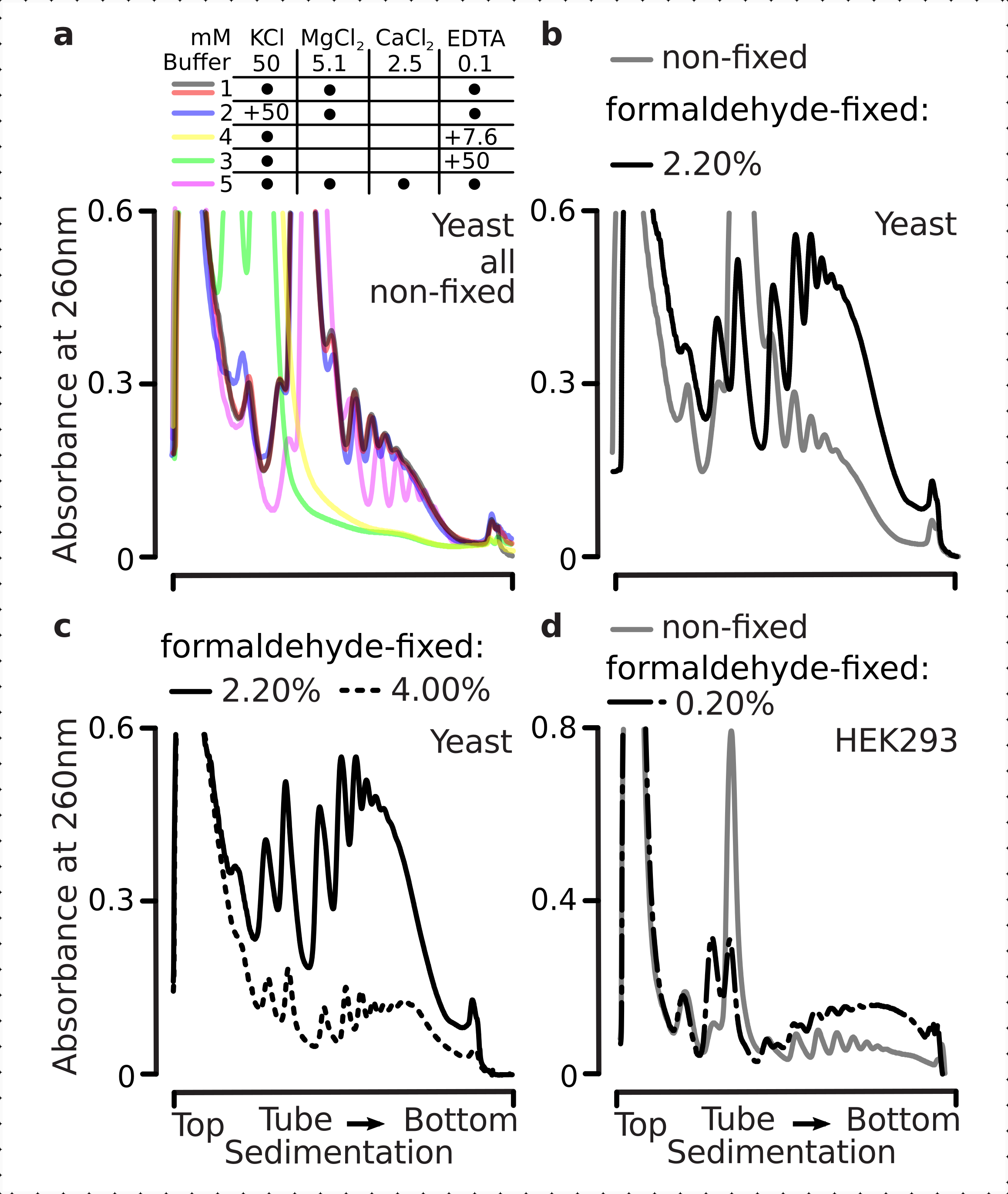
转化复合物对缓冲液的离子组成敏感,这在评估沉降特性的超速离心过程中尤为重要。因此,我们使用从研磨的非固定酵母材料中提取的澄清裂解物测试了几种沉淀缓冲液,以选择最适合解析转化复合物的条件,并在梯度上分离核糖体亚基(SSU,LSU),单体(RS)和多体。所有缓冲液均基于含有25 mM HEPES-KOH pH 7.6和2 mM DTT的核心成分。KCl、MgCl2、CaCl2和EDTA的浓度在缓冲液中进一步修饰(图2a),并且这些组分在梯度上样前被添加到裂解物中,并在梯度浇注前被添加到蔗糖梯度缓冲液中。
在缓冲液1和2中,获得解析良好的转化复合物。缓冲液1导致小核糖体亚基(SSU)的分离更好(图2a)。省略MgCl2 和添加EDTA(缓冲液3,4)导致大多数多微体的高沉降性能丧失,并且可能部分分解(图2a)。虽然添加2.5mM CaCl2 导致多杂体峰更加均匀,但与缓冲液1和2相比,在这种情况下,多粒体物质的总量减少(图2a)。因此,我们选择缓冲区 1 作为首选的工作缓冲区。

图2:用于转化复合物提取的缓冲条件和固定稳定性的评估。 所示是在260nm处收集的以10%-40%w / v蔗糖梯度分离的总酵母细胞裂解物的紫外吸光度曲线。(a)单盐和二价盐以及镁离子封存对从非固定酵母细胞中提取的物质沉淀的影响。红线和灰线表示典型的重复。(b,c)比较来自非固定(灰线),2.2%(黑线)和4.4%(黑虚线)的裂解物,w / v的甲醛固定酵母细胞。(d) 与来自相同非固定细胞的材料(灰线)相比,通过优化的 0.2% w/v 的 HEK 293T 细胞甲醛固定(黑色虚线和虚线)来稳定多粒体。 请点击此处查看此图的放大版本。
{kind=link}
接下来,我们检查了通过用不同甲醛浓度固定来稳定多体的效果。使用其他相同的细胞材料,缓冲液,细胞处理和定时方法,我们比较了从非固定细胞中提取的材料以及用2.2%和4%w / v的甲醛固定的细胞(图2b,c)。我们发现,2.2%w / v的甲醛更适合固定,因为它很好地保留了多体,可以通过多体与单体的比例来判断(图2b),与4%w / v的甲醛相比,它没有降低核糖体材料的总产量,甲醛表现出明显的过度固定迹象(图2c)。
对于来自哺乳动物细胞的材料,由于基于洗涤剂的提取所需的裂解缓冲液与细胞体积比较大,因此使用缓冲液2(图2a)。这在蔗糖梯度沉降时产生了分辨良好的转化复合物(图2d)。值得注意的是,甲醛浓度低得多,为0.2%w / v,因为较高浓度导致大量的多粒体和核糖体物质损失(数据未显示)。与酵母细胞获得的结果相似,交联稳定的材料显示出更好的多微体保存和更高的多体与单体比率(图2d)。
接下来,我们测试了所选的甲醛固定条件是否足够有效,以稳定交联结果导致多体组分内活性翻译的mRNA,并且多粒体产量的提高不仅仅是抑制酶功能和翻译伸长率的进展的结果。我们使用EDTA和高一价盐(KCl)来破坏多微体和核糖体的稳定性。将这些试剂加入到澄清的酵母细胞裂解物中,并分别包含在缓冲液1组合物顶部的所有后续缓冲液和蔗糖梯度中。
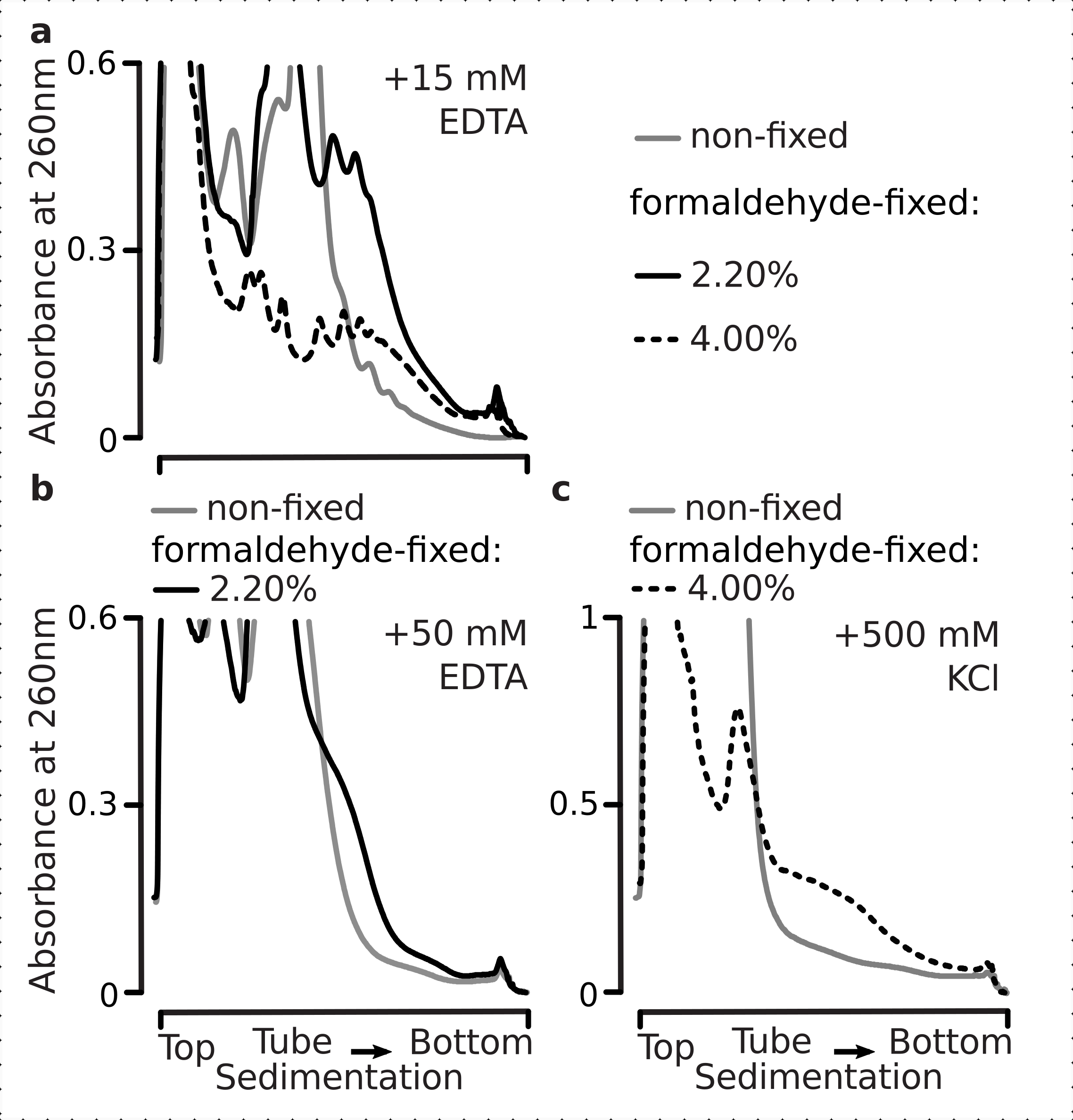
事实上,15 mM EDTA对来自固定细胞的多体组分表现出较小的不稳定效应(图3a),证实了交联的复合物更健壮。EDTA的不稳定作用可以通过增加甲醛的浓度来克服,因为来自4%w / v的甲醛固定细胞的材料抵抗展开更好(图3a)。然而,将EDTA浓度增加到50 mM导致大多数平移配合物在固定和非固定条件下的稳定性,这可以从物质沉降较慢和没有形状良好的峰中推断出来(图3b)。这可以通过结构的部分展开和致密性的整体丧失来解释,而不是通过多体组分从mRNA中完全解离来解释。即使在这种情况下,交联材料也显示出更快的沉降速度(图3b)。

图3:体内酵母甲醛固定对多微体稳定性的影响。所有实验都使用了缓冲液1(见文本和图2a)。数据类型和绘图如图2图例中所述。(a) 比较向细胞裂解物和后续缓冲液中加入15 mM EDTA对非固定(灰线)、2.2%(黑线)和4%(黑虚线)的多微体的稳定性,w/v与甲醛固定细胞。(b) 与 (a) 相同,但用于添加 50 mM EDTA,不包括 4% w/v 的甲醛固定细胞。(c)与(a)相同,但用于添加500 mM KCl,不包括2.2%w / v的甲醛固定细胞。请点击此处查看此图的放大版本。
{kind=link}
与EDTA效应类似,在500 mM KCl下,我们发现4%w / v的甲醛固定显着改善了稳定性(图3c)。在这种情况下,致密性的明显丧失也可以解释为核糖体复合物成分的部分脱离,而不是它们与RNA完全解离。总体而言,来自甲醛固定细胞的多体表现出对展开和结构不稳定的更高抵抗力,这与在这些复合物中形成额外的共价键一致。
在刺激生长条件下,mRNA可以迅速启动,导致多个核糖体在相同的mRNA分子上积累,形成称为多核糖体或多体的结构。多粒体可以通过蔗糖梯度的超速离心分离,在那里它们根据其顺序(mRNA上同时附着的核糖体的数量)沉积。当翻译被抑制时,核糖体无法很快进行另一轮翻译,导致多体的(部分)"反汇编",这表现为向低阶多体的模态转移和单体4,26的积累。
葡萄糖匮乏可以提供可以在多体顺序分布水平上可视化的转化反应模型。葡萄糖消耗引起对酵母1,3,40的最显着和最快速的翻译抑制作用之一。先前的研究证明,在葡萄糖消耗1分钟内,多小体的丢失,单体的积累和翻译起始的抑制可能发生4。在葡萄糖重新补充的5分钟内,翻译迅速恢复,多体3,4明显增加。还观察到,当细胞暴露于含有0.5%(w / v)或更低葡萄糖的培养基时,翻译受到抑制,并且在0.6%(w / v)或更高的葡萄糖水平中看不到效果。
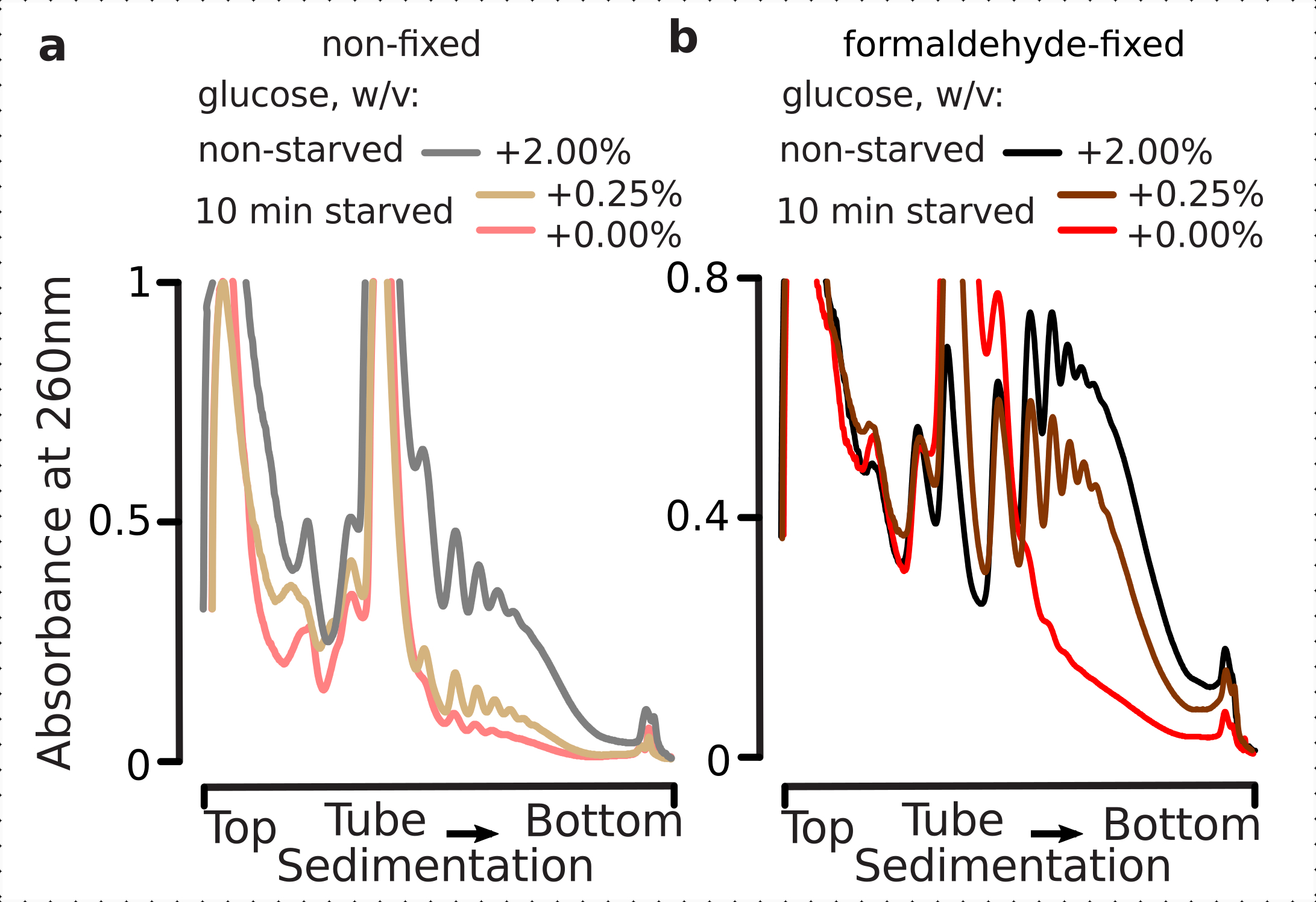
因此,我们希望确定我们的固定条件是否适合于保存葡萄糖应激反应动力学内的翻译差异,这可以通过多体与单体比率来评估。我们将来自在高葡萄糖(2.00%w / v添加)上以中等指数阶段生长的细胞的材料与那些转移到未添加或低添加(分别为0.00%或0.25%w / v)葡萄糖的培养基中10分钟的材料进行了比较。在对照组(非饥饿;用含有2%w/v添加葡萄糖的相同标准培养基快速置换,然后孵育10分钟并固定)和饥饿10分钟(用相同培养基快速置换0.25 w/v或未添加葡萄糖, 然后孵育10分钟并固定)细胞。
与早期的发现一致,我们观察到酵母细胞在葡萄糖饥饿应激时严重抑制翻译(图4a)。无论是无添加的还是低葡萄糖的条件都诱导了多微体的分解,在低添加葡萄糖的情况下,略微但明显地保留了更多的多聚体。因此,酵母葡萄糖去除反应可能不是全开或全关型,而是逐渐调整。肯定了对甲醛交联的稳定作用的期望,来自固定细胞的多体材料已经证明了饥饿细胞和非饥饿细胞之间的更高区别,可以说保留了更高的响应动态范围(图4b)。有趣的是,在来自固定细胞的材料的情况下,与非固定细胞相比,低添加的葡萄糖浓度导致特定的多粒体丰度与未添加的葡萄糖条件相比,其与未添加的葡萄糖条件的区别要好得多(图4a)。这有力地表明了甲醛固定方法在保存和捕获高度动态过程平衡中相对微小和瞬态差异方面的适用性,例如在转化反应期间。

图4:捕获葡萄糖饥饿时酵母翻译的快速变化。 所有实验都使用了缓冲液1(见文本和 图2a)。数据类型和绘图如图 2 图例中所述。(a)从非饥饿(灰线),限制性葡萄糖饥饿(0.25%w / v加入葡萄糖10分钟;棕色线)和葡萄糖消耗(不添加葡萄糖10分钟;红线)非固定酵母细胞获得的细胞裂解物。(b) 与 (a) 相同,但适用于 2.2% w/v 甲醛固定细胞。 请点击此处查看此图的放大版本。
{kind=link}
使用蔗糖梯度沉降("多体分析")监测与主动翻译mRNA相关的核糖体的翻译状态是一种广泛应用的技术26,27,28。结合定量微阵列分析和最近的高通量测序28、44,多体分析提供了有关核糖体相关mRNA转录组的信息。在蛋白质生物合成研究领域,传统上认为多体体的存在表明积极参与相应mRNA的翻译。进一步的结论通常(但并非总是)是合理的,即在给定长度的mRNA上存在的核糖体越多(多体的阶数越高),mRNA参与翻译的次数就越积极。因此,从分离活性翻译RNA的角度来看,从其余材料中分离多体部分可能是有用的。在足迹分析方法中,特别是TCP-seq 10,38,39,它从扫描,开始和停止密码子复合物中产生释放的SSU的单独群体,去除不与完整单体或多体共沉积的核糖体亚基可能具有额外的洞察力。
因此,我们采用了将"非翻译"mRNPs(例如游离SSU(与单个SSU结合的mRNA或没有附着的mRNA的SSU)从mRNA的"主动翻译"池中分离出来。为了实现这一点,我们假设参与与一种(单核)或几种核糖体(多体)相互作用的mRNA可以被主动翻译。这些复合物可以通过其较高的沉淀系数与其他复合物分开。我们还建议将"主动翻译"的mRNA池分离到蔗糖垫(50%w / v蔗糖)中,而不是直接在管壁上沉淀材料。将快速沉淀的配合物离心到垫子中,使我们能够使用吸光度剖面读数来监测分离,并与造粒和再增溶相比,实现溶解,非聚集和非变性材料的更高输出10,38。
总体而言,为了纯化单个SSU,核糖体,二组和可能紧凑堆积的高阶多微体,固定澄清的裂解物进行了两阶段超速离心过程(图5)。在第一个蔗糖梯度中,超速离心导致梯度顶部(蔗糖的10%-20%w / v)部分分离的游离SSU和LSU,而交联的翻译池包括与一个完全核糖体相关的多微体和mRNA集中在梯度的底部(蔗糖的50%w / v)(图5a).然后将含有翻译的mRNA池的蔗糖层的底部50%w / v浓缩其RNA,并用RNase I消化其RNA,然后进行第二次蔗糖梯度超速离心以获得单独的SSU,LSU,RS,RNase抗性二组(DS)和小分数的高阶核酸酶抗性多微体(图5b)。用乙酸铀酰进行阴性染色和用透射电子显微镜成像证实了在每个沉降阶段分离的复合物的身份(图5)。

图5:从未翻译的RNA中分离出总翻译的RNA级分(a,c)示意图(左)和相应的代表性结果(右;数据类型和绘图如图2图例中所述)(a)第一次不连续蔗糖梯度分离非翻译的细胞质基质组分,包括游离SSU和翻译的mRNA库,通过与核糖体和多微体共沉降鉴定, 和(c)通过受控的RNase I消化和超速离心分离从翻译的mRNA池中释放的单个核糖体复合物,通过第二个线性蔗糖梯度进入SSU,LSU,核糖体(RS)和核酸酶抗性二体(DS)级分。包括高(15 AU260)和低(8 AU 260)未饥饿消化物质的量,以证明当对次要馏分感兴趣时增加超速离心负荷的可能性。高阶核酸酶抗性多体也可以被鉴定(例如,所提供的例子中的三体)。(b,d)来自(a,c)的醋酸铀酰对比部分的代表性TEM图像,如标记。请点击此处查看此图的放大版本。
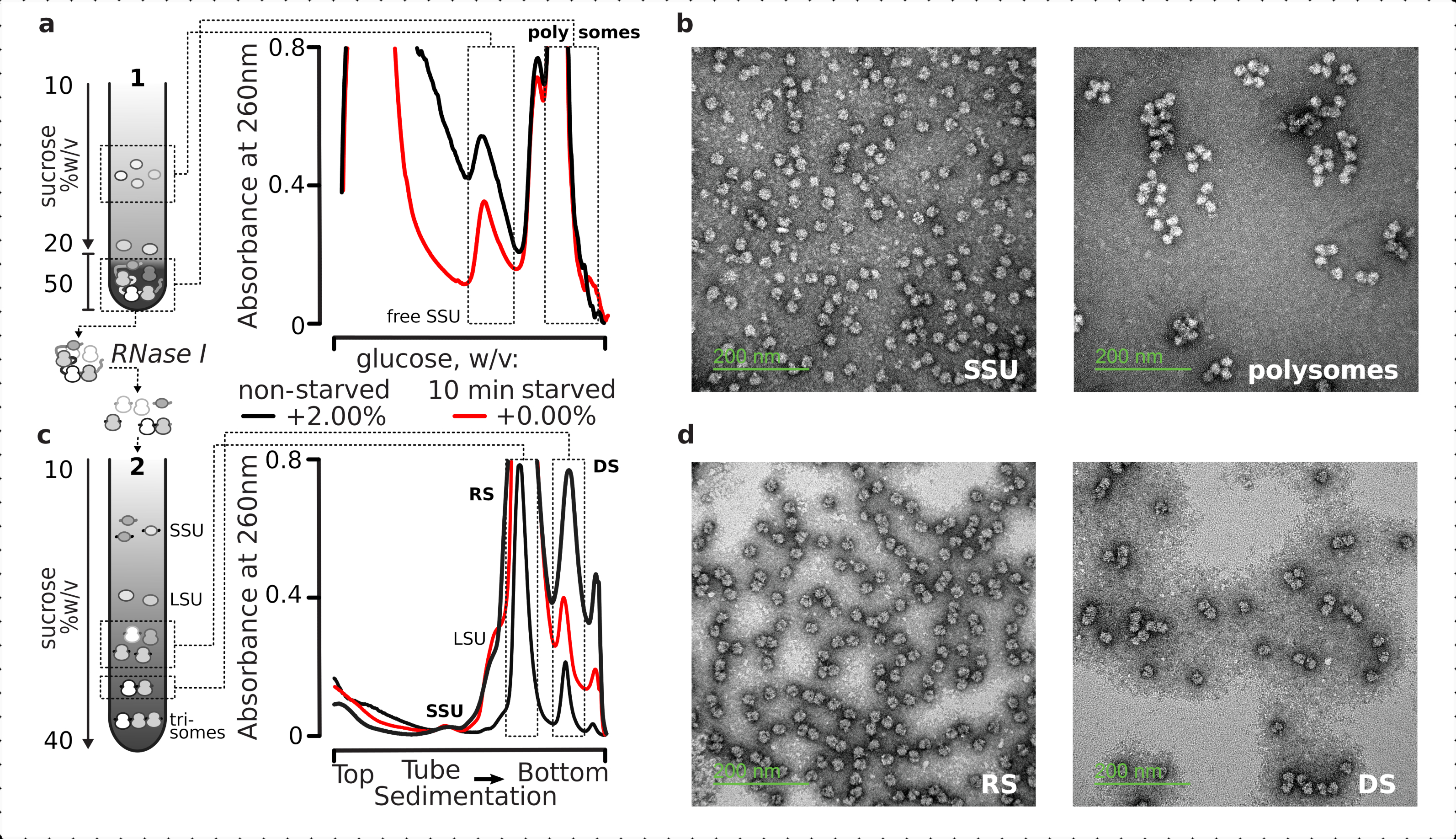{kind=link}
为了检查固定方案对短暂核糖体相关蛋白(特别是eIF)保留的适用性,我们测试了eIF4A的共沉淀,eIF4A是一种不稳定的eIF,动态结合到核糖体组分。我们利用eIF4A串联亲和纯化(TAP)标记的酵母菌株(TIF1-TAP),并使用抗TAP抗体研究了来自固定细胞与非固定细胞的材料中的eIF4A存在,与Pab1p的丰度相比作为额外的RNA结合对照,使用SDS-PAGE然后进行蛋白质印迹(图6)。

图6:体内甲醛固定后翻译复合物中瞬时蛋白的稳定性。(a,b)(顶部图)(a)非固定和(b)2.2%甲醛固定eIF4A-TAP酵母细胞的全细胞裂解物(WCL)通过超速离心分离并可视化如图2图例中所述。(底图)在相应梯度(顶图)中分析的材料分离时,对各自的蔗糖梯度分数进行蛋白质印迹成像,并将WCL作为对照。(c)固定和非固定材料分数中eIF4A或Pab1p丰度之间的平均比率。根据(a,b)(底图)的数据计算了eIF4A(黑条)和Pab1p(灰条)的相对比例(归一化为所有2-7个分数的信号),并得出了它们与固定到非固定比率的2,3(SSU,LSU),4,5(RS,轻多聚体)和6,7(重多聚体)。误差线表示与均值之比的标准差,合并分数(虚线框)被视为重复项。请点击此处查看此图的放大版本。
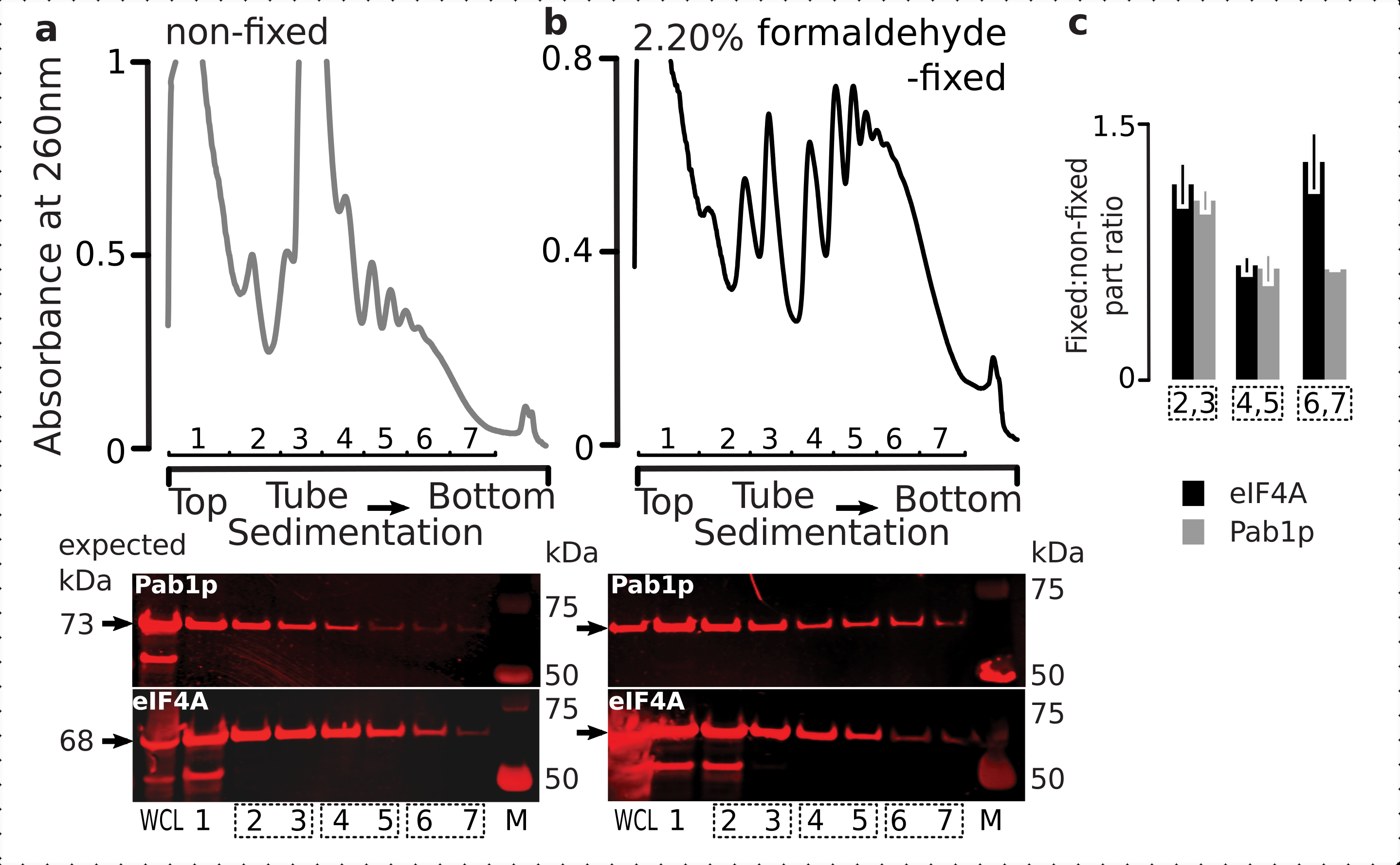{kind=link}
与它们在细胞中的高丰度一致,我们观察到来自全细胞裂解物(WCL)中的蛋白质和来自非固定细胞的较慢沉降组分的高强度信号(图6a,下图)。我们还在WCL中检测到大量来自固定细胞的这些蛋白质,并保证了交联材料提取的效率和没有意外损失(图6b,底图)。然而,与非固定细胞相比,与Pab1p相比,来自固定细胞的材料在沉降较快的核糖体组分中显示出eIF4A的相对存在升高(图6c)。这一结果表明,eIF4A与甲醛交联材料中的聚体仍然更紧密地结合。
在确认交联对核糖体级分中eIF4A存在的积极和特异性稳定作用后,我们使用来自eIF4A标记(TIF1-TAP)酵母菌株的固定材料,通过用磁性IgG磁珠的亲和纯化来捕获和富集含eIF4A的配合物。在通过蔗糖梯度(例如,酵母方案的第1.3节)进行第一次沉降后,我们有亲和力富集的WCL,游离SSU和多体(翻译的mRNA池)级分,以及从翻译池中拆卸成具有RNase I的单个复合物(例如,酵母方案的第1.4节)时第二次沉降的SSU,LSU和RS级分(图7).在所有情况下,除了LSU级分外,我们都能够观察到纯化馏分(洗脱液,E)中eIF4A的选择性富集,与源材料(输入,I)中存在β肌动蛋白(图7)相比。

图7:通过瞬时相关的eIF4A选择性免疫纯化 体内 甲醛稳定的转化复合物。该示意图说明了不同翻译复合物和eIF4A表位的来源,包括eIF4A-TAP酵母细胞的非分馏澄清WCL;游离SSU和翻译的RNA库(多体)在第一次超速离心中分离;SSU,LSU和RS级分通过RNase I消化从翻译的RNA中释放出来,并使用第二次超速离心分离(见文本)。蛋白质印迹图像提供了与同时染色的β肌动蛋白对照的丰度相比,馏分中eIF4A丰度的可视化。 请点击此处查看此图的放大版本。
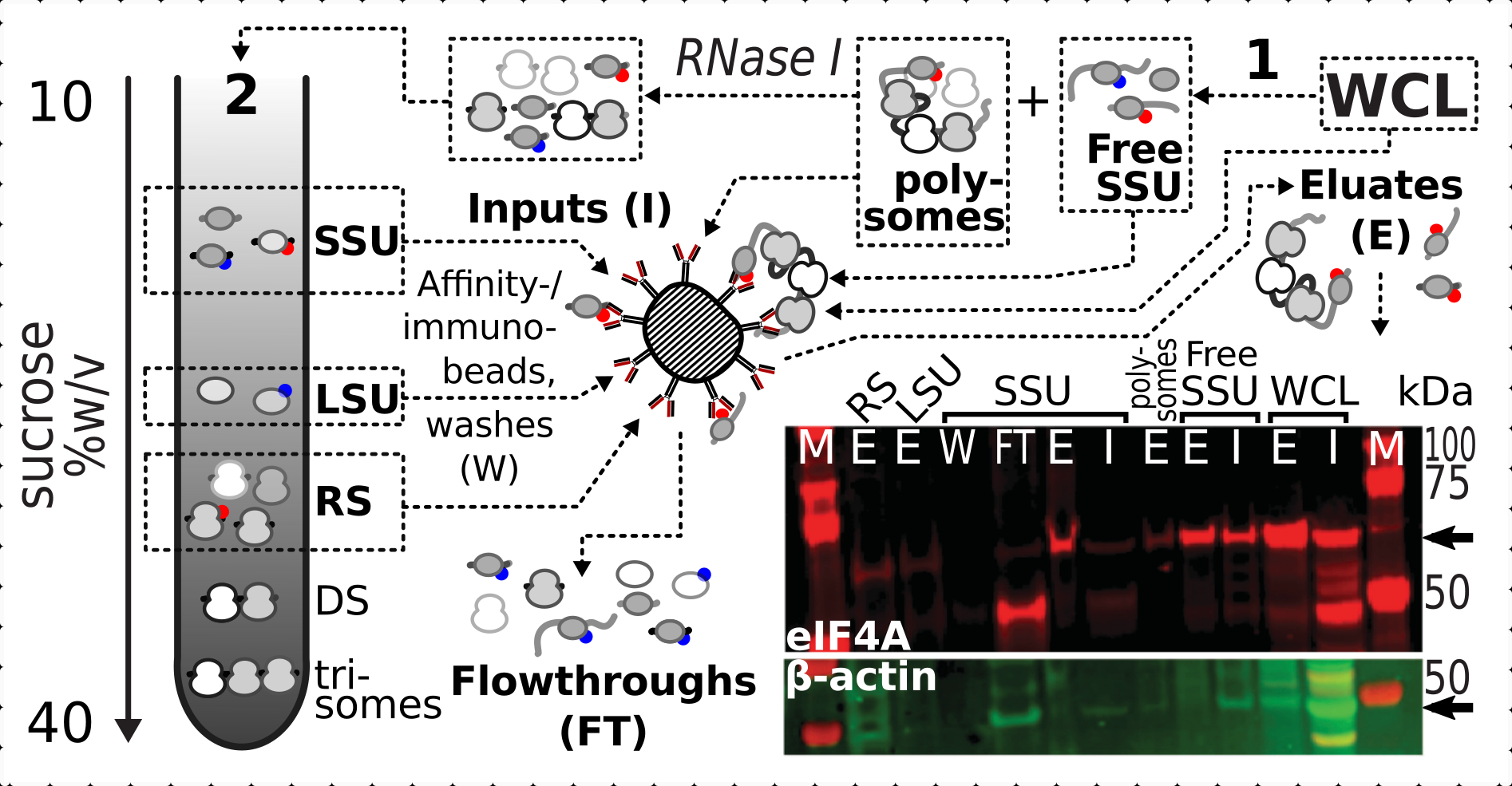{kind=link}
附表1。请点击此处下载此表格。
讨论
甲醛固定是实现生物分子10、36、45、46、47、48体内快速交联的一种方便而流行的方法。与其他潜在的生物分子靶标相比,成功捕获转化复合物需要在细胞或其他材料的快速冷却期间立即固定。如果没有未完成的稳定性,则有可能继续进行与翻译相关的不同过程,从而将复杂分布从未扰动的体内状态49转移开来。与其他转化停滞和核糖体复合物稳定化方法相比,甲醛作用在细胞膜上的迅速性和交联的不加区分性保证了翻译复合物中间体的最大多样性,更接近其天然分布状态50。
这里介绍的方法已经在酵母和哺乳动物细胞中建立和优化,并且现在已经被其他组衍生出来用于更多样化的生物材料,例如在整个脊椎动物(例如,斑马鱼胚胎)10,38,39,49,51,52.虽然这些工作共同保证了该方法的多功能性和广泛适用性,但由于需要优化和调整,翻译复合物的快速甲醛交联可以被认为是难以转置到新型生物材料上的。
该方法成功的首要要求是重新优化甲醛浓度以及细胞收集和破坏技术。渗透性较差,小而圆的酵母细胞需要更高(至少10倍)甲醛浓度和固定细胞的物理破坏。相比之下,培养物中大型和扁平的贴壁哺乳动物细胞很容易过度固定,并且在固定时需要轻柔处理,而固定复合物的提取可以使用洗涤剂通过膜破坏进行化学操作。交联不足可能允许不太稳定或寿命较短的中间体解离或泄漏到较晚的状态。过度交联可能会对分离和研究核糖体组分的能力产生负面影响,并可能产生选择性偏倚,例如重质复合物的更深层消耗。在我们的观察中,即使是微小的改变,例如所使用的贴壁人细胞的类型,也会影响回收的交联复合物的产量,并且可能需要重新优化交联方案。我们还可以预测具有实质上不同渗透性性质的细胞,例如植物细胞,将需要对固定条件进行额外的广泛优化52。然而,很难想象一种生物材料会与这种方法完全不相容。
与哺乳动物固定方案相关的一个考虑因素是用作输入的细胞材料的密度和数量。建议让细胞连续生长,无需重新接种或其他干扰至少2天,以避免对细胞翻译动力学的外部影响。适用于大多数细胞类型,但对于大多数贴壁细胞,始终如一地达到不超过70%的汇合水平将确保没有主要的接触抑制作用,这些效应会对翻译速率产生负面影响和不可预测的影响。
甲醛固定的另一个有趣且可能特别方便的特征源于其不分青红皂白的反应性,即混合分类学系统中对转化复合物的稳定作用。细菌,尤其是线粒体、叶绿体和不同细胞内寄生虫的转化复合物,一直难以用特定的翻译抑制剂靶向。相反,在TCP-seq数据中,映射到线切割转录组的足迹在数据38,39,50中很容易观察到。一个有趣的后续发展可能是使用这种方法来研究整个微群落中的翻译,例如在土壤,水或肠道样本中,可靠的快速翻译停滞和使用任何其他手段的复杂稳定将是有问题的。
还应该提到的是,对于最复杂的材料(例如坚硬和/或笨重的组织),没有什么可以阻止在细胞破碎和材料均质化后立即使用甲醛稳定。这种方法已经经常用于消除用特定的小分子抑制剂33,53,54,55稳定翻译复合物时的细胞进入延迟。鉴于甲醛固定传统上在电子显微镜45,56,57,58等应用中用于体外/体外样品稳定具有出色的效果,在这种情况下,我们可以预期更少的负面影响,特别是那些与从完全固定的细胞中提取转化复合物的不良相关的负面影响。
我们的发现证实了快速甲醛固定的可用性,以稳定高度瞬态的复合物,例如那些含有eIF4A的复合物。值得注意的是,与哺乳动物相比,酵母eIF4A与盖结合复合物eIF4F的相关性要弱得多,因此通常与转化复合物相关。eIF4A通常在酵母29,59,60,61,62,63中的核糖体材料的任何广泛纯化过程中丢失。然而,在体内固定酵母材料中,可以在预期其存在的所有转化配合物中实现eIF4A的可靠富集。先前发表的Sel-TCP-seq数据已经证明了eIF2和eIF3的富集,它们与核糖体更强地结合(但也揭示了短暂发生的共翻译蛋白复合物组装)39。因此,该方法适用于检测两者,更强和较弱的附着成分的平移配合物。
总而言之,我们提出了一种方法,主要用于深入了解翻译起始阶段发生的变化,以及何时需要在mRNA上进行最小扰动的核糖体分布。重要的是,该方法适用于平移复合物(如eIF4A)中相对不稳定和动态的组分的稳定,并且可以广泛用于必要的优化。我们还提供了甲醛固定在翻译快速动态变化的情况下的有用性的证据,开辟了研究领域,例如快节奏的细胞对环境变化或应激条件的反应。
披露声明
作者声明没有利益冲突。
致谢
这项工作得到了澳大利亚研究委员会发现项目赠款(DP180100111至T.P.和N.E.S.),国家卫生和医学研究委员会研究员资助(GNT1175388至N.E.S.)和研究奖学金(APP1135928至T.P.)的支持。作者承认澳大利亚国立大学高级显微镜中心的澳大利亚显微镜设施,该设施由大学和联邦政府资助。
材料
| Name | Company | Catalog Number | Comments |
| Yeast extract | Merck, Sigma-Aldrich | 70161 | |
| Peptone | Merck, Sigma-Aldrich | 70178 | |
| D-Glucose (Dextrose) | Merck, Sigma-Aldrich | 49139 | |
| Adenine sulphate | Amresco | 0607-50G | |
| Formaldehyde solution | Merck Sigma-Aldrich | F11635-500ML | ACS reagent, 37 wt. % in H2O, contains 10-15% Methanol as stabiliser (to prevent polymerisation) |
| RNaseOUT™ Recombinant Ribonuclease Inhibitor | Invitrogen™ byThermo Fischer Scientific | 10777019 | |
| cOmplete™, EDTA-free Protease Inhibitor Cocktail | COEDTAF-RO Roche by Merck | 11873580001 | |
| Magnesium chloride solution | (Merck/Sigma-Aldrich) | M1028 | |
| Ethylenediaminetetraacetic acid solution | (Merck/Sigma-Aldrich) | E7889 | |
| Ambion™ RNase I, cloned, 100 U/µL | Ambion | AM2294 | |
| SUPERase•In™ RNase Inhibitor (20 U/μL) | Invitrogen™ by Thermo Fisher Scientific | AM2694 | |
| Acidic phenol:chlorophorm:isoamyl alcohol 125:24:1 (pH 4.0-5.0) | (Merck/Sigma-Aldrich) | P1944-100ML | |
| Dynabeads™ Goat Anti-Mouse IgG | Invitrogen™ by Thermo Fisher Scientific) | 11033 | |
| Sodium Acetate (3 M), pH 5.5 | Invitrogen™ by Thermo Fisher Scientific) | AM9740 | |
| Glycogen (5 mg/ml) | Invitrogen™ by Thermo Fisher Scientific) | AM9510 | |
| Ethyl alcohol, Pure | Merck; Sigma Aldrich | E7023 | |
| Amersham™ Hybond® P Western blotting membranes, PVDF | Merck | GE10600023 | PVDF membrane for western blotting |
| Bolt™ 4 to 12%, Bis-Tris, 1.0 mm, Mini Protein Gel | Invitrogen™ by ThermoFischer Sientific | NW04120BOX | Protein gel |
| 4X Bolt™ LDS Sample Buffer | Invitrogen™ by ThermoFischer Sientific | B0007 | LDS sample loading buffer |
| Precision Plus Protein™ Kaleidoscope™ Prestained Protein Standards | BioRad | 1610375 | Protein ladder |
| 20X Bolt™ MES SDS Running Buffer | ThermoFischer Scientific | B0002 | PAGE runninjg buffer |
| Intercept® (PBS) Blocking Buffer | LI-COR | 927-70001 | Odyssey Blcoking buffer (PBS) |
| IRDye® 800CW Goat anti-Mouse IgG Secondary Antibody | LI-COR | 92632210 | |
| IRDye® 800CW Goat anti-Rabbit IgG Secondary Antibody | LI-COR | 92632211 | |
| TAP Tag Polyclonal Antibody | Invitrogen™ by ThermoFischer Sientific | CAB1001 | |
| Anti-beta Actin antibody | Abcam | ab8227 | |
| Sucrose | (Merck/Sigma-Aldrich) | 84097 | BioUltra, for molecular biology, ≥99.5% (HPLC) |
| DL-Dithiothreitol solution | (Merck/Sigma-Aldrich) | 43816 | BioUltra, for molecular biology, ~1 M in H2O |
| Terumo Syringe 1CC/mL | Terumo Syringe | 878499 | |
| Potassium chloride | (Merck/Sigma-Aldrich) | 60128 | |
| HEPES | (Merck/Sigma-Aldrich) | H3375 | |
| Dulbecco's Modified Eagle's Medium - high glucose | Sigma Aldrich | D5796 | |
| Fetal Bovine Serum | Sigma Aldrich | 12003C | |
| Trypsin-EDTA (0.05%), phenol red | Gibco | 25300062 | |
| Dulbecco's Phosphate Buffered Saline with Calcium and magnesium | Sigma-Aldrich | D8662 | |
| Glycine | Sigma-Aldrich | G7126 | |
| Tris hydrochloride | Merck/Sigma-Aldrich | 10812846001 | |
| Sodium dodecyl sulfate | Merck/Sigma-Aldrich | 436143 | |
| IGEPAL CA-630 | Merck/Sigma-Aldrich | I3021 | |
| Rnasin Ribonuclease Inhibitor | Promega | N2111 | |
| Stainless steel grinding jar | Retsch | 02.462.0059 | |
| MM400 mixer mill | Retsch | 20.745.0001 | |
| Gradient Fractionator | Brandel | BRN-BR-188 | |
| Thermomixer R | Eppendorf | Z605271 | |
| Nanodrop spectrophotometer | Thermo Fisher Scientific | ND-2000 | |
| 0.5-ml microcentrifuge tubes with locking devices | Eppendorf Safe-Lock | 30121023 | |
| Mini Gel Tank | (Thermo Fisher Scientific) | A25977 | PAGE running tank |
| 5 mL, Open-Top Thinwall Ultra-Clear Tube, 13 x 51mm | Beckman-Coulter | 344057 | |
| 13.2 mL, Certified Free Open-Top Thinwall Polypropylene, 14 x 89mm - 50Pk | Beckman-Coulter | 331372 | |
| Amicon Ultra-0.5 ultrafiltration devices | Merck | UFC5030 | Ultracel-30 regenerated cellulose membrane, 0.5 mL sample volume |
| Thermo Sorvall Evolution RC Floor Super Speed Centrifuge | Cambridge Scientific | 15566 | |
| Beckman Coulter Optima L-90K | GMI | 8043-30-1191 | |
| Nunc EasYFlask 175cm2 | Thermofisher Scientific | 159910 | |
| Falcon 50 mL Conical Centrifuge Tubes | Thermofisher Scientific | 14-432-22 | |
| 25 mL Serological Pipette | Sigma-Aldrich | SIAL1250 | |
| 10 mL Serological Pipette | Sigma-Aldrich | SIAL1100 | |
| DNA lobind tubes | Eppendorf | 30108051 | |
| Cold Centrifuge 5810 R | Eppendorf | EP022628188 | for 50 mL tubes |
| Orbital Shaking Incubator | Ratek | OM11 | |
| Frezco 17 Microcentrifuge | Thermofisher Scientific | 75002402 | |
| Eppendorf DNA lo-bind tubes | Merck/Sigma-Aldrich | EP0030108051 | |
| Eppendorf® Protein LoBind tubes | Merck/Sigma-Aldrich | EP0030108116 | |
| SW 41 Ti Swinging bucket rotor | Beckman-Coulter | 331362 | |
| Heracell™ 150i CO2 Incubator, 150 L | Thermofisher Scientific | 51026282 | |
| 0,3 mL ultra-fine II short insulin syringe | BD Medical | 328822 | |
| 3 mL syringe with Luer Lok tip | BD Medical | 302113 | |
| 25 G x 16 mm Hypodermic Needle | Terumo | TUAN2516R1 |
参考文献
- Janapala, Y., Preiss, T., Shirokikh, N. E. Control of translation at the initiation phase during glucose starvation in yeast. International Journal of Molecular Sciences. 20 (16), 4043 (2019).
- Masvidal, L., Hulea, L., Furic, L., Topisirovic, I., Larsson, O. mTOR-sensitive translation: Cleared fog reveals more trees. RNA Biology. 14 (10), 1299-1305 (2017).
- Ashe, M. P., De Long, S. K., Sachs, A. B. Glucose depletion rapidly inhibits translation initiation in yeast. Molecular Biology of the Cell. 11 (3), 833-848 (2000).
- Crawford, R. A., Pavitt, G. D. Translational regulation in response to stress in Saccharomyces cerevisiae. Yeast. 36 (1), 5-21 (2019).
- Melamed, D., Pnueli, L., Arava, Y. Yeast translational response to high salinity: global analysis reveals regulation at multiple levels. RNA. 14 (7), 1337-1351 (2008).
- Hershey, J. W., Sonenberg, N., Mathews, M. B. Principles of translational control: An overview. Cold Spring Harbor Perspectives in Biology. 4 (12), 011528 (2012).
- Mata, J., Marguerat, S., Bähler, J. Post-transcriptional control of gene expression: a genome-wide perspective. Trends in Biochemical Sciences. 30 (9), 506-514 (2005).
- Spriggs, K. A., Bushell, M., Willis, A. E. Translational regulation of gene expression during conditions of cell stress. Molecular Cell. 40 (2), 228-237 (2010).
- Liu, B., Qian, S. B. Translational reprogramming in cellular stress response. Wiley Interdisciplinary Reviews RNA. 5 (3), 301-315 (2014).
- Archer, S. K., Shirokikh, N. E., Beilharz, T. H., Preiss, T. Dynamics of ribosome scanning and recycling revealed by translation complex profiling. Nature. 535 (7613), 570-574 (2016).
- Hinnebusch, A. G., Ivanov, I. P., Sonenberg, N. Translational control by 5'-untranslated regions of eukaryotic mRNAs. Science. 352 (6292), 1413-1416 (2016).
- Dever, T. E., Green, R. The elongation, termination, and recycling phases of translation in eukaryotes. Cold Spring Harbor Perspectives in Biology. 4 (7), 013706 (2012).
- Shirokikh, N. E., Preiss, T. Translation initiation by cap-dependent ribosome recruitment: Recent insights and open questions. Wiley Interdisciplinary Reviews RNA. 9 (4), 1473 (2018).
- Jiménez-Díaz, A., Remacha, M., Ballesta, J. P., Berlanga, J. J. Phosphorylation of initiation factor eIF2 in response to stress conditions is mediated by acidic ribosomal P1/P2 proteins in Saccharomyces cerevisiae. PLoS One. 8 (12), 84219 (2013).
- Sonenberg, N., Hinnebusch, A. G. Regulation of translation initiation in eukaryotes: mechanisms and biological targets. Cell. 136 (4), 731-745 (2009).
- Majmundar, A. J., Wong, W. J., Simon, M. C. Hypoxia-inducible factors and the response to hypoxic stress. Molecular Cell. 40 (2), 294-309 (2010).
- Barraza, C. E., et al. The role of PKA in the translational response to heat stress in Saccharomyces cerevisiae. PLoS One. 12 (10), 0185416 (2017).
- Richter, K., Haslbeck, M., Buchner, J. The heat shock response: Life on the verge of death. Molecular Cell. 40 (2), 253-266 (2010).
- Jamar, N. H., Kritsiligkou, P., Grant, C. M. The non-stop decay mRNA surveillance pathway is required for oxidative stress tolerance. Nucleic Acids Research. 45 (11), 6881-6893 (2017).
- Chen, Z., et al. The complete pathway for thiosulfate utilization in Saccharomyces cerevisiae. Applied and Environmental Microbiology. 84 (22), (2018).
- Marzluf, G. A. Molecular genetics of sulfur assimilation in filamentous fungi and yeast. Annual Review of Microbiology. 51, 73-96 (1997).
- Miller, D., Brandt, N., Gresham, D. Systematic identification of factors mediating accelerated mRNA degradation in response to changes in environmental nitrogen. PLoS Genetics. 14 (5), 1007406 (2018).
- Zhang, W., Du, G., Zhou, J., Chen, J. Regulation of sensing, transportation, and catabolism of nitrogen sources in Saccharomyces cerevisiae. Microbiology and Molecular Biology Reviews. 82 (1), (2018).
- Tokpohozin, S. E., Fischer, S., Becker, T. Selection of a new Saccharomyces yeast to enhance relevant sorghum beer aroma components, higher alcohols, and esters. Food Microbiology. 83, 181-186 (2019).
- Walker, G. M., Stewart, G. G. Saccharomyces cerevisiae in the production of fermented beverages. Beverages. 2 (4), 30 (2016).
- Chassé, H., Boulben, S., Costache, V., Cormier, P., Morales, J. Analysis of translation using polysome profiling. Nucleic Acids Research. 45 (3), 15 (2017).
- Jin, H. Y., Xiao, C. An integrated polysome profiling and ribosome profiling method to investigate in vivo translatome. Methods in Molecular Biology. 1712, 1-18 (2018).
- Arava, Y., et al. Genome-wide analysis of mRNA translation profiles in Saccharomyces cerevisiae. Proceedings of the National Academy of Sciences of the United States of America. 100 (7), 3889-3894 (2003).
- Lackner, D. H., et al. A network of multiple regulatory layers shapes gene expression in fission yeast. Molecular Cell. 26 (1), 145-155 (2007).
- Ingolia, N. T., Ghaemmaghami, S., Newman, J. R. S., Weissman, J. S. Genome-Wide Analysis in Vivo of Translation with Nucleotide Resolution Using Ribosome Profiling. Science. 324 (5924), 218-223 (2009).
- Ingolia, N. T., Hussmann, J. A., Weissman, J. S. Ribosome Profiling: Global Views of Translation. Cold Spring Harbor Perspectives in Biology. 11 (5), (2019).
- Gerashchenko, M. V., Lobanov, A. V., Gladyshev, V. N. Genome-wide ribosome profiling reveals complex translational regulation in response to oxidative stress. Proceedings of the National Academy of Sciences of the United States of America. 109 (43), 17394-17399 (2012).
- Hussmann, J. A., Patchett, S., Johnson, A., Sawyer, S., Press, W. H. Understanding biases in ribosome profiling experiments reveals signatures of translation dynamics in yeast. Proceedings of the National Academy of Sciences Genetics. 11 (12), 1005732 (2015).
- Santos, D. A., Shi, L., Tu, B. P., Weissman, J. S. Cycloheximide can distort measurements of mRNA levels and translation efficiency. Nucleic Acids Research. 47 (10), 4974-4985 (2019).
- Schneider-Poetsch, T., et al. Inhibition of eukaryotic translation elongation by cycloheximide and lactimidomycin. Nature Chemical Biology. 6 (3), 209-217 (2010).
- Hoffman, E. A., Frey, B. L., Smith, L. M., Auble, D. T. Formaldehyde crosslinking: A tool for the study of chromatin complexes. Journal of Biological Chemistry. 290 (44), 26404-26411 (2015).
- Kage, U., Powell, J. J., Gardiner, D. M., Kazan, K. Ribosome profiling in plants: What is not lost in translation. Journal of Experimental Botany. 71 (18), 5323-5332 (2020).
- Shirokikh, N. E., Archer, S. K., Beilharz, T. H., Powell, D., Preiss, T. Translation complex profile sequencing to study the in vivo dynamics of mRNA-ribosome interactions during translation initiation, elongation and termination. Nature Protocols. 12 (4), 697-731 (2017).
- Wagner, S., et al. Selective translation complex profiling reveals staged initiation and co-translational assembly of initiation factor complexes. Molecular Cell. 79 (4), 546-560 (2020).
- Zlotorynski, E. Profiling ribosome dynamics. Nature Reviews Molecular Cell Biology. 17 (9), 535-535 (2016).
- Sen, N. D., Gupta, N., S, K. A., Preiss, T., Lorsch, J. R., Hinnebusch, A. G. Functional interplay between DEAD-box RNA helicases Ded1 and Dbp1 in preinitiation complex attachment and scanning on structured mRNAs in vivo. Nucleic Acids Research. 47 (16), 8785-8806 (2019).
- Zhao, J., Qin, B., Nikolay, R., Spahn, C. M. T., Zhang, G. Translatomics: The global view of translation. International Journal of Molecular Sciences. 20 (1), 20010212 (2019).
- Luthe, D. S. A simple technique for the preparation and storage of sucrose gradients. Analytical Biochemistry. 135 (1), 230-232 (1983).
- Wang, Z., Gerstein, M., Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nature Review Genetics. 10 (1), 57-63 (2009).
- Orlando, V. Mapping chromosomal proteins in vivo by formaldehyde-crosslinked-chromatin immunoprecipitation. Trends in Biochemical Sciences. 25 (3), 99-104 (2000).
- Schmiedeberg, L., Skene, P., Deaton, A., Bird, A. A Temporal Threshold for Formaldehyde Crosslinking and Fixation. PLoS One. 4 (2), 4636 (2009).
- Solomon, M. J., Varshavsky, A. Formaldehyde-mediated DNA-protein crosslinking: A probe for in vivo chromatin structures. Proceedings of the National Academy of Sciences. 82 (19), 6470-6474 (1985).
- Solomon, M. J., Larsen, P. L., Varshavsky, A. Mapping proteinDNA interactions in vivo with formaldehyde: Evidence that histone H4 is retained on a highly transcribed gene. Cell. 53 (6), 937-947 (1988).
- Bohlen, J., Fenzl, K., Kramer, G., Bukau, B., Teleman, A. A. Selective 40S footprinting reveals cap-tethered ribosome scanning in human cells. Molecular Cell. 79 (4), 561-574 (2020).
- Shirokikh, N. E. Translation complex stabilization on messenger RNA and footprint profiling to study the RNA responses and dynamics of protein biosynthesis in the cells. Critical Reviews in Biochemistry and Molecular Biology. , (2021).
- Giess, A., et al. Profiling of small ribosomal subunits reveals modes and regulation of translation initiation. Cell Reports. 31 (3), 107534 (2020).
- Firmino, A. A. P., et al. Separation and paired proteome profiling of plant chloroplast and cytoplasmic ribosomes. Plants (Basel). 9 (7), (2020).
- Gerashchenko, M. V., Gladyshev, V. N. Translation inhibitors cause abnormalities in ribosome profiling experiments. Nucleic Acids Research. 42 (17), 134 (2014).
- Santos, D. A., Shi, L., Tu, B. P., Weissman, J. S. Cycloheximide can distort measurements of mRNA levels and translation efficiency. Nucleic Acids Research. 47 (10), 4974-4985 (2019).
- Schneider-Poetsch, T., et al. Inhibition of eukaryotic translation elongation by cycloheximide and lactimidomycin. Nature Chemical Biology. 6 (3), 209-217 (2010).
- Plénat, F., et al. Formaldehyde fixation in the third millennium. Annales De Pathologie. 21 (1), 29-47 (2001).
- Salic, A., Mitchison, T. J. A chemical method for fast and sensitive detection of DNA synthesis in vivo. Proceedings of the National Academy of Sciences. 105 (7), 2415-2420 (2008).
- Wang, N. S., Minassian, H. The formaldehyde-fixed and paraffin-embedded tissues for diagnostic transmission electron microscopy: A retrospective and prospective study. Human Pathology. 18 (7), 715-727 (1987).
- Grifo, J. A., et al. Characterization of eukaryotic initiation factor 4A, a protein involved in ATP-dependent binding of globin mRNA. Journal of Biological Chemistry. 257 (9), 5246-5252 (1982).
- Li, Y. Commonly used tag combinations for tandem affinity purification. Biotechnology and Applied Biochemistry. 55 (2), 73-83 (2010).
- Blum, S., et al. ATP hydrolysis by initiation factor 4A is required for translation initiation in Saccharomyces cerevisiae. Proceedings of the National Academy of Sciences. 89 (16), 7664-7668 (1992).
- Merrick, W. C. eIF4F: A Retrospective. Journal of Biological Chemistry. 290 (40), 24091-24099 (2015).
- Rogers, G. W., Komar, A. A., Merrick, W. C. eIF4A: The godfather of the DEAD box helicases. Progress in Nucleic Acid Research and Molecular Biology. 72, 307-331 (2002).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。