
Method Article
啮齿动物组织的霰弹枪脂质组学
摘要
基于鸟枪式质谱的脂质组学可在各种啮齿动物组织的单次测量中同时提供广谱脂质类别的灵敏定量快照。
摘要
脂质作为所有原核细胞和真核细胞的重要组成部分起着至关重要的作用。质谱技术的不断改进使脂质组学成为监测稳态和疾病状态下组织脂质组组成的强大分析工具。本文提出了一种用于鸟枪式脂质分析方法的分步方案,该方法支持以高通量同时检测和定量不同组织和生物流体样品中的数百种分子脂质。该方法利用自动纳米流直接进样加标标记内标的总脂质提取物,无需色谱分离即可进入高分辨率质谱仪。从亚微克量的啮齿动物组织开始,MS分析每个样品需要10分钟,涵盖小鼠肺组织中14种脂质类别的多达400种脂质。这里介绍的方法非常适合研究疾病机制以及识别和量化表明啮齿动物组织内早期毒性或有益作用的生物标志物。
引言
香烟烟雾 (CS) 被认为是与肺部慢性炎症性疾病发展相关的主要危险因素,包括肺癌、支气管炎和慢性阻塞性肺病 (COPD)1。除了肺部影响之外,CS暴露在其他疾病的发展中起着重要作用,例如动脉粥样硬化性冠状动脉疾病和外周血管疾病1。心血管疾病与慢性阻塞性肺病分别是全球第一和第三大死亡原因。毒理学风险评估方法历来依赖于使用啮齿动物等动物模型。 体内 仅鼻子或全身大鼠和小鼠模型通常用于研究暴露于CS的长期影响。
例如,一般来说,6个月的烟雾暴露会诱发小鼠肺的组织学和功能异常,类似于人类疾病,包括肺气肿、气道重塑和肺动脉高压,尽管与长期人类吸烟者相比,这些变化相对较轻2。在动物和人体组织中,研究观察到对CS暴露的反应发生了广泛的分子变化,包括氧化应激反应,炎症和结构组织变化3,4。毫不奇怪,据报道,CS 暴露对肺脂质组也有深远的影响,包括对表面活性剂脂质、脂质信号传导介质和结构脂质的影响4,5,6。
为了表征小鼠肺长期CS暴露引起的大量脂质变化,我们进行了快速定量的霰弹枪直接输注质谱分析。在2005年引入鸟枪式脂质组学方法7后,该方法已被有效地用于扩展我们对酵母11,秀丽隐杆线虫12和黑腹果蝇13等模型系统中脂质细胞代谢8,9,10的知识,以及广泛的哺乳动物样品类型,例如细胞系14, 15、16为各种人类17、18和啮齿动物组织19、20和体液21、22。
在过去的几十年里,研究揭示了细胞对环境变化的反应的复杂性,涉及数千种相互关联的蛋白质、脂质和代谢物。这清楚地表明,使用最先进的分析技术对于深入了解分子机器和揭示外源性生理影响的全部规模至关重要。在这种情况下,由鸟枪式脂质组学方法产生的全面定量脂质指纹图谱可以有效地增加我们对脂质细胞代谢的了解8,9,10。
关于CS暴露作为几种疾病的危险因素,毒理学风险评估方法历来依赖于使用啮齿动物等动物模型。Shotgun-Lipidomics MS 提供了一种快速、灵敏的定量分析工具,用于评估多种样品类型的脂质组扰动。鸟枪法脂质组学的独特之处在于,通过使用产生电喷雾电离(ESI)纳米喷雾23的导电纳米芯片,无需色谱分离即可自动直接直接分析总脂质提取物(加标内标),无需色谱分离即可进入高分辨率MS仪器。
在MS1模式下同时获取的质荷比信息提供了所有完整内源性脂质的总脂质足迹。可选地,MS2/MS3模式(其中母体脂质分子被碎片化和分析)提供了额外的结构信息。数据分析需要专门的软件,包括光谱的反卷积和合并光谱中的峰分配,从而导致脂质鉴定和假定的化学结构解析。此外,可以通过加标标记的内标混合物来进行绝对定量,该混合物含有每个目标脂质类别至少一种脂质标准品。总体而言,使用本技术,每个样品只需几分钟的MS分析可以覆盖啮齿动物组织中14个脂质类别24 中多达800种脂质的鉴定和定量。
研究方案
所有涉及动物的程序均在由国际实验动物护理评估和认证协会认可并获得新加坡农业食品和兽医局许可的设施中进行,并得到机构动物护理和使用委员会的批准,并符合国家实验动物研究咨询委员会关于科学目的护理和使用动物的指南(NACLAR, 2004).
1. 样品采集
- 在暴露于ApoE - / / - 小鼠3个月和6个月后进行小鼠肺解剖。暴露后16-24小时收集组织,在液氮中快速冷冻,并在启动脂质组学工作流程之前储存在-80°C。确保从每个"组学"解剖组中收集总共九个样本。
2. 组织 - 样品的粉碎
- 准备用于组织研磨的磁锤和耗材。预冷纸巾袋、镊子、带盖组织转移管、"V"型塑料刮刀和干冰上的 2 mL 塑料收集管。将相应的管架安装在磁锤仪器上。打开磁锤并将冲击力设置为 高。
- 将样品装入纸巾袋中;如果需要,使用预冷的镊子将样品插入组织袋的顶部开口。将样品放在柔性袋的中心,远离边缘。
- 通过将预冷的收集管拧到纸巾袋的顶部来密封纸巾袋。
- 通过将柔性袋浸入液氮中 10 秒来快速冷冻样品。
- 排气收集管;松开管子 1/4 圈进行排气,以避免在磁锤撞击时使袋子破裂。
- 将冷冻纸巾袋装入磁锤上。
- 按下 激活 按钮应用磁锤以粉碎样品。如果袋中残留有未粉碎的组织碎片,请再次在液氮中快速冷冻样品并重复第二次冲击。
- 从磁锤上取出纸巾袋,将粉碎的样品保持在底部。为防止样品融化,请立即再次快速冷冻。
- 将样品转移到收集管中。快速倒置管,使组织袋位于顶部,并敲击袋以将组织颗粒转移到收集管的底部。
注意:此步骤应快速完成,以避免组织融化。 - 将10mg组织等分试样转移到2 mL管中进行进一步的脂质组学分析,并将样品储存在-80°C,直到执行进一步的步骤。
3. 组织均质化
- 打开组织裂解器仪器,将 振荡频率 设置为 30 Hz , 振荡持续时间 设置为 2分钟。制备组织匀浆缓冲溶液:150mM碳酸氢铵在水中,pH值为~8.2。
- 从储存冰箱中取出样品,放在冰上,然后将四个不锈钢珠子加入装有组织粉末的管中。向每个样品中加入 0.5 mL 的 150 mM 碳酸氢铵缓冲液。
- 将样品管放入组织裂解器支架中。用相同数量的管子平衡两个支架。将支架固定到组织裂解器固定臂中。破坏组织。
- 将试管放在冰上,目视检查试管中是否存在残留的组织聚集体。如果组织破碎不完全(可见聚集体),则通过执行另一个治疗周期重复破坏过程。将样品放在冰上。
- 在专用于蛋白质测定的 1.5 mL 管中制备 100 μL 样品的等 分试样 1 。在10°C下以18,200× g 离心5分钟。
- 通过布拉德福德测定法测定等 分试样1 的蛋白质含量(见第4节)。将样品储存在冰上。
- 在新的 2 mL 微管中制备 20-100 μL 原液匀浆的等 分试样 2 ,用于脂质提取(参见第 5 节)。如果未立即分析样品,请将其保存在冰上,直到进行提取。
注意:最终提取量必须根据总蛋白质量定义。它必须在每总等分试样 0.015-0.045 毫克蛋白质的范围内。
4. 布拉德福德蛋白测定法
注意:研究样品应在分析开始前随机化,并且样品处理的所有步骤都遵循随机化。
- 用碳酸氢铵缓冲液稀释等 分试样1 上清液2x。
注意:稀释系数取决于组织匀浆的浓度。对于更浓缩的溶液,应用更高的因子,使确定的浓度落在测定的动态范围内。 - 根据 表1制备牛血清白蛋白(BSA)标准曲线。涡旋标准管。在室温下以 18,200 × g 离心管 15 秒。
- 从先前制备的标准管中,根据 图1所示的板布局将空白和标准品各转移6 μL到96孔平底板上。
- 根据 图1中描述的板布局将先前制备的样品转移到96孔平底板上。
- 使用多通道移液器和混合物向每个孔中加入 250 μL Bradford 试剂。孵育5分钟。
- 使用读板器测量595nm波长处的吸光度。
- 根据标准曲线计算等分试样中的蛋白质浓度。
注意:在计算蛋白质浓度时,应考虑程序开始时使用的稀释因子。
5. 浮肿提取
注意:避免使用低结合塑料管进行脂质组学提取,因为强有机溶剂会释放增塑剂并在样品分析过程中产生强背景。
- 在总空白 (TB)、内标空白 (ISB) 和质控样品 (QC) 的每个试管中制备 1.5 mL 管中加入 100 μL 碳酸氢铵溶液,考虑在每组 10 个样品之间放置 1 个 QC 样品。将所有样品放在冰上。
- 向所有 QC 样品中加入 10 μL 混合人血浆。将 10 μL 内标溶液加标到所有 ISB、QC 和研究样品(即除 TB 以外的所有样品)中。
注意:对于质量控制材料,请使用任何混合的市售K2EDTA等离子体。在收到时,使用上述方案量化混合血浆中的脂质浓度,并将这些范围作为使用QC材料的所有分析的参考。NIST血浆仅用于更有针对性的应用或需要解决分析准确性的全局方法验证。 - 向每个样品中加入 300 μL −20 °C 丁醇/甲醇混合物 (3/1, v/v)。在室温下在恒温下以450× g 摇动10分钟。
- 加入 300 μL 庚烷/乙酸乙酯混合物 (3/1, v/v)。在室温下在恒温下以450× g 摇动10分钟。
- 加入 300 μL 1% 乙酸混合物。在室温下在恒温下在 恒温下摇 动5分钟×。在室温下以2,800× g 离心5分钟。将 360 μL 上相转移到新的 2 mL 自锁管中 2.
注意:液-液萃取可能导致相边界的位置以样品依赖的方式移动,这可能导致上相的体积差异很小。如有必要,调整上层的收集量。 - 向水相管 1 中加入 320 μL 庚烷/乙酸乙酯 (3/1, v/v)。在室温下在恒温下以450 ×g 摇动5分钟。在室温下以2,800× g 离心5分钟。转移320μL上相,并与步骤5.11中的馏分合并。
注意:如有必要,请调整上层的收集量。 - 向管 1 中的水相中加入 250 μL 庚烷/乙酸乙酯 (3/1, v/v)。在室温下在热混合器上以450× g 摇动5分钟。在室温下以2,800× g 离心5分钟。转移 200 μL 上相,并与管 2 中步骤 5.11 和步骤 5.15 中的馏分合并。
注意:如有必要,请调整上层的收集量。 - 在真空浓缩器中蒸发至35°C干燥。
注意:如果需要,干燥的样品可以储存在-20°C直至分析。 - 重新溶解在 300 μL MS 混合物溶液(7.5 mM 乙酸铵在异丙醇/甲醇/氯仿中的溶液,4:2:1 溶液)中。涡旋每个管5秒以确保所有东西都溶解。在4°C下以18,200× g 离心5分钟。
- 根据 图 2 中的板布局将 50 μL 的等分试样转移到微升板中,以进行正电离模式分析(第 1-6 列),并用 50 μL MS 混合溶液稀释。
- 根据 图 2 中的板布局将 20 μL 的等分试样转移到 MTP 板中,以进行负电离模式分析(第 7-12 列),并用 80 μL MS 混合物稀释。用箔纸包裹板,并在分析前保持在4°C。
6. 质谱分析
- 在分析前 5 天或更短时间内,按照制造商的说明在正负模式下校准质谱仪 (MS)。
- 预先安装直接输液纳米源,确保其与MS的转移毛细管正确对齐。 将4.1μm喷嘴芯片安装到芯片支架中并在软件中进行配置。
- 使用以下参数进行正负模式方法设置:气体压力为 1.25 psi;源电压为 1.1 kV;5 μL 样品进样体积;输出触点闭合 Rel1 持续 2.5 秒;5分钟的交货时间;板在4°C下冷却。
- 为正模式的直接输液机器人设置分析序列队列。
- 使用以下 MS 设置将 MS 方法设置为 正模式 :
- 将毛细管温度设置为 250 °C,将 S 透镜射频电平设置为 65.0。以 140,000 分辨率在 550-1000 m/z 范围内执行 0 至 1 分钟的全扫描 MS 采集,自动增益控制为 1 ×106,最长进样时间为 50 ms。应用锁定质量 680.48022。
- 在 17,500 分辨率下在 1 到 5 分钟的时间范围内设置数据无关的 MS/MS 采集方法,固定的第一质量为 250 m/z。在 1 × 105 和最大注入时间为 64 ms、碰撞能量为 20 NCE 和 1 m/z 的隔离窗口下,对 MS2 使用自动增益控制。使用夹杂物质量列表,范围为 550 至 1,000 m/z,质量步长为 1 Da。
- 对于 负模式下的采集,请在MS调谐文件中将 毛细管温度 设置为 250°C , 将S-Lens RF电平 设置为 65.0 。
- 使用以下MS方法设置:全扫描采集模式从0到1分钟,分辨率为140,000,覆盖400-940 m/z范围,自动增益控制为1×106;最长注入时间为50毫秒;使用锁质量 529.46262。
- 在运行 1 到 5 分钟之间以 17,500 的分辨率在 DIA 模式下设置 MS2 采集,固定的第一质量为 150 m/z;自动增益控制 1 × 105;64毫秒的最长注射时间;和35NCE的碰撞能量。使用从 400 到 940 的夹杂物质量列表,质量步长为 1 Da。
- 在MS软件中以 正 模式创建分析序列队列。等待样品采集由来自每个样品的直接输液机器人的接触闭合信号触发。
- 正模式分析完成后,为 负 模式的直接输液机器人创建序列队列。
- 在 负 模式下在MS软件中设置分析序列队列。
7. 数据处理
- 要将正负模式中的数据文件从.raw格式转换为 .mzML 格式,请使用转换器软件。
注意:正模式和负模式的数据文件必须分开转换(因为它们的采集设置和噪声阈值不同)。- 填写以下设置以执行文件转换:MS 和 MS2 的噪声阈值因子分别为 3 和 5;激活MS和MS2的噪声消除功能,数据压缩,平均MS2扫描和峰值拾取。为正负模式设置TIC阈值(例如,10,000和5,000;根据特定仪器产生的特定样本的噪声水平定义)。在处理结束时生成 XLC 和 PDF 报告。
- 进行脂质鉴定。定义以下(示例)文件导入设置参数:选择窗口±0.5 Da(取决于MS方法中的选择窗口);时间范围 2-300 秒(取决于 MS 方法中的扫描时间);没有MS和MS2的校准质量数;从逐峰软件元数据文件(最小质量 [MS])和(最小质量 [MS2])传输并四舍五入质量范围;MS和MS2的容差分别为3 ppm和10 ppm;将 MS 和 MS2 阈值字段、频率滤波器、MS1 偏移量和 PMO 留空;选择MS和MS2的同位素校正;转换器元数据文件中的分辨率梯度作为(分辨率线性拟合 [MS])和(分辨率线性拟合 [MS2])传输。
注意:由于采集设置不同,阳性和阴性模式下的脂质鉴定必须分开进行。 - 导入数据后,转到 "运行 "菜单并上传MFQL文件以进行脂质鉴定。
注:有关MFQL结构的详细信息,请参阅Herzog等人25的出版物。请参阅 https://wiki.mpi-cbg.de/lipidx/MFQL_library 中的 MFQL 文件的一些示例并查看讨论。用于数据处理的所有MFQL均在补充材料中提供。 - 通过将脂质特征的母离子质量强度除以标记内标的相应强度,然后将商乘以标记内标的浓度,在MS水平上量化鉴定的物质。标准化布拉德福德测定法测量的每量总蛋白质的最终脂质浓度。
8. 质量控制检查程序
注意:质量检查程序是验证方法技术重现性的重要步骤。为此,分析商业混合血浆以确定每批15个相同等分试样(共5批)在5天内不同脂质的内源性水平。请注意,在本例中,数据评估管道是使用 R 统计软件环境创建为 Shiny Web 应用程序。
- 将参考验收范围定义为为所有五个批次计算的平均值±3个标准差。
- 对每个分析批次应用"通过"验收规则:考虑到一种参比化合物代表一种脂质类别,90%的参比靶标必须通过。例如,如果 QC 验证中包含 15 个参考目标,请确保 13 个目标通过才能接受该批次。
注意:换句话说,每种参比化合物必须通过68%的QC样品才能接受该脂质类别的数据。 - 如果研究批次不符合QC检查的验收标准,请重复该过程。
9. (相对)共轭脂肪酸量的估计
- 对于每个脂质类别,根据其MS2强度估计共轭脂肪酸的量。
注意:由于碎裂和电离差异,推导值仅用于样品组之间的相对丰度比较,而不是例如估计特定脂肪酸对脂质类别的总共轭脂肪酸库的相对贡献。 - 通过相应内标脂肪酸片段的平均强度来标准化脂肪酸片段的MS2强度。根据内标的浓度和测量的组织重量,得出共轭脂肪酸的"标准化浓度"。将这些值汇总每个脂质类别,以估计每个样品中每种共轭脂肪酸的总量。
10. 统计分析
- 执行统计分析。为每个条件和相应的对照组拟合线性模型,并根据 log2 转换数据的 t 统计量计算 p 值。使用 Benjamini-Hochberg 错误发现率方法纠正多种测试效应。
注意:调整后的p值<0.05的脂质被认为是差异丰度的。这里,在R统计环境26中进行统计分析。
结果
在这里,作为代表性的结果,我们提供了说明如何使用鸟枪脂质组学来研究CS对小鼠肺部的影响的数据。霰弹枪脂质分析方案已成功应用于两组样品的 体内 研究:从暴露于CS的9只雌性 Apoe− / - 小鼠(3R4F组;阳性对照)解剖肺组织,以及在新鲜空气条件下维持的相同数量的小鼠(假组)作为3个月时收集的阴性对照, 4个月和6个月的暴露时间点。将动物保持在22°C±2°C温度和55%±15%湿度的过滤和调节新鲜空气中,并用γ辐照的颗粒饲料喂养。光照状态维持在每天12小时。每个笼子最多饲养八只小鼠。从肯塔基大学购买了用于暴露处理的 3R4F 参考香烟,以产生 3R4F CS 气溶胶(600 μg 总颗粒物 TPM/L 气溶胶),目标暴露浓度相当于 28 μg 尼古丁/L.CS根据 Phillips 等人的说法,使用 30 端口旋转吸烟机产生 3R4F 香烟的烟雾27.根据加拿大卫生部强化吸烟协议,对 3R4F 香烟施加 55 mL 的抽吸量、每 30 秒一口和 100% 堵塞通风孔以提供足够的暴露。关于研究设计的所有其他细节已在之前报告过27,28。
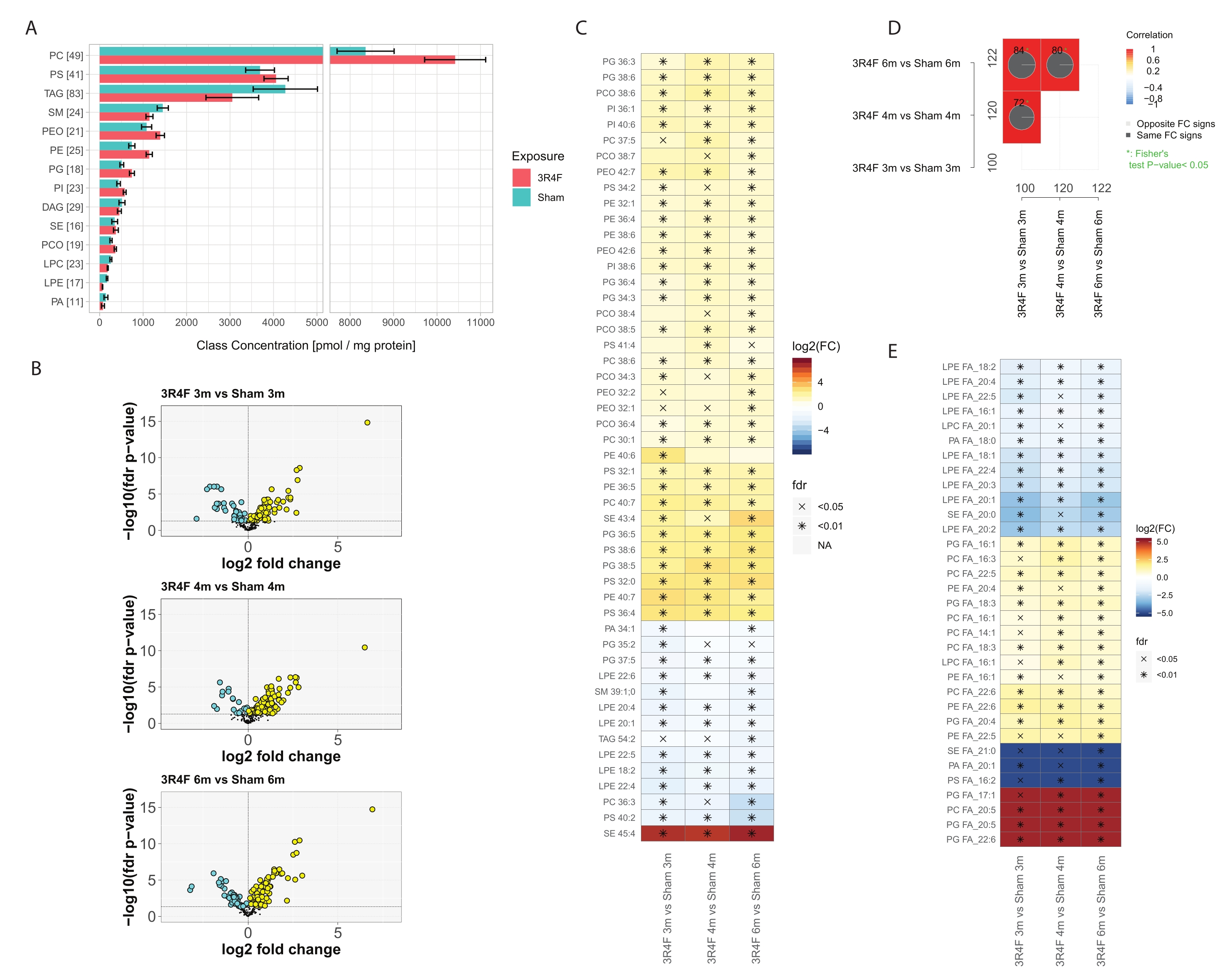
总体而言,鉴定并定量了来自14种最丰富的脂质类别的~400种分子脂质物种(图3)。PE、PEO、PC、PCO、PI 和 PG 脂质类别的每类总归一化脂质浓度略有升高,SM、LPE 和 PA 脂质观察到一些下调(图 3A),而 TAG 和 PS 没有差异。 有趣的是,据报道,CS 组中 PCO 和 PEO 缩醛磷脂脂质水平升高在炎症细胞中29.缩醛磷脂脂质的细胞内功能之一是抗氧化活性。在暴露于活性氧(ROS)和氮种(RNS)下,缩醛磷脂与二酰磷脂相比的选择性氧化报道了30。缩醛磷脂化学结构中的烯基醚键对氧化应激的自由基攻击敏感31。自由基攻击的主要产物是羟基化二十碳四烯酸、2-单酰基甘油磷脂、十五烷醇、甲酸、各种链长的α-羟基醛、1-甲酰基-2-花生四烯酰甘油磷脂和溶血磷脂。
这些结果表明,在基于总分子式的分子特征中,有100-120种化合物受到CS暴露的显着影响(图3D)。MS2片段信息的详细反卷积和片段信号强度的归一化允许近似定义每个脂质类别中表示的每种共轭脂肪酸(FA)的总量(图3E),并在暴露组和对照组之间比较这些数据。观察到完全饱和的FA和只有少量不饱和的FA明显减少,主要是在溶血脂质的背景下。相比之下,CS组PC、PE和PG磷脂类组成中的多不饱和脂肪酸(PUFA)在所有时间点均升高。PC和PG与二十碳五戊酸(EPA)和二十二碳六烯酸(DHA)结合的极端情况仅在3R4F CS暴露组中检测到(图3E,红色)。

图 1:布拉德福德测定中样品分布的板布局。 空白和标准样品一式三份放置在第1-3列中;未知样品一式两份放在第 4-11 列中。 请点击此处查看此图的大图。
{kind=link}

图 2:用于鸟枪式质谱分析的 96 孔微量滴定板的板布局。 在正电离模式下采集的空白、标准、未知和QC样品的分布映射到板的左侧(第1-6列);负电离模式的所有样品都放在右侧(第7-12列)。缩写:pos = 正数;质量控制 = 质量控制;负 = 负;TB = 总空白。 请点击此处查看此图的大图。
{kind=link}

图 3:暴露于香烟烟雾的小鼠肺部的霰弹枪脂质组学分析。 Apoe−/− 小鼠暴露于来自3R4F参考香烟或新鲜空气(假)的CS。(A)脂类浓度和每类脂类种类的数量。对于每个类别,定量脂质种类的数量在括号中给出。每个脂质类别的平均置信区间和 95% 置信区间,分别针对每种暴露类型(跨三个时间点聚合)。(B) 火山图显示三个时间点 3R4F CS 与假比较的量化脂质种类的效应大小(log2 倍变化)和显著性(-log10 FDR 调整的 p 值)。显著升高的脂质以黄色标记;显著降低的脂质以青色标记(FDR调整的p值<0.05)。(C)绝对平均倍数变化最大的25种脂质物种的差异丰度。3R4F CS 与假比较的 Log2 倍变化以颜色编码,并指示统计显著性:*:FDR 调整后的 p 值< 0.01;X:FDR 调整后的 p 值< 0.05。(四)比较图。倍数变化比较的相关系数用颜色编码,并指示共享的差异丰度脂质的数量(边缘中的总数)。饼图显示了具有相同变化方向(FC 符号)的共享差异丰度脂质的百分比。星号表示不同丰度脂质的显着重叠。(E)每个脂质类别的共轭FA的不同丰度。图 C 所示的共轭脂肪酸在所有三个时间点的3R4F和假暴露之间具有显着丰度差异(FDR调整的p值<0.05)。根据脂肪酸片段的MS2强度估计每种脂质类别的脂肪酸量。缩写:CS = 香烟烟雾;FDR = 错误发现率;FC = 倍数变化;FA = 脂肪酸;PC = 磷脂酰胆碱;PS = 磷脂酰丝氨酸;TAG = 三酰基甘油;SM = 鞘磷脂;PEO = 缩醛磷脂酰乙醇胺;PE = 磷脂酰乙醇胺;PG = 磷脂酰甘油;PI = 磷脂酰肌醇;DAG = 二酰基甘油;SE = 甾醇/胆固醇酯;PCO = 缩醛磷脂酰胆碱;LPC = 溶血磷脂酰胆碱;LPE = 溶血磷脂酰乙醇胺;PA = 磷脂酸。 请点击此处查看此图的大图。
{kind=link}
| BSA终浓度(毫克/毫升) | 2 毫克/毫升 (μL) 的 BSA 溶液 | 碳酸氢铵缓冲液(μL) |
| 1 | 50 | 50 |
| 0.75 | 37.5 | 62.5 |
| 0.5 | 25 | 75 |
| 0.25 | 12.5 | 87.5 |
| 0.125 | 12.5 | 187.5 |
| 0.0625 | 50 份 0.125 mg/mL 溶液 | 50 |
| 0 | 0 | 100 |
表1:布拉德福德测定校准曲线的BSA内标稀释方案。 缩写:BSA = 牛血清白蛋白。
补充信息:用于脂质探索者脂质鉴定的MFQL文件。 .zip包由各个必需的 MFQL 文件组成,每个文件都按照以下模式命名:<脂质类缩写>_<电离模式>_MS2.mfql(例如,PE_neg_MS2.mfql)。标记的内标标识的文件名包括<脂质缩写>中的D7(D9)标签(例如,PED7_neg_MS2.mfql)。这些MFQL文件用于验证内标中的氘量。 请点击此处下载此文件。
讨论
尽管MS的进步已经产生了多种方法来准确监测许多脂质种类,但以前对各种哺乳动物组织的脂质分析并未显示出一致的结果,因此,脂质的特定组织特异性功能仍不清楚。与蛋白质功能分析相比,蛋白质功能分析可以使用更可靠的方法来敲除特定化合物的表达,大多数脂质不能选择性地关闭或在组织中过表达,这使得脂质的功能评估变得困难。组织脂质组浓度的高级分析可能为识别循环脂质与人类疾病之间的关联提供了一种替代方法。
在评估能够定性和定量覆盖任何小鼠组织的内源性脂质谱的综合脂质组学方法时,我们优先考虑鸟枪式脂质组学方法。通常,可以进行两种相反类型的样品分析:使用基于液相色谱(LC)的分离对脂质进行完全非靶向筛选,并进一步进行质谱检测以揭示样品的总脂质复杂性,或者靶向方法,该方法通常允许非常精确地定量感兴趣的特定脂质。相比之下,所提出的Shotgun法脂质组学工作流程的强大之处在于能够快速全面覆盖来自预定义脂质类别的数百种内源性脂质,这仍然可以以稳健的半定量方式进行。
不同脂质类别的电离效率取决于结构,并且根据不同的实验电离条件可能会有很大差异。与基于 LC 的分离方法相比,霰弹枪法分析最大限度地减少了这些差异,因为在相同的电离条件下将整个脂质提取物直接同时输注到 MS 仪器中。使用同位素标记的脂质类似物或结构相似的非内源性标准品可以对所有脂质类别进行半定量。霰弹枪式脂质组学在 MS 分析期间提供低运行间和运行中变异性;因此,与基于非靶向液相色谱的方法相比,该方法产生的变异系数21 更低,后者需要多种标准品才能充分定量。重要的是,虽然当前方法中没有使用外部校准曲线,但该方法仍被认为是完全定量的32。
每个脂质类别一个水平的标记内标(或未内源性表达的未标记标准品)足以定量大多数脂质。只有少数出版物报道了霰弹枪法脂质组学方法的部分方法验证。例如,在Gryzbek等人17 和Surma等人21中,使用内标和固定量的样品基质制备反向校准曲线。通过对数变换脂质量及其强度的线性回归评估线性,并分别报告为 R2 和斜率。检测限(LOD)和定量限(LOQ)通过加权线性回归确定,基于LOD的信噪比为3,LOQ的信噪比为10。对于大多数脂质类别,脂肪组织的LOQ定义为2-9.8pmol和血浆中的0.05-5μM之间。在这两种情况下,均使用每类非内源性单一内标来估算该类中所有脂质的估计值。然而,在这项工作中,由于几个问题,我们不提供LOD / LOQ:内源性基质不是无化合物的,并且组织的替代基质不可用 - 因此,不可能尖峰少量已知量的标准品。由于不存在纯标准品且同位素标记脂质的可用性非常有限,因此我们不使用由其相同的同位素标记标准品归一化的特定化合物的校准曲线系列来执行经典的靶向定量方法。Orbitrap探测器通过应用傅里叶变换自动执行瞬态信号的转换,并且某些信号已经被取代 - 因此,较低的浓度范围将仅线性下降到某个最小信号,低于该范围将不再可检测到分子。Xcalibur软件信噪比值取决于分子的m / z比;因此,含有不同脂肪酸组合的每个脂质类别的化合物将具有不同的噪声值。此外,LOQ/LOQ值与基质类型严格相关,当在不同的啮齿动物组织中对脂质组进行定量时,应通过单独评估每种组织类型的LOQ来反映。
事实上,该方法提供了高达4个数量级33的高线性动态定量范围和非常好的灵敏度,可以覆盖最重要的内源性结构脂质,这可以通过MS采集32的技术改进进一步提高。平均脂质浓度的变异系数大多低于15%,这意味着霰弹枪式脂质组学符合美国食品和药物管理局(FDA)的要求,是良好实验室规范(GLP)和良好临床实践(GCLP)研究需要考虑的方法34。
然而,必须注意的是,由于它们的极性不同,某些脂质类别受到特定共轭FA的贡献的影响更大。这导致含有广泛共轭FA的等摩尔混合物中的强度响应失真,导致定量偏差,正如Koivusalo等人35 对磷脂类别所强调的那样。值得注意的是,这些作者提供了广泛的FA的数据,从24-48链长度,这可能不能反映真实生物样本中的情况。多不饱和物种的响应比完全饱和物种高40%,但仅在较高浓度下观察到这种效应。当混合物逐渐稀释时,不饱和度的影响逐渐减弱,几乎以每物质0.1 pmol/μL消失。此外,所有测量都是在离子阱和三重四极仪器上进行的,而不是在Q-Exactive设备上进行的。
所呈现的工作流程的另一个优点是其技术灵活性,可以适应特定的项目要求。例如,任何脂质提取方案——例如改性的Bligh和Dyer36、甲基叔丁基醚37或丁醇-甲醇38的方法——都可用于在MS分析之前制备总脂质提取物。氯仿-甲醇萃取的主要局限性是下层含有脂质级分,这使得日常工作复杂化,尤其是自动化;此外,还必须考虑氯仿的毒性。叔丁基醚萃取广泛用于血浆样品的脂质分析37,并提出了自动化版本21。在这种情况下,我们选择了BUME方法,因为它对加标的PG、PI、PA和PS脂质等级38提供了更好的回收率,降低了溶剂消耗,并具有自动化39的可能性,而所有三种方法提取的组织样品的总浓度相当。此外,虽然样品提取在当前工作中是手动进行的,但通过自动样品制备和 96 孔格式的脂质提取40,41,也可以从大型样品集中获得可重现和精确的结果。这允许人们在大规模的临床和毒理学研究中实施脂质组学分析。
在当前的工作中,我们分别进行了正负模式的MS采集,而无需像Schuhmann等人42所描述的极性切换。对于浓度略低的溶液,纳米酸盐信号在负模式下的稳定性优于正模式。此外,我们使用转换器软件开发了一个完全可追溯的程序,从.raw文件到 mzML,它提供了要在 LipidXplorer 软件中指定的值 - 这样,不需要手动分辨率斜率计算。我们还应用了不同的噪声设置替换,因为在正模式下,频谱的噪声水平高于负模式。所有步骤都针对可追溯的高通量常规分析进行了优化。
对于鉴定,鸟枪式脂质组学分析可以利用不同脂质类别的独特行为,这些行为在不同极性模式下形成独特的加合物。在该方法中,在正电离模式下重叠的具有相同分子量的PC和PE物种在负电离模式下可以完全分离,因为PC形成乙酸盐或甲酸盐加合物,而PE以去质子化形式电离。此外,(部分)结构反卷积对于不仅利用分子式而且利用块状脂肪酸结构的方法都是可能的。例如,总碳和总不饱和度计数水平的FA鉴定适用于所有磷脂,DAG,TAG和溶血磷脂。对于某些磷脂类别43 ,可以部分执行每种异构体形式的自下而上定量,但由于MS2谱图中不同FA的信号响应不相等,因此对于DAG和TAG来说要复杂得多。
还必须强调需要实施适当的质量控制程序,与该领域最近的举措完全一致44.由于我们希望确保实验室之间和实验室内适当的数据可追溯性和可重复性,因此已经采取了许多步骤,例如在分析的所有步骤中对样品进行适当的随机化,使用供应商认证的标准混合物,包括质量控制样品,验证批次接受或拒绝的程序,以及创建内部数据库以长期跟踪QC性能。与这些举措相一致的还有需要一种标准化的方法来解决样品稳定性问题。一般来说,大多数结构脂质不受脂质氧化的影响,这与脂质氧化、氧化脂质和多不饱和脂肪酸更相关,因此对储存和处理条件更敏感。然而,正确评估样品稳定性仍然是一项技术上具有挑战性的任务。
然而,在确定化合物的亚分子水平时,该协议具有局限性。考虑到总提取物没有分离,在MS分析中合并了具有相同分子量但不同脂肪酸组成的脂质的所有亚型。对于大多数类,通过使用MS2中残留脂肪酸片段的碎裂率可以实现结构的部分反卷积。然而,由于不同亚型的碎裂行为存在巨大差异,并且纯化学标准品的种类不足以定义补偿值,因此每种亚型的独立定量仍然是一项特别具有挑战性的任务。另一个限制是ESI过程不可避免地会产生伪影,导致某些脂质(如DAG、Pa和FA)人为产生峰,这可能导致错误的定量。
接下来,我们根据我们的经验总结协议中最关键的部分。第一个与以下事实有关:每种小鼠组织类型在脂质量和类别比率方面都具有独特的脂质谱。因此,必须仔细确定基于提取前总蛋白质含量的组织起始量,以免使MS信号饱和,并且不会由于高浓度33 的脂质聚集而离开动态定量范围,或者在相反的极端中提供足够的MS信号以覆盖每个脂质类别的主要脂质化合物。
第二个关键方面是确保直接输液纳米源芯片出口的位置与质谱仪的转移毛细管正确对齐。考虑到质谱仪在两种模式下的完全校准每周进行一次,校准源和纳米源芯片设置之间的切换可能是由于安装过程中未对准而导致信号强度急剧变化的原因。
该协议的另一个关键部分是仔细处理内标混合物。由于这种混合物含有大量的二氯甲烷,一旦打开,应迅速食用,以避免长时间储存和多次使用导致蒸发和人为浓度变化。此外,从−20 °C储存中取出标准混合物后,对标准混合物的一致处理非常重要,因为使用气垫移液器移液时,温差会导致体积不一致。一种选择是用纯甲醇代替标准重悬缓冲液中的二氯甲烷,这可以提高操作的便利性,但可能会对某些脂质类别的溶解度产生负面影响,从而降低这些脂质类别的定量准确性。
最后一个关键部分是数据处理。数据处理工作流程结合了逐峰软件从 .raw 到 .mzML 格式的逐峰转换,应用 MS2 扫描平均和 MS1 和 MS2 噪声过滤,以及峰拾取和数据压缩。作为替代方案,Proteowizard软件也可用于数据转换,但在这种情况下,必须手动定义LipidXplorer中的几个设置。霰弹枪式脂质组学的所有复杂性都特别集中在直接注入MS1和MS2光谱的反卷积步骤上。开源 LipidXplorer 软件根据质量精度、质量分辨率及其随 m/z 增加而变化的斜率,从 mzML 文件格式导入转换后的光谱。该软件合并了在分析运行中获取的多个单独的MS和MS / MS谱图。之后,它对齐不同样品运行中的单个峰,并且在每组对齐峰中,用其单个强度加权平均质量数替换其质量,而每个数据文件中的丰度保持不变。对齐峰簇和单个峰强度的代表性质量数存储在主扫描数据库中。主扫描数据库包含为批次中的所有样品生成的所有 MS1 和 MS2 光谱,并且可以通过用分子碎裂查询语言 (MFQL) 编写的查询对脂质鉴定进行反卷积。
总体而言,该方法涵盖了基于正模式的DAG、TAG和SE脂质的鉴定,以及基于负模采集的PC、PE、PS、PI、PA、PG、SM、LPC和LPE脂质的鉴定。在脂质鉴定过程中,对MS1和MS2进行同位素校正,并在.xlsx输出文件中报告调整后的强度。或者,其他几种软件可用于处理霰弹枪数据,例如ALEX45 和LipidHunter46。
脂肪酸 - 主要的核和细胞膜成分 - 以磷脂的形式储存,以进一步转化为生物活性分子。它们可以通过Lands途径47被溶血磷脂酰基转移酶转化为溶血磷脂。例如,已知LPCAT3酶显示出将AA掺入溶血磷脂酰胆碱和溶血磷脂酰丝氨酸中间体的高特异性。据报道,这些酶在炎症细胞(例如肺泡巨噬细胞和支气管上皮细胞47)中表达相对较高,导致在这些细胞中释放更大比例的含AA磷脂。然而,在磷脂酶A2从膜磷脂中释放出来后,许多酶催化了PUFAs转化为各种类型的脂质介质48。此外,这些PUFAs可以通过环加氧酶(COX)-1和COX-2酶的作用 成为 生产促炎和抗炎前列腺素的底物47。磷脂是脂质介质在特定位置释放的来源之一,在这些位置需要这些介质来证明其生物效应。
肺是一个复杂的器官,由多种细胞类型组成,每种细胞类型在促进正常肺发育和功能方面发挥着重叠和生态位的作用。只有少数研究用于分离和分析小鼠或人肺中的不同细胞类型(例如,肺泡2型细胞)49,50;其他主要的肺细胞类型尚未表征。另一项有趣的研究51 用于分离和进行小鼠肺中的内皮,上皮,间充质和混合免疫细胞的脂质分析。观察到PCO和PG中掺入的PUFAs的浓度在免疫细胞中富集。考虑到CS诱导肺内免疫细胞增加(4-5倍)的事实,如支气管肺泡灌洗(BAL)52评估的那样,本研究中观察到的总脂质变化可以通过小鼠肺中免疫细胞募集的累积增加来解释。
总之,观察到的磷脂中掺入的单不饱和脂肪酸和多不饱和脂肪酸的扭曲可以反映细胞内某些FA的过量和/或构成在氧化应激和炎症条件下过度产生的氧磷脂前体的细胞内资源。然而,关于游离EPA、DHA和其他脂磷脂的额外数据对于澄清这一点至关重要,并且超出了当前方法的应用范围。
披露声明
所有作者都是菲利普莫里斯国际公司的员工。菲利普莫里斯国际是该项目的唯一资金来源和赞助商。
致谢
作者要感谢研究团队,并特别感谢PMI R&D,菲利普莫里斯国际研究实验室私人有限公司生物研究和气溶胶团队的技术援助和支持。Ltd.,新加坡,PMI研发,菲利普莫里斯产品有限公司,纳沙泰尔,瑞士。作者感谢Sam Ansari管理生物样本库,并感谢Sindhoora Bhargavi Gopala Reddy编辑手稿的支持。
材料
| Name | Company | Catalog Number | Comments |
| 1, 5, and 2 mL self-lock tubes | Eppendorf | 30120086, 30120094 | |
| 3 mm stainless still beads | Qiagen | 69997 | |
| 4.1 µm nozzle chip | Advion | HD-D-384 | |
| Acetic acid | Sigma Aldrich | 45754-100ML-F | |
| Ammonium acetate | Honeywell | 14267-25G | |
| Ammonium bicarbonate | Sigma Aldrich | 09830-500G | |
| Bovine serum albumin standard, 2 mg/mL | Thermo Scientific | 23209 | |
| Butanol | Honeywell | 33065-2.5L | |
| Chloroform | Sigma Aldrich | 650498-1L | |
| Dichloromethane | Honeywell | 34856-1L | |
| Ethyl acetate | Honeywell | 33211 | |
| Greiner CELLSTAR 96 well plates | Sigma | M9686 | |
| Heptane | Sigma Aldrich | 34873-2.5L | |
| Isopropanol | Fisher Scientific | A461 | |
| Methanol | Fisher Scientific | A456 | |
| Mouse pooled plasma | BioIvt | ||
| Mouse SPLASH standard | Avanti Polar Lipids | 330710X | Internal standard |
| Nunc 96-flat bottom well transparent plates | VWR | 62409-068 | |
| Plastic spatula | Sigma | Z560049-300EA | |
| Quick Start Bradford 1x Dye reagent | BioRad | 5000205 | |
| Serum diluent | Sigma Aldrich | D5197 | |
| Equipment/software | |||
| CryoPrep CP02 impactor instrument | Covaris | Magnetic hammer | |
| Centrifuge 5427R | Eppendorf | . | Centrifuge |
| ChipSoft 8.3 | Advion Biosciences | . | Software to set up method and acquisition on the Triversa nanomate robot |
| LipidXplorer 1.2.8.1 | N/A | . | Software to identify lipids |
| Peak By Peak | SpectroSwiss | . | Software to convert .raw data from MS to .mzml format |
| ProteoWizard | ProteoWizard | . | Alternative (open source) software to convert .raw data from MS to .mzml format |
| Q-Exactive MS | Thermo Fisher | . | High resolution orbitrap mass spectrometer |
| Qiagen Tissue Lyser II | Qiagen | . | Tissue lyser |
| SpeedVac SPD140DDA | Thermo Fisher | . | Vacuum concentrator |
| Tecan Infinite M nano plus | Tecan | . | Plate reader |
| ThermoMixer C | Eppendorf | . | Thermomixer |
| TriVersa Nanomate | Advion Biosciences | . | Direct infusion nano-source |
| Xcalibur 4.3 | Thermo Scientific | . | Software to set up method and acquisition on the Q-Exactive MS |
参考文献
- Salehi, N., Janjani, P., Tadbiri, H., Rozbahani, M., Jalilian, M. Effect of cigarette smoking on coronary arteries and pattern and severity of coronary artery disease: A review. Journal of International Medical Research. 49 (12), 3000605211059893 (2021).
- Churg, A., Sin, D. D., Wright, J. L. Everything prevents emphysema: Are animal models of cigarette smoke-induced chronic obstructive pulmonary disease any use. American Journal of Respiratory Cell and Molecular Biology. 45 (6), 1111-1115 (2011).
- Lee, G., Walser, T. C., Dubinett, S. M. Chronic inflammation, chronic obstructive pulmonary disease, and lung cancer. Current Opinion in Pulmonary Medicine. 15 (4), 303-307 (2009).
- Titz, B., et al. Effects of cigarette smoke, cessation, and switching to two heat-not-burn tobacco products on lung lipid metabolism in C57BL/6 and Apoe-/- mice-An integrative systems toxicology analysis. Toxicological Sciences. 149 (2), 441-457 (2016).
- Morissette, M. C., Shen, P., Thayaparan, D., Stampfli, M. R. Disruption of pulmonary lipid homeostasis drives cigarette smoke-induced lung inflammation in mice. European Respiratory Journal. 46 (5), 1451-1460 (2015).
- Jubinville, E., et al. Interplay between cigarette smoking and pulmonary reverse lipid transport. European Respiratory Journal. 50 (3), 1700681 (2017).
- Han, X., Gross, R. W. Shotgun lipidomics: Electrospray ionization mass spectrometric analysis and quantitation of cellular lipidomes directly from crude extracts of biological samples. Mass Spectrometry Reviews. 24 (3), 367-412 (2005).
- Titz, B., et al. Multi-omics systems toxicology study of mouse lung assessing the effects of aerosols from two heat-not-burn tobacco products and cigarette smoke. Computational and Structural Biotechnology Journal. 18, 1056-1073 (2020).
- Hartung, T., et al. Systems toxicology: Real world applications and opportunities. Chemical Research in Toxicology. 30 (4), 870-882 (2017).
- Talikka, M., et al., Lyubimov, A., et al. Systems Toxicology. Encyclopedia of Drug Metabolism and Interactions. , (2016).
- Ejsing, C. S., et al. Global analysis of the yeast lipidome by quantitative shotgun mass spectrometry. Proceedings of the National Academy of Sciences of the United States of America. 106 (7), 2136-2141 (2009).
- Papan, C., et al. Systematic screening for novel lipids by shotgun lipidomics. Analytical Chemistry. 86 (5), 2703-2710 (2014).
- Carvalho, M., et al. Effects of diet and development on the Drosophila lipidome. Molecular Systems Biology. 8, 600 (2012).
- Strittmatter, N., et al. Shotgun lipidomic profiling of the NCI60 cell line panel using rapid evaporative ionization mass spectrometry. Analytical Chemistry. 88 (15), 7507-7514 (2016).
- Lydic, T. A., et al. Rapid and comprehensive 'shotgun' lipidome profiling of colorectal cancer cell derived exosomes. Methods. 87, 83-95 (2015).
- Sampaio, J. L., et al. Membrane lipidome of an epithelial cell line. Proceedings of the National Academy of Sciences of the United States of America. 108 (5), 1903-1907 (2011).
- Grzybek, M., et al. Comprehensive and quantitative analysis of white and brown adipose tissue by shotgun lipidomics. Molecular Metabolism. 22, 12-20 (2019).
- Stegemann, C., et al. Comparative lipidomics profiling of human atherosclerotic plaques. Circulation: Cardiovascular Genetics. 4 (3), 232-242 (2011).
- Tajima, Y., et al. Lipidomic analysis of brain tissues and plasma in a mouse model expressing mutated human amyloid precursor protein/tau for Alzheimer's disease. Lipids in Health and Disease. 12, 68 (2013).
- Gross, R. W., Han, X. Shotgun lipidomics of neutral lipids as an enabling technology for elucidation of lipid-related diseases. American Journal of Physiology: Endocrinology and Metabolism. 297 (2), 297-303 (2009).
- Surma, M. A., et al. An automated shotgun lipidomics platform for high throughput, comprehensive, and quantitative analysis of blood plasma intact lipids. European Journal of Lipid Science and Technology. 117 (10), 1540-1549 (2015).
- Heiskanen, L. A., Suoniemi, M., Ta, H. X., Tarasov, K., Ekroos, K. Long-term performance and stability of molecular shotgun lipidomic analysis of human plasma samples. Analytical Chemistry. 85 (18), 8757-8763 (2013).
- Zhang, S., Van Pelt, C. K. Chip-based nanoelectrospray mass spectrometry for protein characterization. Expert Review of Proteomics. 1 (4), 449-468 (2004).
- Surma, M. A., et al. Mouse lipidomics reveals inherent flexibility of a mammalian lipidome. Scientific Reports. 11 (1), 19364 (2021).
- Herzog, R., Schwudke, D., Shevchenko, A. LipidXplorer: Software for quantitative shotgun lipidomics compatible with multiple mass spectrometry platforms. Current Protocols in Bioinformatics. 43, 11-30 (2013).
- R: A language and environment for statistical computing. R: Foundation for Statistical Computing Available from: https://www.R-project.org/ (2018)
- Phillips, B., et al. A 7-month cigarette smoke inhalation study in C57BL/6 mice demonstrates reduced lung inflammation and emphysema following smoking cessation or aerosol exposure from a prototypic modified risk tobacco product. Food and Chemical Toxicology. 80, 328-345 (2015).
- Phillips, B., et al. A six-month systems toxicology inhalation/cessation study in ApoE(-/-) mice to investigate cardiovascular and respiratory exposure effects of modified risk tobacco products, CHTP 1.2 and THS 2.2, compared with conventional cigarettes. Food and Chemical Toxicology. 126, 113-141 (2019).
- Zemski Berry, K. A., Murphy, R. C. Electrospray ionization tandem mass spectrometry of glycerophosphoethanolamine plasmalogen phospholipids. Journal of the American Society for Mass Spectrometry. 15 (10), 1499-1508 (2004).
- Engelmann, B. Plasmalogens: Targets for oxidants and major lipophilic antioxidants. Biochemical Society Transactions. 32, 147-150 (2004).
- Nagan, N., Zoeller, R. A. Plasmalogens: Biosynthesis and functions. Progress in Lipid Research. 40 (3), 199-229 (2001).
- Southam, A. D., Weber, R. J., Engel, J., Jones, M. R., Viant, M. R. A complete workflow for high-resolution spectral-stitching nanoelectrospray direct-infusion mass-spectrometry-based metabolomics and lipidomics. Nature Protocols. 12 (2), 310-328 (2016).
- Yang, K., Han, X. Accurate quantification of lipid species by electrospray ionization mass spectrometry - Meet a key challenge in lipidomics. Metabolites. 1 (1), 21-40 (2011).
- Zullig, T., Trotzmuller, M., Kofeler, H. C. Lipidomics from sample preparation to data analysis: a primer. Analytical and Bioanalytical Chemistry. 412 (10), 2191-2209 (2020).
- Koivusalo, M., Haimi, P., Heikinheimo, L., Kostiainen, R., Somerharju, P. Quantitative determination of phospholipid compositions by ESI-MS: Effects of acyl chain length, unsaturation, and lipid concentration on instrument response. Journal of Lipid Research. 42 (4), 663-672 (2001).
- Bligh, E. G., Dyer, W. J. A rapid method of total lipid extraction and purification. Canadian Journal of Biochemistry and Physiology. 37 (8), 911-917 (1959).
- Matyash, V., Liebisch, G., Kurzchalia, T. V., Shevchenko, A., Schwudke, D. Lipid extraction by methyl-tert-butyl ether for high-throughput lipidomics. Journal of Lipid Research. 49 (5), 1137-1146 (2008).
- Lofgren, L., Forsberg, G. B., Stahlman, M. The BUME method: A new rapid and simple chloroform-free method for total lipid extraction of animal tissue. Scientific Reports. 6, 27688 (2016).
- Lofgren, L., et al. The BUME method: A novel automated chloroform-free 96-well total lipid extraction method for blood plasma. Journal of Lipid Research. 53 (8), 1690-1700 (2012).
- Jung, H. R., et al. High throughput quantitative molecular lipidomics. Biochimica et Biophysica Acta. 1811 (11), 925-934 (2011).
- Lavrynenko, O., et al. Ceramide ratios are affected by cigarette smoke but not heat-not-burn or e-vapor aerosols across four independent mouse studies. Life Sciences. 263, 118753 (2020).
- Schuhmann, K., et al. Shotgun lipidomics on a LTQ Orbitrap mass spectrometer by successive switching between acquisition polarity modes. Journal of Mass Spectrometry. 47 (1), 96-104 (2012).
- Schuhmann, K., et al. Quantitative fragmentation model for bottom-up shotgun lipidomics. Analytical Chemistry. 91 (18), 12085-12093 (2019).
- Lipidomics Standards Initiative Consortium. Lipidomics needs more standardization. Nature Metabolism. 1 (8), 745-747 (2019).
- Husen, P., et al. Analysis of lipid experiments (ALEX): A software framework for analysis of high-resolution shotgun lipidomics data. PloS One. 8 (11), 79736 (2013).
- Ni, Z., Angelidou, G., Lange, M., Hoffmann, R., Fedorova, M. LipidHunter identifies phospholipids by high-throughput processing of LC-MS and shotgun lipidomics datasets. Analytical Chemistry. 89 (17), 8800-8807 (2017).
- Zemski Berry, K. A., Murphy, R. C., Kosmider, B., Mason, R. J. Lipidomic characterization and localization of phospholipids in the human lung. Journal of Lipid Research. 58 (5), 926-933 (2017).
- Ghosh, M., Tucker, D. E., Burchett, S. A., Leslie, C. C. Properties of the Group IV phospholipase A2 family. Progress in Lipid Research. 45 (6), 487-510 (2006).
- Besnard, V., et al. Deletion of Scap in alveolar type II cells influences lung lipid homeostasis and identifies a compensatory role for pulmonary lipofibroblasts. Journal of Biological Chemistry. 284 (6), 4018-4030 (2009).
- Plantier, L., et al. Activation of sterol-response element-binding proteins (SREBP) in alveolar type II cells enhances lipogenesis causing pulmonary lipotoxicity. Journal of Biological Chemistry. 287 (13), 10099-10114 (2012).
- Kyle, J. E., et al. Cell type-resolved human lung lipidome reveals cellular cooperation in lung function. Scientific Reports. 8 (1), 13455 (2018).
- Lugg, S. T., Scott, A., Parekh, D., Naidu, B., Thickett, D. R. Cigarette smoke exposure and alveolar macrophages: mechanisms for lung disease. Thorax. 77 (1), 94-101 (2022).
转载和许可
请求许可使用此 JoVE 文章的文本或图形
请求许可探索更多文章
This article has been published
Video Coming Soon
版权所属 © 2025 MyJoVE 公司版权所有,本公司不涉及任何医疗业务和医疗服务。